Палеоботаника - Paleobotany
Эта статья нужны дополнительные цитаты для проверка. (Октябрь 2011 г.) (Узнайте, как и когда удалить этот шаблон сообщения) |

| Часть серии по |
| Палеонтология |
|---|
 |
История палеонтологии |
Отрасли палеонтологии |
| Палеонтологический портал Категория |
Палеоботаника, который также пишется как палеоботаника, это ветвь ботаника занимается восстановлением и идентификацией растение остается от геологический контекстов и их использование для биологической реконструкции прошлые среды (палеогеография), а эволюционная история растений, имеющий отношение к эволюция жизни в целом. Синоним палеофитология. Это компонент палеонтология и палеобиология. Префикс палео- означает "древний, старый",[1] и получен из Греческий прилагательное παλαιός, палаи.[2] Палеоботаника включает изучение наземное растение окаменелости, а также изучение доисторический морской фотоавтотрофы, Такие как фотосинтетический водоросли, водоросли или же ламинария. Тесно родственная область - это палинология, который является исследованием окаменелый и сохранившийся споры и пыльца.
Палеоботаника играет важную роль в реконструкции древних экологический системы и климат, известный как палеоэкология и палеоклиматология соответственно; и имеет фундаментальное значение для изучения зеленого растение разработка и эволюция. Палеоботаника также стала важной в области археология, в первую очередь для использования фитолиты в относительное свидание И в палеоэтноботаника.
Возникновение палеоботаники как научной дисциплины можно увидеть в начале 19 века, особенно в работах немецкого палеонтолога. Эрнст Фридрих фон Шлотхайм, чешский (богемский) дворянин и ученый Каспар Мария фон Штернберг, и французский ботаник Адольф-Теодор Бронгниар.[3][4]
Обзор палеоботанической записи
Макроскопические остатки истинного сосудистые растения впервые встречаются в ископаемое запись во время Силурийский Период Палеозой эпоха. Некоторые дисперсные, фрагментарные окаменелости спорного сродства, в первую очередь споры и кутикула, были найдены в породах из Ордовик Период в Оман, и считаются производными от печеночник - или же мох ископаемые растения класса (Веллман, Остерлофф и Мохиуддин, 2003 г. ).

Важным местонахождением окаменелостей ранних наземных растений является Райни Черт, найдено за пределами села Райни в Шотландия. Rhynie Chert - ранний Девонский спекать (горячий источник ) депозит состоит в основном из кремнезем. Это исключительное место благодаря сохранению нескольких различных клады растений, из мхи и ликофиты к более необычным, проблемным формам. Многие ископаемые животные, в том числе членистоногие и паукообразные, также находятся в Райни Черте, и он предлагает уникальное окно в историю ранней земной жизни.
Растительного происхождения окаменелости в конце Девонский и включать дерево стволы листья, и корни. Считалось, что самое раннее дерево Археоптерис, который несет простой, папоротник -подобно листья расположены по спирали на ветвях на вершине хвойное дерево -подобно хобот (Мейер-Берто, Шеклер и Вендт, 1999 г. ), хотя теперь известно, что это недавно открытый Wattieza.[5]
Широко распространен каменный уголь отложения болот в Северной Америке и Европе во время Каменноугольный Период содержит множество окаменелостей, содержащих древовидные ликоподы до 30 метров высотой, обильный семенные растения, Такие как хвойные породы и семенные папоротники, и бесчисленное множество меньше, травянистый растения.
Покрытосеменные (цветущие растения ) возникла в Мезозойский, а пыльца и листья цветковых растений впервые появляются в период раннего Меловой, примерно 130 миллионов лет назад.
Окаменелости растений
А растение ископаемое какая-либо сохранившаяся часть растение который давно умер. Такие окаменелости могут быть доисторическими оттисками, которым много миллионов лет, или кусочками уголь которым всего несколько сотен лет. Доисторические растения - это различные группы растений, которые жили до того, как зарегистрировано. история (раньше примерно 3500 г. до н.э. ).
Сохранение окаменелостей растений

Ископаемые остатки растений можно сохранить разными способами, каждый из которых может дать различную информацию об исходном родительском растении. Эти способы сохранения обсуждаются на общих страницах на окаменелости но в палеоботаническом контексте их можно резюмировать следующим образом.
- Adpressions (сжатия - впечатления). Это наиболее часто встречающийся тип окаменелостей растений. Они обеспечивают хорошую морфологическую детализацию, особенно дорсивентральных (уплощенных) частей растения, таких как листья. Если кутикула сохранена, они также могут дать прекрасные анатомические детали эпидермиса. Остальные детали клеточной анатомии обычно сохраняются.
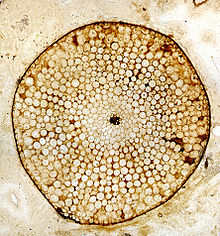 Риния, Нижний девонский Райни Черт, Шотландия, Великобритания. Поперечный разрез ствола, сохранившийся в виде окаменения кремнезема, демонстрирующий сохранение ячеистой структуры.
Риния, Нижний девонский Райни Черт, Шотландия, Великобритания. Поперечный разрез ствола, сохранившийся в виде окаменения кремнезема, демонстрирующий сохранение ячеистой структуры. - Окаменение (перминерализация или окаменелости анатомической сохранности). Они обеспечивают прекрасную детализацию клеточной анатомии растительной ткани. Морфологические детали также можно определить с помощью серийных срезов, но это занимает много времени и сложно.
- Формы и отливки. Они имеют тенденцию сохранять только более прочные части растения, такие как семена или древесные стебли. Они могут предоставить информацию о трехмерной форме растения, а в случае слепков пней могут предоставить доказательства плотности исходной растительности. Однако они редко сохраняют какие-либо мелкие морфологические детали или анатомию клеток. Подмножество таких окаменелостей пробковые слепки, где центр стебля либо полый, либо с тонкой сердцевиной. После смерти попадает осадок и образует слепок центральной полости стебля. Наиболее известные образцы сердцевинных слепков встречаются у Sphenophyta каменноугольного периода (Каламиты) и кордаиты (Artisia).
 Crossotheca hughesiana Кидстон, Средний Пенсильвания, Козли, недалеко от Дадли, Великобритания. Орган пыльцы lyginopteridalean, сохранившийся как аутигенная минерализация (минерализованный на месте). Образец в музее Седжвик, Кембридж, Великобритания.
Crossotheca hughesiana Кидстон, Средний Пенсильвания, Козли, недалеко от Дадли, Великобритания. Орган пыльцы lyginopteridalean, сохранившийся как аутигенная минерализация (минерализованный на месте). Образец в музее Седжвик, Кембридж, Великобритания. - Аутигенные минерализации. Они могут обеспечить очень мелкие трехмерные морфологические детали и оказались особенно важными при изучении репродуктивных структур, которые могут сильно искажаться при адпрессиях. Однако, поскольку они образуются в минеральных конкрециях, такие окаменелости редко бывают большого размера.
- Фусаин. Огонь обычно разрушает растительную ткань, но иногда остатки древесного угля могут сохранить мелкие морфологические детали, которые теряются при других способах сохранения; Некоторые из лучших свидетельств ранних цветов сохранились в фузине. Окаменелости фузаинов хрупкие и часто маленькие, но из-за их плавучести могут часто дрейфовать на большие расстояния и, таким образом, могут свидетельствовать о растительности вдали от зон отложений.


Ископаемые таксоны
Окаменелости растений почти всегда представляют собой разрозненные части растений; даже небольшие травянистые растения редко сохраняются целыми. Те несколько примеров окаменелостей растений, которые кажутся остатками целых растений, на самом деле неполны, поскольку внутренняя клеточная ткань и мелкие микроморфологические детали обычно теряются во время окаменелости. Остатки растений можно сохранить различными способами, каждый из которых раскрывает различные особенности исходного родительского растения.
Из-за этих трудностей палеоботаники обычно присваивают разные таксономические названия разным частям растения в разных режимах сохранения. Например, в субарборесцентном палеозое сфенофиты, отпечаток листа можно отнести к роду Annularia, сжатие конуса, связанное с Палеостахия, и основа, назначенная либо Каламиты или же Артроксилон в зависимости от того, сохранился ли он в виде слепка или окаменения. Все эти окаменелости, возможно, произошли от одного и того же родительского растения, но каждой из них дано собственное таксономическое название. Такой подход к названию окаменелостей растений возник в результате работ Адольф Бронниар[6] и выдержал испытание временем.
В течение многих лет этот подход к названию окаменелостей растений принимался палеоботаниками, но не получил официального оформления в рамках Международные правила ботанической номенклатуры.[7] В итоге, Томас (1935) и Йонгманс, Галле и Готан (1935) предложил комплекс формальных положений, суть которых была внесена в 1952 г. Международный кодекс ботанической номенклатуры.[8] Эти ранние положения позволяли отнести окаменелости, представляющие определенные части растений в определенном состоянии сохранности, к родам органов. Кроме того, небольшая подгруппа родов органов, известная как роды форм, была признана на основе искусственных таксонов, введенных Brongniart (1822) в основном для окаменелостей листвы. С годами концепции и правила, касающиеся родов органов и форм, претерпели изменения в рамках последовательных кодов номенклатуры, что отражает неспособность палеоботанического сообщества прийти к соглашению о том, как этот аспект таксономической номенклатуры растений должен работать (история, рассмотренная Клил и Томас (2010) ). От использования родов органов и ископаемых отказались Кодекс Сент-Луиса (Greuter et al. 2000 г. ), замененный на «морфотакса».
Ситуация в Венский кодекс 2005 г.[9] было то, что любой таксон растений, тип которого является ископаемым, кроме Диатомеи, можно описать как морфотаксон, определенная часть растения, сохраненная определенным образом. Хотя название всегда привязано к типу экземпляра, его объем (т.е. диапазон экземпляров, которые могут быть включены в таксон) определяется таксономистом, который использует это имя. Такое изменение ограничения может привести к расширению диапазона частей растений и / или состояний сохранности, которые могут быть включены в таксон. Например, ископаемый род, изначально основанный на сжатии семяпочек, может быть использован для включения многояйцевых купул, внутри которых семяпочки изначально родились. Если в данном случае существует уже названный ископаемый род этих купул, может возникнуть сложность. Если бы палеоботаники были уверены, что тип ископаемого рода яйцеклеток и ископаемого рода купул может быть включен в один и тот же род, то эти два названия будут соревноваться за то, чтобы быть правильным для недавно измененного рода.
Morphotaxa были введены, чтобы попытаться преодолеть проблему конкурирующих названий, которые представляли различные части растений и / или состояния сохранности. Что бы вы сделали, если бы название вида пыльцевого органа предшествовало названию вида того типа пыльцы, который производит этот орган пыльцы. Утверждалось, что палеоботаники были бы недовольны, если бы органы пыльцы были названы с использованием таксономического названия, типовым образцом которого является пыльцевое зерно. Как указал Клил и Томас (2010) Однако риск того, что название пыльцевого зерна заменит название пыльцевого органа, маловероятен. Палеоботаники должны быть полностью уверены в том, что типовой образец вида пыльцы, который обычно представляет собой рассредоточенное зерно, определенно произошел от того же растения, которое произвело орган пыльцы. Мы знаем по современным растениям, что близкородственные, но разные виды могут производить пыльцу практически неотличимой. Казалось бы, морфотаксы не дают палеоботаникам реального преимущества перед обычными ископаемыми таксонами, и эта концепция была отброшена на ботаническом конгрессе 2011 г. и конференции 2012 г. Международный кодекс номенклатуры водорослей, грибов и растений.
Ископаемые группы растений



Некоторые растения остались почти неизменными на протяжении всей геологической временной шкалы Земли. Хвощи развились к позднему девону,[10] рано папоротники был разработан Миссисипец, хвойные породы посредством Пенсильванский. Некоторые из доисторических растений такие же, как и сегодня, и поэтому живые окаменелости, Такие как Гинкго билоба и Sciadopitys verticillata. Другие растения радикально изменились или вымерли.
Примеры доисторических растений:
- Араукария мирабилис
- Археоптерис
- Каламиты
- Dillhoffia
- Глоссоптерис
- Hymenaea protera
- Nelumbo aureavallis
- Пахиптерис
- Палеораф
- Пелтандра примаева
- Протосальвиния
- Троходендрон наста
Известные палеоботаники
- Эдвард В. Берри (1875–1945), палеоэкология и фитогеография
- Уильям Гилберт Чалонер (1928–2016)
- Изабель Куксон (1893–1973), ранние сосудистые растения, палинология
- Дайан Эдвардс (1942–), колонизация земли ранними наземными флорами
- Томас Максвелл Харрис (1903–1983), мезозойские растения Земли Джеймсон (Гренландия) и Йоркшира.
- Роберт Кидстон (1852–1924), ранние наземные растения, флоры девона и карбона и их использование в стратиграфии.
- Этель Ида Санборн (1883–1952), вымершая флора Орегона и западной части Соединенных Штатов
- Бирбал Сахни (1891–1949), пересмотр растений индийской гондваны
- Данкинфилд Генри Скотт (1854–1934), анализ строения ископаемых растений.
- Константин фон Эттингсхаузен (1826–1897), третичные флоры
- Каспар Мария фон Штернберг (1761–1838), «отец палеоботаники»
- Франц Унгер (1800–1870), пионер физиологии растений, фитотомии и почвоведения.
- Джек А. Вулф (1936–2005), Третичный палеоклимат западной части Северной Америки.
- Гилберт Артур Лейсман (1924–1996), известный своими работами по ликофитам каменноугольного периода центральной Северной Америки.
Смотрите также
- Криптоспоры
- Эволюционная история растений
- Палеофикология
- Доисторическая жизнь
- Хронология эволюции растений
Рекомендации
- ^ Стерн, W.T. (2004). Ботаническая латынь (4-е (п / б) изд.). Портленд, Орегон: Timber Press. п. 460. ISBN 978-0-7153-1643-6.
- ^ Лидделл, Генри Джордж и Скотт, Роберт (1940). "παλαιός". Греко-английский лексикон. Оксфорд: Clarendon Press. Получено 2019-07-16.
- ^ "Бронгниар, Адольф-Теодор". www.encyclopedia.com. Encyclopedia.com: БЕСПЛАТНЫЙ онлайн-словарь. Получено 22 февраля 2017.
- ^ Клил, Кристофер Дж .; Лазарь, Морин; Таунсенд, Аннет (2005). «Иллюстрации и иллюстраторы в« золотой век »палеоботаники: 1800–1840». В Bowden, A.J .; Burek, C.V .; Wilding, R. (ред.). История палеоботаники: избранные очерки. Лондон: Геологическое общество Лондона. п.41. ISBN 9781862391741.
- ^ Спир, Брайан Р. (10 июня 1995 г.), Девонский период, получено 12 мая 2012
- ^ Brongniart (1822)
- ^ Брике, Дж. (1906), Международные правила ботанической номенклатуры, принятые в рамках Международного конгресса по ботанике в Вене, 1905 г., Йена: Фишер, OCLC 153969885
- ^ Lanjouw et al. 1952 г.
- ^ Макнил 2006
- ^ Эльгорриага, А .; Escapa, I.H .; Rothwell, G.W .; Tomescu, A.M.F .; Кунео, Н. (2018). "Происхождение Equisetum: эволюция хвоща (Equisetales) в пределах основной клады эвфиллофитов Sphenopsida". Американский журнал ботаники. 105 (8): 1286–1303. Дои:10.1002 / ajb2.1125. PMID 30025163.
дальнейшее чтение
- Brongniart, A. (1822), "Sur la classification et la distribution des végétaux fossiles en général, et sur ceux des terrains de supérieur en specific", Mém. Mus. Natl. Hist. Nat., 8: 203–240, 297–348
- Клил, С.Дж. и Томас, Б.А. (2010), «Ботаническая номенклатура и окаменелости растений», Таксон, 59: 261–268, Дои:10.1002 / налог.591024
- Greuter, W .; McNeill, J .; Barrie, F R .; Burdet, H.M .; Демулин, В .; Filgueiras, T.S .; Nicolson, D.H .; Silva, P.C .; Skog, J.E .; Терланд, штат Нью-Джерси, и Хоксворт, Д.Л. (2000), Международный кодекс ботанической номенклатуры (Кодекс Сент-Луиса), Кенигштейн: Научные книги Кельца, ISBN 978-3-904144-22-3
- Jongmans, W.J .; Галле, Т. И Готан, В. (1935), Предлагаемые дополнения к Международным правилам ботанической номенклатуры, принятые пятым Международным ботаническим конгрессом в Кембридже 1930 г., Херлен, OCLC 700752855
- Lanjouw, J .; Baehni, C .; Merrill, E.D .; Rickett, H.W .; Робинс, В .; Спраг, Т.А. И Стафлеу, Ф.А. (1952), Международный кодекс ботанической номенклатуры: принят седьмым Международным ботаническим конгрессом; Стокгольм, июль 1950 г., Regnum Vegetabile 3, Утрехт: Международное бюро таксономии растений Международной ассоциации таксономии растений, OCLC 220069027
- McNeill, J .; и др., ред. (2006), Международный кодекс ботанической номенклатуры (Венский кодекс), принятый семнадцатым Международным ботаническим конгрессом, Вена, Австрия, июль 2005 г. (электронное издание), Вена: Международная ассоциация таксономии растений, архив из оригинал 6 октября 2012 г., получено 2011-02-20
- Мейер-Берто, Бриджит; Шеклер, С. И Вендт, Дж. (1999) "Археоптерис является самым ранним современным деревом », Природа, 398 (6729): 700–701, Bibcode:1999Натура.398..700М, Дои:10.1038/19516, S2CID 4419663
- Томас, HH (1935), «Предлагаемые дополнения к Международным правилам ботанической номенклатуры, предложенные британскими палеоботаниками» (PDF), Журнал ботаники, 73: 111
- Wellman, Charles H .; Остерлофф, Питер Л. и Мохиуддин, Узма (2003), «Фрагменты древнейших наземных растений» (PDF), Природа, 425 (6955): 282–285, Bibcode:2003 Натур. 425..282Вт, Дои:10.1038 / природа01884, PMID 13679913, S2CID 4383813
- Уилсон Н. Стюарт и Гар В. Ротвелл. 2010 г. Палеоботаника и эволюция растений, Второе издание. Издательство Кембриджского университета, Кембридж, Великобритания. ISBN 978-0-521-38294-6.
- Томас Н. Тейлор, Эдит Л. Тейлор и Майкл Крингс. 2008 г. Палеоботаника: биология и эволюция ископаемых растений, 2-е изд. Academic Press (отпечаток Elsevier): Берлингтон, Массачусетс; Нью-Йорк, штат Нью-Йорк; Сан-Диего, Калифорния, США, Лондон, Великобритания. 1252 страницы. ISBN 978-0-12-373972-8.
внешняя ссылка
- Международная организация палеоботаники
- Ботаническое общество Америки - палеоботаническая секция
- Группа исследований палеоботаники, Университет Мюнстера, Германия
- Биота ранних наземных экосистем: Райни Черт, Университет Абердина, Великобритания
- Библиография палеоботаники
- Проект Штернберга
- PaleoNet - рассылки и ссылки по палеонтологии
- Растения Парка Юрского периода Растения, которые жили, когда по земле бродили динозавры
- Глобальный реестр научных названий ископаемых организмов
- Ссылки для палеоботаников
| Субдисциплины | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Растение группы | |||||||||||
| |||||||||||
| |||||||||||
| Рост и привычка растений | |||||||||||
| Размножение | |||||||||||
| Таксономия растений | |||||||||||
| Упражняться | |||||||||||
| |||||||||||
| |||||||||||