Органелла - Organelle
| Органелла | |
|---|---|
| Подробности | |
| Произношение | /ɔːrɡəˈпɛл/ |
| Часть | Клетка |
| Идентификаторы | |
| латинский | Органелла |
| MeSH | D015388 |
| TH | H1.00.01.0.00009 |
| FMA | 63832 |
| Анатомические термины микроанатомии | |
В клеточная биология, органелла является специализированным подразделением, обычно внутри клетка, имеющий определенную функцию. Название органелла исходит из идеи, что эти структуры являются частями клеток, так как органы относятся к тело, следовательно органелла суффикс -elle быть миниатюрный. Органеллы либо отдельно заключены в свои собственные липидные бислои (также называемые мембраносвязанными органеллами) или пространственно отдельные функциональные единицы без окружающего липидного бислоя (органеллы, не связанные с мембраной). Хотя большинство органелл являются функциональными единицами внутри клеток, некоторые функциональные единицы, выходящие за пределы клеток, часто называют органеллами, например реснички, то жгутик и архаеллум, а трихокиста.
Органеллы идентифицируются по микроскопия, а также может быть очищен фракционирование клеток. Существует много типов органелл, особенно в эукариотические клетки. К ним относятся конструкции, составляющие внутренние эндомембранная система (такой как ядерная оболочка, эндоплазматический ретикулум, и аппарат Гольджи ) и другие структуры, такие как митохондрии и пластиды. Пока прокариоты не обладают эукариотическими органеллами, некоторые содержат белок -очищенный бактериальные микрокомпартменты, которые считаются примитивными прокариотические органеллы;[1] есть также свидетельства наличия других структур, связанных с мембраной.[2] Также прокариотический жгутик который выступает за пределы клетки и ее мотора, а также в значительной степени внеклеточный пилус, часто называют органеллами.
История и терминология
| Клеточная биология | |
|---|---|
| В животная клетка | |
 Компоненты типичной животной клетки:
|
В биологии органы определяются как ограниченные функциональные единицы внутри организм.[3] В аналогия от органов тела до микроскопических клеточных субструктур очевидно, поскольку даже в ранних работах авторы соответствующих учебников редко уточняют различие между ними.
В 1830-х годах Феликс Дюжарден опровергнутый Эренберг теория, которая гласила, что микроорганизмы имеют те же органы, что и у многоклеточных животных, только второстепенные.[4]
Признан первым[5][6][7] использовать миниатюрный из орган (то есть маленький орган) для клеточных структур был немецкий зоолог Карл Август Мёбиус (1884), который использовал термин органула (множественное число от орган, уменьшительное от латинский орган).[8] В сноске, которая была опубликована в качестве исправления в следующем номере журнала, он обосновал свое предложение называть органы одноклеточных организмов «органеллами», поскольку они являются лишь по-разному сформированными частями одной клетки, в отличие от многоклеточных органов многоклеточных организмов. .[8][9]
Типы
Хотя большинство клеточных биологов считают, что термин органелла быть синонимом отсек для кювет, пространство, часто ограниченное одним или двумя липидными бислоями, некоторые клеточные биологи предпочитают ограничить этот термин включением только тех клеточных компартментов, которые содержат дезоксирибонуклеиновая кислота (ДНК), произошедшая от ранее автономных микроскопических организмов, приобретенных через эндосимбиоз.[10][11][12]
Согласно этому определению, может быть только два широких класса органелл (то есть те, которые содержат свою собственную ДНК и происходят из эндосимбиотических организмов). бактерии ):
- митохондрии (почти у всех эукариот)
- пластиды[13] (например, в растениях, водорослях и некоторых простейших).
Предполагается, что другие органеллы имеют эндосимбиотическое происхождение, но не содержат собственной ДНК (особенно жгутика - см. эволюция жгутиков ).
Второе, менее ограничительное определение органелл состоит в том, что они являются мембраносвязанными структурами. Однако даже при использовании этого определения некоторые части клетки, которые, как было показано, являются отдельными функциональными единицами, не квалифицируются как органеллы. Поэтому использование органелл для обозначения не связанных с мембранами структур, таких как рибосомы, является обычным и приемлемым.[14][15][16] Это привело к тому, что многие тексты разграничили мембраносвязанный и немембранный связанные органеллы.[17] Органеллы, не связанные с мембраной, также называемые большими биомолекулярные комплексы, большие собрания макромолекулы которые выполняют определенные и специализированные функции, но у них отсутствуют мембранные границы. Многие из них называются «белковые органеллы», поскольку многие структуры состоят из белков. К таким клеточным структурам относятся:
- большие РНК и белковые комплексы: рибосома, сплайсосома, свод
- большой белковые комплексы: протеасома, Холофермент ДНК-полимераза III, Холофермент РНК-полимеразы II, симметричный вирусный капсиды, комплекс GroEL и GroES; мембранные белковые комплексы: фотосистема I, АТФ-синтаза
- большие комплексы ДНК и белков: нуклеосома
- центриоль и центр организации микротрубочек (MTOC)
- цитоскелет
- жгутик
- ядрышко
- стрессовая гранула
- половая клетка гранула
- нейрональная транспортная гранула
Механизмы, с помощью которых такие органеллы, не связанные с мембраной, образуются и сохраняют свою пространственную целостность, можно сравнить с жидко-жидким. разделение фаз.[18]
Эукариотические органеллы
Эукариотический клетки имеют сложную структуру и по определению частично организованы внутренними отсеками, которые сами по себе окружены липидными мембранами, которые напоминают самые внешние клеточная мембрана. Более крупные органеллы, такие как ядро и вакуоли, хорошо видны оптический микроскоп. Они были одними из первых биологических открытий, сделанных после изобретения микроскоп.
Не все эукариотические клетки имеют каждую из перечисленных ниже органелл. У исключительных организмов есть клетки, которые не содержат некоторых органелл, которые в противном случае могли бы считаться универсальными для эукариот (например, митохондрии).[19] Есть также случайные исключения из числа мембран, окружающих органеллы, перечисленных в таблицах ниже (например, некоторые из них, перечисленные как двухмембранные, иногда встречаются с одинарными или тройными мембранами). Кроме того, количество отдельных органелл каждого типа, обнаруженных в данной клетке, варьируется в зависимости от функции этой клетки.
| Органелла | Основная функция | Структура | Организмы | Примечания |
|---|---|---|---|---|
| клеточная мембрана | отделяет внутренность всех клеток от внешней среды (внеклеточное пространство), что защищает клетку от окружающей среды. | двумерная жидкость | все эукариоты | |
| клеточная стенка | Клеточная стенка состоит из пептидогликана и является жесткой, придает клетке форму, помогает удерживать органеллы внутри клетки и не позволяет клетке разорваться из-за изменений осмотического давления. | целлюлоза | растения, простейшие, редкие клептопластические организмы | |
| хлоропласт (пластида ) | фотосинтез, улавливает энергию солнечного света | двухмембранный отсек | растения, простейшие, редкие клептопластические организмы | имеет собственную ДНК; предполагается, что он будет поглощен предковой эукариотической клеткой (эндосимбиоз) |
| эндоплазматический ретикулум | трансляция и сворачивание новых белков (грубый эндоплазматический ретикулум), экспрессия липидов (гладкий эндоплазматический ретикулум) | одномембранный отсек | все эукариоты | грубая эндоплазматическая сеть покрыта рибосомами, имеет складки, представляющие собой плоские мешочки; гладкая эндоплазматическая сеть имеет трубчатые складки |
| жгутик | передвижение, сенсорный | белок | некоторые эукариоты | |
| аппарат Гольджи | сортировка, упаковка, переработка и модификация белков | одномембранный отсек | все эукариоты | цис-лицевой (выпуклый), ближайший к шероховатой эндоплазматической сети; транс-лицевой (вогнутый) дальше всего от грубого эндоплазматического ретикулума |
| митохондрия | производство энергии за счет окисления веществ глюкозы и высвобождения аденозинтрифосфат | двухмембранный отсек | большинство эукариот | составляющий элемент хондриом; имеет собственную ДНК; предполагается, что его поглотила предковая эукариотическая клетка (эндосимбиоз)[20] |
| ядро | Поддержание ДНК, контролирует всю деятельность клетки, РНК транскрипция | двухмембранный отсек | все эукариоты | содержит большую часть геном |
| вакуоль | хранение, транспортировка, помогает поддерживать гомеостаз | одномембранный отсек | эукариоты |
Митохондрии и пластиды, в том числе хлоропласты, имеют двойные мембраны и собственные ДНК. Согласно эндосимбиотическая теория, они, как полагают, возникли из не полностью потребленных или вторгшихся прокариотические организмы.
| Органелла / Макромолекула | Основная функция | Структура | Организмы |
|---|---|---|---|
| акросома | помогает сперматозоидам слиться с яйцеклеткой | одномембранный отсек | большинство животных |
| аутофагосома | везикула, которая изолирует цитоплазматический материал и органеллы для деградации | двухмембранный отсек | все эукариоты |
| центриоль | якорь для цитоскелет, организует деление клеток, образуя волокна веретена | Микротрубочка белок | животные |
| ресничка | движение во внешней среде или во внешней среде; «важнейший сигнальный путь развития».[21] | Микротрубочка белок | животные, простейшие, немного растений |
| книдокиста | жалящий | свернутый полый каналец | книдарийцы |
| прибор для проверки зрения | обнаруживает свет, позволяя фототаксис иметь место | зеленые водоросли и другие одноклеточные фотосинтетический организмы, такие как эвглениды | |
| гликосома | выполняет гликолиз | одномембранный отсек | Немного простейшие, Такие как Трипаносомы. |
| глиоксисома | преобразование жира в сахар | одномембранный отсек | растения |
| гидрогеносома | производство энергии и водорода | двухмембранный отсек | несколько одноклеточных эукариот |
| лизосома | распад больших молекул (например, белков + полисахаридов) | одномембранный отсек | животные |
| меланосома | хранение пигмента | одномембранный отсек | животные |
| митосома | вероятно играет роль в Железно-серный кластер (Fe-S) сборка | двухмембранный отсек | несколько одноклеточных эукариот без митохондрий |
| миофибрилла | миоцит сокращение | связанные волокна | животные |
| ядрышко | пре-рибосомная продукция | белок-ДНК-РНК | большинство эукариот |
| оцеллоид | обнаруживает свет и, возможно, формы, позволяя фототаксис иметь место | двухмембранный отсек | члены семьи Warnowiaceae |
| в скобках | не охарактеризован | не охарактеризован | грибы |
| пероксисома | расщепление метаболической перекиси водорода | одномембранный отсек | все эукариоты |
| протеасома | деградация ненужных или поврежденных белков протеолизом | очень большой белковый комплекс | все эукариоты, все археи и некоторые бактерии |
| рибосома (80S) | перевод РНК в белки | РНК-белок | все эукариоты |
| стрессовая гранула | хранение мРНК[22] | безмембранный (мРНП комплексы) | большинство эукариот |
| TIGER домен | белки, кодирующие мРНК | безмембранный | большинство организмов |
| везикул | транспортировка материалов | одномембранный отсек | все эукариоты |
Другие связанные структуры:
Прокариотические органеллы
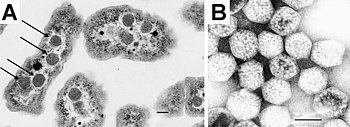

Прокариоты не так сложны по структуре, как эукариоты, и когда-то считалось, что у них нет внутренних структур, окруженных липидные мембраны. В прошлом они часто рассматривались как имеющие слабую внутреннюю организацию и отсутствие клеточные отсеки; но постепенно появляются подробности о внутренних структурах прокариот. Ранним ошибочным поворотом была идея, разработанная в 1970-х годах, что бактерии могут содержать клеточная мембрана складки, названные мезосомы, но позже было показано, что это артефакты, произведенные химическими веществами, используемыми для подготовки клеток к электронная микроскопия.[24]
Однако появляется все больше доказательств компартментализации, по крайней мере, у некоторых прокариот.[2] Недавние исследования показали, что по крайней мере у некоторых прокариот есть микрокамеры, Такие как карбоксисомы. Эти субклеточные компартменты имеют диаметр 100–200 нм и окружены белковой оболочкой.[1] Еще более поразительным является описание мембраносвязанного магнитосомы в бактериях, о которых сообщалось в 2006 году.[25][26]
Бактериальный тип Планктомицеты выявил ряд особенностей компартментализации. Планктомицеты имеют внутрицитоплазматические мембраны, которые разделяют цитоплазму на парфоплазму (внешнее пространство, свободное от рибосом) и пиреллулосому (или рибоплазму, внутреннее пространство, содержащее рибосомы).[27] Мембранный анаммоксосомы были обнаружены у пяти родов Planctomycetes anammox.[28] У планктомицетов Gemmata obscuriglobus сообщалось о подобной ядру структуре, окруженной липидными мембранами.[27][29]
Компартментализация - это особенность прокариотических фотосинтетический конструкции.[2] Фиолетовые бактерии имеют "хроматофоры", которые являются реакционными центрами в инвагинации клеточной мембраны.[2] Зеленые серные бактерии имеют хлоросомы, которые являются фотосинтетическими антенные комплексы обнаружено, что они связаны с клеточными мембранами.[2] Цианобактерии иметь внутренний тилакоид мембраны для светозависимый фотосинтез; исследования показали, что клеточная мембрана и мембраны тилакоидов не являются непрерывными друг с другом.[2]
| Органелла / макромолекула | Основная функция | Структура | Организмы |
|---|---|---|---|
| анаммоксосома | анаэробное окисление аммония | Ladderane липидная мембрана | "Candidatus "бактерии внутри Планктомицеты |
| карбоксисомный | фиксация углерода | белок-оболочка бактериальный микрокомпьютер | некоторые бактерии |
| хлоросома | фотосинтез | светособирающий комплекс, прикрепленный к клеточной мембране | зеленые серные бактерии |
| жгутик | движение во внешней среде | белковая нить | некоторые прокариоты и эукариоты |
| магнитосома | магнитная ориентация | неорганический кристалл, липидная мембрана | магнитотактические бактерии |
| нуклеоид | Поддержание ДНК, транскрипция к РНК | ДНК-белок | прокариоты |
| пилус | Адгезия к другим клеткам для конъюгации или к твердому субстрату для создания подвижных сил. | волосовидный отросток, торчащий (хотя и частично встроенный) в плазматическую мембрану | прокариотические клетки |
| плазмида | Обмен ДНК | кольцевая ДНК | некоторые бактерии |
| рибосома (70S) | перевод РНК в белки | РНК-белок | бактерии и археи |
| тилакоид мембраны | фотосинтез | белки и пигменты фотосистемы | по большей части цианобактерии |
Смотрите также
- Гипотеза CoRR
- Эжектосома
- Эндосимбиотическая теория
- Биогенез органелл
- Транспортировка мембранных везикул
- Интерфейс "хозяин-патоген"
Рекомендации
- ^ а б Kerfeld CA, Sawaya MR, Tanaka S, Nguyen CV, Phillips M, Beeby M, Yeates TO (август 2005 г.). «Белковые структуры, образующие оболочку примитивных органелл». Наука. 309 (5736): 936–8. Bibcode:2005Наука ... 309..936K. CiteSeerX 10.1.1.1026.896. Дои:10.1126 / science.1113397. PMID 16081736.
- ^ а б c d е ж Мурат, Дороти; Бирн, Меган; Комейли, Араш (01.10.2010). «Клеточная биология прокариотических органелл». Перспективы Колд-Спринг-Харбор в биологии. Дои:10.1101 / cshperspect.a000422. ЧВК 2944366. PMID 20739411. Получено 2020-07-11.
- ^ Петерсон Л. (17 апреля 2010 г.). «Освоение частей клетки». Планета уроков. Получено 2010-04-19.
- ^ Ди Грегорио MA (2005). Отсюда в вечность: Эрнст Геккель и научная вера. Гёттинген: Vandenhoeck & Ruprecht. п. 218.
- ^ Bütschli O (1888). Klassen u. Доктора Х. Г. Бронна Ordnungen des Thier-Reichs wissenschaftlich dargestellt in Wort und Bild. Эрстер Бэнд. Простейшие. Dritte Abtheilung: Infusoria und System der Radiolaria. п. 1412.
Die Vacuolen sind demnach in strengem Sinne keine beständigen Organe oder O r g a n u l a (wie Möbius die Organe der Einzelligen im Gegensatz zu denen der Vielzelligen zu nennen vorschlug).
- ^ Райдер JA, изд. (Февраль 1889 г.). «Эмбриология: строение сперматозоида человека». Американский натуралист. 23: 184.
Возможно, будет полезно использовать здесь слово «органелла» вместо «орган», следуя предложению Мебиуса. Функционально дифференцированные многоклеточные агрегаты в многоклеточных формах или многоклеточных являются в этом смысле органами, тогда как для функционально дифференцированных частей одноклеточные организмы или для таких дифференцированных частей одноклеточных зародышевых элементов многоклеточных животных подходит крохотный органула.
- ^ Робин C, Пуше G, Дюваль MM, Retterrer E, Tourneux F (1891). Journal de l'anatomie et de la Physiologie normales et patologiques de l'homme et des animaux. Ф. Алкан.
- ^ а б Мёбиус К (Сентябрь 1884 г.). "Das Sterben der einzelligen und der vielzelligen Tiere. Vergleichend betrachtet". Biologisches Centralblatt. 4 (13, 14): 389–392, 448.
Während die Fortpflanzungszellen der vielzelligen Tiere Unthätig fortleben bis sie sich loslösen, wandern und entwickeln, treten die einzelligen Tiere auch durch die an der Fortpflanzung beteiligten Foot Leibesmasse in Au der Mit der Veßencher. "Die Organe der Heteroplastiden bestehen aus vereinigten Zellen. Da die Organe der Monoplastiden nur verschieden ausgebildete Teile e i n e r Zelle sind schlage ich vor, sie «Organula» zu nennen
- ^ Уокер, Патрик (2009). Ядерный импорт гистонового складчатого мотива, содержащего гетеродимеры, импортином 13. Niedersächsische Staats-und Universitätsbibliothek Göttingen.
- ^ Килинг П.Дж., Арчибальд Дж.М. (апрель 2008 г.). «Эволюция органелл: что в названии?». Текущая биология. 18 (8): R345-7. Дои:10.1016 / j.cub.2008.02.065. PMID 18430636.
- ^ Иманян Б., Карпентер К.Дж., Килинг П.Дж. (март – апрель 2007 г.). «Митохондриальный геном третичного эндосимбионта сохраняет гены белков транспорта электронов». Журнал эукариотической микробиологии. 54 (2): 146–53. Дои:10.1111 / j.1550-7408.2007.00245.x. PMID 17403155.
- ^ Маллинз С (2004). «Теория биогенеза органелл: историческая перспектива». Биогенез клеточных органелл. Springer Science + Business Media, Национальные институты здоровья. ISBN 978-0-306-47990-8.
- ^ Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П. «Генетические системы митохондрий и пластид». Молекулярная биология клетки (4-е изд.). ISBN 978-0-8153-3218-3.
- ^ Кэмпбелл Н.А., Рис Дж. Б., Митчелл Л.Г. (2002). Биология (6-е изд.). Бенджамин Каммингс. ISBN 978-0-8053-6624-2.
- ^ Нотт Т.Дж., Петсалаки Э., Фарбер П., Джервис Д., Фасснер Э., Плоховец А., Краггс Т.Д., Базетт-Джонс Д.П., Поусон Т., Форман-Кей Д.Д., Болдуин А.Дж. (март 2015 г.). «Фазовый переход неупорядоченного белка nuage генерирует экологически чувствительные безмембранные органеллы». Молекулярная клетка. 57 (5): 936–947. Дои:10.1016 / j.molcel.2015.01.013. ЧВК 4352761. PMID 25747659.
- ^ Банани С.Ф., Ли ХО, Хайман А.А., Розен М.К. (май 2017 г.). «Биомолекулярные конденсаты: организаторы клеточной биохимии». Обзоры природы Молекулярная клеточная биология. 18 (5): 285–298. Дои:10.1038 / nrm.2017.7. ЧВК 7434221. PMID 28225081.
- ^ Кормак Д.Х. (1984). Введение в гистологию. Липпинкотт. ISBN 978-0-397-52114-2.
- ^ Brangwynne CP, Eckmann CR, Courson DS, Rybarska A, Hoege C, Gharakhani J, Jülicher F, Hyman AA (июнь 2009 г.). «Гранулы зародышевой линии P представляют собой жидкие капли, которые локализуются за счет контролируемого растворения / конденсации». Наука. 324 (5935): 1729–32. Bibcode:2009Sci ... 324.1729B. Дои:10.1126 / science.1172046. PMID 19460965.
- ^ Фэи Р.К., Ньютон Г.Л., Аррик Б., Оверданк-Богарт Т., Алей С.Б. (апрель 1984 г.). «Entamoeba histolytica: эукариот без метаболизма глутатиона». Наука. 224 (4644): 70–2. Bibcode:1984 Наука ... 224 ... 70F. Дои:10.1126 / science.6322306. PMID 6322306.
- ^ Альбертс Б., Джонсон А., Льюис Дж., Морган Д., Рафф М.С., Робертс К., Уолтер П., Уилсон Дж. Х., Хант Т. (18 ноября 2014 г.). Молекулярная биология клетки (Шестое изд.). Наука о гирляндах. п. 679. ISBN 978-0815345244.
- ^ Бадано Дж. Л., Мицума Н., Билс П. Л., Катсанис Н. (сентябрь 2006 г.). «Цилиопатии: новый класс генетических заболеваний человека». Ежегодный обзор геномики и генетики человека. 7: 125–48. Дои:10.1146 / annurev.genom.7.080505.115610. PMID 16722803.
- ^ Андерсон П., Кедерша Н. (март 2008 г.). «Стресс-гранулы: Дао сортировки РНК». Тенденции в биохимических науках. 33 (3): 141–50. Дои:10.1016 / j.tibs.2007.12.003. PMID 18291657.
- ^ Цай Ю., Савая М.Р., Cannon GC, Цай Ф., Уильямс Э.Б., Хайнхорст С., Керфельд, Калифорния, Йейтс, ТО (июнь 2007 г.). «Структурный анализ CsoS1A и белковой оболочки карбоксисомы Halothiobacillus neapolitanus». PLoS Биология. 5 (6): e144. Дои:10.1371 / journal.pbio.0050144. ЧВК 1872035. PMID 17518518.
- ^ Райтер А. (январь – февраль 1988 г.). «Вклад новых криометодов в лучшее знание анатомии бактерий». Annales de l'Institut Pasteur. Микробиология. 139 (1): 33–44. Дои:10.1016/0769-2609(88)90095-6. PMID 3289587.
- ^ Комейли А., Ли З., Ньюман Д. К., Дженсен Дж. Дж. (Январь 2006 г.). «Магнитосомы - это инвагинации клеточной мембраны, организованные актин-подобным белком MamK» (PDF). Наука. 311 (5758): 242–5. Bibcode:2006Научный ... 311..242K. Дои:10.1126 / science.1123231. PMID 16373532.
- ^ Scheffel A, Gruska M, Faivre D, Linaroudis A, Plitzko JM, Schüler D (март 2006 г.). «Кислый белок выравнивает магнитосомы вдоль нитчатой структуры магнитотактических бактерий». Природа. 440 (7080): 110–4. Bibcode:2006Натура.440..110С. Дои:10.1038 / природа04382. PMID 16299495.
- ^ а б Линдси, М. Р .; Webb, R.I .; Строус, М; Jetten, M. S .; Батлер, М. К .; Forde, R.J .; Фуэрст, Дж. А. (2001). «Компартментализация клеток в планктомицетах: новые типы структурной организации бактериальной клетки». Архив микробиологии. 175 (6): 413–29. Дои:10.1007 / s002030100280. PMID 11491082.
- ^ Jetten, Mike S.M .; Нифтрик, Лаура ван; Строус, Марк; Картал, Боран; Кельтьенс, Ян Т .; Оп ден Камп, Хуб Дж. М. (2009-06-01). «Биохимия и молекулярная биология анаммокс-бактерий». Критические обзоры в биохимии и молекулярной биологии. С. 65–84. Дои:10.1080/10409230902722783. PMID 19247843. Получено 2020-08-03.
- ^ Fuerst JA (13 октября 2005 г.). «Внутриклеточная компартментация у планктомицетов». Ежегодный обзор микробиологии. 59: 299–328. Дои:10.1146 / annurev.micro.59.030804.121258. PMID 15910279.
внешняя ссылка
| Библиотечные ресурсы о Органелла |
 СМИ, связанные с Органеллы в Wikimedia Commons
СМИ, связанные с Органеллы в Wikimedia Commons- Проект "Древо жизни": эукариоты
- Базы данных органелл
| Авторитетный контроль |
|---|