Рост клеток - Cell growth
Рост клеток относится к увеличение общего масса из клетка, включая оба цитоплазматический, ядерный и органелла объем.[1] Рост клеток происходит, когда общая скорость клеточного биосинтез (изготовление биомолекулы или анаболизм) больше, чем общая скорость клеточной деградации (разрушение биомолекулы через протеасома, лизосома или же аутофагия, или катаболизм).[2][3][4]
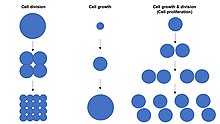
Рост клеток не следует путать с деление клеток или клеточный цикл, которые представляют собой отдельные процессы, которые могут происходить наряду с ростом клеток в процессе распространение клеток, где клетка, известная как «материнская клетка», растет и делится, чтобы произвести две »дочерние клетки ".[1] Важно отметить, что рост клеток и деление клеток также могут возникать независимо друг от друга. В начале эмбриональное развитие (расщепление из зигота сформировать морула и бластодерма ), деления клеток происходят многократно без роста клеток. И наоборот, некоторые клетки могут расти без деление клеток или без какой-либо прогрессии клеточный цикл, например, рост нейроны в течение аксональный поиск пути в нервная система разработка.
В многоклеточный организмы рост тканей редко происходит исключительно за счет роста клеток без деление клеток, но чаще всего происходит через распространение клеток.[1] Это потому, что одна ячейка только с одной копией геном в ядро клетки может выполнять биосинтез и, таким образом, рост клеток происходит вдвое быстрее, чем у двух клеток. Следовательно, две клетки растут (накапливают массу) в два раза быстрее, чем одна клетка, а четыре клетки растут в 4 раза быстрее, чем одна клетка. Этот принцип приводит к экспоненциальный увеличение в рост тканей скорость (накопление массы) во время пролиферации клеток за счет экспоненциальный увеличение количества ячеек.
Размер клетки зависит как от роста клеток, так и от деление клеток с непропорциональным увеличением скорости роста клеток, приводящим к образованию более крупных клеток, и непропорциональным увеличением скорости деления клеток, ведущим к образованию множества более мелких клеток. Распространение клеток обычно включает сбалансированный рост клеток и деление клеток скорости, которые поддерживают примерно постоянный размер клеток в экспоненциально пролиферирующей популяции клеток.
Некоторые особые клетки могут вырасти до очень больших размеров за счет необычного "эндорепликация "клеточный цикл, в котором геном тиражируется во время S-фаза но нет последующего митоза (M-фаза ) или деление клеток (цитокинез ). Эти большие эндорепликация клетки имеют много копий геном, так что очень полиплоид.
Ооциты могут быть необычно большими клетками у видов, у которых эмбриональное развитие происходит вдали от тела матери в яйце, откладываемом извне. Большого размера некоторых яиц можно достичь либо путем перекачки цитозольных компонентов из соседних клеток через цитоплазматические мостики, называемые кольцевыми каналами (Дрозофила ) или интернализацией гранул, хранящих питательные вещества (гранулы желтка), посредством эндоцитоз (лягушки ).
Механизмы контроля роста клеток
Клетки может расти за счет увеличения общего количества сотовых биосинтез такое, что производство биомолекулы превышает общую скорость клеточной деградации биомолекулы через протеасома, лизосома или же аутофагия.
Биосинтез из биомолекулы инициируется выражением гены которые кодируют РНК и / или белки, включая ферменты которые катализируют синтез липиды и углеводы.
Индивидуальный гены обычно выразил через транскрипция в информационная РНК (мРНК) и перевод в белки, и экспрессия каждого гена происходит на разных уровнях в зависимости от типа клетки (в ответ на сети регуляции генов ).
Чтобы стимулировать рост клеток, можно увеличить глобальную скорость экспрессии генов. за счет увеличения общей скорости транскрипция к РНК-полимераза II (для активных генов) или общая скорость мРНК перевод в белок за счет увеличения обилия рибосомы и тРНК, чей биогенез зависит от РНК-полимераза I и РНК-полимераза III. В Мой с фактор транскрипции является примером регуляторного белка, который может индуцировать общую активность РНК-полимераза I, РНК-полимераза II и РНК-полимераза III управлять глобальным транскрипция и перевод и тем самым рост клеток.
Кроме того, активность отдельных рибосомы могут быть увеличены для повышения глобальной эффективности мРНК перевод посредством регуляции факторов инициации трансляции, включая «фактор инициации удлинения трансляции 4E» (eIF4E ) комплекс, который связывает и закрывает 5'-конец мРНК. Протеин TOR, часть TORC1 комплекс, является важным регулятором перевод инициация, а также биогенез рибосом.[5] TOR представляет собой серин / треонин киназа которые могут непосредственно фосфорилировать и инактивировать общий ингибитор eIF4E, названный 4E-связывающий белок (4E-BP), чтобы повысить эффективность перевода. TOR также непосредственно фосфорилирует и активирует рибосомный протеин S6-киназу (S6K ), что способствует биогенез рибосом.
Чтобы подавить рост клеток, можно уменьшить глобальную скорость экспрессии генов или глобальную скорость биомолекулярный деградация может быть увеличена за счет увеличения скорости аутофагия. TOR обычно напрямую подавляет функцию аутофагия индуцирование киназы Atg1 / ULK1. Таким образом, уменьшая TOR активность снижает глобальную скорость перевод и увеличивает степень аутофагия для уменьшения роста клеток.
Регуляция роста клеток у животных
Многие сигнальные молекулы, контролирующие рост клеток, называются факторы роста, многие из которых вызывают преобразование сигнала через Путь PI3K / AKT / mTOR, который включает предшествующую липидкиназу PI3K и нижестоящий серин / треониновый белок киназа Акт, который способен активировать другую протеинкиназу TOR, который продвигает перевод и подавляет аутофагия для стимулирования роста клеток.
Доступность питательных веществ влияет на производство факторы роста из Инсулин /IGF-1 семьи, которые циркулируют в организме животных в виде гормонов для активации Путь PI3K / AKT / mTOR в камерах для продвижения TOR активность, так что, когда животные хорошо питаются, они будут расти быстро, а когда они не могут получать достаточное количество питательных веществ, они уменьшат скорость своего роста.
Кроме того, наличие аминокислоты в отдельные клетки также напрямую способствует TOR активности, хотя этот способ регуляции более важен для одноклеточных организмов, чем для многоклеточный организмы, такие как животные, которые всегда поддерживают изобилие аминокислоты в обращении.
Одна спорная теория предполагает, что многие различные клетки млекопитающих претерпевают переходы в зависимости от размера во время клеточного цикла. Эти переходы контролируются циклин-зависимой киназой Cdk1.[6] Хотя белки, которые контролируют Cdk1, хорошо изучены, их связь с механизмами, контролирующими размер клеток, остается неуловимой. Постулируемая модель контроля размера млекопитающих рассматривает массу как движущую силу клеточного цикла. Клетка не может вырасти до аномально большого размера, потому что при определенном размере клетки или клеточной массе инициируется S-фаза. Фаза S запускает последовательность событий, ведущих к митозу и цитокинезу. Клетка не может стать слишком маленькой, потому что более поздние события клеточного цикла, такие как S, G2 и M, задерживаются до тех пор, пока масса не увеличится достаточно, чтобы начать S-фазу.[7]
Популяции клеток
Популяции клеток проходят через определенный тип экспоненциальный рост называется удвоением или распространение клеток. Таким образом, каждый поколение ячеек должно быть вдвое больше, чем у предыдущего поколения. Однако количество поколений дает только максимальную цифру, поскольку не все клетки выживают в каждом поколении. Клетки могут воспроизводиться на стадии митоза, когда они удваиваются и разделяются на две генетически равные клетки.
Размер ячейки
Размер клеток у разных организмов сильно различается, например, у некоторых водорослей. Caulerpa taxifolia являясь одиночной камерой длиной несколько метров.[8] Растительные клетки намного больше, чем клетки животных, и простейшие, такие как Парамеций может быть 330 мкм в длину, в то время как типичная человеческая клетка может иметь длину 10 мкм. Как эти клетки «решают», какого размера они должны быть перед делением, остается открытым вопросом. Известно, что отчасти ответственны химические градиенты, и предполагается, что обнаружение механического напряжения путем цитоскелет структур. Работа над этой темой обычно требует организма, клеточный цикл которого хорошо охарактеризован.
Регулирование размера дрожжевых клеток
Связь между размером ячейки и деление клеток был широко изучен в дрожжи. Для некоторых клеток существует механизм, по которому деление клеток не начинается, пока клетка не достигнет определенного размера. Если подача питательных веществ ограничена (после времени t = 2 на диаграмме ниже), и скорость увеличения размера клеток замедляется, период времени между делениями клеток увеличивается.[9] Были выделены мутанты размером с дрожжевые клетки, которые начинают деление клеток до достижения нормального / обычного размера (крошечный мутанты).[10]

Wee1 белок - это тирозинкиназа который в норме фосфорилирует регуляторный белок клеточного цикла Cdc2 (гомолог CDK1 у человека), циклин-зависимая киназа, на остатке тирозина. Cdc2 запускает митоз путем фосфорилирования широкого круга мишеней. Этот ковалентный модификация молекулярной структуры Cdc2 подавляет ферментативную активность Cdc2 и предотвращает деление клеток. Wee1 удерживает Cdc2 неактивным во время раннего G2 когда клетки еще маленькие. Когда клетки достигают достаточного размера во время G2, фосфатаза Cdc25 устраняет ингибирующее фосфорилирование и, таким образом, активирует Cdc2, чтобы разрешить митотический вход. Баланс активности Wee1 и Cdc25 с изменениями размера клеток координируется системой контроля митотического входа. На мутантах Wee1, клетках с ослабленной активностью Wee1, было показано, что Cdc2 становится активным, когда клетка меньше. Таким образом, митоз происходит до того, как дрожжи достигают своего нормального размера. Это предполагает, что деление клеток может частично регулироваться разбавлением белка Wee1 в клетках по мере их роста.
Связывание Cdr2 с Wee1
Протеинкиназа Cdr2 (который негативно регулирует Wee1) и родственную Cdr2 киназу Cdr1 (который непосредственно фосфорилирует и ингибирует Wee1 in vitro)[11] локализуются в связке корковых узлов в середине интерфазных клеток. После вступления в митоз факторы цитокинеза, такие как миозин II набираются в аналогичные узлы; эти узлы в конечном итоге конденсируются, образуя цитокинетический звенеть.[12] Ранее не охарактеризованный белок, Blt1, было обнаружено, что они колокализуются с Cdr2 в медиальных интерфазных узлах. Клетки с нокаутом Blt1 имели увеличенную длину при делении, что согласуется с задержкой митотического входа. Это открытие связывает физическое местоположение, полосу корковых узлов, с факторами, которые, как было показано, непосредственно регулируют митотический вход, а именно Cdr1, Cdr2 и Blt1.
Дальнейшие эксперименты с GFP -маркированные белки и мутантные белки указывают на то, что медиальные кортикальные узлы сформированы упорядоченной, зависимой от Cdr2 сборкой множества взаимодействующих белков во время интерфазы. Cdr2 находится на вершине этой иерархии и работает выше Cdr1 и Blt1.[13] Митозу способствует негативная регуляция Wee1 с помощью Cdr2. Также было показано, что Cdr2 рекрутирует Wee1 в медиальный кортикальный узел. Механизм этой вербовки еще предстоит выяснить. Мутант киназы Cdr2, который способен правильно локализоваться, несмотря на потерю функции фосфорилирования, нарушает рекрутирование Wee1 в медиальную кору и задерживает вступление в митоз. Таким образом, Wee1 локализуется со своей ингибирующей сетью, что демонстрирует, что митоз контролируется посредством Cdr2-зависимой негативной регуляции Wee1 в медиальных кортикальных узлах.[13]
Факторы полярности клеток
Факторы полярности клеток, расположенные на концах клеток, обеспечивают пространственные ориентиры для ограничения распределения Cdr2 в середине клетки. В делящихся дрожжах Schizosaccharomyces pombe (С. Помбе), клетки делятся до определенного воспроизводимого размера во время митоза из-за регулируемой активности Cdk1.[14] Протеинкиназа клеточной полярности Пом1, член семейства киназ, регулируемых фосфорилированием тирозина (DYRK) с двойной специфичностью, локализуется на концах клеток. В клетках, нокаутированных по Pom1, Cdr2 больше не ограничивался серединой клетки, а диффузно наблюдался через половину клетки. Из этих данных становится очевидным, что Pom1 обеспечивает тормозящие сигналы, которые ограничивают Cdr2 серединой клетки. Далее было показано, что Pom1-зависимые сигналы приводят к фосфорилированию Cdr2. Было также показано, что клетки с нокаутом Pom1 делятся с меньшим размером, чем клетки дикого типа, что указывает на преждевременное вступление в митоз.[13]
Pom1 формирует полярные градиенты, которые достигают максимума на концах ячейки, что показывает прямую связь между факторами контроля размера и конкретным физическим местоположением в ячейке.[15] По мере увеличения размера клетки градиент в Pom1 увеличивается. Когда клетки маленькие, Pom1 распространяется диффузно по телу клетки. По мере увеличения размера ячейки концентрация Pom1 уменьшается в середине и становится концентрированной на концах ячейки. Маленькие клетки в ранней стадии G2, которые содержат достаточные уровни Pom1 во всей клетке, имеют неактивный Cdr2 и не могут вступать в митоз. Только когда клетки перерастут в поздний G2, когда Pom1 ограничен клеточными концами, Cdr2 в медиальных кортикальных узлах активируется и может начать ингибирование Wee1. Это открытие показывает, как размер клетки играет прямую роль в регуляции начала митоза. В этой модели Pom1 действует как молекулярное звено между ростом клеток и митотическим входом через путь Cdr2-Cdr1-Wee1-Cdk1.[13] Полярный градиент Pom1 успешно передает информацию о размере и геометрии клеток регуляторной системе Cdk1. Благодаря этому градиенту клетка гарантирует, что она достигла определенного, достаточного размера для вступления в митоз.
Другие экспериментальные системы для изучения регуляции размера клеток
Одним из распространенных способов производства очень больших клеток является слияние клеток с образованием синцития. Например, очень длинные (несколько дюймов) скелетные мышцы клетки образуются путем слияния тысяч миоциты. Генетические исследования плодовой мушки Дрозофила выявили несколько генов, которые необходимы для образования многоядерных мышечных клеток путем слияния миобласты.[16] Некоторые из ключевых белков важны для клеточная адгезия между миоцитами, а некоторые из них участвуют в зависимой от адгезии межклеточной преобразование сигнала что допускает каскад событий слияния клеток. клетки растений осложняются тем, что практически все растительные клетки находятся внутри твердого клеточная стенка. Под влиянием некоторых гормонов растений клеточная стенка может быть реконструирована, что позволяет увеличить размер клеток, что важно для роста некоторых тканей растений.
Большинство одноклеточных организмов имеют микроскопические размеры, но есть и гигантские бактерии и простейшие которые видны невооруженным глазом. Видеть: Таблица размеров ячеек —Плотные популяции гигантской серной бактерии в отложениях шельфа Намибии[17]— Крупные протисты рода Хаос, близкородственный к роду Амеба
В палочковидных бактериях Кишечная палочка, Caulobacter crescentus и Б. subtilis Размер клеток контролируется простыми механизмами, в которых деление клеток происходит после добавления постоянного объема с момента предыдущего деления.[18][19] Всегда растя на одну и ту же величину, клетки, рожденные меньше или больше среднего, естественным образом сходятся к среднему размеру, эквивалентному количеству, добавляемому в течение каждого поколения.
Деление клеток
Размножение клеток бесполый. Для большинства составляющих клетки рост - это устойчивый, непрерывный процесс, прерывающийся лишь на короткое время при Фаза M когда ядро, а затем и клетка делятся на две части.
Процесс деления клеток, называемый клеточный цикл, состоит из четырех основных частей, называемых фазами. Первая часть, называемая грамм1 фаза отмечен синтезом различных ферменты которые необходимы для репликации ДНК. Вторая часть клеточного цикла - это Фаза S, куда Репликация ДНК производит два идентичных набора хромосомы. Третья часть - это грамм2 фаза в котором значительный синтез белка происходит, в основном связанное с производством микротрубочки которые требуются в процессе разделения, называемые митоз Четвертая фаза, Фаза M, состоит из ядерного подразделения (кариокинез ) и цитоплазматическое деление (цитокинез ), сопровождающийся образованием нового клеточная мембрана. Это физическое разделение «материнской» и «дочерней» клеток. Фаза M была разбита на несколько отдельных фаз, последовательно известных как профаза, прометафаза, метафаза, анафаза и телофаза приводящий к цитокинезу.
Клеточное деление более сложное в эукариоты чем у других организмов. Прокариотический клетки, такие как бактериальный клетки воспроизводятся двойное деление, процесс, который включает репликацию ДНК, сегрегацию хромосом и цитокинез. Деление эукариотических клеток включает митоз или более сложный процесс, называемый мейоз. Митоз и мейоз иногда называют двумя "ядерный деление ». Двоичное деление похоже на размножение эукариотических клеток, которое включает митоз. Оба приводят к образованию двух дочерних клеток с тем же числом хромосом, что и родительская клетка. Мейоз используется для особого процесса воспроизводства клеток диплоид организмы. Он производит четыре особые дочерние клетки (гаметы ), которые содержат половину нормального клеточного количества ДНК. А мужчина и женский затем гамета может объединиться, чтобы произвести зигота, клетка, которая снова имеет нормальное количество хромосом.
Остальная часть статьи представляет собой сравнение основных характеристик трех типов клеточного воспроизводства, которые включают бинарное деление, митоз или мейоз. На диаграмме ниже показаны сходства и различия этих трех типов размножения клеток.

Сравнение трех типов деления клеток
Содержимое ДНК клетки дублируется в начале процесса воспроизводства клеток. До Репликация ДНК, содержание ДНК в клетке можно представить как количество Z (в клетке есть Z-хромосомы). После процесса репликации ДНК количество ДНК в клетке равно 2Z (умножение: 2 x Z = 2Z). Во время бинарного деления и митоза дублированное содержимое ДНК воспроизводящей родительской клетки разделяется на две равные половины, которым суждено попасть в две дочерние клетки. Заключительная часть процесса размножения клеток - это деление клеток, когда дочерние клетки физически отделяются от родительской клетки. Во время мейоза происходит два этапа деления клеток, которые вместе производят четыре дочерние клетки.
После завершения бинарного деления или размножения клеток с участием митоза каждая дочерняя клетка имеет одинаковое количество ДНК (Z) как то, что было у родительской клетки до репликации своей ДНК. Эти два типа воспроизводства клеток дали две дочерние клетки, которые имеют такое же количество хромосом, что и родительская клетка. Хромосомы дублируются до деления клеток при формировании новых клеток кожи для воспроизводства. После размножения мейотических клеток четыре дочерние клетки имеют половину количества хромосом, которое изначально было у родительской клетки. Это гаплоидный количество ДНК, часто обозначаемое как N. Мейоз используется диплоид организмы для производства гаплоидных гамет. В диплоидном организме, таком как человеческий организм, большинство клеток тела имеют диплоидное количество ДНК, 2N. Используя это обозначение для подсчета хромосом, мы говорим, что человеческий соматический клетки имеют 46 хромосом (2N = 46) в то время как человек сперма и яйца имеют 23 хромосомы (N = 23). У людей есть 23 различных типа хромосом, из которых 22 аутосомы и особая категория половые хромосомы. Есть две различные половые хромосомы, Х-хромосома и Y-хромосома. Диплоидная человеческая клетка имеет 23 хромосомы от отца этого человека и 23 от матери. То есть в вашем теле есть две копии хромосомы номер 2 человека, по одной от каждого из ваших родителей.

Сразу после репликации ДНК в клетке человека будет 46 «двойных хромосом». В каждой двойной хромосоме есть две копии молекулы ДНК этой хромосомы. Во время митоза двойные хромосомы расщепляются с образованием 92 «одиночных хромосом», половина из которых попадает в каждую дочернюю клетку. Во время мейоза существует два этапа разделения хромосом, которые гарантируют, что каждая из четырех дочерних клеток получит по одной копии каждого из 23 типов хромосом.
Половое размножение
Хотя размножение клеток, использующее митоз, может воспроизводить эукариотические клетки, эукариоты беспокоятся о более сложном процессе мейоза, потому что половое размножение например, мейоз дает селективное преимущество. Обратите внимание, что когда начинается мейоз, две копии сестринских хроматид номер 2 соседствуют друг с другом. За это время может быть генетическая рекомбинация События. Информация из ДНК хромосомы 2, полученная от одного родителя (красный цвет), будет передаваться молекуле ДНК хромосомы 2, полученной от другого родителя (зеленый цвет). Обратите внимание, что в митозе две копии хромосомы номер 2 не взаимодействуют. Рекомбинация генетической информации между гомологичными хромосомами в течение мейоз это процесс для восстановление повреждений ДНК. Этот процесс может также производить новые комбинации генов, некоторые из которых могут быть адаптивно полезными и влиять на ход эволюции. Однако у организмов с более чем одним набором хромосом на основной стадии жизненного цикла пол также может дать преимущество, потому что при случайном спаривании он производит гомозиготы и гетерозиготы согласно Отношение Харди – Вайнберга.
Расстройства
На клеточном уровне может произойти ряд нарушений роста, которые, следовательно, лежат в основе большей части последующего течения болезни. рак, в которых группа клеток демонстрирует неконтролируемый рост и деление за пределы нормы, вторжение (вторжение и разрушение прилегающих тканей), а иногда метастаз (распространяется на другие части тела через лимфу или кровь). Несколько ключевых детерминант роста клеток, например плоидность и регуляция клеточного метаболизм, обычно нарушаются в опухоли.[20] Следовательно, гетерогенный рост клеток и плеоморфизм является одним из самых ранних признаков рак прогрессия.[21][22] Несмотря на преобладание плеоморфизма в патологии человека, его роль в прогрессировании заболевания неясна. В эпителиальный В тканях плеоморфизм клеточного размера может вызывать дефекты упаковки и диспергировать аберрантные клетки.[23] Но последствия атипичного роста клеток в других тканях животных неизвестны.
Методы измерения
Рост клеток можно обнаружить разными методами. рост размера клетки можно визуализировать микроскопия, используя подходящие морилки. Но увеличение количества ячеек обычно более значительна. Его можно измерить путем ручного подсчета клеток под микроскопическим наблюдением, используя метод исключения красителя (т. Е. трипановый синий ) для подсчета только жизнеспособных клеток. Менее требовательные, масштабируемые методы включают использование цитометры, пока проточной цитометрии позволяет комбинировать количество клеток («события») с другими конкретными параметрами: флуоресцентные зонды для мембран, цитоплазмы или ядер позволяют различать мертвые / жизнеспособные клетки, типы клеток, дифференцировку клеток, экспрессию биомаркер Такие как Ki67.
Помимо увеличения количества ячеек, можно оценить рост метаболической активности, это CFDA и кальцеин -AM измеряет (флуориметрически) не только функциональность мембраны (удерживание красителя), но также функциональность цитоплазматических ферментов (эстераз). В МТТ анализы (колориметрический) и Ресазурин анализ (флуориметрический) дозирования окислительно-восстановительного потенциала митохондрий.
Все эти анализы могут хорошо коррелировать или нет, в зависимости от условий роста клеток и желаемых аспектов (активности, пролиферации). Задача еще более усложняется с популяциями разных клеток, к тому же при объединении помех роста клеток или токсичность.
Смотрите также
- Бактериальный рост
- Двойное деление
- Клеточный цикл
- Клон (генетика)
- Биология развития
- Мейоз
- Митоз
- Плеоморфизм
- Стволовая клетка
Рекомендации
- ^ а б c Конлон, Ян; Рафф, Мартин (1999). «Контроль размера в развитии животных». Клетка. 96 (2): 235–244. Дои:10.1016 / S0092-8674 (00) 80563-2. ISSN 0092-8674. PMID 9988218. S2CID 15738174.
- ^ Grewal, Savraj S; Эдгар, Брюс А. (2003). «Контроль деления клеток у дрожжей и животных: имеет ли значение размер?». Журнал биологии. 2 (1): 5. Дои:10.1186/1475-4924-2-5. ISSN 1475-4924. ЧВК 156596. PMID 12733996.
- ^ Neufeld, Thomas P; де ла Крус, Аида Флора А; Джонстон, Лаура А; Эдгар, Брюс А. (1998). «Координация роста и деления клеток в крыле дрозофилы». Клетка. 93 (7): 1183–1193. Дои:10.1016 / S0092-8674 (00) 81462-2. ISSN 0092-8674. PMID 9657151. S2CID 14608744.
- ^ Томпсон, Барри Дж. (2010). «Контроль развития роста и деления клеток у дрозофилы». Текущее мнение в области клеточной биологии. 22 (6): 788–794. Дои:10.1016 / j.ceb.2010.08.018. PMID 20833011.
- ^ Хафен, Э. (2004). «Взаимодействие между фактором роста и питательными веществами: уроки TOR дрозофилы». TOR. Актуальные темы микробиологии и иммунологии. 279. С. 153–167. Дои:10.1007/978-3-642-18930-2_10. ISBN 978-3-642-62360-8. ISSN 0070-217X. PMID 14560957.
- ^ Митчисон Дж. М. (2003). «Рост во время клеточного цикла». Int. Преподобный Цитол. Международный обзор цитологии. 226: 165–258. Дои:10.1016 / S0074-7696 (03) 01004-0. ISBN 978-0-12-364630-9. PMID 12921238.
- ^ Купер, Стивен (2004). «Контроль и поддержание размера клеток млекопитающих». BMC Cell Biology. 5 (1): 35. Дои:10.1186/1471-2121-5-35. ЧВК 524481. PMID 15456512.
- ^ Пеплоу, Марк (23 марта 2005 г.). «Водоросли создают клей для восстановления повреждений клеток». Nature.com. Получено 4 июля 2016.
- ^ Славов Н .; Ботштейн Д. (июнь 2011 г.). «Связь между реакцией скорости роста, метаболическим циклом и циклом деления клеток в дрожжах». Молекулярная биология клетки. 22 (12): 1997–2009. Дои:10.1091 / mbc.E11-02-0132. ЧВК 3113766. PMID 21525243.
- ^ Wee1 мутанты С. Помбе имеют маленький размер ячейки и гомологичные белки у людей также регулируют вступление клетки в митоз; в Лодиш Х.Ф., Берк А., Зипурски Л.С., Мацудаира П. и др., Ред. (2000). Молекулярная клеточная биология (4-е изд.). Нью-Йорк: W.H. Фримен. ISBN 978-0-7167-3136-8.
- ^ Ву Л., Рассел П. (июнь 1993 г.). «Киназа Nim1 способствует митозу, инактивируя тирозинкиназу Wee1». Природа. 363 (6431): 738–41. Bibcode:1993Натура.363..738Вт. Дои:10.1038 / 363738a0. PMID 8515818. S2CID 4320080.
- ^ Ву Дж.К., Кун-младший, Ковар Д.Р., Поллард Т.Д. (ноябрь 2003 г.). «Пространственный и временной путь сборки и сжатия сократительного кольца при цитокинезе делящихся дрожжей». Dev. Клетка. 5 (5): 723–34. Дои:10.1016 / S1534-5807 (03) 00324-1. PMID 14602073.
- ^ а б c d Мозли Дж. Б., Майё А., Паолетти А., медсестра П. (июнь 2009 г.). «Пространственный градиент координирует размер клетки и митотический вход у делящихся дрожжей». Природа. 459 (7248): 857–60. Bibcode:2009Натура.459..857М. Дои:10.1038 / природа08074. PMID 19474789. S2CID 4330336.
- ^ Rupes I (сентябрь 2002 г.). «Проверка размера клеток дрожжей». Тенденции Genet. 18 (9): 479–85. Дои:10.1016 / S0168-9525 (02) 02745-2. PMID 12175809.
- ^ Падте Н.Н., Мартин С.Г., Ховард М., Чанг Ф. (декабрь 2006 г.). «Фактор конца клетки pom1p ингибирует mid1p в спецификации плоскости клеточного деления у делящихся дрожжей». Curr. Биол. 16 (24): 2480–7. Дои:10.1016 / j.cub.2006.11.024. PMID 17140794.
- ^ Менон С.Д., Осман З., Ченчилл К., Чиа В. (июнь 2005 г.). «Положительная обратная связь между Dumbfounded и Rolling pebbles приводит к увеличению мышечной трубки у дрозофилы». J. Cell Biol. 169 (6): 909–20. Дои:10.1083 / jcb.200501126. ЧВК 2171639. PMID 15955848.
- ^ Schulz HN, Brinkhoff T, Ferdelman TG, Mariné MH, Teske A, Jorgensen BB (апрель 1999 г.). «Плотные популяции гигантской серной бактерии в отложениях шельфа Намибии». Наука. 284 (5413): 493–5. Bibcode:1999Научный ... 284..493С. Дои:10.1126 / science.284.5413.493. PMID 10205058. S2CID 32571118.
- ^ Тахери-Араги, S; Брэдд, S; Sauls, J. T .; Hill, N. S .; Левин, П. А .; Paulsson, J; Вергассола, М; Июнь, С. (февраль 2015 г.). «Контроль размера клеток и гомеостаз у бактерий». Текущая биология. 25 (3): 385–391. Дои:10.1016 / j.cub.2014.12.009. ЧВК 4323405. PMID 25544609.
- ^ Кампос, М; Суровцев, И. В .; Като, S; Пайнтдахи, А; Beltran, B; Ebmeier, S.E .; Джейкобс-Вагнер, К. (декабрь 2014 г.). «Расширение постоянного размера способствует гомеостазу размера бактериальных клеток». Клетка. 159 (6): 1433–1446. Дои:10.1016 / j.cell.2014.11.022. ЧВК 4258233. PMID 25480302.
- ^ Schmoller, Kurt M .; Скотейм, Ян М. (декабрь 2015 г.). «Биосинтетические основы контроля размера клеток». Тенденции Cell Biol. 25 (12): 793–802. Дои:10.1016 / j.tcb.2015.10.006. ЧВК 6773270. PMID 26573465.
- ^ Трэвис, W.D .; Brambilla, B .; Берк, А.П .; Маркс, А .; Николсон, А.Г. (2015). Классификация ВОЗ опухолей легких, плевры, тимуса и сердца. Лион: Международное агентство по изучению рака. ISBN 978-92-832-2436-5.
- ^ Эль-Наггар, А.К .; Chan, J.C.K .; Grandis, J.R .; Takata, T .; Slootweg, P.J. (2017-01-23). Классификация опухолей головы и шеи ВОЗ. Лион: Международное агентство по изучению рака. ISBN 978-92-832-2438-9. Архивировано из оригинал на 2019-10-31. Получено 2019-10-31.
- ^ Ramanathan, Subramanian P .; Крайнц, Матей; Гибсон, Мэтью С. (октябрь 2019 г.). «Плеоморфизм размера клетки приводит к аберрантному рассредоточению клонов в пролиферирующем эпителии». Клетка развития. 51 (1): 49–61.e4. Дои:10.1016 / j.devcel.2019.08.005. ЧВК 6903429. PMID 31495693.
Книги
- Морган, Дэвид О. (2007). Клеточный цикл: принципы контроля. Лондон: Сандерленд, Массачусетс. ISBN 978-0-9539181-2-6.
внешняя ссылка
- Сравнение поколенческой и экспоненциальной моделей роста клеточной популяции
- Локальный рост в массиве дисков Вольфрам Демонстрационный проект.
Результат изображения для роста клеток
Рост клеток (или интерфаза) - это сокращение от идеи «роста популяций клеток» посредством размножения клеток. Это этап, на котором клетки готовятся к следующему делению, происходят биохимические активности и реакции, однако на этом этапе не видно никаких явных изменений.