Сотовая модель Поттса - Cellular Potts model
В вычислительная биология, а Сотовая модель Поттса (CPM, также известная как модель Глазье-Гранера-Хогевега) - это вычислительная модель клеток и тканей. Он используется для моделирования индивидуального и коллективного поведения клеток, тканей. морфогенез и рак разработка. CPM описывает ячейки как деформируемые объекты с определенным объемом, которые могут придерживаться друг к другу и к среде, в которой они живут. Формализм можно расширить, включив в него такие поведения клеток, как миграция клеток, рост и разделение, и клеточная сигнализация. Первый CPM был предложен для моделирования сортировка ячеек к Франсуа Гранер и Джеймс Глейзер как модификация большой Q Модель Поттса.[1] CPM тогда популяризировали Полин Хогевег для изучения морфогенеза.[2]Хотя модель была разработана для описания биологические клетки, его также можно использовать для моделирования отдельных частей биологической клетки или даже областей жидкости.
Описание модели
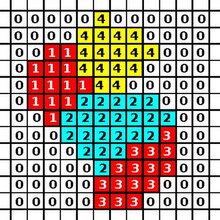
CPM состоит из прямоугольного Евклидово решетка, где каждая ячейка представляет собой подмножество узлов решетки, разделяющих одни и те же идентификатор ячейки (аналогично спину в моделях Поттса в физике). Средой являются участки решетки, не занятые клетками. Динамика модели определяется энергетической функцией: Гамильтониан который описывает энергию определенной конфигурации ячеек в решетке. В базовой CPM эта энергия является результатом адгезии между клетками и устойчивости клеток к изменениям объема. Алгоритм обновления CPM сводит к минимуму эту энергию.
Для развития модели Мегаполис -стилевые обновления выполняются, то есть:
- выбрать случайный узел решетки я
- выбрать случайный соседний узел решетки j скопировать его ID в я.
- вычислить разницу в энергии () между исходной и предлагаемой новой конфигурацией.
- принять или отклонить это событие копирования на основе изменения энергии , следующее:
- если новая энергия ниже, всегда принимайте копию;
- если новая энергия выше, примите копию с вероятностью (в Температура Больцмана Т определяет вероятность энергетически невыгодных колебаний.


Гамильтониан
Первоначальная модель, предложенная Гранером и Глейзером, содержит клетки двух типов, с разными энергиями адгезии для клеток одного типа и клеток другого типа. Каждый тип ячейки также имеет разную энергию контакта со средой, и предполагается, что объем ячейки остается близким к целевому значению. Гамильтониан формулируется как:

куда я, j узлы решетки, σя - ячейка в узле i, τ (σ) - тип ячейки ячейки σ, J - коэффициент, определяющий сцепление между двумя ячейками типов τ (σ), τ (σ '), δ - Дельта Кронекера, v (σ) - объем ячейки σ, V (σ) - целевой объем, λ - Множитель Лагранжа определение силы ограничения объема.
Клетки с более низким значением J для их мембранного контакта будут более прочно слипаться. Следовательно, можно моделировать различные модели сортировки ячеек, варьируя значения J.
Расширения
Со временем CPM превратилась из конкретной модели сортировки ячеек в общую структуру с множеством расширений, некоторые из которых частично или полностью являются внерешеточными.[3] Различное поведение клеток, например хемотаксис, удлинение и гаптотаксис можно включить, расширив гамильтониан H или изменение энергии . Вспомогательные подрешетки могут использоваться для включения дополнительной пространственной информации, такой как концентрации химических веществ.
Хемотаксис
В CPM клетки можно заставить двигаться в направлении более высоких хемокин концентрация, за счет увеличения вероятности копирования ID сайта j на сайт я когда концентрация хемокина выше при j. Это делается путем изменения изменения энергии со слагаемым, пропорциональным разнице концентраций при я и j:[2]

Где сила хемотаксического движения, и и представляют собой концентрацию хемокина в сайте i и j, соответственно. Градиент хемокина обычно реализуется на отдельной решетке тех же размеров, что и решетка клетки.



Мультимасштабное и гибридное моделирование с использованием CPM
Алгоритм Core GGH (или CPM), который определяет эволюцию структур клеточного уровня, может быть легко интегрирован с динамикой внутриклеточной сигнализации, динамикой диффузии реакций и моделью, основанной на правилах, для учета процессов, которые происходят в более низком (или более высоком) масштабе времени.[4] Программное обеспечение с открытым исходным кодом Bionetsolver можно использовать для интеграции внутриклеточной динамики с алгоритмом CPM.[5]
Рекомендации
- ^ Гранер, Франсуа; Стекольщик, Джеймс (1992). «Моделирование биологической сортировки клеток с использованием двумерной расширенной модели Поттса» (PDF). Phys. Rev. Lett. 69 (13): 2013–7. Bibcode:1992ПхРвЛ..69.2013Г. Дои:10.1103 / PhysRevLett.69.2013. PMID 10046374.
- ^ а б Сэвилл, Николас Дж .; Хогевег, Полин (1997). «Моделирование морфогенеза: от одиночных клеток до ползающих слизней». J. Theor. Биол. 184 (3): 229–235. Дои:10.1006 / jtbi.1996.0237. PMID 31940735.
- ^ Балтер, Ариэль; Merks, Roeland M.H .; Popławski, Nikodem J .; Сват, Мацей; Стекольщик, Джеймс А. (2007). «Модель Глейзера-Гранера-Хогевега: расширения, будущие направления и возможности для дальнейшего изучения». Одноклеточные модели в биологии и медицине. Математика и биологические науки во взаимодействии. С. 151–167. Дои:10.1007/978-3-7643-8123-3_7. ISBN 978-3-7643-8101-1.
- ^ Сабо, А; Меркс, РМ (2013). «Клеточное моделирование роста опухоли, инвазии опухоли и эволюции опухоли». Границы онкологии. 3. Дои:10.3389 / fonc.2013.00087. ЧВК 3627127. PMID 23596570.
- ^ Андасари, Виви; Ропер, Райан Т; Сват, Maciej H; Капеллан, Массачусетс (2012). «Интеграция внутриклеточной динамики с использованием CompuCell3D и Bionetsolver: приложения для многомасштабного моделирования роста и инвазии раковых клеток». PLOS ONE. 7 (3): e33726. Bibcode:2012PLoSO ... 733726A. Дои:10.1371 / journal.pone.0033726. ЧВК 3312894. PMID 22461894.
- Чен, Нан; Стекольщик, Джеймс А .; Izaguirre, Jesus A .; Альбер, Марк С. (2007). «Параллельная реализация модели Cellular Potts для моделирования клеточного морфогенеза». Компьютерная физика Коммуникации. 176 (11–12): 670–681. Bibcode:2007CoPhC.176..670C. Дои:10.1016 / j.cpc.2007.03.007. ЧВК 2139985. PMID 18084624.
внешняя ссылка
- Джеймс Глейзер (профессиональный сайт)
- CompuCell3D, среда моделирования CPM: Sourceforge
- SimTK
- Сайт разработки Notre Dame
- Модель искусственной жизни многоклеточного морфогенеза с автономно генерируемыми градиентами для позиционной информации с использованием модели Cellular Potts
- Стохастические клеточные автоматы