Морфология растений - Plant morphology
Фитоморфология это исследование физическая форма и внешнее строение из растения.[1] Обычно это считается отличным от анатомия растений,[1] который является изучением внутреннего структура растений, особенно на микроскопическом уровне.[2] Морфология растений полезна для визуальной идентификации растений. Недавние исследования в молекулярная биология начал исследовать молекулярные процессы, участвующие в определении сохранения и разнообразия морфологии растений. В этих исследованиях транскриптом образцы сохранения были обнаружены, чтобы отметить важные онтогенетический переходы во время завод жизненный цикл, который может привести к эволюционным ограничениям, ограничивающим диверсификацию[3].

Объем


Морфология растений «представляет собой исследование развития, формы и структуры растений и, как следствие, попытку интерпретировать их на основе сходства плана и происхождения».[4] Морфология растений состоит из четырех основных областей, каждая из которых частично совпадает с другой областью исследований. Биологические науки.
Прежде всего, морфология сравнительныйЭто означает, что морфолог изучает структуры многих разных растений одного или разных видов, затем проводит сравнения и формулирует идеи о сходстве. Когда считается, что структуры у разных видов существуют и развиваются в результате общих, унаследованных генетический пути, эти структуры называются гомологичный. Например, уходит из сосна, дуб, и капуста все они выглядят по-разному, но имеют общую структуру и расположение частей. О гомологии листьев легко сделать. Морфолог растений идет дальше и обнаруживает, что шипы из кактус также имеют ту же базовую структуру и развитие, что и листья у других растений, поэтому колючки кактуса также гомологичны листьям. Этот аспект морфологии растений частично совпадает с изучением растений. эволюция и палеоботаника.
Во-вторых, морфология растений учитывает как вегетативный (соматический) конструкции растений, а также репродуктивный конструкции. Вегетативные структуры сосудистые растения включает изучение системы побегов, состоящей из стебли и уходит, так же хорошо как корень система. Репродуктивные структуры более разнообразны и обычно характерны для определенной группы растений, таких как цветы и семена, папоротник сори, и мох капсулы. Детальное изучение репродуктивных структур растений привело к открытию смена поколений содержится во всех растениях и большинстве водоросли. Эта область морфологии растений частично совпадает с изучением биоразнообразие и систематика растений.
В-третьих, морфология растений изучает структуру растений в различных масштабах. В самых маленьких масштабах ультраструктура, общие структурные особенности клеток видны только с помощью электронный микроскоп, и цитология, исследование клеток с помощью оптическая микроскопия. В этом масштабе морфология растений пересекается с анатомия растений как область исследования. В самом крупном масштабе - это изучение растений. привычка к росту, общая архитектура завода. Схема ветвления в дерево будет варьироваться от вида к виду, как и внешний вид растения как дерево, трава, или трава.
В-четвертых, морфология растений изучает характер развитие, процесс, посредством которого структуры возникают и созревают по мере роста растения. В то время как животные производят все части тела, которые они когда-либо будут иметь с раннего возраста, растения постоянно производят новые ткани и структуры на протяжении всей своей жизни. У живого растения всегда есть зародышевые ткани. На то, как созревают новые структуры по мере их создания, может влиять момент в жизни растения, когда они начинают развиваться, а также среда, которой подвергаются эти структуры. Морфолог изучает этот процесс, причины и его результат. Эта область морфологии растений пересекается с физиология растений и экология.
Сравнительная наука
Морфолог растений сравнивает структуры многих растений одного или разных видов. Проведение таких сравнений между подобными структурами на разных заводах решает вопрос Зачем конструкции похожи. Вполне вероятно, что подобные основные причины генетики, физиологии или реакции на окружающую среду привели к этому внешнему сходству. Результат научного исследования этих причин может привести к одному из двух открытий лежащей в основе биологии:
- Гомология - структура схожа у двух видов из-за общего происхождения и общей генетики.
- Конвергенция - структура похожа у двух видов из-за независимой адаптации к общим воздействиям окружающей среды.
Понимание того, какие характеристики и структуры принадлежат каждому типу, является важной частью понимания растений. эволюция. Биолог-эволюционист полагается на морфолога растений для интерпретации структур и, в свою очередь, дает филогении взаимоотношений растений, которые могут привести к новым морфологическим открытиям.
Гомология
Когда считается, что структуры у разных видов существуют и развиваются в результате общих наследственных генетических путей, эти структуры называют гомологичный. Например, листья сосны, дуба и капусты выглядят по-разному, но имеют общие основные структуры и расположение частей. О гомологии листьев легко сделать. Морфолог растений идет дальше и обнаруживает, что колючки кактуса также имеют ту же основную структуру и развитие, что и листья других растений, и поэтому колючки кактуса также гомологичны листьям.
Конвергенция
Когда считается, что структуры у разных видов существуют и развиваются в результате общих адаптивных реакций на давление окружающей среды, эти структуры называют сходящийся. Например, листья Bryopsis plumosa и основы Спаржа щетинистая оба имеют одинаковые перистые ветвления, хотя одна из них - водоросль, а другая - цветущее растение. Сходство в общей структуре происходит независимо в результате конвергенции. Форма роста многих кактусы и виды Молочай очень похожи, хотя и принадлежат к очень далеким семьям. Сходство является результатом общих решений проблемы выживания в жаркой и сухой среде.
Вегетативные и репродуктивные характеристики
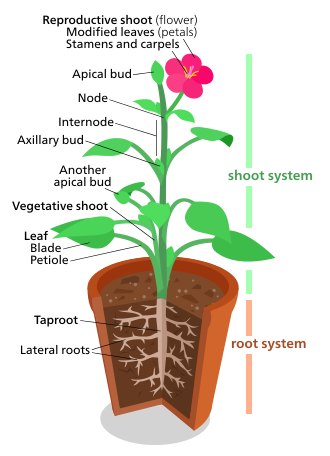
Морфология растений касается как вегетативных структур растений, так и репродуктивных структур.
В вегетативный (соматический) структуры сосудистые растения включают две основные системы органов: (1) система стрельбы, состоящий из стебли и уходит, и (2) a корневая система. Эти две системы являются общими почти для всех сосудистых растений и представляют собой объединяющую тему для изучения морфологии растений.
Напротив, репродуктивный структуры разнообразны и обычно характерны для определенной группы растений. Такие структуры как цветы и фрукты находятся только в покрытосеменные; сори находятся только в папоротники; и шишки находятся только в хвойные породы и другие голосеменные. Поэтому репродуктивные признаки считаются более полезными для классификации растений, чем вегетативные признаки.
Использование в идентификации
Биологи растений используют морфологические признаки растений, которые можно сравнивать, измерять, подсчитывать и описывать, чтобы оценить различия или сходства в таксонах растений и использовать эти признаки для идентификации, классификации и описания растений.
Когда символы используются в описаниях или для идентификации, они называются диагностический или ключевые персонажи которые могут быть как качественными, так и количественными.
- Количественные признаки - это морфологические признаки, которые можно подсчитать или измерить, например, у вида растений лепестки цветов шириной 10–12 мм.
- Качественные признаки - это морфологические признаки, такие как форма листьев, цвет цветка или опушение.
Оба типа символов могут быть очень полезны для идентификации растений.
Смена поколений
Детальное изучение репродуктивных структур растений привело к открытию смена поколений, встречается во всех растениях и большинстве водоросли, немецкий ботаник Вильгельм Хофмайстер. Это открытие - одно из самых важных, сделанных во всей морфологии растений, поскольку оно обеспечивает общую основу для понимания жизненного цикла всех растений.
Пигментация у растений
Основная функция пигментов в растениях - фотосинтез, который использует зеленый пигмент хлорофилл вместе с несколькими красными и желтыми пигментами, которые помогают улавливать как можно больше световой энергии. Пигменты также являются важным фактором привлечения насекомые к цветам, чтобы ободрить опыление.
Растительные пигменты включают множество различных видов молекул, в том числе: порфирины, каротиноиды, антоцианы и беталаины. Все биологические пигменты избирательно поглощают определенные длины волн свет в то время как отражающий другие. Поглощаемый свет может использоваться растением для стимуляции химических реакций, а длина отраженной волны света определяет цвет, который пигмент будет казаться глазом.
Морфология в развитии
Завод развитие это процесс, посредством которого структуры возникают и созревают по мере роста растения. Это предмет изучения в анатомия растений и физиология растений а также морфология растений.
Процесс развития растений принципиально отличается от того, что наблюдается у растений. позвоночное животное животные. Когда животное эмбрион начинает развиваться, он очень рано производит все части тела, которые когда-либо будут у него в жизни. Когда животное рождается (или вылупляется из яйца), у него есть все части тела, и с этого момента оно будет только расти и становиться более зрелым. Напротив, растения на протяжении всей своей жизни постоянно производят новые ткани и структуры из меристемы[5] расположены на концах органов или между зрелыми тканями. Таким образом, у живого растения всегда есть зародышевые ткани.
Свойства организации, наблюдаемые в растении: эмерджентные свойства которые больше, чем сумма отдельных частей. «Сборка этих тканей и функций в единый многоклеточный организм дает не только характеристики отдельных частей и процессов, но и совершенно новый набор характеристик, которые нельзя было бы предсказать на основе исследования отдельных частей».[6] Другими словами, знания всего о молекулах в растении недостаточно, чтобы предсказать характеристики клеток; а знание всех свойств клеток не позволяет предсказать все свойства структуры растения.
Рост
А сосудистое растение начинается с одноклеточной зигота, образована оплодотворение яйцеклетки с помощью сперматозоида. С этого момента он начинает делиться, образуя растение. эмбрион через процесс эмбриогенез. Когда это произойдет, полученные клетки будут организованы так, что один конец станет первым корнем, а другой конец - верхушкой побега. В семя растения, у эмбриона разовьется один или несколько «семенных листков» (семядоли ). К концу эмбриогенеза у молодого растения будут все части, необходимые для начала его жизни.
Однажды эмбрион прорастает из своего семени или родительского растения он начинает производить дополнительные органы (листья, стебли и корни) в процессе органогенез. Новые корни растут из корня меристемы расположен на кончике корня, и из побега вырастают новые стебли и листья меристемы находится на кончике побега.[7] Ветвление происходит, когда небольшие скопления клеток, оставленные меристемой и еще не подвергшиеся клеточная дифференциация чтобы сформировать специализированную ткань, начинают расти как верхушка нового корня или побега. Рост из любой такой меристемы на кончике корня или побега называется первичный рост и приводит к удлинению этого корня или побега. Вторичный рост приводит к расширению корня или побега из делений клеток в камбий.[8]
Помимо роста на ячейка деление, растение может прорасти удлинение ячейки. Это происходит, когда отдельные клетки или группы клеток становятся длиннее. Не все растительные клетки вырастают до одинаковой длины. Когда клетки на одной стороне ствола растут дольше и быстрее, чем клетки на другой стороне, в результате ствол будет изгибаться в сторону более медленно растущих клеток. Этот направленный рост может происходить через реакцию растения на определенный стимул, такой как свет (фототропизм ), сила тяжести (гравитропизм ), воды, (гидротропизм ) и физический контакт (тигмотропизм ).
Рост и развитие растений опосредуются специфическими гормоны растений и регуляторы роста растений (ГРР) (Росс и др., 1983).[9] Уровни эндогенных гормонов зависят от возраста растений, морозостойкости, состояния покоя и других метаболических состояний; фотопериод, засуха, температура и другие внешние условия окружающей среды; и экзогенные источники ГРР, например, применяемые извне и ризосферного происхождения.
Морфологическая вариация
Растения демонстрируют естественные вариации по форме и строению. В то время как все организмы различаются от человека к человеку, растения демонстрируют дополнительный тип изменений. В пределах одного человека повторяются части, которые могут отличаться по форме и структуре от других подобных частей. Это изменение наиболее легко увидеть на листьях растения, хотя другие органы, такие как стебли и цветы, могут иметь аналогичные изменения. У этой вариации есть три основные причины: позиционные эффекты, влияние окружающей среды и молодость.
Эволюция морфологии растений
Факторы транскрипции и сети регуляции транскрипции играют ключевую роль в морфогенезе растений и их эволюции. Во время посадки растений появилось много новых семейств факторов транскрипции, которые преимущественно связаны с сетями многоклеточного развития, репродукции и развития органов, внося вклад в более сложный морфогенез наземных растений.[10]
Позиционные эффекты

Хотя растения производят множество копий одного и того же органа в течение своей жизни, не все копии одного и того же органа будут идентичными. Существуют различия между частями зрелого растения, обусловленные относительным положением органа. Например, на новой ветке листья могут иметь одинаковый узор вдоль ветки. Форма листьев, образующихся у основания ветки, будет отличаться от листьев, образующихся на верхушке растения, и эта разница сохраняется от ветки к ветке на данном растении и в данном виде. Это различие сохраняется после того, как листья на обоих концах ветки созрели, и не является результатом того, что одни листья моложе других.
Экологические последствия
На то, как созревают новые структуры по мере их создания, может влиять момент в жизни растений, когда они начинают развиваться, а также среда, которой подвергаются эти структуры. Это можно увидеть в водные растения и надводные растения.
Температура
Температура оказывает множество воздействий на растения в зависимости от множества факторов, включая размер и состояние растения, а также температуру и продолжительность воздействия. Чем меньше и больше сочные растения, тем выше подверженность повреждению или смерти от слишком высоких или слишком низких температур. Температура влияет на скорость биохимических и физиологических процессов, скорость обычно (в определенных пределах) увеличивается с температурой. Однако соотношение Вант-Гоффа для мономолекулярных реакций (которое гласит, что скорость реакции удваивается или утроится при повышении температуры на 10 ° C) не строго соблюдается для биологических процессов, особенно при низких и высоких температурах.
Последствия замерзания воды в растениях во многом зависят от того, происходит ли замерзание внутриклеточно (внутри клеток) или вне клеток в межклеточном (внеклеточном) пространстве.[11] Внутриклеточное замораживание обычно приводит к гибели клетки вне зависимости от морозостойкости растения и его тканей.[12] Внутриклеточное замораживание в природе встречается редко, но умеренные скорости снижения температуры, например, от 1 ° C до 6 ° C / час, вызывают образование межклеточного льда, и этот «внеклеточный лед»[13] может быть или не быть летальным, в зависимости от прочности ткани.
При отрицательных температурах сначала замерзает вода в межклеточных пространствах тканей растений, хотя вода может оставаться незамерзшей до тех пор, пока температура не упадет ниже 7 ° C.[11] После первоначального межклеточного образования льда клетки сжимаются, поскольку вода теряется в сегрегированном льду. Клетки подвергаются сублимационной сушке, причем обезвоживание является основной причиной обморожения.
Показано, что скорость охлаждения влияет на морозостойкость тканей,[14] но фактическая скорость замораживания будет зависеть не только от скорости охлаждения, но также от степени переохлаждения и свойств ткани.[15] Сакаи (1979a)[14] продемонстрировал сегрегацию льда в зачатках побегов белой и черной елей Аляски при медленном охлаждении до 30–40 ° C. Эти замороженные и обезвоженные почки пережили погружение в жидкий азот при медленном согревании. Аналогичным образом отреагировали и цветочные зачатки. Внеорганное промерзание в зачатках объясняет способность наиболее выносливых из северных хвойных деревьев переживать зимы в регионах, где температура воздуха часто опускается до -50 ° C и ниже.[13] Зимостойкость почек таких хвойные породы усиливается за счет небольшого размера почек, более быстрого перемещения воды и способности переносить интенсивное обезвоживание замораживанием. У бореальных видов Picea и Pinus, морозостойкость однолетних сеянцев на уровне зрелых растений,[16] учитывая подобные состояния покоя.
Юность

Органы и ткани, производимые молодым растением, например, саженец, часто отличаются от тех, которые производятся тем же растением, когда они стареют. Это явление известно как юность или гетеробластика. Например, молодые деревья будут давать более длинные и тонкие ветви, которые будут расти вверх больше, чем ветви, которые они будут производить как полностью выросшее дерево. Кроме того, листья, образовавшиеся на раннем этапе роста, обычно крупнее, тоньше и неправильной формы, чем листья взрослого растения. Образцы молодых растений могут настолько отличаться от взрослых растений того же вида, что яйцекладущие насекомые не распознают растение как пищу для своего потомства. Различия видны в укоренении и цветении, а также на одном и том же зрелом дереве. Молодые черенки, взятые у основания дерева, образуют корни гораздо легче, чем черенки, исходящие из средней и верхней кроны. Цветение у основания дерева отсутствует или менее обильно, чем цветение на более высоких ветвях, особенно когда молодое дерево впервые достигает возраста цветения.[17]
Переход от ранних форм роста к поздним обозначается как «изменение вегетативной фазы ', но есть некоторые разногласия по поводу терминологии.[18]
Современная морфология растений
Рольф Саттлер пересмотрел фундаментальные концепции сравнительной морфологии, такие как концепция гомология. Он подчеркнул, что гомология должна также включать частичную гомологию и количественную гомологию.[19][20] Это приводит к континуальной морфологии, которая демонстрирует континуум между морфологическими категориями корня, побега, стебля (каулом), листа (филлом) и волос (трихома). Как лучше всего описываются промежуточные звенья между категориями, обсуждали Bruce K. Kirchoff et al.[21] Недавнее исследование, проведенное Stalk Institute, извлекло координаты, соответствующие основанию и листьям каждого растения в трехмерном пространстве. Когда растения на графике были размещены в соответствии с их фактическим расстоянием перемещения питательных веществ и общей длиной ветвей, растения почти идеально попадали на кривую Парето. «Это означает, что способ выращивания растений в своей архитектуре также оптимизирует очень распространенный компромисс при проектировании сети. В зависимости от окружающей среды и вида, завод выбирает различные способы достижения компромиссов для этих конкретных условий окружающей среды». [22]
В честь Агнес Арбер, автора теория частичного побега листа, Рутисхаузер и Ислер назвали континуальный подход нечеткой арберовской морфологией (FAM). «Нечеткий» относится к нечеткая логика, «Арбериан» в Агнес Арбер. Рутисхаузер и Ислер подчеркнули, что этот подход подтверждается не только многими морфологическими данными, но и данными молекулярной генетики.[23] Более свежие данные молекулярной генетики предоставляют дополнительную поддержку морфологии континуума. Джеймс (2009) пришел к выводу, что «в настоящее время широко признано, что ... радиальность [характеристика большинства стеблей] и дорсивентральность [характеристика листьев] - всего лишь крайности непрерывного спектра. Фактически, это просто синхронизация гена KNOX. выражение !. "[24] Эккардт и Баум (2010) пришли к выводу, что «сейчас общепринято, что сложные листья выражают свойства как листьев, так и побегов».[25]
Морфология процесса описывает и анализирует динамический континуум формы растений. Согласно этому подходу конструкции не имеют процесс (ы), они находятся процесс (ы).[26][27][28] Таким образом, дихотомия структура / процесс преодолевается «расширением нашего понятия« структура », чтобы включить и признать, что в живом организме это не просто вопрос пространственной структуры с« активностью »как чем-то сверх или против. это, но что конкретный организм - это пространственно-временный структура и что эта пространственно-временная структура и есть сама деятельность ".[29]
Для Жен, Барабе и Лакруа классическая морфология (то есть основная морфология, основанная на концепции качественной гомологии, подразумевающей взаимоисключающие категории) и морфология континуума являются подклассами более всеобъемлющей морфологии процесса (динамической морфологии).[30]
Классическая морфология, морфология континуума и морфология процесса очень важны для эволюции растений, особенно в области эволюционной биологии растений (evo-DevO), которая пытается объединить морфологию растений и молекулярную генетику растений.[31] В подробном тематическом исследовании необычных морфологий Рутисхаузер (2016) проиллюстрировал и обсудил различные темы эволюции растений, такие как нечеткость (непрерывность) морфологических концепций, отсутствие взаимно однозначного соответствия между структурными категориями и экспрессией генов. , понятие морфопространства, адаптивная ценность особенностей bauplan по сравнению с патио ludens, физиологические адаптации, обнадеживающие монстры и сальтационная эволюция, значение и пределы устойчивости развития и т. д.[32]
Нравится нам это или нет, но морфологические исследования находятся под влиянием философских допущений, таких как либо / или логика, нечеткая логика, дуализм структуры / процесса или его трансцендентность. А эмпирические данные могут повлиять на философские предположения. Таким образом, существует взаимодействие между философией и эмпирическими открытиями. Эти взаимодействия являются предметом того, что было названо философией морфологии растений.[33]
Смотрите также
- Словарь морфологии растений
- Анатомия растений
- Идентификация растений
- Физиология растений
- Эволюционная биология развития растений
- Таксономия
использованная литература
- ^ а б Рэйвен П. Х., Р. Ф. Эверт и С. Э. Эйххорн. Биология растений, 7-е изд., Стр. 9. (Нью-Йорк: У. Х. Фриман, 2005). ISBN 0-7167-1007-2.
- ^ Эверт, Рэй Франклин и Исав, Кэтрин (2006) Анатомия растения Исава: меристемы, клетки и ткани растительного тела - их структура, функции и развитие Уайли, Хобокен, Нью-Джерси, страница xv, ISBN 0-471-73843-3
- ^ Дрост, Хайк-Георг; Белльштедт, Джулия; Ó'Maoiléidigh, Diarmuid S .; Сильва, Андерсон Т .; Габель, Александр; Вайнхольдт, Клаус; Райан, Патрик Т .; Dekkers, Bas J.W .; Бенцинк, Леони; Hilhorst, Henk W.M .; Лигтеринк, Уилко; Веллмер, Фрэнк; Гроссе, Иво; Квинт, Марсель (23 февраля 2016 г.). «Постэмбриональные модели песочных часов отмечают онтогенетические переходы в развитии растений». Молекулярная биология и эволюция. 33 (5): 1158–1163. Дои:10.1093 / molbev / msw039. PMID 26912813.
- ^ Гарольд К. Болд, К. Дж. Алексопулос и Т. Делеворяс. Морфология растений и грибов, 5-е изд., Стр. 3. (Нью-Йорк: Харпер-Коллинз, 1987). ISBN 0-06-040839-1.
- ^ Bäurle, I; Ло, Т. (2003). «Апикальные меристемы: фонтан молодости растений». BioEssays. 25 (10): 961–70. Дои:10.1002 / bies.10341. PMID 14505363. Обзор.
- ^ Леопольд, А.С. Рост и развитие растений, стр. 183. (Нью-Йорк: McGraw-Hill, 1964).
- ^ Бренд, U; Хобе, М; Саймон, Р. (2001). «Функциональные домены в меристемах побегов растений». BioEssays. 23 (2): 134–41. Дои:10.1002 / 1521-1878 (200102) 23: 2 <134 :: AID-BIES1020> 3.0.CO; 2-3. PMID 11169586. Обзор.
- ^ Барлоу, П. (2005). «Узорчатое определение клеток в растительной ткани: вторичная флоэма деревьев». BioEssays. 27 (5): 533–41. Дои:10.1002 / bies.20214. PMID 15832381.
- ^ Ross, S.D .; Pharis, R.P .; Биндер, У. Д. 1983. Регуляторы роста и хвойные деревья: их физиология и потенциальное использование в лесном хозяйстве. п. 35–78 в Никелл, Л. (Ред.), Химические вещества, регулирующие рост растений. Vol. 2, CRC Press, Бока-Ратон, Флорида.
- ^ Jin JP; и другие. (Июль 2015 г.). «Карта регуляторов транскрипции Arabidopsis выявляет отличительные функциональные и эволюционные особенности новых факторов транскрипции». Молекулярная биология и эволюция. 32 (7): 1767–1773. Дои:10.1093 / molbev / msv058. ЧВК 4476157. PMID 25750178.
- ^ а б Glerum, C. 1985. Морозостойкость саженцев хвойных пород: принципы и применение. п. 107–123 в Дурья, М. (Ред.). Ход работы: Оценка качества рассады: принципы, процедуры и прогностические возможности основных тестов. Workshop, октябрь 1984 г., Университет штата Орегон, For. Res. Lab., Corvallis OR.
- ^ Lyons, J.M .; Raison, J.K .; Степонкус, П. 1979. Растительная мембрана в ответ на низкую температуру: обзор. п. 1–24 в Lyons, J.M .; Graham, D .; Raison, J.K. (Ред.). Низкотемпературный стресс у сельскохозяйственных культур. Academic Press, Нью-Йорк, штат Нью-Йорк.
- ^ а б Sakai, A .; Ларчер, В. (ред.) 1987. Морозостойкость растений. Springer-Verlag.
- ^ а б Сакаи, A. 1979a. Механизм предотвращения промерзания примордиальных побегов почек хвойных. Physiol растительных клеток. 20: 1381–1390.
- ^ Левитт, Дж. 1980. Реакция растений на стрессы окружающей среды. Том 1. Переохлаждение, замерзание и высокотемпературные напряжения, 2-е изд. Academic Press, Нью-Йорк, штат Нью-Йорк. 497 с.
- ^ Sakai, A .; Окада, С. 1971. Морозостойкость хвойных пород. Silvae Genet. 20 (3): 91–97.
- ^ Майкл А. Дирр; Чарльз Хойзер-младший (2006). «2». Справочное руководство по размножению древесных растений (Второе изд.). Varsity Press Inc., стр. 26, 28, 29. ISBN 0942375092.
- ^ Джонс, Синтия С. (1999-11-01). «Очерк молодости, фазового изменения и гетеробластики семенных растений». Международный журнал наук о растениях. 160 (S6): –105 – S111. Дои:10.1086/314215. ISSN 1058-5893. PMID 10572025.
- ^ Саттлер Р. (1984). «Гомология - непрекращающийся вызов». Систематическая ботаника. 9 (4): 382–394. Дои:10.2307/2418787. JSTOR 2418787.
- ^ Саттлер Р., 1994, Гомология, гомеоз и морфология процессов у растений. В: Б.К. Холл (ред.) Гомология: иерархическая основа сравнительной морфологии. Нью-Йорк: Academic Press, стр. 423-475.
- ^ Кирхгоф, Б. К.; Pfeifer, E; Рутисхаузер, Р. (2008). «Онтология структуры растений: как мы должны маркировать структуры растений с сомнительной или смешанной идентичностью?». Zootaxa. 1950: 103–122. Дои:10.11646 / zootaxa.1950.1.10.
- ^ Конн, Адам; Педмале, Уллас; Чори, Джоанн (2017). «Лазерное сканирование с высоким разрешением показывает, что архитектура предприятия отражает универсальные принципы проектирования сети». Сотовые системы. 5 (1): 103–122. Дои:10.1016 / j.cels.2017.06.017. PMID 28750198.
- ^ Rutishauser, R .; Ислер, Б. (2001). «Генетика развития и морфологическая эволюция цветковых растений, особенно пузырчатых (Utricularia): нечеткая арберовская морфология дополняет классическую морфологию» (PDF). Анналы ботаники. 88 (6): 1173–1202. Дои:10.1006 / anbo.2001.1498.
- ^ Джеймс, П. Дж. (2009). "'Дерево и лист: Другой ракурс ». Линней. 25: 13–19.
- ^ Eckardt, NA; Баум, Д. (2010). «Загадка Podostemad: эволюция необычной морфологии у Podostemaceae». Растительная клетка. 22 (7): 2131–2140. Дои:10.1105 / tpc.110.220711. ЧВК 2929115. PMID 20647343.
- ^ Саттлер, Р. (1992). «Морфология процессов: структурная динамика в развитии и эволюции». Канадский журнал ботаники. 70 (4): 708–714. Дои:10.1139 / b92-091.
- ^ Вергара-Силва, Ф. (2003). "Растения и концептуальная артикуляция эволюционной биологии развития". Биология и философия. 18 (2): 261–264. Дои:10.1023 / А: 1023936102602. S2CID 81013686.
- ^ Саттлер, Р. 2019. Структурные и динамические подходы к развитию и эволюции формы растений. В: Fusco, G. (ed) Перспективы эволюционной биологии и биологии развития. Очерки для Алессандро Минелли. Глава 6, с. 57-70 [1]
- ^ Вудгер, Дж. 1967 г. Биологические принципы. Лондон: Рутледж и Кегоан Пол (переиздан с новым Введением).
- ^ Jeune, B; Barabé, D; Лакруа, С. (2006). «Классическая и динамическая морфология: к синтезу через пространство форм». Acta Biotheoretica. 54 (4): 277–293. Дои:10.1007 / s10441-007-9007-8. PMID 17486414. S2CID 25928998.
- ^ Минелли, А. 2018. Эволюционная биология развития растений. Эволюционируемость фенотипа. Нью-Йорк: Издательство Кембриджского университета.
- ^ Рутисхаузер, Р. (2016). «Эволюция необычных морфологий у Lentibulariaceae (пузырчатые и родственные им) и Podostemonaceae (речные сорняки): графическое изображение на стыке биологии развития и морфологической диверсификации». Анналы ботаники. 117 (5): 811–832. Дои:10.1093 / aob / mcv172. ЧВК 4845801. PMID 26589968.
- ^ Саттлер, Р. (2018). «Философия морфологии растений». Elemente der Naturwissenschaft. 108: 55–79.(расширенную версию этой статьи см. [2] )
внешние ссылки
| Поля |  | |
|---|---|---|
| Бактерии | ||
| Протисты |
| |
| Растения | ||
| Беспозвоночные | ||
| Млекопитающие | ||
| Другие позвоночные | ||
| Другие темы | ||
| ||
| Субдисциплины | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Завод группы | |||||||||||
| |||||||||||
| |||||||||||
| Рост и привычка растений | |||||||||||
| Размножение | |||||||||||
| Таксономия растений | |||||||||||
| Практика | |||||||||||
| |||||||||||
| |||||||||||