Хлоропласт - Chloroplast


Хлоропласты /ˈkлɔːrəˌплæsтs,-плɑːsтs/[1][2] находятся органеллы это поведение фотосинтез, где фотосинтетический пигмент хлорофилл фиксирует энергия из Солнечный свет, преобразует ее и хранит в молекулах-накопителях энергии АТФ и НАДФН освобождая кислород из воды в растение и водоросль клетки. Затем они используют АТФ и НАДФН для создания органических молекул из углекислый газ в процессе, известном как Цикл Кальвина. Хлоропласты выполняют ряд других функций, в том числе: синтез жирных кислот, много аминокислота синтез, и иммунная реакция в растениях. Количество хлоропластов на клетку варьируется от одного у одноклеточных водорослей до 100 у растений, таких как Арабидопсис и пшеница.
Хлоропласт - это органеллы, известные как пластида, был характеризован его две мембраны и высокая концентрация хлорофилл. Другие типы пластид, такие как лейкопласт и хромопласт, содержат мало хлорофилла и не осуществляют фотосинтез.
Хлоропласты очень динамичны - они циркулируют и перемещаются внутри растительных клеток, а иногда и ущипнуть пополам воспроизвести. На их поведение сильно влияют такие факторы окружающей среды, как цвет и интенсивность света. Хлоропласты, подобные митохондрии, содержат свои собственные ДНК, который считается унаследованным от их предка - фотосинтетического цианобактерии что было поглотил ранним эукариотический клетка.[3] Хлоропласты не могут быть произведены растительной клеткой и должны быть унаследованы каждой дочерней клеткой во время деления клетки.
За одним исключением ( амебовидный Паулинелла хроматофора), все хлоропласты, вероятно, можно проследить до одного эндосимбиотическое событие, когда цианобактерия была поглощена эукариотом. Несмотря на это, хлоропласты можно найти у очень широкого круга организмов, некоторые из которых даже не связаны напрямую друг с другом - следствие многих вторичный и даже третичные эндосимбиотические события.
Слово хлоропласт происходит от греческих слов хлор (χλωρός), что означает зеленый, и пластыри (πλάστης), что означает «тот, кто формирует».[4]
Открытие
Первое окончательное описание хлоропласта (Хлорофиллкёрнен, «зерно хлорофилла») было дано Хьюго фон Моль в 1837 г. в виде отдельных тел внутри клетки зеленого растения.[5] В 1883 г. Андреас Франц Вильгельм Шимпер назвал бы эти тела «хлоропластидами» (Хлоропластиден).[6] В 1884 г. Эдуард Страсбургер приняли термин «хлоропласты» (Хлоропластен).[7][8][9]
Происхождение и эволюция
Хлоропласты - это один из многих типов органелл в растительной клетке. Считается, что они произошли от эндосимбиотический цианобактерии. Митохондрии предположительно возникли в результате аналогичного случая эндосимбиоза, когда аэробный прокариот был охвачен.[10] Такое происхождение хлоропластов было впервые предположено русским биологом. Константин Мерещковский в 1905 г.[11] после Андреас Франц Вильгельм Шимпер заметил в 1883 г., что хлоропласты очень похожи на цианобактерии.[6] Хлоропласты встречаются только в растения, водоросли,[12] и амебовидный Паулинелла хроматофора.
Родительская группа: Цианобактерии
Хлоропласты считаются эндосимбиотическими цианобактериями.[13] Цианобактерии иногда называют сине-зелеными водоросли хотя они прокариоты. Они разнообразны филюм из бактерии способный выполнять фотосинтез, и являются грамотрицательный, что означает, что они имеют две клеточные мембраны. Цианобактерии также содержат клеточная стенка пептидогликана, который толще, чем у других грамотрицательных бактерий, и расположен между их клеточными мембранами.[14] Как и хлоропласты, они имеют тилакоиды в.[15] На тилакоидных мембранах имеются фотосинтетические пигменты, включая хлорофилл а.[16] Фикобилины также являются распространенными пигментами цианобактерий, обычно организованными в полусферические фикобилисомы прикреплены к внешней стороне мембран тилакоидов (однако фикобилины не являются общими для всех хлоропластов).[16][17]
Первичный эндосимбиоз
![Первичный эндосимбиоз. Эукариот с митохондриями поглотил цианобактерии в случае серийного первичного эндосимбиоза, создав линию клеток с обеими органеллами. [10] Важно отметить, что у цианобактериального эндосимбионта уже была двойная мембрана - мембрана, образованная фагосомной вакуолью, была утрачена. [18]](http://upload.wikimedia.org/wikipedia/commons/thumb/b/b8/Chloroplast_endosymbiosis_simple.svg/440px-Chloroplast_endosymbiosis_simple.svg.png)
Эукариот с митохондрии поглотил цианобактерии в случае серийный первичный эндосимбиоз, создающий родословная клеток с обеими органеллами.[10] Важно отметить, что цианобактериальный эндосимбионт уже имел двойную мембрану - фагосомная вакуоль -производная мембрана была потеряна.[18]
Где-то 1-2 миллиарда лет назад[19][20][21]свободная жизнь цианобактерии вошел рано эукариотический клетка, либо в качестве пищи, либо в качестве внутреннего паразит,[10] но удалось избежать фагоцитарная вакуоль он содержался в.[16] Два самых сокровенных липидно-двухслойные мембраны[22] окружающие все хлоропласты, соответствуют внешнему и внутреннему мембраны предковых цианобактерий грамм отрицательный клеточная стенка[18][23][24] а не фагосомный мембрана от хозяина, которая, вероятно, была потеряна.[18]Новый клеточный резидент быстро стал преимуществом, предоставив пищу эукариотическому хозяину, что позволило ему жить внутри него.[10] Со временем цианобактерия ассимилировалась, и многие ее гены были потеряны или переданы ядро хозяина.[25] Из геномов, которые, вероятно, первоначально содержали более 3000 генов, в хлоропластах современных растений осталось только около 130 генов.[20] Затем некоторые из его белков были синтезированы в цитоплазме клетки-хозяина и импортированы обратно в хлоропласт (ранее цианобактерии).[25][26] По отдельности, где-то около 90–140 миллионов лет назад это случилось снова и привело к амебовидный Паулинелла хроматофора.[21]
Это событие называется эндосимбиоз, или «клетка, живущая внутри другой клетки с взаимной выгодой для обоих». Внешнюю ячейку обычно называют хозяин а внутренняя ячейка называется эндосимбионт.[10]
Считается, что хлоропласты возникли после митохондрии, поскольку все эукариоты содержат митохондрии, но не все имеют хлоропласты.[10][27] Это называется серийный эндосимбиоз - ранний эукариот, поглощающий митохондрия предка, и некоторые его потомки затем поглотили предка хлоропластов, создав клетку с хлоропластами и митохондриями.[10]
Вопрос о том, произошли ли первичные хлоропласты в результате единственного эндосимбиотического события или многих независимых поглощений в различных эукариотических клонах, уже давно обсуждается. В настоящее время принято считать, что организмы с первичными хлоропластами разделяют единственный предок это заняло цианобактерии 600–2000 миллионов лет назад.[19][21] Было высказано предположение, что ближайшим живым родственником этой бактерии является Gloeomargarita lithophora.[28][29][30] Исключение составляет амебовидный Паулинелла хроматофора, который происходит от предка, который принял Прохлорококк цианобактерии 90–500 миллионов лет назад.[31][30][21]
Эти хлоропласты, восходящие непосредственно к предкам цианобактерий, известны как начальный пластиды[32] ("пластида " в данном контексте означает почти то же, что и хлоропласт.[10]). Все первичные хлоропласты принадлежат к одной из четырех ветвей хлоропластов - глаукофит линия хлоропластов, амебовидный Паулинелла хроматофора происхождение, родофит (красные водоросли) линия хлоропластов, или хлоропластидан (зеленый) линия хлоропластов.[33] В родофит и хлоропластидан родословные самые крупные,[18] с хлоропластидан (зеленый) - тот, который содержит наземные растения.[18]
Глаукофита
Обычно считается, что эндосимбиоз произошел в Archaeplastida, внутри которого глаукофита являясь возможной самой ранней расходящейся линией.[26][13] Группа хлоропластов глаукофита является самой маленькой из трех первичных ветвей хлоропластов, обнаруживаемая только у 13 видов,[18] и считается одним из первых.[18][19][34] Глаукофиты имеют хлоропласты, которые сохраняют пептидогликан стенка между их двойными мембранами,[32] как их цианобактерий родитель.[14] По этой причине хлоропласты глаукофита также известны как «муропласты».[32] (кроме «цианопластов» или «цианелл»). Хлоропласты глаукофита также содержат концентрический разложенный тилакоиды, которые окружают карбоксисомный - ан икосаэдр структура, в которой хлоропласты глаукофитов и цианобактерии сохраняют свои фиксация углерода фермент RuBisCO in. Крахмал, который они синтезируют, собирается вне хлоропласта.[16] Как и цианобактерии, тилакоиды хлоропластов глаукофитов и родофитов усеяны светособирающими структурами, называемыми фикобилисомы.[16][32] По этим причинам хлоропласты глаукофита считаются примитивным промежуточным звеном между цианобактериями и более развитыми хлоропластами в красные водоросли и растения.[32]




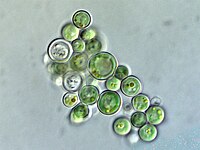
Rhodophyceae (красные водоросли)
В родофит, или же красные водоросли Группа хлоропластов - еще одна большая и разнообразная ветвь хлоропластов.[18] Хлоропласты родофитов также называют родопласты,[32] буквально «красные хлоропласты».[36]
Родопласты имеют двойную мембрану с межмембранным пространством и фикобилин пигменты организованы в фикобилисомы на тилакоидных мембранах, предотвращая накопление их тилакоидов.[16] Некоторые содержат пиреноиды.[32] Родопласты имеют хлорофилл а и фикобилины[34] для фотосинтетических пигментов; фикобилин фикоэритрин отвечает за то, что многие красные водоросли приобретают характерный красный цвет.[35] Однако, поскольку они также содержат сине-зеленый хлорофилл а и другие пигменты, многие из них от красноватого до пурпурного цвета.[32] Красный пигмент фикоэритерин - это адаптация, помогающая красным водорослям улавливать больше солнечного света в глубокой воде.[32]- поэтому некоторые красные водоросли, обитающие на мелководье, имеют меньше фикоэритрина в родопластах и могут казаться более зеленоватыми.[35] Родопласты синтезируют крахмал, называемый флоридский крахмал,[32] который собирается в гранулы вне родопласта, в цитоплазме красной водоросли.[16]
Хлоропластида (зеленые водоросли и растения)
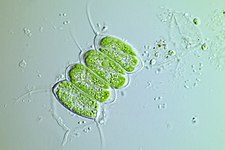
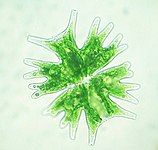



В хлоропластида хлоропласты, или зеленый хлоропласты - еще одна крупная, очень разнообразная линия первичных хлоропластов. Их организмы-хозяева широко известны как зеленые водоросли и наземные растения.[37] Они отличаются от хлоропластов глаукофитов и красных водорослей тем, что потеряли фикобилисомы, и содержать хлорофилл б вместо.[16] Большинство зеленых хлоропластов (очевидно) зеленый, хотя некоторые нет, например, некоторые формы Hæmatococcus pluvialis из-за дополнительных пигментов, которые перекрывают зеленый цвет хлорофиллов. Хлоропласты Chloroplastida потеряли пептидогликан стенка между их двойной мембраной, оставляя межмембранное пространство.[16] Немного растения кажется, сохранил гены для синтеза пептидогликановый слой, хотя они были перепрофилированы для использования в деление хлоропласта вместо.[38]
Большинство хлоропластов, изображенных в этой статье, являются зелеными хлоропластами.
Зеленые водоросли и растения сохраняют свой крахмал внутри их хлоропласты,[16][34][37] а у растений и некоторых водорослей тилакоиды хлоропластов расположены в стеках гран. Некоторые хлоропласты зеленых водорослей содержат структуру, называемую пиреноид,[16] который функционально похож на глаукофит карбоксисомный в том, что это где RuBisCO и CO2 концентрируются в хлоропласте.[39]

Helicosporidium это род нефотосинтетических паразитических зеленых водорослей, которые, как полагают, содержат рудиментарный хлоропласт.[34] Гены из хлоропласта[40] и ядерные гены, указывающие на присутствие хлоропласта, были обнаружены в Helicosporidium[34] даже если никто не видел самого хлоропласта.[34]
Паулинелла хроматофора
Хотя большинство хлоропластов происходит из первого набора эндосимбиотических событий, Паулинелла хроматофора является исключением, которое недавно приобрело фотосинтетический цианобактериальный эндосимбионт. Неясно, имеет ли этот симбионт близкое родство с предковым хлоропластом других эукариот.[18] Находясь на ранних стадиях эндосимбиоза, Паулинелла хроматофора может дать некоторое представление о том, как эволюционировали хлоропласты.[25][41] Паулинелла клетки содержат одну или две сине-зеленые фотосинтезирующие структуры в форме колбасы, называемые хроматофорами,[25][41] произошел от цианобактерии Синехококк. Хроматофоры не могут выжить вне своего хозяина.[25] Хроматофорная ДНК составляет около миллиона пар оснований длинный, содержащий около 850 кодирующих белков гены - намного меньше трех миллионов пар оснований Синехококк геном[25] но намного больше, чем геном из примерно 150 000 пар оснований более ассимилированного хлоропласта.[42][43][44] Хроматофоры передали ядру своего хозяина гораздо меньше своей ДНК. Около 0,3–0,8% ядерной ДНК в Паулинелла из хроматофоров, по сравнению с 11–14% из хлоропластов растений.[41]
Вторичный и третичный эндосимбиоз
Многие другие организмы получили хлоропласты от первичных клонов хлоропластов через вторичный эндосимбиоз - поглотив красную или зеленую водоросль, содержащую хлоропласт. Эти хлоропласты известны как вторичные пластиды.[32]
В то время как первичные хлоропласты имеют двойную мембрану от их цианобактериальный предком, вторичные хлоропласты имеют дополнительные мембраны за пределами двух исходных, в результате вторичного эндосимбиотического события, когда нефотосинтетический эукариот поглотил водоросль, содержащую хлоропласт, но не смог ее переварить, как цианобактерия в начале этой истории.[18] Поглотившаяся водоросль разрушилась, оставив только свой хлоропласт, а иногда и свой клеточная мембрана и ядро, образующий хлоропласт с тремя или четырьмя мембранами[45]- две цианобактериальные мембраны, иногда клеточная мембрана съеденной водоросли и фагосомная вакуоль от клеточной мембраны хозяина.[18]


Гены в ядре фагоцитированного эукариота часто переносятся в ядро вторичного хозяина.[18]Криптомонады и хлорарахниофиты сохраняют ядро фагоцитированного эукариота, объект, называемый нуклеоморф,[18] расположен между второй и третьей мембранами хлоропласта.[16][26]
Все вторичные хлоропласты происходят из зеленый и красные водоросли - нет вторичных хлоропластов из глаукофиты наблюдались, вероятно потому, что глаукофиты относительно редки в природе, что снижает вероятность того, что они были поглощены другим эукариотом.[18]
Хлоропласты, полученные из зеленых водорослей
Зеленые водоросли были подняты эвглениды, хлорарахниофиты, родословная динофлагелляты,[34] и, возможно, предок линии CASH (криптомонады, альвеолиты, страменопилы и гаптофиты )[46] в трех или четырех отдельных захватах.[47] Многие хлоропласты, полученные из зеленых водорослей, содержат пиреноиды, но в отличие от хлоропластов своих зеленых водорослей-предков, запасы продуктов накапливаются в гранулах за пределами хлоропластов.[16]
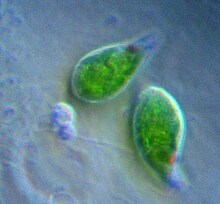
Эвгленофиты
Эвгленофиты - это группа обычных бичеванный протисты которые содержат хлоропласты, полученные из зеленой водоросли.[18] Эвгленофит хлоропласты имеют три мембраны - считается, что мембрана первичного эндосимбионта была потеряна, оставив цианобактериальные мембраны и фагосомную мембрану вторичного хозяина.[18] Хлоропласты эвгленофитов имеют пиреноид и тилакоиды сложены группами по три штуки. Фотосинтетический продукт хранится в виде парамилон, который содержится в мембраносвязанных гранулах в цитоплазме эвгленофита.[16][34]

Хлорарахниофиты
Хлорарахниофиты /ˌkлɔːrəˈрækпяoʊˌжаɪтs/ представляют собой редкую группу организмов, которые также содержат хлоропласты, полученные из зеленых водорослей,[18] хотя их история более сложна, чем у эвгленофитов. Считается, что предком хлорарахниофитов был эукариот с красный хлоропласт, полученный из водорослей. Затем считается, что он потерял свой первый хлоропласт красных водорослей, а позже поглотил зеленую водоросль, дав ей второй, полученный из зеленых водорослей хлоропласт.[34]
Хлоропласты хлорахниофитов ограничены четырьмя мембранами, за исключением клеточной мембраны, где мембраны хлоропластов сливаются в двойную мембрану.[16] Их тилакоиды расположены в рыхлых стопках по три штуки.[16] Хлорарахниофиты имеют форму полисахарида, называемую хризоламинарин которые они хранят в цитоплазме,[34] часто собираются вокруг хлоропласта пиреноид, который выпячивается в цитоплазму.[16]
Хлоропласты Chlorarachniophyte примечательны тем, что зеленая водоросль, из которой они происходят, не была полностью разрушена - ее ядро все еще сохраняется в виде нуклеоморф[18] находится между второй и третьей мембранами хлоропласта[16]- перипластидное пространство, что соответствует цитоплазме зеленой водоросли.[34]
Динофитный хлоропласт, производный от празинофитов
Лепидодиниум вирид и его близкими родственниками являются динофиты (см. ниже), которые утратили свой первоначальный хлоропласт перидинина и заменили его хлоропластом, полученным из зеленых водорослей (более конкретно, празинофит ).[16][48] Лепидодиниум - единственный динофит, у которого хлоропласт не из родопласт происхождение. Хлоропласт окружен двумя мембранами и не имеет нуклеоморфа - все гены нуклеоморфа переданы динофиту. ядро.[48] Эндосимбиотическое событие, которое привело к этому хлоропласту, было серийный вторичный эндосимбиоз а не третичный эндосимбиоз - эндосимбионт был зеленая водоросль содержащий первичный хлоропласт (образующий вторичный хлоропласт).[34]
Хлоропласты, полученные из красных водорослей
Криптофиты
Криптофиты, или криптомонады - это группа водорослей, которые содержат хлоропласты, полученные из красных водорослей. Хлоропласты криптофитов содержат нуклеоморф, внешне напоминающий нуклеоморф хлорарахниофиты.[18] Хлоропласты криптофитов имеют четыре мембраны, самая внешняя из которых непрерывна шероховатой эндоплазматической сети. Они синтезируют обычные крахмал, который хранится в гранулах, находящихся в перипластидное пространство - вне исходной двойной мембраны, в месте, соответствующем цитоплазме красной водоросли. Внутри хлоропластов криптофита находится пиреноид и тилакоиды в стопках по две штуки.[16]
В их хлоропластах нет фикобилисомы,[16] но у них есть фикобилиновые пигменты которые они держат в своем тилакоидном пространстве, а не закрепляются снаружи тилакоидных мембран.[16][18]
Криптофиты могли сыграть ключевую роль в распространении хлоропластов на основе красных водорослей.[49][50]

Гаптофиты
Гаптофиты похожи и тесно связаны с криптофитами или гетероконтофитами.[34] В их хлоропластах отсутствует нуклеоморф,[16][18] их тилакоиды расположены в стопках по три, и они синтезируют хризоламинарин сахар, который они хранят полностью вне хлоропласта, в цитоплазме гаптофита.[16]
Гетероконтофиты (страменопилы)

В гетероконтофиты, также известные как страменопилы, представляют собой очень большую и разнообразную группу эукариот. Фотоавтотрофная линия, Охрофита, в том числе диатомеи и бурые водоросли, золотые водоросли,[35] и желто-зеленые водоросли, также содержит хлоропласты, полученные из красных водорослей.[34]
Хлоропласты гетероконта очень похожи на хлоропласты гаптофитов, содержащие пиреноид, триплетные тилакоиды и, за некоторыми исключениями,[16] имеющий четырехслойную пластидную оболочку, самая внешняя эпипластидная мембрана, соединенная с эндоплазматический ретикулум. Как и гаптофиты, гетероконтофиты хранят сахар в хризоламинарин гранулы в цитоплазме.[16] Хлоропласты гетероконтофитов содержат хлорофилл а и за некоторыми исключениями[16] хлорофилл c,[18] но также есть каротиноиды которые придают им множество цветов.[35]
Apicomplexans, хромериды и динофиты
Альвеолаты являются основной кладой одноклеточных эукариот как автотрофных, так и гетеротрофных членов. Наиболее заметной общей характеристикой является наличие корковых (внешних) альвеол (мешочков). Это сплюснутые пузырьки (мешочки), упакованные в непрерывный слой непосредственно под мембраной и поддерживающие ее, как правило, образующие гибкую пленку (тонкую пленку). У динофлагеллят они часто образуют броневые пластины. Многие члены содержат пластиду, полученную из красных водорослей. Одной из примечательных характеристик этой разнообразной группы является частая потеря фотосинтеза. Однако большинство этих гетеротрофов продолжают обрабатывать нефотосинтетические пластиды.[51]
- Apicomplexans
Apicomplexans представляют собой группу альвеолят. Словно геликоспроидия, они паразитируют и имеют нефотосинтетический хлоропласт.[34] Когда-то считалось, что они связаны с геликоспроидами, но теперь известно, что геликоспроиды - это зеленые водоросли, а не часть линии CASH.[34] К апикомплексам относятся: Плазмодий, то малярия паразит. Многие apicomplexans сохраняют рудиментарный хлоропласт, полученный из красных водорослей[52][34] называется апикопласт, которые они унаследовали от своих предков. Другие апикомплексаны, такие как Криптоспоридиум полностью потеряли хлоропласт.[52] Apicomplexans хранят свою энергию в амилопектин гранулы, которые расположены в их цитоплазме, даже если они нефотосинтетические.[16]
Апикопласты утратили фотосинтетические функции и не содержат фотосинтетических пигментов или настоящих тилакоидов. Они ограничены четырьмя мембранами, но мембраны не связаны с эндоплазматический ретикулум.[16] Тот факт, что apicomplexans по-прежнему сохраняют свой нефотосинтетический хлоропласт, демонстрирует, как хлоропласт выполняет важные функции, кроме фотосинтез. Растение хлоропласты обеспечивают клетки растений многими важными вещами, помимо сахара, и апикопласты ничем не отличаются - они синтезируют жирные кислоты, изопентенилпирофосфат, железо-серные кластеры, и выполнить часть гем путь.[52] Это делает апикопласт привлекательной мишенью для лекарств для лечения заболеваний, связанных с апикомплексом.[32] Важнейшей функцией апикопласта является изопентенилпирофосфат синтез - фактически, apicomplexans умирают, когда что-то мешает этой функции апикопласта, и когда apicomplexans выращивают в среде, богатой изопентенилпирофосфатом, они сбрасывают органеллы.[52]
- Хромериды
В Chromerida недавно обнаруженная группа водорослей из австралийских кораллов, которая включает некоторых близких фотосинтезирующих родственников апикомплексанс. Первый член, Chromera velia, был обнаружен и впервые выделен в 2001 году. Chromera velia со структурой, аналогичной апикомплексам, представляет собой важное звено в эволюционной истории апикомплексов и динофитов. Их пластиды имеют четыре мембраны, лишены хлорофилла с и используют форму типа II. RuBisCO полученный из события горизонтальной передачи.[53]
- Динофиты
В динофлагелляты еще одна очень большая и разнообразная группа протисты, около половины из которых (не менее частично ) фотосинтетический.[35][48]
Большинство хлоропластов динофитов вторичны. красные водоросли производные хлоропластов. Многие другие динофиты утратили хлоропласт (став нефотосинтетическим видом динофлагеллятов) или заменили его. высшее эндосимбиоз[54]- захват другой эукариотической водоросли, содержащей хлоропласт, полученный из красных водорослей. Другие заменили исходный хлоропласт на зеленые водоросли производный.[18][34][48]
Большинство хлоропластов динофитов содержат форму II RuBisCO, по крайней мере, фотосинтетические пигменты хлорофилл а, хлорофилл c2, бета-каротин, и по крайней мере один уникальный динофит ксантофилл (перидинин, диноксантин, или же диадиноксантин ), давая многим золотисто-коричневый цвет.[51][48] Все динофиты хранят крахмал в своей цитоплазме, и у большинства из них хлоропласты с тилакоидами расположены в стопки по три.[16]

Самый распространенный хлоропласт динофитов - это перидинин -типа хлоропласта, характеризующегося каротиноид пигмент перидинин в их хлоропластах вместе с хлорофилл а и хлорофилл c2.[18][48] Перидинин не обнаружен ни в одной другой группе хлоропластов.[48] Хлоропласт перидинина ограничен тремя мембранами (иногда двумя),[16] потеряв исходную клеточную мембрану эндосимбионта красных водорослей.[18][34] Самая внешняя мембрана не связана с эндоплазматическим ретикулумом.[16][48] Они содержат пиреноид, и имеют тилакоиды с триплетным стэком. Крахмал находится вне хлоропласта.[16] Важной особенностью этих хлоропластов является то, что их хлоропластная ДНК очень уменьшенный и разбиты на множество маленьких кружков. Большая часть генома переместилась в ядро, и только важные гены, связанные с фотосинтезом, остались в хлоропласте.[48]
Хлоропласт перидинина считается «оригинальным» хлоропластом динофитов,[48] который был утерян, сокращен, заменен или имеет компанию в нескольких других линиях динофитов.[34]
Фукоксантин-содержащие (происходящие от гаптофита) хлоропласты динофитов

В фукоксантин линии динофитов (включая Карлодиниум и Карения )[34] потеряли свой первоначальный хлоропласт, полученный из красных водорослей, и заменили его новым хлоропластом, полученным из гаптофит эндосимбионт. Карлодиниум и Карения вероятно занялись разными гетероконтофитами.[34] Поскольку хлоропласт гаптофита имеет четыре мембраны, ожидается, что третичный эндосимбиоз создаст шестимембранный хлоропласт, добавив гаптофитный хлоропласт. клеточная мембрана и динофит фагосомная вакуоль.[56] Однако гаптофит был сильно редуцирован, лишился нескольких мембран и ядра, остался только хлоропласт (с его исходной двойной мембраной) и, возможно, одна или две дополнительные мембраны вокруг него.[34][56]
Хлоропласты, содержащие фукоксантин, характеризуются наличием пигмента фукоксантин (фактически 19'-гексаноилокси-фукоксантин и / или 19'-бутаноилокси-фукоксантин ) и без перидинина. Фукоксантин также содержится в хлоропластах гаптофитов, что свидетельствует о его происхождении.[48]

Хлоропласты диатомовых водорослей
Некоторые динофиты, например Криптоперидиний и Дуринская[34] есть диатомовые водоросли (гетероконтофит ) производное хлоропласта.[18] Эти хлоропласты ограничены до пять мембраны,[18] (в зависимости от того, считается ли весь эндосимбионт диатомовых водорослей хлоропластом или только хлоропласт, полученный из красных водорослей). Эндосимбионт диатомовых водорослей был уменьшен относительно немного - он все еще сохраняет свой первоначальный вид митохондрии,[34] и имеет эндоплазматический ретикулум, рибосомы, а ядро и, конечно же, хлоропласты, полученные из красных водорослей - практически полный клетка,[57] все внутри хозяина просвет эндоплазматического ретикулума.[34] Однако эндосимбионт диатомовых водорослей не может хранить свою собственную пищу - вместо этого его запасающий полисахарид находится в гранулах в цитоплазме динофита-хозяина.[16][57] Ядро эндосимбионта диатомовых присутствует, но его, вероятно, нельзя назвать нуклеоморф потому что он не показывает никаких признаков сокращение генома, и, возможно, даже был расширенный.[34] Диатомовые водоросли были поглощены динофлагеллятами не менее трех раз.[34]
Эндосимбионт диатомей ограничен единственной мембраной,[48] внутри него находятся хлоропласты с четырьмя мембранами. Как и диатомовый предок диатомового эндосимбионта, хлоропласты имеют триплетные тилакоиды и пиреноиды.[57]
В некоторых из них роды, хлоропласты диатомового эндосимбионта - не единственные хлоропласты динофита. Первоначальный трехкомпонентный хлоропласт перидинина все еще существует, преобразованный в глазное пятно.[18][34]
Клептопластиды
В некоторых группах миксотрофный протисты, как некоторые динофлагелляты (например. Динофиз ) хлоропласты отделяются от пойманной водоросли и используются временно. Эти клепто-хлоропласты могут иметь срок службы всего несколько дней, а затем заменяются.[58][59]
Динофитный хлоропласт, полученный из криптофитов
Члены рода Динофиз есть фикобилин -содержащий[56] хлоропласт, взятый из криптофит.[18] Однако криптофит не является эндосимбионтом - кажется, что был взят только хлоропласт, а хлоропласт лишен своего нуклеоморф и две крайние мембраны, оставляя только двухмембранный хлоропласт. Хлоропластам криптофитов требуется их нуклеоморф, чтобы поддерживать себя, и Динофиз виды, выращенные в культура клеток в одиночку не может выжить, поэтому возможно (но не подтверждено), что Динофиз хлоропласт - это клептопласт -если так, Динофиз хлоропласты изнашиваются и Динофиз виды должны постоянно поглощать криптофиты, чтобы получить новые хлоропласты взамен старых.[48]
Хлоропластная ДНК
У хлоропластов свои ДНК,[60] часто сокращенно втДНК,[61] или же хпДНК.[62] Он также известен как пластом. Его существование было впервые доказано в 1962 году.[42] и впервые секвенировали в 1986 году, когда две японские исследовательские группы секвенировали ДНК хлоропластов печеночник и табак.[63] С тех пор сотни ДНК хлоропластов разных видов были последовательный, но в основном они принадлежат наземные растения и зеленые водоросли —глаукофиты, красные водоросли, а другие группы водорослей крайне недопредставлены, потенциально представляя некоторые предвзятость с точки зрения «типичной» структуры и содержания ДНК хлоропластов.[64]
Молекулярная структура

За некоторыми исключениями, в большинстве хлоропластов весь геном хлоропластов объединен в одну большую кольцевую молекулу ДНК,[64] обычно 120 000–170 000 пар оснований длинный.[42][43][44][20] Они могут иметь длину контура около 30–60 микрометров и массу около 80–130 миллионов человек. дальтон.[65]
Хотя обычно их считают круговой молекулой, есть некоторые свидетельства того, что молекулы хлоропластной ДНК чаще принимают линейный форма.[64][66]
Инвертированные повторы
Многие ДНК хлоропластов содержат два перевернутые повторы, которые отделяют длинную секцию единственной копии (LSC) от короткой секции единственной копии (SSC).[44]Хотя данная пара инвертированных повторов редко бывает полностью идентична, они всегда очень похожи друг на друга, очевидно, в результате согласованная эволюция.[64]
Инвертированные повторы сильно различаются по длине, от 4000 до 25000. пар оснований каждый из них содержит от четырех до более 150 генов.[64] Инвертированные повторы у растений, как правило, находятся в верхней части этого диапазона, каждый из которых имеет длину 20 000–25 000 пар оснований.[44][67]
Области инвертированного повтора очень консервированный среди наземных растений и накапливают мало мутаций.[44][67] Подобные инвертированные повторы существуют в геномах цианобактерий и двух других линий хлоропластов (глаукофита и rhodophyceae ), предполагая, что они предшествуют хлоропласту,[64] хотя некоторые ДНК хлоропластов с тех пор потеряли[67][68] или перевернул перевернутые повторы (сделав их прямыми повторами).[64] Возможно, что инвертированные повторы помогают стабилизировать остальную часть генома хлоропластов, поскольку ДНК хлоропластов, которые потеряли некоторые из сегментов инвертированных повторов, имеют тенденцию больше перестраиваться.[68]
Нуклеоиды
Новые хлоропласты могут содержать до 100 копий своей ДНК,[42] хотя количество копий ДНК хлоропластов уменьшается примерно до 15–20 с возрастом хлоропластов.[69] Обычно они упаковываются в нуклеоиды, который может содержать несколько одинаковых колец ДНК хлоропластов. В каждом хлоропласте можно найти множество нуклеоидов.[65]В примитивном красные водоросли, нуклеоиды ДНК хлоропласта сгруппированы в центре хлоропласта, в то время как у зеленых растений и зеленые водоросли, нуклеоиды рассредоточены по всей строма.[70]
Хотя ДНК хлоропластов не связана с истинным гистоны,[10] в красные водоросли, подобные белки, которые плотно упаковывают каждое кольцо ДНК хлоропласта в нуклеоид были найдены.[70]
Ремонт ДНК
В хлоропластах мха Physcomitrella patens, то Ремонт несоответствия ДНК белок Msh1 взаимодействует с рекомбинационный ремонт белки RecA и RecG для поддержания хлоропластов геном стабильность.[71] В хлоропластах растения Arabidopsis thaliana то RecA белок поддерживает целостность ДНК хлоропласта с помощью процесса, который, вероятно, включает рекомбинационную репарацию Повреждение ДНК.[72]
Репликация ДНК
Ведущая модель репликации хпДНК

Механизм репликации хлоропластной ДНК (хпДНК) окончательно не определен, но были предложены две основные модели. Ученые попытались наблюдать репликацию хлоропластов с помощью электронная микроскопия с 1970-х гг.[73][74] Результаты экспериментов с микроскопией привели к идее, что ДНК хлоропластов реплицируется с использованием петли двойного смещения (D-петля). По мере того, как D-петля движется через кольцевую ДНК, она принимает промежуточную форму тета, также известную как промежуточное звено репликации Кэрнса, и завершает репликацию с помощью механизма катящегося круга.[73][66] Транскрипция начинается в определенных точках происхождения. Открываются множественные репликационные вилки, позволяя репликационному аппарату транскрибировать ДНК. По мере продолжения репликации вилки разрастаются и в конечном итоге сходятся. Новые структуры хпДНК разделяются, образуя дочерние хромосомы хпДНК.
В дополнение к ранним экспериментам по микроскопии, эта модель также подтверждается количеством дезаминирование видно в хпДНК.[73] Дезаминирование происходит при потере аминогруппы и представляет собой мутацию, которая часто приводит к изменениям оснований. Когда аденин дезаминируется, он становится гипоксантин. Гипоксантин может связываться с цитозином, и когда пара оснований XC реплицируется, он становится GC (таким образом, происходит изменение основания A → G).[75]

Дезаминирование
В хпДНК существует несколько градиентов дезаминирования A → G. ДНК становится восприимчивой к событиям дезаминирования, когда она является одноцепочечной. Когда образуются репликационные вилки, не копируемая нить является однонитевой и, следовательно, подвержена риску дезаминирования A → G. Следовательно, градиенты дезаминирования указывают на то, что вилки репликации, скорее всего, присутствовали, и направление, в котором они изначально открывались (самый высокий градиент, скорее всего, находится ближе всего к месту старта, потому что он был одноцепочечным в течение самого длительного периода времени).[73] Этот механизм до сих пор является ведущей теорией; однако вторая теория предполагает, что большая часть хпДНК на самом деле линейна и реплицируется посредством гомологичной рекомбинации. Далее он утверждает, что только меньшая часть генетического материала хранится в кольцевых хромосомах, тогда как остальная часть находится в разветвленных, линейных или других сложных структурах.[73][66]
Альтернативная модель репликации
Одна из конкурирующих моделей репликации хпДНК утверждает, что большая часть хпДНК является линейной и участвует в гомологичная рекомбинация и репликационные структуры, подобные линейным и кольцевым структурам ДНК бактериофаг Т4.[66][76] Было установлено, что некоторые растения имеют линейную хпДНК, например кукурузу, и что многие виды все еще содержат сложные структуры, которые ученые еще не понимают.[66] Когда были выполнены оригинальные эксперименты с хпДНК, ученые действительно заметили линейные структуры; однако они приписали эти линейные формы ломаным кругам.[66] Если разветвленные и сложные структуры, наблюдаемые в экспериментах с хпДНК, являются реальными, а не артефактами конкатенированной кольцевой ДНК или разорванных кругов, то механизма репликации D-петли недостаточно, чтобы объяснить, как эти структуры будут реплицироваться.[66] В то же время гомологичная рекомбинация не расширяет множественные градиенты A -> G, наблюдаемые в пластомах.[73] Из-за неспособности объяснить градиент дезаминирования, а также многочисленных видов растений, которые, как было показано, имеют кольцевую хпДНК, преобладающая теория продолжает утверждать, что большая часть хпДНК является кольцевой и, скорее всего, реплицируется через механизм D-петли.
Содержание генов и синтез белка
Геном хлоропласта обычно включает около 100 генов.[26][43] этот код для множества вещей, в основном связанных с протеиновый трубопровод и фотосинтез. Как в прокариоты, гены в ДНК хлоропластов организованы в опероны.[26] В отличие от прокариотический Молекулы ДНК, молекулы ДНК хлоропластов содержат интроны (растение митохондриальные ДНК тоже, но не мтДНК человека).[77]
Среди наземных растений состав генома хлоропластов довольно схож.[44]
Редукция генома хлоропласта и перенос генов
Со временем многие части генома хлоропластов были перенесены в ядерный геном хозяина,[42][43][78] процесс, называемый перенос эндосимбиотического гена. В результате геном хлоропласта сильно поврежден. уменьшенный по сравнению со свободноживущими цианобактериями. Хлоропласты могут содержать 60–100 генов, тогда как цианобактерии часто имеют более 1500 генов в своем геноме.[79] Недавно была обнаружена пластида без генома, демонстрирующая, что хлоропласты могут потерять свой геном во время эндосимбиотического процесса переноса гена.[80]
Перенос эндосимбиотического гена - вот откуда мы знаем о потерянные хлоропласты во многих линиях CASH. Даже если хлоропласт в конечном итоге потерян, гены, которые он передал ядру бывшего хозяина, сохраняются, что свидетельствует о существовании утраченного хлоропласта. Например, пока диатомеи (а гетероконтофит ) теперь есть хлоропласт, полученный из красных водорослей, наличие многих зеленые водоросли гены в ядре диатомовых водорослей свидетельствуют о том, что предок диатомовых водорослей имел хлоропласт, полученный из зеленых водорослей в какой-то момент, который впоследствии был заменен красным хлоропластом.[46]
У наземных растений около 11–14% ДНК в их ядрах можно проследить до хлоропластов,[41] до 18% в Арабидопсис, что соответствует примерно 4500 генам, кодирующим белок.[81] В последнее время произошло несколько переносов генов из ДНК хлоропластов в ядерный геном наземных растений.[43]
Из примерно 3000 белков, обнаруженных в хлоропластах, около 95% кодируются ядерными генами. Многие белковые комплексы хлоропласта состоят из субъединиц как генома хлоропласта, так и ядерного генома хозяина. Как результат, синтез белка должны быть скоординированы между хлоропластом и ядром. Хлоропласт в основном находится под ядерным контролем, хотя хлоропласты также могут подавать сигналы, регулирующие экспрессия гена в ядре, называемом ретроградная сигнализация.[82]
Синтез белка
Синтез белка в хлоропластах зависит от двух РНК-полимеразы. Один кодируется ДНК хлоропласта, другой - ядерный источник. Две РНК-полимеразы могут распознавать и связываться с разными видами промоутеры в геноме хлоропласта.[83] В рибосомы в хлоропластах похожи на бактериальные рибосомы.[84]
Эта секция нуждается в расширении с: Различия в размере генома между водорослями и наземными растениями, хлоропластный материал, кодируемый ядром. Вы можете помочь добавляя к этому. (Январь 2013) |
Нацеливание на белок и импорт
Поскольку так много генов хлоропластов было перемещено в ядро, многие белки это первоначально было бы переведено в хлоропласте теперь синтезируются в цитоплазме растительной клетки. Эти белки должны быть возвращены в хлоропласт и импортированы как минимум через две хлоропластные мембраны.[85]
Любопытно, что около половины белковых продуктов перенесенных генов даже не направляются обратно в хлоропласт. Многие стали Exaptations, беря на себя новые функции, такие как участие в деление клеток, маршрутизация белков, и даже устойчивость к болезням. Несколько генов хлоропластов нашли новые дома в митохондриальный геном - большинство стало нефункциональным псевдогены хотя несколько тРНК гены все еще работают в митохондрия.[79] Некоторые перенесенные белковые продукты хлоропластной ДНК направляются в секреторный путь,[79] хотя многие вторичные пластиды ограничены самой внешней мембраной, происходящей из клеточная мембрана, и поэтому топологически вне клетки, чтобы добраться до хлоропласта из цитозоль, то клеточная мембрана должен быть пересечен, что означает вход в внеклеточное пространство. В этих случаях белки, нацеленные на хлоропласты, первоначально перемещаются по секреторному пути.[34]
Потому что клетка, приобретающая хлоропласт уже имел митохондрии (и пероксисомы, а клеточная мембрана секреции), новый хозяин хлоропласта должен был развить уникальную система нацеливания на белок чтобы избежать попадания белков хлоропластов не туда органелла.[85]
![Два конца полипептида называются N-концом, или амино-концом, и С-концом, или карбоксильным концом. [86] Этот полипептид состоит из четырех аминокислот, связанных вместе. Слева находится N-конец с его аминогруппой (H2N) зеленым цветом. Синий С-конец с его карбоксильной группой (CO2H) находится справа.](http://upload.wikimedia.org/wikipedia/commons/thumb/c/c0/Tetrapeptide_structural_formulae.svg/370px-Tetrapeptide_structural_formulae.svg.png)
В большинстве, но не во всех случаях, белки хлоропластов, кодируемые ядром, являются переведено с расщепляемый транзитный пептид это добавлено к N-концу предшественника белка. Иногда транзитная последовательность обнаруживается на С-конце белка,[87] или в функциональной части белка.[85]
Транспортные белки и мембранные транслоконы
После хлоропласта полипептид синтезируется на рибосома в цитозоль, фермент специфический к белкам хлоропластов[88] фосфорилаты, или добавляет фосфатная группа многим (но не всем) из них в их транзитных последовательностях.[85]Фосфорилирование помогает многим белкам связывать полипептид, не давая ему складывание преждевременно.[85] Это важно, потому что это не позволяет белкам хлоропластов принимать свою активную форму и выполнять свои функции хлоропластов в неправильном месте - цитозоль.[89][90] В то же время они должны сохранять форму, достаточную для того, чтобы их можно было распознать по хлоропласту.[89] Эти белки также помогают полипептиду проникать в хлоропласт.[85]
Отсюда белки хлоропластов, связанные со стромой, должны проходить через два белковых комплекса - Комплекс ТОС, или же тRnslocon на оматка cхлоропластная мембрана, а ТИЦ транслокон, или же тranslocon на яннер cхлоропластная мембрана Translocon.[85] Полипептидные цепи хлоропластов, вероятно, часто проходят через два комплекса одновременно, но комплекс TIC может также извлекать препротеины, потерянные в межмембранное пространство.[85]
Структура
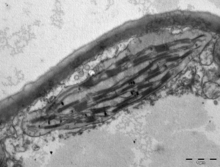
В наземные растения хлоропласты обычно имеют линзовидную форму, диаметр 3–10 мкм и толщину 1–3 мкм.[91][20] Размер хлоропластов проростков кукурузы ≈20 мкм.3 в объеме.[20] Большее разнообразие форм хлоропластов существует среди водоросли, которые часто содержат единственный хлоропласт[16] которые могут иметь форму сети (например, Эдогониум ),[92] чашка (например, Хламидомонада ),[93] ленточная спираль по краям ячейки (например, Спирогира ),[94] или слегка закрученные полосы по краям ячеек (например, Сирогониум ).[95] Некоторые водоросли имеют по два хлоропласта в каждой ячейке; они в форме звезды Зигнема,[96] или может повторять форму половины ячейки в порядок Desmidiales.[97] У некоторых водорослей хлоропласт занимает большую часть клетки с карманами для ядро и другие органеллы,[16] например, некоторые виды Хлорелла имеют чашевидный хлоропласт, занимающий большую часть клетки.[98]
Все хлоропласты имеют по крайней мере три мембранные системы - внешнюю мембрану хлоропласта, внутреннюю мембрану хлоропласта и тилакоид система. Хлоропласты, являющиеся продуктом вторичный эндосимбиоз могут иметь дополнительные мембраны, окружающие эти три.[45] Внутри внешней и внутренней мембран хлоропласта находится хлоропласт. строма, полужелеподобная жидкость[32] который составляет большую часть объема хлоропласта и в котором плавает тилакоидная система.


Существует несколько распространенных заблуждений о внешней и внутренней мембранах хлоропластов. Тот факт, что хлоропласты окружены двойной мембраной, часто приводится как доказательство того, что они являются потомками эндосимбиотических цианобактерии. Это часто интерпретируется как то, что внешняя мембрана хлоропласта является продуктом организма хозяина. клеточная мембрана складывается, образуя пузырек, окружающий предков цианобактерии - что неверно - обе мембраны хлоропластов гомологичный к исходным двойным мембранам цианобактерий.[18]
Двойную мембрану хлоропласта также часто сравнивают с митохондриальный двойная мембрана. Это неверное сравнение - внутренняя мембрана митохондрий используется для работы протонные насосы и выполнять окислительного фосфорилирования поперек, чтобы произвести АТФ энергия. Единственная структура хлоропласта, которую можно рассматривать аналогичный это внутренняя тилакоидная система. Тем не менее, с точки зрения «внутрь-наружу», направление хлоропласта ЧАС+ ионный поток имеет противоположное направление по сравнению с окислительным фосфорилированием в митохондриях.[32][99] Кроме того, с точки зрения функции внутренняя мембрана хлоропласта, которая регулирует прохождение метаболитов и синтезирует некоторые материалы, не имеет аналогов в митохондриях.[32]
Наружная хлоропластная мембрана
Наружная мембрана хлоропласта представляет собой полупористую мембрану, в которой небольшие молекулы и ионы может легко распространяться.[100] Однако он не проницаем для более крупных белки, так хлоропласт полипептиды синтезируется в клетке цитоплазма должны транспортироваться через внешнюю мембрану хлоропласта Комплекс ТОС, или же тRnslocon на оматка cхлоропласт мембрана.[85]
Мембраны хлоропластов иногда выступают в цитоплазму, образуя стромула, или же Stromа -содержащая ваннауле. Стромулы очень редки в хлоропластах и гораздо чаще встречаются в других пластиды подобно хромопласты и амилопласты в лепестках и корнях соответственно.[101][102] Они могут существовать для увеличения хлоропластов. площадь поверхности для кросс-мембранного транспорта, потому что они часто разветвляются и запутываются эндоплазматический ретикулум.[103] Когда они были впервые обнаружены в 1962 году, некоторые биологи растений отклонили эти структуры как артефакты, заявив, что стромулы представляют собой просто хлоропласты странной формы с суженными участками. деление хлоропластов.[104] Однако появляется все больше свидетельств того, что стромулы являются функциональными, неотъемлемыми характеристиками пластид растительных клеток, а не просто артефактами.[105]
Межмембранное пространство и пептидогликановая стенка

Обычно тонкое межмембранное пространство около 10–20 нанометры толстая существует между внешней и внутренней мембранами хлоропластов.[106]
Глаукофитные водоросли хлоропласты имеют пептидогликан слой между хлоропластными мембранами. Это соответствует клеточная стенка пептидогликана от их цианобактериальный предков, который расположен между двумя их клеточными мембранами. Эти хлоропласты называются муропласты (от латинского "мура", что означает «стена»). Другие хлоропласты утратили цианобактериальную стенку, оставив межмембранное пространство между двумя оболочками хлоропластов.[32]
Внутренняя хлоропластная мембрана
Внутренняя мембрана хлоропласта граничит со стромой и регулирует прохождение материалов в хлоропласт и из него. Пройдя через Комплекс ТОС в наружной мембране хлоропласта, полипептиды должен пройти через ТИЦ комплекс (тRnslocon на яннер cхлоропластная мембрана) который расположен во внутренней мембране хлоропласта.[85]
Помимо регулирования прохождения материалов, внутренняя мембрана хлоропласта находится там, где жирные кислоты, липиды, и каротиноиды синтезированы.[32]
Периферический ретикулум
Некоторые хлоропласты содержат структуру, называемую периферический ретикулум хлоропластов.[106] Часто встречается в хлоропластах C4 растения, хотя он также был обнаружен в некоторых C3 покрытосеменные,[32] и даже некоторые голосеменные.[107] Периферический ретикулум хлоропластов состоит из лабиринта перепончатых трубок и пузырьков, продолжающихся внутренняя хлоропластная мембрана что простирается во внутреннюю стромальный жидкость хлоропласта. Считается, что его цель состоит в том, чтобы увеличить количество хлоропластов. площадь поверхности для межмембранного транспорта между стромой и клеткой цитоплазма. Иногда наблюдаемые мелкие пузырьки могут служить транспортные везикулы перебрасывать вещи между тилакоиды и межмембранное пространство.[108]
Строма
В белок -богатые,[32] щелочной,[99] водный жидкость внутри внутренней мембраны хлоропласта и за пределами тилакоидного пространства называется стромой,[32] что соответствует цитозоль оригинала цианобактерии. Нуклеоиды из хлоропластная ДНК, хлоропласт рибосомы, тилакоидная система с пластоглобулы, крахмал гранулы и многие белки можно найти плавающим в нем. В Цикл Кальвина, который исправляет CO2 в G3P проходит в строме.
Рибосомы хлоропластов

У хлоропластов есть собственные рибосомы, которые они используют для синтеза небольшой части своих белков. Рибосомы хлоропласта составляют примерно две трети размера цитоплазматические рибосомы (около 17 нм и 25 нм ).[106] Они берут мРНК переписано с хлоропластная ДНК и переведите их в белок. Хотя похоже на бактериальные рибосомы,[10] Трансляция хлоропластов более сложна, чем у бактерий, поэтому рибосомы хлоропластов обладают некоторыми уникальными особенностями хлоропластов.[109]Малая единица рибосомные РНК в нескольких Хлорофита и эвгленида в хлоропластах отсутствуют мотивы для последовательность шайн-далгарно признание,[110] что считается важным для перевод инициация в большинстве хлоропластов и прокариоты.[111][112] Такая потеря также редко наблюдается в других пластиды и прокариоты.[110][113]
Пластоглобулы
Пластоглобулы (единственное число пластоглобурус, иногда пишется пластоглобула (ы)), представляют собой сферические пузыри липиды и белки[32] около 45–60 нанометров в поперечнике.[114] Они окружены липидный монослой.[114] Пластоглобулы встречаются во всех хлоропластах,[106] но становятся более распространенными, когда хлоропласт находится под окислительный стресс,[114] или когда он стареет и переходит в геронтопласт.[32] Пластоглобулы также демонстрируют большее изменение размера в этих условиях.[114] Они также распространены в этиопласты, но количество уменьшается по мере созревания этиопластов в хлоропласты.[114]
Пластоглубули содержат как структурные белки, так и ферменты, участвующие в липидный синтез и метаболизм. Они содержат много видов липиды включая пластохинон, витамин Е, каротиноиды и хлорофиллы.[114]
Когда-то считалось, что пластоглобулы свободно плавают в строма, но теперь считается, что они постоянно прикреплены либо к тилакоид или к другому пластоглобулусу, присоединенному к тилакоиду, конфигурация, которая позволяет пластоглобулу обмениваться своим содержимым с сетью тилакоидов.[114] В нормальных зеленых хлоропластах подавляющее большинство пластоглобул встречаются по отдельности, прикрепленные непосредственно к своему родительскому тилакоиду. В старых или подверженных стрессу хлоропластах пластоглобулы имеют тенденцию возникать в связанных группах или цепочках, все еще всегда прикрепленных к тилакоиду.[114]
Пластоглобулы образуются при появлении пузыря между слоями липидный бислой тилакоидной мембраны или зачатка существующих пластоглубули, хотя они никогда не отслаиваются и не всплывают в строму.[114] Практически все пластоглобулы образуются на сильно изогнутых краях или вблизи них. тилакоид диски или листы. Они также чаще встречаются на стромальных тилакоидах, чем на Granal ед.[114]
Гранулы крахмала
Гранулы крахмала очень распространены в хлоропластах, обычно занимая 15% объема органелл,[115] хотя в некоторых других пластидах, например амилопласты, они могут быть достаточно большими, чтобы исказить форму органеллы.[106] Гранулы крахмала - это просто скопления крахмала в строме, которые не ограничены мембраной.[106]
Гранулы крахмала появляются и растут в течение дня, поскольку хлоропласт синтезирует сахара, а ночью расходуются на топливо дыхание и продолжить экспорт сахара в флоэма,[116] хотя в зрелых хлоропластах гранула крахмала полностью израсходуется или накапливается новая гранула.[115]
Гранулы крахмала различаются по составу и расположению в разных линиях хлоропластов. В красные водоросли, гранулы крахмала находятся в цитоплазма а не в хлоропласте.[117] В C4 растения, мезофилл хлоропласты, не синтезирующие сахара, лишены гранул крахмала.[32]
RuBisCO
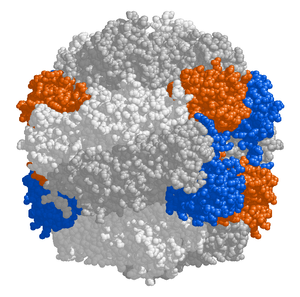
Строма хлоропластов содержит много белков, но наиболее распространенным и важным является RuBisCO, который, вероятно, также является самым распространенным белком на планете.[99] RuBisCO это фермент, который фиксирует CO2 в молекулы сахара. В C3 растения, RuBisCO присутствует во всех хлоропластах, но в C4 растения, он ограничен связка ножен хлоропласты, где Цикл Кальвина выполняется в C4 растения.[118]
Пиреноиды
Хлоропласты некоторых роголистник[119] а водоросли содержат структуры, называемые пиреноиды. Их нет у высших растений.[120] Пиреноиды представляют собой примерно сферические тела с высокой преломляющей способностью, которые являются местом накопления крахмала в растениях, которые их содержат. Они состоят из матрицы, непрозрачной для электронов, окруженной двумя полусферическими крахмальными пластинами. Крахмал накапливается по мере созревания пиреноидов.[121] В водорослях с механизмы концентрации углерода, фермент RuBisCO находится в пиреноидах. Крахмал также может накапливаться вокруг пиреноидов, когда CO2 мало.[120] Пиреноиды могут делиться с образованием новых пиреноидов или образовываться. "de novo".[121][122]
Тилакоидная система

(Вверху) STEM томографический срез хлоропласта салата толщиной 10 нм. Стеки граны связаны между собой несъёмными стромальными тилакоидами, называемыми «ламеллами стромы». Круглые включения, связанные с тилакоидами, представляют собой пластоглобулы. Масштабная шкала = 200 нм. Видеть.[123]
(Внизу) Крупномасштабная 3D-модель, созданная в результате сегментации томографических реконструкций с помощью STEM. грана = желтый; ламели стромы = зеленые; пластоглобулы = пурпурный; Оболочка хлоропласта = синяя. Видеть [123].
Тилакоиды (иногда пишут тилакоиды),[124] представляют собой небольшие взаимосвязанные мешочки, содержащие мембраны, которые легкие реакции фотосинтеза. Слово тилакоид происходит от греческого слова тилакос что означает «мешок».[125]
В строме хлоропласта находится тилакоид система, высокодинамичный набор мембранных мешков, называемый тилакоиды куда хлорофилл найден и легкие реакции из фотосинтез случаться.[15]В большинстве сосудистое растение хлоропласты тилакоиды расположены в стопки, называемые грана,[126] хотя в некоторых C4 растение хлоропласты[118] и немного водоросль хлоропласты тилакоиды свободно плавают.[16]
Тилакоидная структура

Используя оптический микроскоп, едва видны крошечные зеленые гранулы, названные грана.[106] С электронная микроскопия стало возможным более подробно рассмотреть тилакоидную систему, обнаружив, что она состоит из стопок плоских тилакоиды из которых состоит грана, и длинные соединяющие стромальные тилакоиды, которые связывают разные граны.[106]в просвечивающий электронный микроскоп, тилакоидные мембраны выглядят как чередующиеся светлые и темные полосы толщиной 8,5 нанометров.[106]
Долгое время трехмерная структура мембранной системы тилакоидов была неизвестна или оспаривалась. Было предложено много моделей, наиболее распространенной из которых является спиральный модель, в которой гранульные стеки тилакоидов обернуты спиральными стромальными тилакоидами.[127] Другая модель, известная как «модель бифуркации», которая была основана на первом электронном томографическом исследовании тилакоидных мембран растений, изображает стромальные мембраны как широкие пластинки, перпендикулярные столбцам граны, которые раздваиваются на несколько параллельных дисков, образующих сборку гранум-строма.[128] Винтовая модель была поддержана несколькими дополнительными работами,[126][129] но в конечном итоге в 2019 году было определено, что особенности как спиральной, так и бифуркационной модели объединены недавно обнаруженными левосторонними спиральными мембранными переходами.[123] Вероятно, для простоты, тилакоидная система все еще часто изображается в более старых моделях «ступица и спица», где граны соединены друг с другом трубками стромальных тилакоидов.[130]
Грана состоит из стопок уплощенных круглых тилакоидов, напоминающих блинчики. Каждая гранула может содержать от двух до ста тилакоидов,[106] хотя чаще всего встречаются граны с 10–20 тилакоидами.[126] Вокруг граны находится множество параллельных правосторонних спиральных стромальных тилакоидов, также известных как лады или ламеллярные тилакоиды. Спирали поднимаются под углом ~ 20 °, соединяясь с каждым тилакоидом гранала мостиковым щелевым соединением.[126][129][123]
Пластинки стромы простираются в виде больших пластин, перпендикулярных столбцам граны. Эти листы соединены с правыми спиралями либо напрямую, либо через бифуркации, которые образуют левые спиральные поверхности мембраны.[123] Левые винтовые поверхности имеют такой же угол наклона, что и правые спирали (~ 20 °), но ¼ шаг. Приблизительно 4 левых спиральных соединения присутствуют на гранум, что приводит к сбалансированному по шагу массиву правых и левых спиральных мембранных поверхностей разного радиуса и шага, которые консолидируют сеть с минимальной энергией поверхности и изгиба.[123] В то время как разные части тилакоидной системы содержат разные мембранные белки, тилакоидные мембраны непрерывны, а тилакоидное пространство, которое они окружают, образуют единый непрерывный лабиринт.[126]
Тилакоидный состав
Встроенные в тилакоидные мембраны важны белковые комплексы которые выполняют легкие реакции из фотосинтез. Фотосистема II и фотосистема I содержать светоуборочные комплексы с хлорофилл и каротиноиды которые поглощают световую энергию и используют ее для возбуждения электронов. Молекулы в тилакоидной мембране используют заряженные электроны для накачки ионы водорода в тилакоидное пространство, уменьшая pH и превращая его в кислоту. АТФ-синтаза это большой белковый комплекс, который использует градиент концентрации ионов водорода в тилакоидном пространстве для генерации АТФ энергия, когда ионы водорода текут обратно в строму - очень похоже на плотинную турбину.[99]
Существует два типа тилакоидов - тилакоиды граналов, которые расположены в гранах, и тилакоиды стромы, которые контактируют с строма. Гранальные тилакоиды представляют собой круглые диски в форме блинов диаметром около 300–600 нанометров. Стромальные тилакоиды геликоид листы, которые закручиваются вокруг граны.[126] Плоские вершины и днище тилакоидов граналя содержат только относительно плоские фотосистема II белковый комплекс. Это позволяет им плотно складываться, образуя грану с множеством слоев плотно прижатой мембраны, называемой гранальной мембраной, что увеличивает стабильность и площадь поверхности для светового захвата.[126]
В отличие, фотосистема I и АТФ-синтаза представляют собой большие белковые комплексы, которые выступают в строму. Они не могут поместиться в прижатых гранальных мембранах, и поэтому обнаруживаются в стромальной тилакоидной мембране - краях тилакоидных дисков граналей и стромальных тилакоидах. Эти большие белковые комплексы могут действовать как разделители между листами стромальных тилакоидов.[126]
На количество тилакоидов и общую площадь тилакоидов хлоропласта влияет воздействие света. Затененные хлоропласты содержат больше и больше грана с большей площадью тилакоидной мембраны, чем хлоропласты, освещенные ярким светом, которые имеют все меньше и меньше гран и меньшую площадь тилакоидов. Степень тилакоида может измениться в течение нескольких минут после воздействия или удаления света.[108]
Пигменты и красители хлоропластов
Внутри фотосистем, встроенных в тилакоидные мембраны хлоропластов, различны фотосинтетические пигменты, которые поглощают и переносят Световая энергия. Типы обнаруженных пигментов различны в разных группах хлоропластов и отвечают за большое разнообразие окраски хлоропластов.

Хлорофиллы
Хлорофилл а содержится во всех хлоропластах, а также в их цианобактериальный предки. Хлорофилл а это цвет морской волны пигмент[131] частично отвечает за окраску большинства цианобактерий и хлоропластов. Существуют и другие формы хлорофилла, такие как вспомогательные пигменты хлорофилл б, хлорофилл c, хлорофилл d,[16] и хлорофилл ж.
Хлорофилл б является оливково-зеленый пигмент содержится только в хлоропластах растения, зеленые водоросли, любые вторичные хлоропласты, полученные вторичный эндосимбиоз зеленой водоросли и несколько цианобактерии.[16] Это хлорофиллы а и б вместе они делают большую часть хлоропластов растений и зеленых водорослей зелеными.[131]
Хлорофилл c в основном обнаруживается во вторичных эндосимбиотических хлоропластах, происходящих из красная водоросль, хотя в самих хлоропластах красных водорослей он не обнаружен. Хлорофилл c также встречается в некоторых зеленые водоросли и цианобактерии.[16]
Хлорофиллы d и ж пигменты встречаются только у некоторых цианобактерий.[16][132]
Каротиноиды
![У красной водоросли Delesseria sanguinea есть хлоропласты, содержащие красные пигменты, такие как фикоэритерин, которые маскируют их сине-зеленый хлорофилл а [35].](http://upload.wikimedia.org/wikipedia/commons/thumb/1/19/Delesseria_sanguinea_Helgoland.JPG/250px-Delesseria_sanguinea_Helgoland.JPG)
В добавление к хлорофиллы, другая группа желтый –апельсин[131] пигменты под названием каротиноиды также встречаются в фотосистемах. Есть около тридцати фотосинтетических каротиноидов.[133] Они помогают передавать и рассеивать избыточную энергию,[16] и их яркие цвета иногда преобладают над зеленым хлорофиллом, как во время Осень, когда листья некоторые наземные растения сменить цвет.[134] β-каротин представляет собой ярко-красно-оранжевый каротиноид, обнаруженный почти во всех хлоропластах, например хлорофилл а.[16] Ксантофиллы, особенно оранжево-красный зеаксантин, также распространены.[133] Существуют многие другие формы каротиноидов, которые встречаются только в определенных группах хлоропластов.[16]
Фикобилины
Фикобилины третья группа пигментов, содержащихся в цианобактерии, и глаукофит, красные водоросли, и криптофит хлоропласты.[16][135] Однако фикобилины бывают всех цветов. фикоэритерин является одним из пигментов, придающих красный цвет многим красным водорослям.[136] Фикобилины часто образуют относительно большие белковые комплексы размером около 40 нанометров, называемые фикобилисомы.[16] Нравиться фотосистема I и АТФ-синтаза фикобилисомы выступают в строму, предотвращая накопление тилакоидов в хлоропластах красных водорослей.[16] Криптофит В хлоропластах и некоторых цианобактериях фикобилиновые пигменты не организованы в фикобилисомы, а вместо этого они удерживаются в тилакоидном пространстве.[16]
| Фотосинтетические пигменты Таблица наличия различных пигментов в группах хлоропластов. Цветные клетки представляют присутствие пигмента.[16][133][135] | |||||||||
| Хлорофилла | Хлорофиллб | Хлорофиллc | Хлорофилл d и ж | Ксантофиллы | α-каротин | β-каротин | Фикобилины | ||
| Наземные растения | |||||||||
| Зеленые водоросли | |||||||||
| Эвгленофиты и Хлорарахниофиты | |||||||||
| Многоклеточные красные водоросли | |||||||||
| Одноклеточные красные водоросли | |||||||||
| Гаптофиты и Динофиты | |||||||||
| Криптофиты | |||||||||
| Глаукофиты | |||||||||
| Цианобактерии | |||||||||
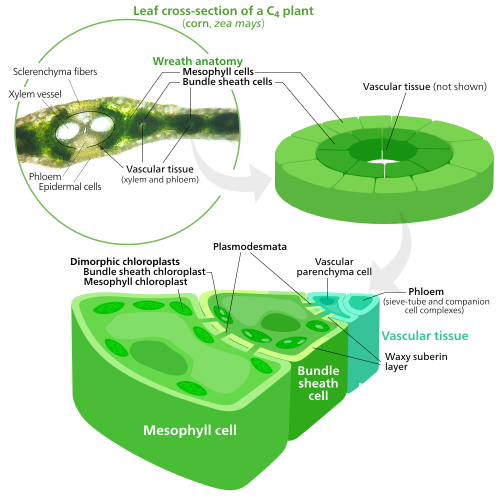
Специализированные хлоропласты в C4 растения
Чинить углекислый газ в молекулы сахара в процессе фотосинтез, хлоропласты используют фермент, называемый RuBisCO. У RuBisCO есть проблема - ему трудно различать углекислый газ и кислород, поэтому при высоких концентрациях кислорода RuBisCO начинает случайно добавлять кислород к прекурсорам сахара. Это имеет конечный результат АТФ энергия тратится и CO
2 выпускается без производства сахара. Это большая проблема, поскольку O2 производится начальным легкие реакции фотосинтеза, вызывая проблемы в Цикл Кальвина который использует RuBisCO.[137]
C4 растения разработали способ решить эту проблему - пространственно разделив световые реакции и цикл Кальвина. Световые реакции, которые накапливают световую энергию в АТФ и НАДФН, сделаны в мезофилл ячейки C4 лист. Цикл Кальвина, который использует накопленную энергию для производства сахара с помощью RuBisCO, выполняется в связка клеток оболочки, слой клеток, окружающий вена в лист.[137]
В результате хлоропласты в C4 Клетки мезофилла и клетки оболочки пучка специализированы для каждой стадии фотосинтеза. В клетках мезофилла хлоропласты специализируются на световых реакциях, поэтому в них отсутствуют RuBisCO, и нормальный грана и тилакоиды,[118] которые они используют для производства АТФ и НАДФН, а также кислорода. Они хранят CO
2 в четырехуглеродном соединении, поэтому процесс называется C4 фотосинтез. Затем четырехуглеродное соединение транспортируется к хлоропластам оболочки пучка, где оно выпадает. CO
2 и возвращается к мезофиллу. Хлоропласты связки оболочки не проводят световые реакции, препятствуя накоплению в них кислорода и нарушая активность RuBisCO.[137] Из-за этого в них отсутствуют тилакоиды, организованные в грана стеки - хотя хлоропласты оболочек пучков все еще имеют свободно плавающие тилакоиды в строме, где они все еще проводят циклический поток электронов, световой метод синтеза АТФ для питания цикла Кальвина без образования кислорода. Им не хватает фотосистема II, и только фотосистема I - единственный белковый комплекс, необходимый для циклического электронного потока.[118][137] Поскольку задача хлоропластов связки оболочки - выполнять цикл Кальвина и производить сахар, они часто содержат большое количество крахмал зерна.[118]
Оба типа хлоропластов содержат большое количество периферический ретикулум хлоропластов,[118] которые они используют, чтобы получить больше площадь поверхности для перевозки вещей в них и из них.[107][108] Хлоропласты мезофилла имеют немного больший периферический ретикулум, чем хлоропласты пучковой оболочки.[138]
Место расположения
Распространение на заводе
Не все клетки многоклеточного растения содержат хлоропласты. Все зеленые части растения содержат хлоропласты - хлоропласты, а точнее, хлорофилл в них то, что делает фотосинтетические части растения зелеными.[15] В клетки растений которые содержат хлоропласты, обычно паренхима клетки, хотя хлоропласты также можно найти в колленхима ткань.[139] Клетка растения, содержащая хлоропласты, известна как хлоренхима клетка. Типичный хлоренхима клетка наземного растения содержит от 10 до 100 хлоропластов.

У некоторых растений, например кактусы, хлоропласты находятся в стебли,[140] хотя у большинства растений хлоропласты сосредоточены в листья. Один квадрат миллиметр ткани листа может содержать полмиллиона хлоропластов.[15] Внутри листа хлоропласты в основном находятся в мезофилл слои лист, а замыкающие клетки из устьица. Палисадный мезофилл клетки могут содержать 30–70 хлоропластов на клетку, в то время как замыкающие клетки устьиц содержат только около 8–15 на клетку, а также гораздо меньше хлорофилл. Хлоропласты также можно найти в связка ножен клетки листа, особенно в C4 растения, которые проводят Цикл Кальвина в их клетках-оболочках пучка. Они часто отсутствуют в эпидермис листа.[141]
Сотовая связь
Хлоропластное движение
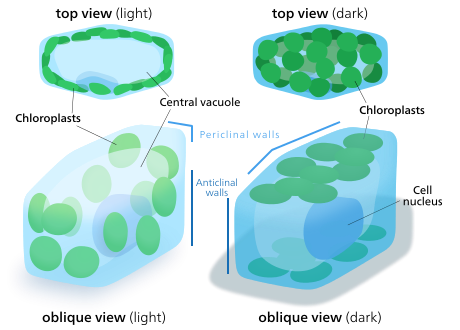
Хлоропласты клеток растений и водорослей могут ориентироваться в соответствии с доступным светом. В условиях низкой освещенности они раскладываются по листу, увеличивая площадь поверхности для поглощения света. При ярком свете они будут искать укрытие, выстраиваясь вертикальными колоннами вдоль растительной клетки. клеточная стенка или поворачиваясь боком, чтобы свет падал на них с ребра. Это снижает воздействие и защищает их от фотоокислительное повреждение.[142] Эта способность распределять хлоропласты таким образом, чтобы они могли укрываться друг за другом или распространяться, может быть причиной того, что наземные растения эволюционировали, чтобы иметь много маленьких хлоропластов вместо нескольких больших.[143]Движение хлоропластов считается одной из наиболее строго регулируемых систем стимул-реакция, которые можно найти у растений.[144] Митохондрии также наблюдали за движением хлоропластов.[145]
У высших растений движением хлоропластов управляет фототропины, синий свет фоторецепторы также отвечает за завод фототропизм. В некоторых водорослях мхи, папоротники, и цветущие растения, на движение хлоропластов помимо синего влияет красный свет,[142] хотя очень длинные волны красного цвета скорее препятствуют движению, чем ускоряют его. Синий свет обычно заставляет хлоропласты искать убежища, а красный свет вытягивает их, чтобы максимизировать поглощение света.[145]
Исследования Валлиснерия гигантская, водный цветущее растение, показали, что хлоропласты могут двигаться в течение пяти минут после воздействия света, хотя изначально они не проявляют никакой чистой направленности. Они могут двигаться дальше микрофиламент треков, и тот факт, что сетка из микрофиламентов меняет форму, образуя сотовую структуру, окружающую хлоропласты после их перемещения, предполагает, что микрофиламенты могут помочь закрепить хлоропласты на месте.[144][145]
Функция и химия
Хлоропласты охранных клеток
Эта секция нуждается в расширении с: определенные функции, противоречивые функции, характеристики и совокупность. Вы можете помочь добавляя к этому. (август 2013) |
В отличие от большинства клеток эпидермиса, замыкающие клетки растения устьица содержат относительно хорошо развитые хлоропласты.[141] Однако то, что они делают, вызывает споры.[146]
Врожденный иммунитет растений
Растения не хватает специализированных иммунные клетки - все клетки растения участвуют в растении иммунная реакция. Хлоропласты вместе с ядро, клеточная мембрана, и эндоплазматический ретикулум,[147] ключевые игроки в возбудитель защита. Из-за своей роли в иммунном ответе растительной клетки патогены часто нацелены на хлоропласт.[147]
У растений есть два основных иммунных ответа: гиперчувствительный ответ, в которых инфицированные клетки изолируются и подвергаются запрограммированная гибель клеток, и системная приобретенная резистентность, где инфицированные клетки излучают сигналы, предупреждающие остальную часть растения о присутствии патогена. Хлоропласты стимулируют оба ответа, намеренно повреждая их фотосинтетическую систему, производя активные формы кислорода. Высокий уровень активных форм кислорода вызовет гиперчувствительный ответ. Активные формы кислорода также напрямую убивают любые патогены внутри клетки. Более низкие уровни активных форм кислорода инициируют системная приобретенная резистентность, запуская продукцию защитных молекул в остальной части растения.[147]
Известно, что у некоторых растений хлоропласты приближаются к очагу инфекции и ядро во время инфекции.[147]
Хлоропласты могут служить клеточными сенсорами. После обнаружения стресса в клетке, который может быть вызван патогеном, хлоропласты начинают производить такие молекулы, как салициловая кислота, жасмоновая кислота, оксид азота и активные формы кислорода которые могут служить защитными сигналами. В качестве клеточных сигналов активные формы кислорода представляют собой нестабильные молекулы, поэтому они, вероятно, не покидают хлоропласт, а вместо этого передают свой сигнал неизвестной молекуле второго мессенджера. Все эти молекулы инициируют ретроградная сигнализация - сигналы от хлоропласта, регулирующие экспрессия гена в ядре.[147]
Помимо защитной сигнализации, хлоропласты с помощью пероксисомы,[148] помочь синтезировать важную защитную молекулу, жасмонат. Хлоропласты синтезируют все жирные кислоты в растительной клетке[147][149]—линолевая кислота, жирная кислота, является предшественником жасмоната.[147]
Фотосинтез
Одна из основных функций хлоропласта - это его роль в фотосинтез, процесс, с помощью которого свет превращается в химическую энергию, чтобы впоследствии производить пищу в виде сахара. Вода (ЧАС2O) и углекислый газ (CO2) используются в фотосинтезе, а сахар и кислород (O2) производится с использованием Световая энергия. Фотосинтез делится на два этапа: легкие реакции, где вода расщепляется для производства кислорода, а темные реакции, или же Цикл Кальвина, который строит молекулы сахара из углекислого газа. Две фазы связаны энергоносителями аденозинтрифосфат (АТФ) и никотинамидадениндинуклеотидфосфат (НАДФ+).[150][151]
Легкие реакции

На тилакоидных мембранах происходят световые реакции. Они берут Световая энергия и хранить в НАДФН, форма НАДФ+, и АТФ заправлять темные реакции.
Энергоносители
АТФ - это фосфорилированная версия аденозиндифосфат (АДФ), который накапливает энергию в клетке и обеспечивает большую часть клеточной активности. АТФ - это активированная форма, а АДФ - (частично) обедненная форма. НАДФ+ является переносчиком электронов, который переносит электроны высокой энергии. В легких реакциях он становится уменьшенный, что означает, что он собирает электроны, становясь НАДФН.
Фотофосфорилирование
Как и митохондрии, хлоропласты используют потенциальная энергия хранится в ЧАС+, или градиент ионов водорода для выработки энергии АТФ. Два фотосистемы захватить световую энергию, чтобы зарядить энергией электроны взято из воды, и отпустите их электронная транспортная цепь. В молекулы между фотосистемами используют энергию электронов для закачки ионов водорода в тилакоидное пространство, создавая градиент концентрации, с большим количеством ионов водорода (до тысячи раз)[99] внутри тилакоидной системы, чем в строме. Ионы водорода в тилакоидном пространстве затем размытый обратно вниз по градиенту концентрации, возвращаясь обратно в строму через АТФ-синтаза. АТФ-синтаза использует энергию протекающих ионов водорода фосфорилировать аденозиндифосфат в аденозинтрифосфат, или АТФ.[99][152] Поскольку АТФ-синтаза хлоропластов проецируется в строму, АТФ синтезируется там, чтобы использовать ее в темновых реакциях.[153]
НАДФ+ снижение
Электроны часто удаляются из цепи переноса электронов заряжать НАДФ+ с электронами, сокращение это к НАДФН. Подобно АТФ-синтазе, ферредоксин-НАДФ+ редуктаза, фермент, который снижает НАДФ+, высвобождает НАДФН, который он производит, в строму именно там, где он необходим для темных реакций.[153]
Потому что НАДП+ восстановление удаляет электроны из цепей переноса электронов, они должны быть заменены - работа фотосистема II, который разбивает воды молекулы (H2O), чтобы получить электроны из его атомы водорода.[99][150]
Циклическое фотофосфорилирование
Пока фотосистема II фотолизирует вода для получения и возбуждения новых электронов, фотосистема I просто восстанавливает энергию обедненных электронов в конце цепи переноса электронов. Обычно повторно заряженные электроны забираются НАДФ.+, хотя иногда они могут стекать обратно больше H+- накачивание цепей переноса электронов для транспортировки большего количества ионов водорода в тилакоидное пространство для генерации большего количества АТФ. Это называется циклическое фотофосфорилирование потому что электроны рециркулируют. Циклическое фотофосфорилирование распространено в C4 растения, которым нужно больше АТФ чем НАДФН.[137]
Темные реакции
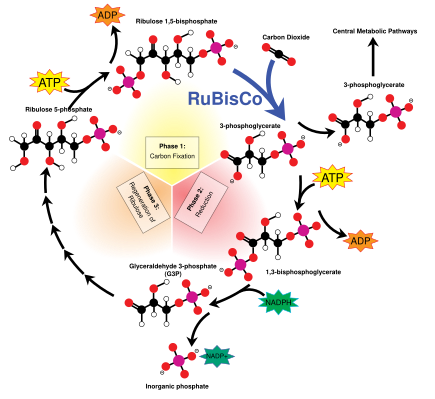
В Цикл Кальвина, также известный как темные реакции, представляет собой серию биохимических реакций, фиксирующих CO2 в G3P молекул сахара и использует энергию и электроны из АТФ и НАДФН сделано в легких реакциях. Цикл Кальвина протекает в строме хлоропласта.[137]
Пока назван "темные реакции", у большинства растений они протекают на свету, поскольку темные реакции зависят от продуктов световых реакций.[15]
Фиксация углерода и синтез G3P
Цикл Кальвина начинается с использования фермента RuBisCO исправить CO2 в пятиуглеродный Рибулоза бисфосфат (RuBP) молекулы. В результате возникают нестабильные молекулы из шести атомов углерода, которые сразу же распадаются на молекулы из трех атомов углерода, называемые 3-фосфоглицериновая кислота, или 3-PGA. АТФ и НАДФН произведенный в световых реакциях используется для преобразования 3-PGA в глицеральдегид-3-фосфат, или молекулы сахара G3P. Большинство молекул G3P рециркулируют обратно в RuBP, используя энергию большего количества АТФ, но каждая шестая произведенная молекула выходит из цикла - конечный продукт темных реакций.[137]
Сахара и крахмалы

Глицеральдегид-3-фосфат может удваиваться, образуя более крупные молекулы сахара, такие как глюкоза и фруктоза. Эти молекулы перерабатываются, и от них тем больше сахароза, а дисахарид широко известный как столовый сахар, производится, хотя этот процесс происходит вне хлоропласта, в цитоплазма.[154]
В качестве альтернативы глюкоза мономеры в хлоропласте могут быть связаны вместе, чтобы сделать крахмал, который накапливается в крахмальные зерна находится в хлоропласте.[154]В таких условиях, как высокое содержание CO в атмосфере2 концентрации, эти зерна крахмала могут вырасти очень большими, искажая грану и тилакоиды. Гранулы крахмала вытесняют тилакоиды, но оставляют их нетронутыми.[155]Заболоченный корни также может вызвать крахмал накопление в хлоропластах, возможно, из-за меньшего сахароза вывозится из хлоропласта (точнее, растительная клетка ). Это истощает растение свободный фосфат снабжение, которое косвенно стимулирует синтез крахмала хлоропластов.[155]Хотя это связано с низкой скоростью фотосинтеза, сами зерна крахмала не обязательно могут существенно влиять на эффективность фотосинтеза,[156] и может быть просто побочным эффектом другого фактора, подавляющего фотосинтез.[155]
Фотодыхание
Фотодыхание может произойти при слишком высокой концентрации кислорода. RuBisCO не может хорошо отличить кислород от углекислого газа, поэтому может случайно добавить O2 вместо CO2 к RuBP. Этот процесс снижает эффективность фотосинтеза - он потребляет АТФ и кислород, выделяет CO.2, и не производит сахара. Он может тратить до половины углерода, зафиксированного в цикле Кальвина.[150] Несколько механизмов развились в разных ветвях, которые повышают концентрацию углекислого газа по сравнению с кислородом в хлоропласте, повышая эффективность фотосинтеза. Эти механизмы называются механизмы концентрации углекислого газа, или СКК. К ним относятся Метаболизм крассуловой кислоты, C4 фиксация углерода,[150] и пиреноиды. Хлоропласты в C4 растения примечательны тем, что демонстрируют отчетливый диморфизм хлоропластов.
pH
Из-за ЧАС+ градиент через тилакоидную мембрану, внутренняя часть тилакоида кислый, с pH около 4,[157] в то время как строма слегка щелочная, с pH около 8.[158]Оптимальный pH стромы для цикла Кальвина составляет 8,1, при этом реакция почти останавливается, когда pH падает ниже 7,3.[159]
CO2 в воде может образоваться угольная кислота, которые могут нарушать pH изолированных хлоропластов, препятствуя фотосинтезу, даже если CO2 является использовал в фотосинтезе. Однако хлоропласты в живых клетки растений это не так сильно влияет.[158]
Хлоропласты могут перекачивать K+ и H+ ионы внутрь и наружу с помощью плохо изученной световой транспортной системы.[158]
В присутствии света pH просвета тилакоидов может упасть до 1,5 единиц pH, в то время как pH стромы может повыситься почти на одну единицу pH.[159]
Синтез аминокислот
Только хлоропласты составляют почти все клетки растений. аминокислоты в их строма[160] кроме серосодержащий такие как цистеин и метионин.[161][162] Цистеин производится в хлоропласте ( пропластид тоже), но он также синтезируется в цитозоль и митохондрии, вероятно, потому, что у него проблемы с пересечением мембран, чтобы добраться туда, где это необходимо.[162] Известно, что хлоропласт производит предшественников метионина, но неясно, выполняет ли органелла последний этап пути или это происходит в цитозоль.[163]
Другие соединения азота
Хлоропласты составляют все клетки пурины и пиримидины - азотистые основания нашел в ДНК и РНК.[160] Они также конвертируют нитрит (НЕТ2−) в аммиак (NH3) который снабжает завод азот сделать его аминокислоты и нуклеотиды.[160]
Прочие химические продукты
Эта секция нуждается в расширении с: нужно больше о липидах, а также о парамилоне. Вы можете помочь добавляя к этому. (Март 2013 г.) |
Пластида - это место разнообразных и сложных липид синтез в растениях.[164][165] Углерод, используемый для образования большей части липидов, получен из ацетил-КоА, который является продуктом декарбоксилирования пируват.[164] Пируват может проникать в пластиду из цитозоля путем пассивной диффузии через мембрану после образования в гликолиз.[166] Пируват также производится в пластиде из фосфоенолпирувата, метаболита, образующегося в цитозоле из пирувата или PGA.[164] Ацетат в цитозоле недоступен для биосинтеза липидов в пластиде.[167] Типичная длина жирных кислот, образующихся в пластиде, составляет 16 или 18 атомов углерода, с 0-3 СНГ двойные связи.[168]
Биосинтез жирных кислот из ацетил-КоА в первую очередь требует двух ферментов. Ацетил-КоА-карбоксилаза создает малонил-КоА, используемый как на первом этапе, так и на этапах расширения синтеза. Синтаза жирных кислот (FAS) представляет собой большой комплекс ферментов и кофакторов, в том числе белок-носитель ацила (ACP), который удерживает ацильную цепь в процессе ее синтеза. Инициирование синтеза начинается с конденсации малонил-АПФ с ацетил-КоА с образованием кетобутирил-АПФ. 2 скидки, связанные с использованием НАДФН и одно обезвоживание создает бутирил-АСР. Увеличение количества жирных кислот происходит в результате повторяющихся циклов конденсации, восстановления и дегидратации малонил-АСР.[164]
Другие липиды получают из Путь метил-эритритолфосфата (MEP) и состоит из гибберелины, стеролы, абсцизовая кислота, фитол, и бесчисленное множество вторичные метаболиты.[164]
Дифференциация, репликация и наследование

Хлоропласты - это особый тип органелл растительной клетки, называемый пластида, хотя эти два термина иногда используются как синонимы. Есть много других видов пластид, которые выполняют различные функции. Все хлоропласты в растении происходят от недифференцированных пропластидов, обнаруженных в зигота,[160] или оплодотворенное яйцо. Пропластиды обычно встречаются у взрослых растений. апикальные меристемы. Хлоропласты обычно не развиваются из пропластидов в меристемы кончиков корней[169]- взамен образования крахмалакопительных амилопласты встречается чаще.[160]
В стреляет, пропластиды из стрелять апикальной меристемой могут постепенно превращаться в хлоропласты в фотосинтетический ткани листа по мере созревания листа при воздействии необходимого света.[12] Этот процесс включает инвагинации внутренней пластидной мембраны, формирующие листы мембраны, которые выступают во внутреннюю оболочку. строма. Эти листы мембраны затем складываются, чтобы тилакоиды и грана.[170]
Если покрытосеменные побеги не подвергаются воздействию света, необходимого для образования хлоропластов, пропластиды могут развиться в этиопласт стадия перед превращением в хлоропласты. Этиопласт - это пластида, в которой отсутствует хлорофилл, и имеет инвагинации внутренней мембраны, которые образуют решетку трубок в их строме, называемую проламеллярное тело. Хотя этиопластам не хватает хлорофилла, они имеют желтый хлорофилл. предшественник на складе.[12] В течение нескольких минут воздействия света проламеллярное тело начинает реорганизовываться в стопки тилакоидов, и начинает вырабатываться хлорофилл. Этот процесс, при котором этиопласт становится хлоропластом, занимает несколько часов.[170] Голосеменные не требуют света для образования хлоропластов.[170]
Однако свет не гарантирует, что пропластид превратится в хлоропласт. Превратится ли пропластида в хлоропласт или другой вид пластиды, в основном контролируется ядро[12] и во многом зависит от того, в какой ячейке он находится.[160]

Пластидное взаимопревращение
Пластидная дифференциация не постоянна, фактически возможно множество взаимопревращений. Хлоропласты могут быть преобразованы в хромопласты, которые пигмент -заполненные пластиды, отвечающие за яркие цвета, видимые в цветы и спелые фрукты. Хранение крахмала амилопласты также могут быть преобразованы в хромопласты, и пропластиды могут развиваться прямо в хромопласты. Хромопласты и амилопласты также могут стать хлоропластами, как это происходит, когда морковь или картофель горит. Если растение повреждено или что-то еще заставляет растительную клетку вернуться в меристематический состояние, хлоропласты и другие пластиды могут снова превратиться в пропластиды. Хлоропласт, амилопласт, хромопласт, пропласт и т. Д. Не являются абсолютными состояниями - часто встречаются промежуточные формы.[160]
Разделение
Эта секция нуждается в расширении с: функциями, Z-образным кольцом динамической сборки, регуляторами, такими как Giant Chloroplast 1. Вы можете помочь добавляя к этому. (Февраль 2013) |
Большинство хлоропластов в фотосинтетической клетке не развиваются непосредственно из пропластидов или этиопластов. Фактически, типичный стрелять меристематически растительная клетка содержит всего 7–20 пропластиды. Эти пропластиды дифференцируются в хлоропласты, которые делятся с образованием 30–70 хлоропластов, обнаруженных в зрелой фотосинтезирующей растительной клетке. Если ячейка разделяет деление хлоропластов обеспечивает дополнительные хлоропласты для разделения между двумя дочерними клетками.[171]
В одноклеточных водоросли, деление хлоропластов - единственный способ образования новых хлоропластов. Дифференциации пропластид не происходит - когда клетка водоросли делится, ее хлоропласт делится вместе с ней, и каждый дочерняя ячейка получает зрелый хлоропласт.[170]
В клетке делятся почти все хлоропласты, а не небольшая группа быстро делящихся хлоропластов.[172] Хлоропласты не имеют определенного S-фаза - их репликация ДНК не синхронизирована и не ограничена репликацией их клеток-хозяев.[173]Многое из того, что мы знаем о делении хлоропластов, получено в результате изучения таких организмов, как Арабидопсис и красная водоросль Cyanidioschyzon merolæ.[143]
![Большинство хлоропластов в растительных клетках и все хлоропласты в водорослях возникают в результате деления хлоропластов. [170] Ссылки на изображения, [143] [174]](http://upload.wikimedia.org/wikipedia/commons/thumb/d/df/Chloroplast_division.svg/800px-Chloroplast_division.svg.png)
Процесс деления начинается, когда белки FtsZ1 и FtsZ2 собрать в нити и с помощью протеина ARC6, образуют структуру, называемую Z-кольцом, в строме хлоропласта.[143][174] В Мин система управляет размещением Z-образного кольца, обеспечивая более или менее равномерное расщепление хлоропласта. Протеин Разум предотвращает соединение FtsZ и образование волокон. Другой белок ARC3 также могут быть задействованы, но это не очень хорошо понимается. Эти белки активны на полюсах хлоропласта, предотвращая образование там Z-кольца, но вблизи центра хлоропласта, Мой подавляет их, позволяя формировать Z-образное кольцо.[143]
Далее два пластид-делительные кольца, или образуются кольца ПД. Внутреннее кольцо, разделяющее пластиды, расположено на внутренней стороне внутренней мембраны хлоропласта и образуется первым.[143] Наружное кольцо, разделяющее пластиды, обернуто вокруг внешней мембраны хлоропласта. Он состоит из нитей диаметром около 5 нанометров,[143] расположены рядами на расстоянии 6,4 нанометра друг от друга и сжимаются, чтобы сжать хлоропласт. Именно тогда начинается сужение хлоропластов.[174]
У некоторых видов, таких как Cyanidioschyzon merolæ, хлоропласты имеют третье кольцо, разделяющее пластиды, расположенное в межмембранном пространстве хлоропласта.[143][174]
В конце фазы сжатия, динамин белки собираются вокруг внешнего пластид-делящего кольца,[174] помогая придать силу сдавливанию хлоропласта.[143] Между тем Z-образное кольцо и внутреннее кольцо, разделяющее пластиды, ломаются.[174] На этом этапе множество плазмид ДНК хлоропластов, плавающих в строме, разделяются и распределяются между двумя образующимися дочерними хлоропластами.[175]
Позже динамины мигрируют под внешнее разделяющее кольцо пластид в прямой контакт с внешней мембраной хлоропласта,[174] расщепить хлоропласт в двух дочерних хлоропластах.[143]
Остаток внешнего пластидного разделяющего кольца остается плавающим между двумя дочерними хлоропластами, а остаток динаминового кольца остается прикрепленным к одному из дочерних хлоропластов.[174]
Из пяти или шести колец, участвующих в делении хлоропластов, только внешнее разделяющее пластиды кольцо присутствует на протяжении всей фазы сжатия и деления - тогда как Z-кольцо образуется первым, сжатие не начинается до тех пор, пока не сформируется внешнее разделяющее пластиды кольцо.[174]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Регулирование
У видов водоросли которые содержат один хлоропласт, регуляция деления хлоропластов чрезвычайно важна для обеспечения того, чтобы каждая дочерняя клетка получила хлоропласт - хлоропласты не могут быть созданы с нуля.[77][143] У таких организмов, как растения, клетки которых содержат несколько хлоропластов, координация более слабая и менее важна. Вероятно, что деление хлоропластов и клеток в какой-то мере синхронизировано, хотя механизмы этого в основном неизвестны.[143]
Было показано, что свет необходим для деления хлоропластов. Хлоропласты могут расти и проходить через некоторые стадии сжатия под некачественный зеленый свет, но медленно завершают деление - им требуется яркий белый свет для полного деления. Было обнаружено, что листья шпината, выращенные при зеленом освещении, содержат много крупных хлоропластов в форме гантелей. Воздействие белого света может стимулировать эти хлоропласты к делению и уменьшать популяцию хлоропластов в форме гантелей.[172][175]
Наследование хлоропластов
Нравиться митохондрии, хлоропласты обычно наследуются от одного родителя. Наследование двух родительских хлоропластов, при котором пластидные гены наследуются от обоих родительских растений, у некоторых цветковых растений встречается в очень низких количествах.[176]
Многие механизмы предотвращают наследование ДНК хлоропластов от двух родителей, включая избирательное разрушение хлоропластов или их генов внутри гамета или же зигота, а хлоропласты одного родителя исключаются из эмбриона. Родительские хлоропласты можно отсортировать так, чтобы в каждом потомстве присутствовал только один тип.[177]
Голосеменные, Такие как сосны, в основном передают хлоропласты по отцовской линии,[178] пока цветущие растения часто наследуют хлоропласты по материнской линии.[179][180] Когда-то считалось, что цветковые растения наследуют хлоропласты только по материнской линии. Однако в настоящее время имеется много задокументированных случаев покрытосеменные наследующие хлоропласты по отцовской линии.[176]
Покрытосеменные, которые передают хлоропласты по материнской линии, есть много способов предотвратить отцовское наследование. Большинство из них производят сперматозоиды не содержащие пластид. Существует множество других задокументированных механизмов, предотвращающих отцовское наследование у этих цветковых растений, например, различная скорость репликации хлоропластов внутри эмбриона.[176]
Среди покрытосеменных чаще наблюдается наследование отцовских хлоропластов. гибриды чем в потомстве от родителей того же вида. Это предполагает, что несовместимые гибридные гены могут вмешиваться в механизмы, предотвращающие отцовское наследование.[176]
Транспластомные растения
В последнее время хлоропласты привлекли внимание разработчиков генетически модифицированные культуры. Поскольку у большинства цветковых растений хлоропласты не наследуются от мужского родителя, трансгены в этих пластидах не могут распространяться пыльца. Это делает пластидная трансформация ценный инструмент для создания и выращивания генетически модифицированных растений, содержащихся в биологической среде, что значительно снижает экологические риски. Этот биологическое сдерживание Таким образом, стратегия подходит для создания сосуществование традиционного и органического сельского хозяйства. Хотя надежность этого механизма еще не была изучена для всех соответствующих видов сельскохозяйственных культур, недавние результаты по табачным растениям являются многообещающими, показывая, что степень неудачной локализации транспластомных растений составляет 3 из 1000000.[180]
Рекомендации
- ^ Джонс Д. (2003) [1917]. Роуч П., Хартманн Дж., Сеттер Дж. (Ред.). Словарь английского произношения. Кембридж: Издательство Кембриджского университета. ISBN 3-12-539683-2.
- ^ «Хлоропласт». Словарь Merriam-Webster.
- ^ Основы биологии (18 марта 2016 г.). «Бактерии».
- ^ «хлоропласт». Интернет-словарь этимологии.
- ^ фон Моль, Х. (1835/1837). Ueber die Vermehrung der Pflanzen-Zellen durch Teilung. Диссертация. Тюбинген 1835 г. Флора 1837, .
- ^ а б Шимпер, АФ (1883). "Über die Entwicklung der Chlorophyllkörner und Farbkörper" [О развитии зерен и пятен хлорофилла]. Бот. Zeitung (на немецком). 41: 105–14, 121–31, 137–46, 153–62. Архивировано из оригинал 19 октября 2013 г.
- ^ Страсбургер Э (1884). Das botanische Praktikum (1-е изд.). Йена: Густав Фишер.
- ^ Ганнинг Б., Кениг Ф., Говинджи П. (2006). «Посвящение пионерам исследований структуры хлоропластов». В Wise RR, Hoober JK (ред.). Строение и функции пластид. Нидерланды: Спрингер. стр. xxiii – xxxi. ISBN 9781402065705.
- ^ Хубер Дж. К. (1984). Хлоропласты. Нью-Йорк: Пленум. ISBN 9781461327677.
- ^ а б c d е ж грамм час я j Кэмпбелл Н.А., Рис Дж. Б., Урри Л. А., Кейн М. Л., Вассерман, Минорский П. В., Джексон Р. Б. (2009). Биология (8-е изд.). Бенджамин Каммингс (Пирсон). п. 516. ISBN 978-0-8053-6844-4.
- ^ Мерещковский С (1905). "Über Natur und Ursprung der Chromatophoren im Pflanzenreiche" [О природе и происхождении хроматофоров в растительном мире]. Биол Централбл (на немецком). 25: 593–604.
- ^ а б c d Альбертс Б. (2002). Молекулярная биология клетки (4-е изд.). Нью-Йорк [u.a.]: Гарланд. ISBN 0-8153-4072-9.
- ^ а б Мур KR, Magnabosco C, Momper L, Gold DA, Bosak T, Fournier GP (2019). «Расширенная рибосомная филогения цианобактерий поддерживает глубокое размещение пластидов». Границы микробиологии. 10: 1612. Дои:10.3389 / fmicb.2019.01612. ЧВК 6640209. PMID 31354692.
- ^ а б Кумар К., Мелла-Эррера Р.А., Golden JW (апрель 2010 г.). «Цианобактериальные гетероцисты». Перспективы Колд-Спринг-Харбор в биологии. 2 (4): а000315. Дои:10.1101 / cshperspect.a000315. ЧВК 2845205. PMID 20452939.
- ^ а б c d е Кэмпбелл Н.А., Рис Дж. Б., Урри Л. А., Кейн М. Л., Вассерман, Минорский П. В., Джексон Р. Б. (2009). Биология (8-е изд.). Бенджамин Каммингс (Пирсон). С. 186–187. ISBN 978-0-8053-6844-4.
- ^ а б c d е ж грамм час я j k л м п о п q р s т ты v ш Икс у z аа ab ac объявление ае аф аг ах ай эй ак аль являюсь ан ао ap водный ар в качестве в au средний ау Ким Э, Арчибальд Дж. М. (2009). «Разнообразие и эволюция пластидов и их геномов». В Sandelius AS, Aronsson H (ред.). Хлоропласт. Монографии по растительной клетке. 13. С. 1–39. Дои:10.1007/978-3-540-68696-5_1. ISBN 978-3-540-68692-7.
- ^ Bryant DA, Guglielmi G, de Marsac NT, Castets AM, Cohen-Bazire G (1979). «Структура фикобилисом цианобактерий: модель». Архив микробиологии. 123 (2): 311–34. Дои:10.1007 / BF00446810. S2CID 1589428.
- ^ а б c d е ж грамм час я j k л м п о п q р s т ты v ш Икс у z аа ab ac объявление ае аф Килинг П.Дж. (октябрь 2004 г.). «Разнообразие и эволюционная история пластид и их хозяев». Американский журнал ботаники. 91 (10): 1481–93. Дои:10.3732 / ajb.91.10.1481. PMID 21652304. S2CID 17522125.
- ^ а б c d Макфадден Г.И., ван Дурен Г.Г. (июль 2004 г.). «Эволюция: геном красных водорослей подтверждает общее происхождение всех пластид». Текущая биология. 14 (13): R514-6. Дои:10.1016 / j.cub.2004.06.041. PMID 15242632. S2CID 18131616.
- ^ а б c d е Майло Р., Филлипс Р. "Клеточная биология в цифрах: насколько велики хлоропласты?". book.bionumbers.org. Получено 7 февраля 2017.
- ^ а б c d Санчес-Баракальдо П., Рэйвен Дж. А., Пизани Д., Нолл А. Х. (сентябрь 2017 г.). «Ранние фотосинтезирующие эукариоты населяли среды обитания с низкой соленостью». Труды Национальной академии наук Соединенных Штатов Америки. 114 (37): E7737 – E7745. Дои:10.1073 / pnas.1620089114. ЧВК 5603991. PMID 28808007.
- ^ Fuks B, Homblé F (октябрь 1996 г.). «Механизм проникновения протонов через липидные мембраны хлоропластов». Физиология растений. 112 (2): 759–66. Дои:10.1104 / стр.112.2.759. ЧВК 158000. PMID 8883387.
- ^ Джойард Дж., Блок М.А., Дус Р. (август 1991 г.). «Молекулярные аспекты биохимии пластидных оболочек». Европейский журнал биохимии. 199 (3): 489–509. Дои:10.1111 / j.1432-1033.1991.tb16148.x. PMID 1868841.
- ^ «Хлоропласт». Энциклопедия науки. Получено 27 декабря 2012.
- ^ а б c d е ж Накаяма Т., Арчибальд Дж. М. (апрель 2012 г.). «Развитие фотосинтетической органеллы». BMC Биология. 10 (1): 35. Дои:10.1186/1741-7007-10-35. ЧВК 3337241. PMID 22531210.
- ^ а б c d е Макфадден Г.И. (январь 2001 г.). «Происхождение и интеграция хлоропластов». Физиология растений. 125 (1): 50–3. Дои:10.1104 / стр.125.1.50. ЧВК 1539323. PMID 11154294.
- ^ Арчибальд Дж. М. (январь 2009 г.). «Загадка эволюции пластид». Текущая биология. 19 (2): R81-8. Дои:10.1016 / j.cub.2008.11.067. PMID 19174147. S2CID 51989.
- ^ Понсе-Толедо Р.И., Дешам П., Лопес-Гарсия П., Живанович Ю., Бензерара К., Морейра Д. (февраль 2017 г.). «Пресноводная цианобактерия с ранним ветвлением и у истоков пластид». Текущая биология. 27 (3): 386–391. Дои:10.1016 / j.cub.2016.11.056. ЧВК 5650054. PMID 28132810.
- ^ де Фриз Дж, Арчибальд Дж. М. (февраль 2017 г.). «Эндосимбиоз: развились ли пластиды из пресноводных цианобактерий?». Текущая биология. 27 (3): R103 – R105. Дои:10.1016 / j.cub.2016.12.006. PMID 28171752.
- ^ а б Лопес-Гарсия П., Эме Л., Морейра Д. (декабрь 2017 г.). «Симбиоз в эволюции эукариот». Журнал теоретической биологии. 434: 20–33. Дои:10.1016 / j.jtbi.2017.02.031. ЧВК 5638015. PMID 28254477.
- ^ Делай Л., Валадес-Кано С., Перес-Заморано Б. (март 2016 г.). "Насколько древняя Paulinella Chromatophora?". PLOS токи. 8. Дои:10.1371 / currents.tol.e68a099364bb1a1e129a17b4e06b0c6b. ЧВК 4866557. PMID 28515968.
- ^ а б c d е ж грамм час я j k л м п о п q р s т ты v ш Мудрый Р. Р., Хубер Дж. К. (2006). Строение и функции пластид. Дордрехт: Спрингер. С. 3–21. ISBN 978-1-4020-4061-0.
- ^ Болл С., Коллеони С., Ченчи Ю., Радж Дж. Н., Тиртье С. (март 2011 г.). «Эволюция метаболизма гликогена и крахмала у эукариот дает молекулярные ключи к пониманию возникновения пластидного эндосимбиоза». Журнал экспериментальной ботаники. 62 (6): 1775–801. Дои:10.1093 / jxb / erq411. PMID 21220783.
- ^ а б c d е ж грамм час я j k л м п о п q р s т ты v ш Икс у z аа ab ac объявление ае аф Килинг П.Дж. (март 2010 г.). «Эндосимбиотическое происхождение, разнообразие и судьба пластид». Философские труды Лондонского королевского общества. Серия B, Биологические науки. 365 (1541): 729–48. Дои:10.1098 / rstb.2009.0103. ЧВК 2817223. PMID 20124341.
- ^ а б c d е ж грамм Кэмпбелл Н.А., Рис Дж. Б., Урри Л. А., Кейн М. Л., Вассерман, Минорский П. В., Джексон Р. Б. (2009). Биология (8-е изд.). Бенджамин Каммингс (Пирсон). С. 582–92. ISBN 978-0-8053-6844-4.
- ^ "родо-". Бесплатный словарь. Фарлекс. Получено 7 июн 2013.
- ^ а б Льюис Л.А., Маккорт Р.М. (октябрь 2004 г.). «Зеленые водоросли и происхождение наземных растений». Американский журнал ботаники. 91 (10): 1535–56. Дои:10.3732 / ajb.91.10.1535. PMID 21652308.
- ^ Мачида М., Такечи К., Сато Х., Чунг С.Дж., Куроива Х., Такио С. и др. (Апрель 2006 г.). «Гены пути синтеза пептидогликана необходимы для деления хлоропластов у мха». Труды Национальной академии наук Соединенных Штатов Америки. 103 (17): 6753–8. Bibcode:2006ПНАС..103.6753М. Дои:10.1073 / pnas.0510693103. ЧВК 1458953. PMID 16618924.
- ^ Moroney JV, Somanchi A (январь 1999 г.). «Как водоросли концентрируют CO2, чтобы повысить эффективность фотосинтетической фиксации углерода?». Физиология растений. 119 (1): 9–16. Дои:10.1104 / стр.119.1.9. ЧВК 1539202. PMID 9880340.
- ^ Tartar A, Boucias DG (апрель 2004 г.). «Не фотосинтетическая патогенная зеленая водоросль Helicosporidium sp. Сохранила модифицированный функциональный пластидный геном». Письма о микробиологии FEMS. 233 (1): 153–7. Дои:10.1016 / j.femsle.2004.02.006. PMID 15043882.
- ^ а б c d Nowack EC, Vogel H, Groth M, Grossman AR, Melkonian M, Glöckner G (январь 2011 г.). «Перенос эндосимбиотических генов и регуляция транскрипции перенесенных генов в Paulinella chromatophora». Молекулярная биология и эволюция. 28 (1): 407–22. Дои:10.1093 / molbev / msq209. PMID 20702568.
- ^ а б c d е Данн Л. (2002). Биологические науки - объяснение (PDF). Зеленая ДНК: ОБЪЯСНЕНИЕ БИОЛОГИИ.
- ^ а б c d е Clegg MT, Gaut BS, Learn GH, Morton BR (июль 1994). «Скорость и закономерности эволюции ДНК хлоропластов». Труды Национальной академии наук Соединенных Штатов Америки. 91 (15): 6795–801. Bibcode:1994PNAS ... 91.6795C. Дои:10.1073 / пнас.91.15.6795. ЧВК 44285. PMID 8041699.
- ^ а б c d е ж Шоу Дж., Лики Э. Б., Шиллинг Э. Э., Small RL (март 2007 г.). «Сравнение последовательностей всего хлоропластного генома для выбора некодирующих областей для филогенетических исследований покрытосеменных: черепаха и заяц III». Американский журнал ботаники. 94 (3): 275–88. Дои:10.3732 / ajb.94.3.275. PMID 21636401. S2CID 30501148.
- ^ а б Чаал Б.К., Грин Б.Р. (февраль 2005 г.). «Пути импорта белка в« сложных »хлоропластах, происходящих от вторичного эндосимбиоза с участием предка красных водорослей». Молекулярная биология растений. 57 (3): 333–42. Дои:10.1007 / s11103-004-7848-у. PMID 15830125. S2CID 22619029.
- ^ а б Мустафа А., Бестери Б., Майер У. Г., Боулер С., Валентин К., Бхаттачарья Д. (июнь 2009 г.). «Геномные следы скрытого пластидного эндосимбиоза у диатомовых водорослей» (PDF). Наука. 324 (5935): 1724–6. Bibcode:2009Научный ... 324.1724M. Дои:10.1126 / science.1172983. PMID 19556510. S2CID 11408339.
- ^ Роджерс МБ, Гилсон П.Р., Су В., Макфадден Г.И., Килинг П.Дж. (январь 2007 г.). «Полный хлоропластный геном хлорарахниофита Bigelowiella natans: свидетельство независимого происхождения хлорарахниофитов и вторичных эндосимбионтов эвгленид». Молекулярная биология и эволюция. 24 (1): 54–62. Дои:10.1093 / molbev / msl129. PMID 16990439.
- ^ а б c d е ж грамм час я j k л м п Хакетт Дж. Д., Андерсон Д. М., Эрднер Д. Л., Бхаттачарья Д. (октябрь 2004 г.). «Динофлагелляты: замечательный эволюционный эксперимент». Американский журнал ботаники. 91 (10): 1523–34. Дои:10.3732 / ajb.91.10.1523. PMID 21652307.
- ^ Толедо, Род-Айленд (5 марта 2018 г.). Происхождение и ранняя эволюция фотосинтезирующих эукариот (Тезис). Université Paris-Saclay.
- ^ Bodył A (февраль 2018 г.). «Некоторые пластиды, полученные из красных водорослей, эволюционировали посредством клептопластии? Гипотеза». Биологические обзоры Кембриджского философского общества. 93 (1): 201–222. Дои:10.1111 / brv.12340. PMID 28544184. S2CID 24613863.
- ^ а б Яноушковец Дж., Гавелис Г.С., Бурки Ф., Динь Д., Бахварофф Т.Р., Горник С.Г. и др. (Январь 2017 г.). «Основные переходы в эволюции динофлагеллят, выявленные филотранскриптомикой». Труды Национальной академии наук Соединенных Штатов Америки. 114 (2): E171 – E180. Дои:10.1073 / pnas.1614842114. ЧВК 5240707. PMID 28028238.
- ^ а б c d Наир СК, Стрипен Б. (август 2011 г.). «Что вообще делают человеческие паразиты с хлоропластом?». PLOS Биология. 9 (8): e1001137. Дои:10.1371 / journal.pbio.1001137. ЧВК 3166169. PMID 21912515.
- ^ Куигг А., Котабова Э., Ярешова Дж., Кана Р., Сетлик Дж., Седива Б. и др. (10 октября 2012 г.). «Фотосинтез в Chromera velia представляет собой простую систему с высокой эффективностью». PLOS ONE. 7 (10): e47036. Bibcode:2012PLoSO ... 747036Q. Дои:10.1371 / journal.pone.0047036. ЧВК 3468483. PMID 23071705.
- ^ Доррелл Р.Г., Смит А.Г. (июль 2011 г.). «Делают ли красный и зеленый коричневый цвет ?: перспективы накопления пластид в хромальвеолатах». Эукариотическая клетка. 10 (7): 856–68. Дои:10.1128 / EC.00326-10. ЧВК 3147421. PMID 21622904.
- ^ Мисон Б.В., Чанг С.С., Суини Б.М. (1982). "Характеристика перидин-хлорофилловых α-белков морской динофлагелляты Ceratium furca". Ботаника Марина. 25 (8): 347–50. Дои:10.1515 / botm.1982.25.8.347. S2CID 83867103.
- ^ а б c Тенгс Т., Дальберг О.Дж., Шалчиан-Тебризи К., Klaveness D, Руди К., Делвиче К.Ф., Якобсен К.С. (май 2000 г.). «Филогенетический анализ показывает, что содержащие 19'-гексаноилокси-фукоксантин динофлагелляты имеют третичные пластиды гаптофитного происхождения». Молекулярная биология и эволюция. 17 (5): 718–29. Дои:10.1093 / oxfordjournals.molbev.a026350. PMID 10779532.
- ^ а б c Schnepf E, Elbrächter M (1999). «Хлоропласты динофитов и филогения - обзор». Grana. 38 (2–3): 81–97. Дои:10.1080/00173139908559217.
- ^ Сковгаард А (1998). «Роль удержания хлоропластов в морской динофлагелляте». Экология водных микробов. 15: 293–301. Дои:10.3354 / ame015293.
- ^ Доррелл Р.Г., Хоу Си-Джей (август 2015 г.). «Интеграция пластид с их хозяевами: уроки, извлеченные из динофлагеллят». Труды Национальной академии наук Соединенных Штатов Америки. 112 (33): 10247–54. Bibcode:2015ПНАС..11210247D. Дои:10.1073 / pnas.1421380112. ЧВК 4547248. PMID 25995366.
- ^ Хоган CM (2010). "Дезоксирибонуклеиновая кислота". В Draggan S, Cleveland C (ред.). Энциклопедия Земли. Вашингтон, округ Колумбия: Национальный совет по науке и окружающей среде.
- ^ «ктДНК - хлоропластная ДНК». AllAcronyms.com. Архивировано из оригинал 3 июня 2013 г.. Получено 2 января 2013.
- ^ Оксфордский словарь сокращений. ctDNA - словарное определение. 1998 г.
- ^ «Хлоропласты и другие пластиды». Гамбургский университет. Архивировано из оригинал 25 сентября 2012 г.. Получено 27 декабря 2012.
- ^ а б c d е ж грамм Санделиус А.С. (2009). Хлоропласт: взаимодействие с окружающей средой. Springer. п. 18. ISBN 978-3-540-68696-5.
- ^ а б Берджесс Дж. (1989). Введение в развитие растительных клеток. Кембридж: издательство Кембриджского университета. п. 62. ISBN 0-521-31611-1.
- ^ а б c d е ж грамм Бендич А.Дж. (июль 2004 г.). «Круглые хромосомы хлоропластов: великая иллюзия». Растительная клетка. 16 (7): 1661–6. Дои:10.1105 / tpc.160771. ЧВК 514151. PMID 15235123.
- ^ а б c Колоднер Р., Тевари К.К. (январь 1979 г.). «Инвертированные повторы в ДНК хлоропластов высших растений». Труды Национальной академии наук Соединенных Штатов Америки. 76 (1): 41–5. Bibcode:1979ПНАС ... 76 ... 41К. Дои:10.1073 / pnas.76.1.41. ЧВК 382872. PMID 16592612.
- ^ а б Палмер Дж. Д., Томпсон В. Ф. (июнь 1982 г.). «Перестройки ДНК хлоропластов происходят чаще, когда теряется большая инвертированная повторяющаяся последовательность». Клетка. 29 (2): 537–50. Дои:10.1016/0092-8674(82)90170-2. PMID 6288261. S2CID 11571695.
- ^ Биохимия растений (3-е изд.). Академическая пресса. 2005. с.517. ISBN 978-0-12-088391-2.
количество копий ктДНК на хлоропласт.
- ^ а б Кобаяси Т., Такахара М., Миягишима С.Ю., Куроива Х., Сасаки Н., Охта Н. и др. (Июль 2002 г.). «Обнаружение и локализация HU-подобного белка, кодируемого хлоропластом, который организует нуклеоиды хлоропласта». Растительная клетка. 14 (7): 1579–89. Дои:10.1105 / tpc.002717. ЧВК 150708. PMID 12119376.
- ^ Одахара М., Кишита Ю., Секин Ю. (август 2017 г.). «MSH1 поддерживает стабильность генома органелл и генетически взаимодействует с RECA и RECG в мхе Physcomitrella patens». Журнал растений. 91 (3): 455–465. Дои:10.1111 / tpj.13573. PMID 28407383.
- ^ Роуэн Б.А., Ольденбургский ди-джей, Бендич А.Дж. (июнь 2010 г.). «RecA поддерживает целостность молекул хлоропластной ДНК у Arabidopsis». Журнал экспериментальной ботаники. 61 (10): 2575–88. Дои:10.1093 / jxb / erq088. ЧВК 2882256. PMID 20406785.
- ^ а б c d е ж Кришнан Н.М., Рао Б.Дж. (май 2009 г.). «Сравнительный подход к выяснению репликации генома хлоропластов». BMC Genomics. 10 (237): 237. Дои:10.1186/1471-2164-10-237. ЧВК 2695485. PMID 19457260.
- ^ Хайнхорст С., Cannon GC (1993). «Репликация ДНК в хлоропластах». Журнал клеточной науки. 104: 1–9.
- ^ «Влияние химических мутагенов на нуклеотидную последовательность». Биоциклопедия. Получено 24 октября 2015.
- ^ Бернштейн Х, Бернштейн С (июль 1973 г.). «Круглые и разветвленные кольцевые конкатенации в качестве возможных промежуточных продуктов в репликации ДНК бактериофага Т4». Журнал молекулярной биологии. 77 (3): 355–61. Дои:10.1016/0022-2836(73)90443-9. PMID 4580243.
- ^ а б Альбертс Б. (2002). Молекулярная биология клетки (4-е изд.). Нью-Йорк [u.a.]: Гарланд. ISBN 0-8153-4072-9.
- ^ Хуанг CY, Ayliffe MA, Timmis JN (март 2003 г.). «Прямое измерение скорости переноса ДНК хлоропластов в ядро». Природа. 422 (6927): 72–6. Bibcode:2003Натура 422 ... 72ч. Дои:10.1038 / природа01435. PMID 12594458. S2CID 4319507.
- ^ а б c Мартин В., Ружан Т., Ричли Э., Хансен А., Корнелсен С., Линс Т. и др. (Сентябрь 2002 г.). «Эволюционный анализ геномов арабидопсиса, цианобактерий и хлоропластов выявил филогению пластид и тысячи генов цианобактерий в ядре». Труды Национальной академии наук Соединенных Штатов Америки. 99 (19): 12246–51. Bibcode:2002ПНАС ... 9912246М. Дои:10.1073 / pnas.182432999. ЧВК 129430. PMID 12218172.
- ^ Смит Д.Р., Ли Р.В. (апрель 2014 г.). «Пластида без генома: данные нефотосинтетических зеленых водорослей рода Polytomella». Физиология растений. 164 (4): 1812–9. Дои:10.1104 / стр.113.233718. ЧВК 3982744. PMID 24563281.
- ^ Арчибальд Дж. М. (декабрь 2006 г.). «Геномика водорослей: изучение отпечатка эндосимбиоза». Текущая биология. 16 (24): R1033-5. Дои:10.1016 / j.cub.2006.11.008. PMID 17174910.
- ^ Кусевицкий С., Нотт А., Моклер Т.К., Хонг Ф., Сашетто-Мартинс Г., Сурпин М. и др. (Май 2007 г.). «Сигналы от хлоропластов сходятся, чтобы регулировать экспрессию ядерных генов». Наука. 316 (5825): 715–9. Bibcode:2007Наука ... 316..715K. Дои:10.1126 / наука.1140516 (неактивно 6 ноября 2020 г.). PMID 17395793. Сложить резюме – Ученый (31 марта 2007 г.).CS1 maint: DOI неактивен по состоянию на ноябрь 2020 г. (связь)
- ^ Хедтке Б., Бёрнер Т., Вейхе А. (август 1997 г.). «Митохондриальные и хлоропластные РНК-полимеразы фагового типа у Arabidopsis». Наука. 277 (5327): 809–11. Дои:10.1126 / science.277.5327.809. PMID 9242608.
- ^ Харрис Э. Х., Бойнтон Дж. Э., Гиллхэм Н. В. (декабрь 1994 г.). «Рибосомы хлоропластов и синтез белка». Микробиологические обзоры. 58 (4): 700–54. Дои:10.1128 / MMBR.58.4.700-754.1994. ЧВК 372988. PMID 7854253.
- ^ а б c d е ж грамм час я j Soll J, Schleiff E (март 2004 г.). «Импорт белка в хлоропласты». Обзоры природы Молекулярная клеточная биология. 5 (3): 198–208. Дои:10.1038 / nrm1333. PMID 14991000. S2CID 32453554.
- ^ Кэмпбелл Н.А., Рис Дж. Б., Урри Л. А., Кейн М. Л., Вассерман, Минорский П. В., Джексон Р. Б. (2009). Биология (8-е изд.). Бенджамин Каммингс (Пирсон). п. 340. ISBN 978-0-8053-6844-4.
- ^ Lung SC, Chuong SD (апрель 2012 г.). «Транзитный пептидоподобный сигнал сортировки на С-конце направляет препротеиновый рецептор Toc159 Bienertia sinuspersici к внешней мембране хлоропласта». Растительная клетка. 24 (4): 1560–78. Дои:10.1105 / tpc.112.096248. ЧВК 3398564. PMID 22517318.
- ^ Waegemann K, Soll J (март 1996). «Фосфорилирование транзитной последовательности белков-предшественников хлоропластов». Журнал биологической химии. 271 (11): 6545–54. Дои:10.1074 / jbc.271.11.6545. PMID 8626459.
- ^ а б Май Т., Солл Дж. (Январь 2000 г.). «Белки 14-3-3 образуют направляющий комплекс с белками-предшественниками хлоропластов в растениях». Растительная клетка. 12 (1): 53–64. Дои:10.1105 / tpc.12.1.53. ЧВК 140214. PMID 10634907.
- ^ Джарвис П., Солл Дж. (Декабрь 2001 г.). «Импорт белков Toc, Tic и хлоропластов». Biochimica et Biophysica Acta (BBA) - Исследование молекулярных клеток. 1541 (1–2): 64–79. Дои:10.1016 / S0167-4889 (01) 00147-1. PMID 11750663.
- ^ Мудрый Р. Р., Хубер Дж. К. (2007). Строение и функции пластидов. Springer. С. 32–33. ISBN 978-1-4020-6570-5.
- ^ "Oedogonium Link ex Hirn, 1900: 17". водоросли. Получено 19 мая 2013.
- ^ "Хламидомонада Эренберга, 1833: 288". водоросли. Получено 19 мая 2013.
- ^ "Spirogyra Link, 1820: 5". водоросли. Получено 19 мая 2013.
- ^ "Sirogonium Kützing, 1843: 278". водоросли. Получено 19 мая 2013.
- ^ "Зигнема Агард, 1817: xxxii, 98". водоросли. Получено 19 мая 2013.
- ^ "Micrasterias C.Agardh ex Ralfs, 1848: 68". водоросли. Получено 19 мая 2013.
- ^ Джон Д.М., Брук А.Дж., Уиттон Б.А. (2002). Пресноводная водорослевая флора Британских островов: руководство по идентификации пресноводных и наземных водорослей. Кембридж: Издательство Кембриджского университета. п. 335. ISBN 978-0-521-77051-4.
- ^ а б c d е ж грамм Кэмпбелл Н.А., Рис Дж. Б., Урри Л. А., Кейн М. Л., Вассерман, Минорский П. В., Джексон Р. Б. (2009). Биология (8-е изд.). Бенджамин Каммингс (Пирсон). С. 196–197. ISBN 978-0-8053-6844-4.
- ^ Койке Х., Йошио М, Кашино Й., Сато К. (май 1998 г.). «Полипептидный состав оболочек хлоропластов шпината: два основных белка занимают 90% мембран внешней оболочки». Физиология растений и клеток. 39 (5): 526–32. Дои:10.1093 / oxfordjournals.pcp.a029400. PMID 9664716.
- ^ Köhler RH, Hanson MR (январь 2000 г.). «Пластидные канальцы высших растений тканеспецифичны и регулируются в процессе развития». Журнал клеточной науки. 113 (Пт 1): 81–9. PMID 10591627. В архиве из оригинала от 20 сентября 2016 г.
- ^ Грей Дж. К., Салливан Дж. А., Хибберд Дж. М., Хансен М. Р. (2001). «Стромулы: подвижные выступы и связи между пластидами». Биология растений. 3 (3): 223–33. Дои:10.1055 / с-2001-15204.
- ^ Schattat M, Barton K, Baudisch B, Klösgen RB, Mathur J (апрель 2011 г.). «Ветвление пластидных стромул совпадает с динамикой смежного эндоплазматического ретикулума». Физиология растений. 155 (4): 1667–77. Дои:10.1104 / стр.110.170480. ЧВК 3091094. PMID 21273446.
- ^ Schattat MH, Griffiths S, Mathur N, Barton K, Wozny MR, Dunn N и др. (Апрель 2012 г.). «Дифференциальная окраска показывает, что пластиды не образуют сетей для обмена макромолекул». Растительная клетка. 24 (4): 1465–77. Дои:10.1105 / tpc.111.095398. ЧВК 3398557. PMID 22474180.
- ^ Brunkard JO, Runkel AM, Zambryski PC (август 2015 г.). «Хлоропласты расширяют стромулы независимо и в ответ на внутренние окислительно-восстановительные сигналы». Труды Национальной академии наук Соединенных Штатов Америки. 112 (32): 10044–9. Bibcode:2015ПНАС..11210044Б. Дои:10.1073 / pnas.1511570112. ЧВК 4538653. PMID 26150490.
- ^ а б c d е ж грамм час я j Берджесс Дж. (1989). Введение в развитие растительных клеток (Пбк. Ред.). Кембридж: издательство Кембриджского университета. п. 46. ISBN 0-521-31611-1.
- ^ а б Уотли Дж. М. (5 июля 1994 г.). «Возникновение периферической ретикулума в пластидах голосеменных Welwitschia mirabilis». Новый Фитолог. 74 (2): 215–220. Дои:10.1111 / j.1469-8137.1975.tb02608.x.
- ^ а б c Мудрый RR (2007). Строение и функции пластидов. Springer. С. 17–18. ISBN 978-1-4020-6570-5.
- ^ Мануэль А.Л., Киспе Дж., Мэйфилд С.П. (август 2007 г.). «Структура рибосомы хлоропласта: новые домены для регуляции трансляции». PLOS Биология. 5 (8): e209. Дои:10.1371 / journal.pbio.0050209. ЧВК 1939882. PMID 17683199.
- ^ а б Лим К., Кобаяси И., Накай К. (июль 2014 г.). «Изменения во взаимодействии рРНК-мРНК в процессе эволюции пластид». Молекулярная биология и эволюция. 31 (7): 1728–40. Дои:10.1093 / молбев / мсу120. PMID 24710516.
- ^ Хиросе Т., Сугиура М. (январь 2004 г.). «Функциональные последовательности, подобные Шайну-Дальгарно для инициации трансляции мРНК хлоропластов». Физиология растений и клеток. 45 (1): 114–7. Дои:10.1093 / pcp / pch002. PMID 14749493. S2CID 10774032.
- ^ Ма Дж., Кэмпбелл А., Карлин С. (октябрь 2002 г.). «Корреляции между последовательностями Шайна-Дальгарно и особенностями генов, такими как предсказанные уровни экспрессии и структуры оперонов». Журнал бактериологии. 184 (20): 5733–45. Дои:10.1128 / JB.184.20.5733-5745.2002. ЧВК 139613. PMID 12270832.
- ^ Лим К., Фурута Ю., Кобаяши И. (октябрь 2012 г.). «Большие вариации в генах бактериальной рибосомной РНК». Молекулярная биология и эволюция. 29 (10): 2937–48. Дои:10.1093 / молбев / mss101. ЧВК 3457768. PMID 22446745.
- ^ а б c d е ж грамм час я j Остин JR, Frost E, Vidi PA, Kessler F, Staehelin LA (июль 2006 г.). «Пластоглобулы - это липопротеиновые субкомпоненты хлоропласта, которые постоянно связаны с тилакоидными мембранами и содержат биосинтетические ферменты». Растительная клетка. 18 (7): 1693–703. Дои:10.1105 / tpc.105.039859. ЧВК 1488921. PMID 16731586.
- ^ а б Крамптон-Тейлор М., Грандисон С., Пнг К.М., Бушби А.Дж., Смит А.М. (февраль 2012 г.). «Контроль количества крахмальных гранул в хлоропластах Arabidopsis». Физиология растений. 158 (2): 905–16. Дои:10.1104 / стр.111.186957. ЧВК 3271777. PMID 22135430.
- ^ Zeeman SC, Delatte T, Messerli G, Umhang M, Stettler M, Mettler T, Streb S, Reinhold H, Kötting O (2007). «Распад крахмала: недавние открытия предполагают различные пути и новые механизмы». Функциональная биология растений. 34 (6): 465–73. Дои:10.1071 / FP06313. PMID 32689375. S2CID 15995416.
- ^ Роша Дж. Д. (1998). Молекулярная биология хлоропластов и митохондрий хламидомонады. Дордрехт [u.a.]: Kluwer Acad. Publ. С. 550–565. ISBN 978-0-7923-5174-0.
- ^ а б c d е ж Ганнинг Б. Э., Стир М. В. (1996). Биология растительной клетки: структура и функции. Бостон, Массачусетс: издательство "Джонс и Бартлетт". п.24. ISBN 0-86720-504-0.
- ^ Hanson D, Andrews TJ, Badger MR (2002). "Изменчивость CO на основе пиреноидов2 концентрирующий механизм у роголистников (Anthocerotophyta) ». Функциональная биология растений. 29 (3): 407–16. Дои:10.1071 / PP01210. PMID 32689485.
- ^ а б Ма Й, Поллок С. В., Сяо Й, Куннусами К., Морони СП (июнь 2011 г.). «Идентификация нового гена CIA6, необходимого для нормального образования пиреноидов у Chlamydomonas reinhardtii». Физиология растений. 156 (2): 884–96. Дои:10.1104 / стр.111.173922. ЧВК 3177283. PMID 21527423.
- ^ а б Retallack B, Butler RD (январь 1970 г.). «Развитие и строение пиреноидов Bulbochaete hiloensis». Журнал клеточной науки. 6 (1): 229–41. PMID 5417694.
- ^ Браун MR, Арнотт HJ (1970). «Структура и функции водорослевого пиреноида» (PDF). Журнал психологии. 6: 14–22. Дои:10.1111 / j.1529-8817.1970.tb02350.x. S2CID 85604422. Архивировано из оригинал (PDF) 31 мая 2013 г.. Получено 31 декабря 2012.
- ^ а б c d е ж грамм Bussi Y; Шимони Э; Вайнер А; Капон Р; Чаруви Д; Нево Р; Efrati E; Рейх Z (2019). «Фундаментальная спиральная геометрия укрепляет фотосинтетическую мембрану растений». Proc Natl Acad Sci USA. 116 (44): 22366–22375. Дои:10.1073 / pnas.1905994116. ЧВК 6825288. PMID 31611387.
- ^ Инфангер С., Бишоф С., Хильтбруннер А., Агне Б., Багинский С., Кесслер Ф. (март 2011 г.). «Рецептор импорта хлоропластов Toc90 частично восстанавливает накопление клиентских белков Toc159 в мутанте ppi2 Arabidopsis thaliana» (PDF). Молекулярный завод. 4 (2): 252–63. Дои:10.1093 / mp / ssq071. PMID 21220583.
- ^ «тилакоид». Словарь Merriam-Webster. Мерриам-Вебстер. Получено 19 мая 2013.
- ^ а б c d е ж грамм час Мустарди Л., Баттл К., Штайнбах Г., Гараб Г. (октябрь 2008 г.). «Трехмерная сеть тилакоидных мембран у растений: квазигелическая модель сборки гранум-строма». Растительная клетка. 20 (10): 2552–7. Дои:10.1105 / tpc.108.059147. ЧВК 2590735. PMID 18952780.
- ^ Паолилло-младший, диджей (1970). «Трехмерное расположение межзеренных пластинок в хлоропластах». J Cell Sci. 6 (1): 243–55. PMID 5417695.
- ^ Шимони Э; Рав-Хон О; Охад I; Брамфельд V; Райх Z (2005). «Трехмерная организация тилакоидных мембран хлоропластов высших растений, выявленная методом электронной томографии». Растительная клетка. 17 (9): 2580–6. Дои:10.1105 / tpc.105.035030. ЧВК 1197436. PMID 16055630.
- ^ а б Остин-младший, Стэхелин Л.А. (апрель 2011 г.). «Трехмерная архитектура тилакоидов гран и стромы высших растений, определенная с помощью электронной томографии». Физиология растений. 155 (4): 1601–11. Дои:10.1104 / стр.110.170647. ЧВК 3091084. PMID 21224341.
- ^ «Хлоропласт в растительной клетке». TUMEGGY / НАУЧНАЯ ФОТОБИБЛИОТЕКА. Получено 19 августа 2020.
- ^ а б c Кэмпбелл Н.А., Рис Дж. Б., Урри Л. А., Кейн М. Л., Вассерман, Минорский П. В., Джексон Р. Б. (2009). Биология (8-е изд.). Бенджамин Каммингс (Пирсон). С. 190–193. ISBN 978-0-8053-6844-4.
- ^ «Австралийские ученые открыли первый новый хлорофилл за 60 лет». Сиднейский университет. 20 августа 2010 г.
- ^ а б c Такаичи С. (15 июня 2011 г.). «Каротиноиды в водорослях: распределение, биосинтез и функции». Морские препараты. 9 (6): 1101–18. Дои:10.3390 / md9061101. ЧВК 3131562. PMID 21747749.
- ^ Шепли Д. (15 октября 2012 г.). «Почему листья меняют цвет осенью?». Новости Статьи. Получено 21 мая 2013.
- ^ а б Хоу С.Дж., Барбрук А.С., Нисбет Р.Э., Локхарт П.Дж., Ларкум А.В. (август 2008 г.). «Происхождение пластид». Философские труды Лондонского королевского общества. Серия B, Биологические науки. 363 (1504): 2675–85. Дои:10.1098 / rstb.2008.0050. ЧВК 2606771. PMID 18468982.
- ^ «Знакомство с родофитами». Музей палеонтологии Калифорнийского университета. Получено 20 мая 2013.
- ^ а б c d е ж грамм Кэмпбелл Н.А., Рис Дж. Б., Урри Л. А., Кейн М. Л., Вассерман, Минорский П. В., Джексон Р. Б. (2009). Биология (8-е изд.). Бенджамин Каммингс (Пирсон). С. 200–201. ISBN 978-0-8053-6844-4.
- ^ Лоутон-младший (март 1988 г.). «Ультраструктура хлоропластных мембран в листьях кукурузы и райграсса, выявленная методами селективного окрашивания». Новый Фитолог. 108 (3): 277–283. Дои:10.1111 / j.1469-8137.1988.tb04163.x. JSTOR 2433294.
- ^ Робертс К. (2007). Справочник по растениеводству. Чичестер, Западный Сассекс, Англия: Wiley. п. 16. ISBN 978-0-470-05723-0.
- ^ Кэмпбелл Н.А., Рис Дж. Б., Урри Л. А., Кейн М. Л., Вассерман, Минорский П. В., Джексон Р. Б. (2009). Биология (8-е изд.). Бенджамин Каммингс (Пирсон). п. 742. ISBN 978-0-8053-6844-4.
- ^ а б Лоусон Т. и Дж. И. Л. Морисон. Эссе 10.1 Фотосинтез охранных клеток. Физиология и развитие растений, шестое издание [1]
- ^ а б Wells, C .; Балиш, Э. (1979). «Митогенная активность липополисахарида для клеток селезенки из стерильных, обычных и гнотобиотических крыс». Канадский журнал микробиологии. 25 (9): 1087–93. Дои:10,1139 / м79-166. PMID 540263.
- ^ а б c d е ж грамм час я j k л Глинн Дж. М., Миягишима С. Ю., Йодер Д. В., Остерён К. В., Витха С. (май 2007 г.). «Подразделение хлоропластов». Трафик. 8 (5): 451–61. Дои:10.1111 / j.1600-0854.2007.00545.x. PMID 17451550. S2CID 2808844.
- ^ а б Донг XJ, Нагай Р., Такаги С. (1998). «Микрофиламенты закрепляют хлоропласты вдоль внешней периклинальной стенки в эпидермальных клетках Vallisneria посредством взаимодействия PFR и фотосинтеза». Физиология растений и клеток. 39 (12): 1299–306. Дои:10.1093 / oxfordjournals.pcp.a029334.
- ^ а б c Такаги С. (июнь 2003 г.). «Актиновое фотоориентационное движение хлоропластов в растительных клетках». Журнал экспериментальной биологии. 206 (Pt 12): 1963–9. Дои:10.1242 / jeb.00215. PMID 12756277.
- ^ Зейгер Э., Талботт Л.Д., Фречилла С., Шривастава А., Чжу Дж. (2002). «Хлоропласт замыкающей клетки: перспектива двадцать первого века». Новый Фитолог. 153 (3): 415–424. Дои:10.1046 / j.0028-646X.2001.NPH328.doc.x.
- ^ а б c d е ж грамм Падманабхан М.С., Динеш-Кумар СП (ноябрь 2010 г.). «Все готово - роль хлоропластов, эндоплазматического ретикулума и ядра в управлении врожденным иммунитетом растений». Молекулярные взаимодействия растений и микробов. 23 (11): 1368–80. Дои:10.1094 / MPMI-05-10-0113. PMID 20923348.
- ^ Katsir L, Chung HS, Koo AJ, Howe GA (август 2008 г.). «Жасмонатная передача сигналов: консервативный механизм восприятия гормонов». Текущее мнение в области биологии растений. 11 (4): 428–35. Дои:10.1016 / j.pbi.2008.05.004. ЧВК 2560989. PMID 18583180.
- ^ Schnurr JA, Shockey JM, de Boer GJ, Browse JA (август 2002 г.). «Экспорт жирных кислот из хлоропласта. Молекулярная характеристика основной пластидной ацил-кофермент А синтетазы из Arabidopsis». Физиология растений. 129 (4): 1700–9. Дои:10.1104 / стр.003251. ЧВК 166758. PMID 12177483.
- ^ а б c d Биология - концепции и связи. Пирсон. 2009. С. 108–118.
- ^ Кэмпбелл Н.А., Уильямсон Б., Хейден Р.Дж. (2006). Биология: изучение жизни. Бостон, Массачусетс: Пирсон Прентис Холл. ISBN 978-0-13-250882-7.[страница нужна ]
- ^ Ягендорф А.Т., Урибе Э. (январь 1966 г.). «Образование АТФ, вызванное кислотно-основным переходом хлоропластов шпината». Труды Национальной академии наук Соединенных Штатов Америки. 55 (1): 170–7. Bibcode:1966ПНАС ... 55..170J. Дои:10.1073 / пнас.55.1.170. ЧВК 285771. PMID 5220864.
- ^ а б Берг Дж. М., Тимочко Дж. Л., Страйер Л. (2002). Биохимия (5. изд., 4. печатн. Изд.). Нью-Йорк, штат Нью-Йорк [u.a.]: W. H. Freeman. С. Раздел 19.4. ISBN 0-7167-3051-0.
- ^ а б Берг Дж. М., Тимочко Дж. Л., Страйер Л. (2002). Биохимия (5. изд., 4. печатн. Изд.). Нью-Йорк, штат Нью-Йорк [u.a.]: W. H. Freeman. С. Раздел 20.1. ISBN 0-7167-3051-0.
- ^ а б c Wample RL, Дэвис RW (сентябрь 1983 г.). «Влияние затопления на накопление крахмала в хлоропластах подсолнечника (Helianthus annuus L.)». Физиология растений. 73 (1): 195–8. Дои:10.1104 / стр.73.1.195. ЧВК 1066435. PMID 16663176.
- ^ Карми А., Шомер I (1979). «Накопление крахмала и фотосинтетическая активность в первичных листьях фасоли (Phaseolus vulgaris Л.) ». Анналы ботаники. 44 (4): 479–484. Дои:10.1093 / oxfordjournals.aob.a085756.
- ^ Берг Дж. М., Тимочко Дж. Л., Страйер Л. (2002). Биохимия (5-е изд.). W H Freeman. стр. Раздел 19.4. Получено 30 октября 2012.
- ^ а б c Хаузер М., Эйхельманн Х., Ожа В., Хебер У., Лайск А. (июль 1995 г.). «Стимуляция светом быстрой регуляции pH в строме хлоропласта in vivo, на что указывает солюбилизация CO2 в листьях». Физиология растений. 108 (3): 1059–1066. Дои:10.1104 / стр.108.3.1059. ЧВК 157457. PMID 12228527.
- ^ а б Вердан К., Хельдт Х.В., Милованцев М. (август 1975 г.). «Роль pH в регуляции фиксации углерода в строме хлоропластов. Исследования фиксации CO2 в темноте и свете». Biochimica et Biophysica Acta (BBA) - Биоэнергетика. 396 (2): 276–92. Дои:10.1016/0005-2728(75)90041-9. PMID 239746.
- ^ а б c d е ж грамм Берджесс Дж. (1989). Введение в развитие растительных клеток. Кембридж: издательство Кембриджского университета. п. 56. ISBN 0-521-31611-1.
- ^ Ферро М., Сальви Д., Ривьер-Роллан Х., Верма Т., Сеньорэн-Берни Д., Грюнвальд Д. и др. (Август 2002 г.). «Интегральные мембранные белки оболочки хлоропластов: идентификация и субклеточная локализация новых переносчиков». Труды Национальной академии наук Соединенных Штатов Америки. 99 (17): 11487–92. Bibcode:2002PNAS ... 9911487F. Дои:10.1073 / pnas.172390399. ЧВК 123283. PMID 12177442.
- ^ а б Роллан Н., Дру М., Дус Р. (март 1992 г.). «Субклеточное распределение O-ацетилсерин (тиол) лиазы в соцветиях цветной капусты (Brassica oleracea L.)». Физиология растений. 98 (3): 927–35. Дои:10.1104 / стр.98.3.927. ЧВК 1080289. PMID 16668766.
- ^ Раванель С., Гакиер Б., Джоб Д., Дус Р. (июнь 1998 г.). «Особенности биосинтеза и метаболизма метионина в растениях». Труды Национальной академии наук Соединенных Штатов Америки. 95 (13): 7805–12. Bibcode:1998PNAS ... 95.7805R. Дои:10.1073 / пнас.95.13.7805. ЧВК 22764. PMID 9636232.
- ^ а б c d е Бьюкенен ББ, Груиссем У., Джонс Р.Л. (ред.). 2015. Биохимия и молекулярная биология растений. Вили Блэквелл.
- ^ Джойард Дж., Ферро М., Масселон С., Сеньорэн-Берни Д., Салви Д., Гарин Дж., Роллан Н. (апрель 2010 г.).«Протеомика хлоропластов подчеркивает субклеточную компартментацию липидного обмена». Прогресс в исследованиях липидов. 49 (2): 128–58. Дои:10.1016 / j.plipres.2009.10.003. PMID 19879895.
- ^ Праудлав М.О., Турман Д.А. (1981). «Поглощение 2-оксоглутарата и пирувата изолированными хлоропластами гороха». Новый Фитолог. 88 (2): 255–264. Дои:10.1111 / j.1469-8137.1981.tb01722.x.
- ^ Бао Х, Фокке М., Поллард М., Олрогге Дж. 2000. Понимание снабжения предшественником углерода in vivo для синтеза жирных кислот в ткани листа. Журнал растений 22, 39–50.
- ^ Ольрогге Дж., Обзор Дж. (1995). «Биосинтез липидов». Растительная клетка. 7 (7): 957–970. Дои:10.1105 / tpc.7.7.957. ЧВК 160893. PMID 7640528.
- ^ Ганнинг Б. Э., Стир М. В. (1996). Биология растительной клетки: структура и функции. Бостон, Массачусетс: издательство "Джонс и Бартлетт". п.20. ISBN 0-86720-504-0.
- ^ а б c d е Берджесс Дж. (1989). Введение в развитие растительных клеток (Пбк. Ред.). Кембридж: издательство Кембриджского университета. С. 54–55. ISBN 0-521-31611-1.
- ^ Берджесс Дж. (1989). Введение в развитие растительных клеток (Пбк. Ред.). Кембридж: издательство Кембриджского университета. п. 57. ISBN 0-521-31611-1.
- ^ а б Possingham JV, Rose RJ (1976). «Репликация хлоропластов и синтез ДНК хлоропластов в листьях шпината». Труды Королевского общества B: биологические науки. 193 (1112): 295–305. Bibcode:1976RSPSB.193..295P. Дои:10.1098 / rspb.1976.0047. S2CID 2691108.
- ^ Cannon GC, Heinhorst S (1 января 1993 г.). «Репликация ДНК в хлоропластах». Журнал клеточной науки. 104 (1): 1–9. CiteSeerX 10.1.1.1026.3732.
- ^ а б c d е ж грамм час я Миягишима С.Ю., Нисида К., Мори Т., Мацузаки М., Хигасияма Т., Куроива Н., Куроива Т. (март 2003 г.). «Специфический для растений белок, связанный с динамином, образует кольцо в месте деления хлоропласта». Растительная клетка. 15 (3): 655–65. Дои:10.1105 / tpc.009373. ЧВК 150020. PMID 12615939.
- ^ а б Хашимото Х., Поссингем СП (апрель 1989 г.). «Влияние света на цикл деления хлоропластов и синтез ДНК в культивируемых листовых дисках шпината». Физиология растений. 89 (4): 1178–83. Дои:10.1104 / стр.89.4.1178. ЧВК 1055993. PMID 16666681.
- ^ а б c d Хансен А.К., Эскобар Л.К., Гилберт Л.Е., Янсен Р.К. (январь 2007 г.). «Отцовское, материнское и двупарительское наследование генома хлоропластов у Passiflora (Passifloraceae): значение для филогенетических исследований» (PDF). Американский журнал ботаники. 94 (1): 42–6. Дои:10.3732 / ajb.94.1.42. PMID 21642206.
- ^ Бирки CW (декабрь 1995 г.). «Однородительское наследование митохондриальных и хлоропластных генов: механизмы и эволюция». Труды Национальной академии наук Соединенных Штатов Америки. 92 (25): 11331–8. Bibcode:1995PNAS ... 9211331B. Дои:10.1073 / пнас.92.25.11331. ЧВК 40394. PMID 8524780.
- ^ Пауэлл В., Морганте М., МакДевитт Р., Вендрамин Г.Г., Рафальски Дж. А. (август 1995 г.). «Полиморфные области повторов простых последовательностей в геномах хлоропластов: приложения к популяционной генетике сосен». Труды Национальной академии наук Соединенных Штатов Америки. 92 (17): 7759–63. Bibcode:1995PNAS ... 92.7759P. Дои:10.1073 / пнас.92.17.7759. ЧВК 41225. PMID 7644491.
У сосен геном хлоропластов передается через пыльцу.
- ^ Стегеманн С., Хартманн С., Руф С., Бок Р. (июль 2003 г.). «Высокочастотный перенос генов из генома хлоропласта в ядро». Труды Национальной академии наук Соединенных Штатов Америки. 100 (15): 8828–33. Bibcode:2003ПНАС..100.8828С. Дои:10.1073 / pnas.1430924100. ЧВК 166398. PMID 12817081.
- ^ а б Руф С., Карчер Д., Бок Р. (апрель 2007 г.). «Определение уровня содержания трансгена, обеспечиваемого трансформацией хлоропластов». Труды Национальной академии наук Соединенных Штатов Америки. 104 (17): 6998–7002. Bibcode:2007ПНАС..104.6998Р. Дои:10.1073 / pnas.0700008104. ЧВК 1849964. PMID 17420459.
внешняя ссылка
- Хлоропласт - Клеточная база данных
- Clegg MT, Gaut BS, Learn GH, Morton BR (июль 1994). «Скорость и закономерности эволюции ДНК хлоропластов». Труды Национальной академии наук Соединенных Штатов Америки. 91 (15): 6795–801. Bibcode:1994PNAS ... 91.6795C. Дои:10.1073 / пнас.91.15.6795. ЧВК 44285. PMID 8041699.
- Co-Extra исследование трансформации хлоропластов
- NCBI полный геном хлоропласта
| Субдисциплины | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Растение группы | |||||||||||
| |||||||||||
| |||||||||||
| Рост и привычка растений | |||||||||||
| Размножение | |||||||||||
| Таксономия растений | |||||||||||
| Упражняться | |||||||||||
| |||||||||||
| |||||||||||
| Авторитетный контроль |
|---|