Гидрофобное несоответствие - Hydrophobic mismatch
Гидрофобное несоответствие разница между толщиной гидрофобный регионы трансмембранный белок и из биологическая мембрана он охватывает.[1] Чтобы избежать неблагоприятного воздействия воды на гидрофобные поверхности, ожидается, что гидрофобные области трансмембранных белков будут иметь примерно такую же толщину, что и гидрофобная (липидная ацильная цепь) область окружающей среды. липидный бислой. Тем не менее, один и тот же мембранный белок может встречаться в бислое разной толщины.[2] В эукариотических клетках плазматическая мембрана толще, чем мембраны эндоплазматического ретикулума. Однако все белки, которых много в плазматической мембране, первоначально интегрируются в эндоплазматический ретикулум после синтеза на рибосомах.[3] Трансмембранные пептиды или белки и окружающие липиды могут адаптироваться к гидрофобному несоответствию различными способами.
Возможные приспособления к несоответствию
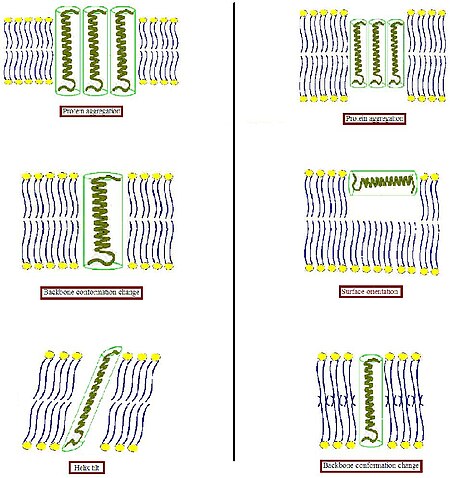
Чтобы избежать неблагоприятного воздействия на гидрофобные поверхности гидрофильной среды, биологическая мембрана имеет тенденцию к некоторой адаптации к такому несоответствию. В различных других системах целостный белок имеет тенденцию окружать себя липидами соответствующего размера и формы.[4] Поскольку белки относительно жесткие, в то время как липидные углеводородные цепи гибкие, условие гидрофобного соответствия может быть выполнено путем растяжения, сжатия и / или наклона липидных цепей.[5][6][7]
- Когда гидрофобная часть трансмембранного белка слишком толстая, чтобы соответствовать толщине гидрофобного бислоя (левая часть рисунка), белок может агрегироваться в мембране, чтобы минимизировать открытую гидрофобную область, или наклонить, чтобы уменьшить их эффективную гидрофобную толщину. Они также могут принять другую конформацию, изменив ориентацию гидрофобных и гидрофильных боковых цепей вблизи границы раздела. Липиды, в свою очередь, могут модулировать толщину мембраны за счет растяжения своих ацильных цепей или даже собираться в другой тип агрегата, тем самым нарушая организацию бислоя.
- Когда гидрофобная часть трансмембранного белка слишком мала, чтобы соответствовать толщине гидрофобного бислоя (правая часть рисунка), это опять же может привести к агрегации белка или изменениям конформации основной цепи и / или ориентации боковой цепи. Кроме того, слишком короткие пептиды могут не включать и вместо этого принимать поверхностную локализацию. Липиды могут уменьшать эффективную толщину бислоя за счет разупорядочения своих ацильных цепей или нарушать организацию бислоя с образованием перевернутой неламеллярной структуры. Возможны также комбинации этих возможных способов адаптации.
Агрегация белков
Так как Моуритсен и Блум[8] предложили детальную термодинамическую модель, которая включает адаптацию липидов и индукцию сегрегации белков при более крайнем несоответствии в своей «Модели матраса», было получено больше дополнительных сведений об агрегации белков, вызванной несовпадением.[9] Также были обнаружены некоторые экспериментальные доказательства того, что гидрофобное несоответствие может приводить к агрегации белков в жидком бислое.[10][11][12] Электронно-микроскопические исследования бактериородопсина, восстановленного в насыщенных и ненасыщенных жидких двойных слоях ПК с различной длиной цепи, показали, что агрегация белков происходит только с довольно большим несоответствием, и что толщина бислоя на 4 Ангстрем толще и на 10 Ангстрем тоньше, чем расчетная гидрофобная толщина слоя. белки допускаются без индукции значительной агрегации.[13]
Наклон спирали
Наклон также возможен, если гидрофобная часть пептида или белка слишком длинная, чтобы охватить мембрану. Предыдущее исследование лактозопермеазы E. coli показало, что при восстановлении белка в липидном бислое PE / PG (3/1) увеличение наклона спирали происходит при увеличении содержания белка.[14] Этот наклон сопровождается уменьшением липидного порядка, что приводит к уменьшению толщины бислоя, подтверждая, что это ответ, связанный с несоответствием. В больших белках, которые многократно охватывают мембрану, могут происходить изменения наклона спирали с небольшим влиянием на упаковку липидов. Однако для одиночной трансмембранной спирали возможно, что наклон вызовет напряжение на окружающие липиды, чтобы разместить спираль в бислое. Таким образом, большая степень наклона может быть менее благоприятным вариантом для отдельных трансмембранных белков.
Ориентация поверхности
Относительно небольшие гидрофобные пептиды могут быть неспособны интегрироваться в мембрану и в ответ принимают ориентацию на поверхности мембраны. Экспериментальные доказательства были продемонстрированы флуоресцентным исследованием искусственного пептида с гидрофобной последовательностью из 19 аминокислот, состоящей в основном из лейцинов и фланкированной с обеих сторон лизинами в качестве якорных остатков.[15] Результаты показали, что преобразование от доминантной трансмембранной к параллельной ориентации пептида может быть вызвано модулированием толщины бислоя посредством добавления холестерина или увеличения длины липидной цепи.
Изменение конформации позвоночника
Для получения подробной информации о последствиях несоответствия конформации пептидов и белков в липидном бислое наиболее подходят небольшие трансмембранные пептиды. Еще нужны исследования.
Теории эффектов рассогласования
Были применены различные теоретические подходы для описания затрат энергии и термодинамических эффектов несоответствия, включая рассмотрение мембраны как эластичного листа.[16][17]или микроскопический подход.
Модель матраса

Модель матраса была предложена в качестве феноменологического теоретического подхода в 1984 году Моуритсеном и Блумом.[18] Это двухкомпонентная теория реальных решений, основанная на теории неидеальных решений и, следовательно, учитывающая разделение фаз. В своей модели они связывают энергию, запасенную в волнообразных волнах поверхности мембраны, вызванную несоответствием, с эластичными свойствами липидов и белков. Они не включают микроскопические детали липидов, но используют в качестве исходных данных известные термодинамические свойства чистой липидной системы. Они также включают непрямые липид-белковые взаимодействия, вызванные несоответствием, а также прямые липид-белковые ван-дер-ваальсовы взаимодействия между гидрофобными частями липидного бислоя и белками. Избыточный «гидрофобный эффект» связан с гидрофобным несоответствием липид-белок и свободной энергией упругой деформации липидных цепей вблизи белка. Потенциалы взаимодействия оцениваются на основе экспериментальных данных, полученных из термодинамических и механических измерений свойств мембраны.
Схема моделирования Монте-Карло
Модель матраса была позже воспроизведена в схеме моделирования Монте-Карло Сперотто и Моуритсеном.[19] Они учитывали различные микросостояния липидов, классифицированные в соответствии с моделью Пинка с 10 состояниями.[20] следовательно, возможен фазовый переход чистого липидного бислоя. Эта версия модели обеспечивает связь между микроскопическими характеристиками системы и ее термодинамическим поведением.
Молекулярная теория
В молекулярной теории липидных цепей мембраны пептиды с их гидрофобной длиной рассматриваются как обеспечивающие граничное условие для конфигурации липидных цепей.[21] Молекулярное моделирование было объединено с феноменологическим вкладом свободной энергии, описывающим отталкивание головной группы липидов и поверхностное натяжение мембранного растворителя. Duque et al.[22].
Экспериментальные исследования гидрофобного несоответствия и наклона спирали
Информация об ответе мембранных белков на несоответствие была получена в результате множества экспериментальных исследований. Различные типы экспериментальных подходов обеспечивают различное понимание вклада вышеупомянутых гипотетических молекулярных ответов. Например, белки или пептиды, снабженные флуоресцентными или парамагнитными метящими группами, можно использовать в флуоресцентной спектроскопии и электронный спиновой резонанс исследования. Они могут выявить молекулярные детали как белок-липидных взаимодействий, так и белок-белковых взаимодействий (характерных для реакции типа агрегации), а также то, как на них влияют (не) условия совпадения. Исследования наклона спирали как функции толщины мембраны также выиграли от использования твердотельный ЯМР методы, в частности, с использованием ориентированных мембран, которые обеспечивают непосредственное понимание угла наклона спирали. Ранние исследования модельных мембранных пептидов (таких как WALP пептид ) предоставили представление о различных факторах, которые влияют на ответ, включая состав мембраны, пептидную последовательность и, в частности, также наличие межфазных якорных остатков. В последние годы большие успехи в Рентгеновская кристаллография и электронная микроскопия Эти методы позволили по-новому взглянуть на липидные взаимодействия более крупных белков. Примером этого является понимание наклона спирали в кристаллизованном белке кальциевого насоса. [23].
Биологическое значение несоответствия
Гидрофобное несоответствие важно для сортировка белков и формирование липидные рафты.
Сортировка белков
В эукариотических клетках уровень холестерина увеличивается по секреторному пути от эндоплазматического ретикулума до Гольджи и плазматической мембраны,[24] предполагая сопутствующее увеличение толщины мембраны. В соответствии с этим, средняя длина трансмембранного сегмента однопролетных белков плазматической мембраны обычно на пять аминокислот больше, чем средняя длина белков Гольджи.[25] Было получено экспериментальное доказательство того, что сортировка белков в Гольджи может быть основана на этой разнице в длине: для нескольких белков, которые обычно находятся в Гольджи, было показано, что увеличение их гидрофобной длины может перенаправить белки к плазматической мембране, или наоборот, что уменьшение гидрофобной длины белков плазматической мембраны может вызвать их задержку в Гольджи.[26][27]
Липидные рафты
Рафты представляют собой мембранные домены, обогащенные холестерином, сфингомиелином (SM) и некоторыми мембранными белками.[28][29] Предполагается, что рафты играют роль во многих физиологических процессах, таких как передача сигналов, эндоцитоз, апоптоз, перенос белков и регуляция липидов.[30][31][32][33][34] Липиды рафта обычно имеют насыщенные углеводородные цепи.[35] Липидные рафты имеют более высокую гидрофобную толщину, чем остальная часть липидного бислоя, что может привести к предпочтительному разделению трансмембранных белков с более высокой гидрофобной толщиной на липидные рафты.[36][37]
Смотрите также
Рекомендации
- ^ Duque, D .; Li, X.J .; Кацов, К .; Schick, M. J. Chem. Phys. 2002, 116, 10478-10484.
- ^ Киллиан, Дж. Biochimica et Biophysica Acta. 1998, 1376, 401-416.
- ^ ван Меер, Г. Анну. Rev. Cell Biol. 1989, 5, 247-275.
- ^ Fattal, D. R .; Бен-Шауль, А. Биофиз. J. 1993, 65, 1795–1809.
- ^ Nezil, F.A .; Блум, М. Биофиз. J. 1992, 61, 1176-1183.
- ^ Куррле, А .; Rieber, P .; Сакманн, Э. Биохимия. 1990, 29, 8274-8282.
- ^ Zhang, Y .; Lewis, A.H .; Hodges, R. S .; МакЭлхейни, Р. Н. Биохимия. 1992, 31, 11579-11588.
- ^ Mouritsen, O.G .; Блум, М. Биофиз. J. 1984, 46, 141–153.
- ^ Mouritsen, O.G .; Биохим. Биофиз. Acta. 1998, 1376, 245-266,
- ^ Lewis, B.A .; Энгельман, Д. J. Mol. Биол. 1983, 166, 203-210.
- ^ Ryba, N.J .; Марш, Д. Биохимия 1992, 31, 7511-7518.
- ^ Кусуми, А .; Хайд, Дж. Biochemistry 1982, 21, 5978-5983.
- ^ Lewis, B.A .; Энгельман, Д. J. Mol. Биол. 1983, 166, 203-210.
- ^ Le Coutre, J .; Нарасимхан, Л. ; Kumar, C .; Patel, N .; Kaback, H.R. Proc. Natl. Акад. Sci. USA 1997, 94, 10167-10171.
- ^ Ren, J .; Lew, S .; Wang, Z .; Лондон, E. Biochemistry 1997, 36, 10213-10220.
- ^ Duque, D .; Li, X.J .; Кацов, К .; Schick, M. J. Chem. Phys. 2002, 116, 10478-10484.
- ^ Fattal, D. R .; Бен-Шауль, А. Биофиз. J. 1993, 65, 1795–1809.
- ^ Mouritsen, O.G .; Блум, М. Биофиз. J. 1984, 46, 141–153.
- ^ Sperotto, M. M .; Mouritsen, O.G. Eur. Биофиз. J. 1991, 19, 157–168.
- ^ Pink, D. A .; Chapman, D. Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 1979, 76, 1542–1546.
- ^ Fattal D. R .; Бен-Шауль, А. Биофиз. J. 1993, 65, 1795–1809
- ^ Duque, D .; Li, X.J .; Кацов, К .; Schick, M. J. Chem. Phys. 2002, 116, 10478–10484.
- ^ Норимацу, Ю., Хасэгава, К., Симидзу, Н., Тойосима, К. Природа 2017, 545, 193-198
- ^ Silvius, J.R .; McElhaney, R.N. Мочь. J. Biochem. 1978, 56, 462-469.
- ^ Bretscher, M.S .; Манро, S. Science 1993, 261, 1280-1281.
- ^ Манро, С. EMBO J. 1995, 14, 4659-4704.
- ^ Масибай А.С .; Balaji, P.V .; Boeggeman, E.E .; Касба, П. J. Biol. Chem. 1993, 268, 9908-9916.
- ^ Саймонс, К .; Тоомре, Д. Нат. Преподобный Мол. Cell Biol. 2000, 1, 31–39.
- ^ Саймонс, К .; Иконен, Э. Природа. 1997, 387, 569–572.
- ^ Саймонс, К .; Иконен, Э. Природа. 1997, 387, 569–572.
- ^ Thomas, J. L .; Holowka, D .; Baird, B .; Webb, W. W. J. Cell Biol. 1994, 125, 795–802.
- ^ Де Йонг, К .; Geldwerth, D .; Кайперс, Ф. А. Биохимия. 1997, 36, 6768–6776.
- ^ Едидин, М. Trends Cell Biol. 2001, 11, 492–496.
- ^ Андерсон, Р. Г. В .; Якобсон К. Наука. 2002, 296, 1821–1825.
- ^ Охво-Рекила, Х .; Ramstedt, B .; Leppimaki, P .; Slotte, J. P. Prog. Lipid Res. 2002, 41, 66–97.]
- ^ Ван Меер, Г. Наука. 2002, 296, 855–856.
- ^ McIntosh, T. J. Chem. Phys. Липиды. 2004, 130, 83–98.