Эволюционная история растений - Evolutionary history of plants


В эволюция из растения привело к широкому диапазону сложности, начиная с самого раннего маты из водорослей через многоклеточные морские и пресноводные зеленые водоросли, наземный мохообразные, ликоподы и папоротники, в комплекс голосеменные и покрытосеменные сегодня. Хотя многие из самых ранних групп продолжают процветать, о чем свидетельствует красный и зеленые водоросли в морской среде, недавно полученные группы вытеснили ранее доминирующие в экологическом отношении, например преобладание цветковых растений над голосеменными в наземных средах.[6]:498
Есть свидетельства того, что цианобактерии и многоклеточные фотосинтезирующие эукариоты жили в пресноводных сообществах на суше еще 1 миллиард лет назад.[7] и что сообщества сложных многоклеточных фотосинтезирующих организмов существовали на суше в конце Докембрийский, около 850 миллион лет назад.[8]
Свидетельства появления эмбриофит наземные растения впервые встречаются в середине ордовика (~470 миллион лет назад), а к середине девона (~390 миллион лет назад), присутствовали многие признаки, присущие сегодня наземным растениям, включая корни и листья. К позднему девону (~370 миллион лет назад) некоторые свободно спорящие растения, такие как Археоптерис было вторичная сосудистая ткань это произвело дерево и образовали леса из высоких деревьев. Также в конце девона, Элькинсия, рано семенной папоротник, развились семена.[9] Эволюционные инновации продолжались в остальной Фанерозой эон и продолжается по сей день. Большинство групп растений не пострадали от Событие пермо-триасового вымирания, хотя структура сообществ изменилась. Это могло создать основу для появления цветущих растений в Триасовый (~200 миллион лет назад), и их более поздняя диверсификация в меловом и Палеоген. Последней крупной группой растений, которые эволюционировали, были травы, который стал важным в середине палеогена, примерно со 40 миллион лет назад. Травы, как и многие другие группы, развили новые механизмы метаболизма, чтобы выжить в условиях низкой CO
2 и теплые, сухие условия тропиков за последние 10 миллион лет.
Колонизация земли
Наземные растения произошли от группы зеленые водоросли, возможно, уже 850 млн лет назад,[8] но похожие на водоросли растения могли появиться еще 1 миллиард лет назад.[7] Ближайшими живыми родственниками наземных растений являются харофиты в частности Charales; если предположить, что привычка Charales мало изменилась с момента расхождения родословных, это означает, что наземные растения произошли от разветвленной нитчатой водоросли, обитающей на мелководье пресной воды,[10] возможно, на краю сезонно высыхающих бассейнов.[11] Однако некоторые недавние данные свидетельствуют о том, что наземные растения могли произойти от одноклеточных наземных растений. харофиты похож на существующие Klebsormidiophyceae.[12] У водоросли бы гаплонтический жизненный цикл. На очень короткое время это было бы парным хромосомы (в диплоид условие), когда яйцо и сперма сначала слился, чтобы сформировать зигота который сразу же разделил бы на мейоз для производства клеток с половиной количества непарных хромосом ( гаплоидный условие). Кооперативное взаимодействие с участием грибы возможно, помогли ранним растениям адаптироваться к стрессам земного царства.[13]


Растения не были первыми фотосинтезаторами на суше. Скорость выветривания предполагает, что организмы, способные к фотосинтезу, уже жили на суше. 1,200 миллион лет назад,[11] и окаменелости микробов были обнаружены в отложениях пресноводных озер из 1,000 миллион лет назад,[14] но запись изотопов углерода предполагает, что их было слишком мало, чтобы повлиять на состав атмосферы, пока около 850 миллион лет назад.[8] Эти организмы, несмотря на филогенетическое разнообразие,[15] были, вероятно, маленькими и простыми, образовав немного больше, чем водорослевую пену.[11]
Свидетельства самого раннего наземные растения встречается намного позже, около 470 млн лет назад, в породах нижнего среднего ордовика из Саудовской Аравии.[16] и Гондвана[17] в виде спор с устойчивыми к гниению стенками. Эти споры, известные как криптоспоры, производились поодиночке (монады), парами (диады) или группами по четыре (тетрады), и их микроструктура напоминает микроструктуру современных печеночник споры, предполагая, что они разделяют эквивалентную степень организации.[18] Их стены содержат спорополленин - еще одно свидетельство эмбриофитного сродства.[19] Возможно, атмосферное «отравление» помешало эукариотам колонизировать землю до этого,[20] или могло просто потребоваться много времени, чтобы необходимая сложность эволюционировала.[21]
Споры трилете, похожие на споры сосудистые растения появляются вскоре после этого в породах верхнего ордовика около 455 миллионов лет назад.[22][23] В зависимости от того, когда именно тетрада расщепляется, каждая из четырех спор может иметь «трилетную метку», Y-образную форму, отражающую точки, в которых каждая клетка прижалась к своим соседям.[24] Однако для этого необходимо, чтобы стенки спор были прочными и стойкими на ранней стадии. Это сопротивление тесно связано с наличием устойчивой к высыханию внешней стенки - признак, который используется только тогда, когда споры должны выжить вне воды. Действительно, даже те эмбриофиты которые вернулись в воду, не имеют прочной стены и не имеют следов трилета.[24] Тщательное изучение спор водорослей показывает, что ни у кого нет спор трилете, либо потому, что их стенки недостаточно устойчивы, либо в тех редких случаях, когда это так, споры рассеиваются, прежде чем они достаточно раздавлены, чтобы развить метку, или не вписываются в тетраэдрическая тетрада.[24]
Самые ранние окаменелости наземных растений были таллоид организмов, которые обитали в речных водно-болотных угодьях и, как установлено, покрыли большую часть ранний силурийский пойма реки. Они могли выжить только тогда, когда земля была заболочена.[25] Были и микробные маты.[26]
Как только растения достигли земли, можно было использовать два подхода к борьбе с высыханием. Современные мохообразные либо избегают этого, либо уступают ему, ограничивая свой ареал влажными условиями, или высыхая и приостанавливая свой метаболизм до тех пор, пока не прибудет больше воды, как в роде печеночников. Targionia. Трахеофиты сопротивляться высыханию, контролируя скорость потери воды. Все они имеют водонепроницаемый внешний вид. кутикула слой везде, где они подвергаются воздействию воздуха (как некоторые мохообразные), чтобы уменьшить потерю воды, но поскольку полное покрытие отрезало бы их от CO
2 в атмосфере трахеофиты используют переменные отверстия, устьица, для регулирования скорости газообмена. Трахеофиты также развили сосудистую ткань, чтобы помочь в перемещении воды внутри организмов (см. ниже ), и отошла от жизненного цикла с преобладанием гаметофитов (см. ниже ). Сосудистая ткань в конечном итоге также способствовала прямому росту без поддержки воды и открыла путь для развития более крупных растений на суше.
А снежный ком земли, приблизительно от 850-630 млн лет назад, как полагают, были вызваны первыми фотосинтезирующими организмами, которые снизили концентрацию углекислого газа и увеличили количество кислорода в атмосфере.[27] Появление наземной флоры увеличило скорость накопления кислорода в атмосфере, поскольку наземные растения производили кислород как отходы. Когда эта концентрация поднялась выше 13%,[когда? ] лесные пожары стали возможны, о чем свидетельствует древесный уголь в летописи окаменелостей.[28] Помимо спорного разрыва в позднем девоне, древесный угль присутствует до сих пор.
Углефикация - важный тафономический Режим. Лесной пожар или захоронение в горячем вулканическом пепле отгоняет летучие соединения, оставляя только чистый углерод. Это не жизнеспособный источник пищи для грибов, травоядных или детритовоедов, поэтому его необходимо консервировать. Он также прочен, поэтому может выдерживать давление и отображать изысканные, иногда субячеистые, детали.
Эволюция жизненных циклов

Все многоклеточные растения имеют жизненный цикл, состоящий из двух поколений или фаз. В гаметофит фаза имеет единый набор хромосом (обозначается1п) и производит гаметы (сперму и яйца). В спорофит фаза имеет парные хромосомы (обозначены2п) и производит споры. Фазы гаметофита и спорофита могут быть гомоморфными, внешне идентичными у некоторых водорослей, таких как Ульва лактука, но очень разные у всех современных наземных растений, состояние, известное как гетероморфия.
В эволюции растений наблюдается переход от гомоморфии к гетероморфности. Водорослевые предки наземных растений почти наверняка были гаплобионтический, будучи гаплоидными на протяжении всех своих жизненных циклов, с одноклеточной зиготой, обеспечивающей стадию 2N. Все наземные растения (т.е. эмбриофиты ) находятся диплобионтический - то есть и гаплоидная, и диплоидная стадии являются многоклеточными.[6] Очевидны две тенденции: мохообразные (печеночники, мхи и роголистник ) развили гаметофит как доминирующую фазу жизненного цикла, и спорофит стал почти полностью зависимым от него; сосудистые растения развили спорофит как доминирующую фазу, причем гаметофиты особенно редки в семенные растения.
Было высказано предположение, что основанием для появления диплоидной фазы жизненного цикла как доминирующей фазы является то, что диплоидия позволяет маскировать выражение вредных мутаций посредством генетическое дополнение.[29][30] Таким образом, если один из родительских геномов в диплоидных клетках содержит мутации приводящие к дефектам в одном или нескольких генные продукты эти недостатки могут быть компенсированы другим родительским геномом (который, тем не менее, может иметь свои собственные дефекты в других генах). Поскольку диплоидная фаза становилась преобладающей, маскирующий эффект, вероятно, позволял размер генома и, следовательно, информационный контент, чтобы увеличиваться без ограничения необходимости повышения точности репликации. Возможность увеличения информационного содержания при невысокой стоимости выгодна, потому что позволяет кодировать новые адаптации. Эта точка зрения была поставлена под сомнение, поскольку свидетельства показывают, что отбор в гаплоиде не более эффективен, чем в диплоидных фазах жизненного цикла мхов и покрытосеменных.[31]
Есть две конкурирующие теории, объясняющие появление диплобионтического жизненного цикла.
В теория интерполяции (также известная как антитетическая или вставная теория)[32] считает, что интерполяция фазы многоклеточного спорофита между двумя последовательными поколениями гаметофитов была нововведением, вызванным предшествующим мейозом в только что проросшей зиготе с одним или несколькими раундами митотического деления, тем самым производя некоторую диплоидную многоклеточную ткань до того, как в конечном итоге мейоз произвел споры. Эта теория предполагает, что первые спорофиты имели очень иную и более простую морфологию по сравнению с гаметофитом, от которого они зависели.[32] Это, по-видимому, хорошо согласуется с тем, что известно о мохообразных, у которых вегетативный таллоидный гаметофит питает простой спорофит, который состоит не более чем из неразветвленного спорангия на стебле. Усложнение предков простого спорофита, включая возможное приобретение фотосинтетических клеток, освободило бы его от зависимости от гаметофита, как это видно у некоторых роголистников (Антоцерос ), и в конечном итоге приводит к тому, что спорофит развивает органы и сосудистую ткань и становится доминирующей фазой, как в трахеофитах (сосудистых растениях).[6] Эта теория может быть подтверждена наблюдениями, которые Cooksonia люди должны были поддерживаться поколением гаметофитов. Наблюдаемое появление более крупных осевых размеров с пространством для фотосинтетической ткани и, следовательно, самодостаточностью, обеспечивает возможный путь для развития самодостаточной фазы спорофитов.[32]
Альтернативная гипотеза, названная теория трансформации (или гомологическая теория), утверждает, что спорофит мог появиться внезапно, задерживая возникновение мейоза до тех пор, пока не сформируется полностью развитый многоклеточный спорофит. Поскольку один и тот же генетический материал будет использоваться как в гаплоидной, так и в диплоидной фазах, они будут выглядеть одинаково. Это объясняет поведение некоторых водорослей, таких как Ульва лактука, которые производят чередующиеся фазы одинаковых спорофитов и гаметофитов. Последующая адаптация к среде иссушающей суши, которая затрудняет половое размножение, могла привести к упрощению сексуально активного гаметофита и развитию фазы спорофита для лучшего рассеивания водонепроницаемых спор.[6] Ткань спорофитов и гаметофитов сосудистых растений, таких как Риния сохранившийся в черне Райни имеет аналогичную сложность, что считается подтверждением этой гипотезы.[32][33][34] Напротив, за исключением Псилотум современные сосудистые растения имеют гетероморфные спорофиты и гаметофиты, у которых гаметофиты редко имеют сосудистую ткань.[35]
Эволюция анатомии растений
Арбускулярный микоризный симбиоз
Нет никаких доказательств того, что у ранних наземных растений силурия и раннего девона были корни, хотя ископаемые свидетельства ризоидов встречаются у нескольких видов, таких как Хорнеофитон. У самых ранних наземных растений также не было сосудистых систем для переноса воды и питательных веществ. Аглаофитон, сосудистое растение без корней, известное по окаменелостям Девона в Рини черт[36] был первым наземным заводом, который обнаружил симбиотический связь с грибами [37] который сформировал арбускулярная микориза, буквально «древовидные грибковые корни», в четко определенном цилиндре клеток (кольцо в поперечном сечении) в коре его стеблей. Грибы питались сахаром растений в обмен на питательные вещества, произведенные или извлеченные из почвы (особенно фосфаты), к которым растение в противном случае не имело бы доступа. Как и другие бескорневые наземные растения силурия и раннего девона Аглаофитон могли полагаться на арбускулярные микоризные грибы для получения воды и питательных веществ из почвы.
Грибы принадлежали к типу Гломеромикота,[38] группа, которая, вероятно, впервые появилась 1 миллиард лет назад и до сих пор образует арбускулярные микоризные ассоциации со всеми основными группами наземных растений от мохообразных до птеридофитов, голосеменных и покрытосеменных, а также с более чем 80% сосудистых растений.[39]
Данные анализа последовательности ДНК показывают, что арбускулярный микоризный мутуализм возник у общего предка этих групп наземных растений во время их перехода на сушу.[40] и, возможно, это даже был решающий шаг, позволивший им колонизировать землю.[41] Микоризные грибы, появившиеся до того, как у этих растений образовались корни, помогали растениям в получении воды и минеральных питательных веществ, таких как фосфор в обмен на органические соединения, которые они не могли синтезировать сами.[39] Такие грибы повышают продуктивность даже простых растений, например печеночников.[42][43]
Кутикула, устьица и межклеточные пространства
Для фотосинтеза растения должны поглощать CO
2 из атмосферы. Однако делая ткани доступными для CO
2 попадание в воду позволяет воде испаряться, так что за это приходится платить.[44] Вода теряется намного быстрее, чем CO
2 впитывается, поэтому растениям необходимо его заменить. Ранние наземные растения переносили воду апопластически внутри пористых стенок их ячеек. Позже они развили три анатомических особенности, которые обеспечили способность контролировать неизбежную потерю воды, которая сопровождала CO
2 приобретение. Во-первых, водонепроницаемое внешнее покрытие или кутикула эволюционировал, что уменьшило потерю воды. Во-вторых, переменные диафрагмы, устьица который может открываться и закрываться для регулирования количества воды, теряемой при испарении во время CO
2 поглощение и, в-третьих, межклеточное пространство между клетками фотосинтетической паренхимы, что позволило улучшить внутреннее распределение CO
2 к хлоропластам. Эта трехкомпонентная система обеспечивала улучшенную гомогидрию, регулирование содержания воды в тканях, обеспечивая особое преимущество, когда подача воды непостоянна. [45] Высота CO
2 Концентрации силурия и раннего девона, когда растения впервые заселили землю, означали, что они относительно эффективно использовали воду. Так как CO
2 был выведен из атмосферы растениями, при его захвате было потеряно больше воды, и возникли более элегантные механизмы сбора и транспортировки воды.[44] Растения, растущие вверх в воздух, нуждались в системе для транспортировки воды из почвы ко всем различным частям надземного растения, особенно к фотосинтезирующим частям. К концу карбона, когда CO
2 концентрация была снижена до уровня, близкого к сегодняшнему, примерно в 17 раз больше воды терялось на единицу CO
2 поглощение.[44] Однако даже в «легкие» первые дни вода всегда была в избытке, и ее приходилось доставлять к частям растения из влажной почвы, чтобы избежать высыхания.[45]
Вода может быть злой капиллярное действие по ткани с небольшими промежутками. В узких столбах воды, например, в стенках растительных клеток или в трахеидах, когда молекулы испаряются с одного конца, они тянут молекулы за собой по каналам. Следовательно, только испарение является движущей силой переноса воды в растениях.[44] Однако без специализированных транспортных судов это сцепление-напряжение Механизм может вызвать отрицательное давление, достаточное для коллапса водопроводящих клеток, ограничивая транспортировку воды до нескольких сантиметров и, следовательно, ограничивая размер самых ранних растений.[44]

Ксилема
Чтобы быть свободным от ограничений небольшого размера и постоянной влажности, которые паренхиматозный транспортная система, заводы нуждались в более эффективной системе водного транспорта. По мере того, как растения росли вверх, специализированный водный транспорт сосудистые ткани развились сначала в виде простых гидроиды типа щетинок спорофитов мха. Эти простые удлиненные клетки были мертвыми и заполненными водой в зрелом возрасте, обеспечивая канал для переноса воды, но их тонкие, неармированные стенки разрушались при небольшом напряжении воды, ограничивая высоту растений. Ксилема трахеиды, более широкие ячейки с лигнин -армированные клеточные стенки, которые были более устойчивы к разрушению при напряжении, вызванном водным стрессом, встречаются в более чем одной группе растений к середине силурия и могут иметь единственное эволюционное происхождение, возможно, в пределах роголистников,[46] объединяющий всех трахеофитов. С другой стороны, они могли развиваться более одного раза.[44] Намного позже, в меловом периоде, за трахеидами последовали сосуды в цветущие растения.[44] По мере развития механизмов переноса воды и водонепроницаемой кутикулы растения могли выжить, не будучи постоянно покрытыми пленкой воды. Этот переход от пойкилогидрия к гомойогидрия открыл новый потенциал для колонизации.[44][45]
Раннедевонские претрахеофиты Аглаофитон и Хорнеофитон имеют неармированные водопроводные трубы со структурой стенок, очень похожей на моховых гидроидов, но они росли вместе с несколькими видами трахеофиты, такие как Риния Gwynne-Vaughanii которые имели трахеиды ксилемы, которые были хорошо усилены полосами лигнина. Самые ранние макрофоссилии, которые, как известно, имели ксилемные трахеиды, были небольшими, среднесилурийскими растениями этого рода. Cooksonia.[47] Однако утолщенные полосы на стенках отдельных фрагментов трубки видны с раннего силурия и далее.[48]
Растения продолжали изобретать способы уменьшения сопротивления потоку внутри своих клеток, постепенно увеличивая эффективность водного транспорта и увеличивая сопротивление трахеид разрушению под напряжением.[49][50] В течение раннего девона максимальный диаметр трахеиды со временем увеличивался, но, возможно, достиг плато в зостерофиллы к среднему девону.[49] Общая скорость переноса также зависит от общей площади поперечного сечения самого пучка ксилемы, и некоторые растения среднего девона, такие как Trimerophytes, имели гораздо более крупные стелы, чем их ранние предки.[49] В то время как более широкие трахеиды обеспечивали более высокую скорость переноса воды, они увеличивали риск кавитации, образования пузырьков воздуха в результате разрыва водяного столба под давлением.[44] Небольшие ямки в стенках трахеиды позволяют воде обходить дефектную трахеиду, предотвращая прохождение пузырьков воздуха[44] но ценой ограниченного расхода. К каменноугольному периоду у голосеменных появились окаймленные ямы,[51][52] клапанные структуры, которые позволяют герметизировать ямки с высокой проводимостью, когда одна сторона трахеиды сбрасывается.
Трахеиды имеют перфорированные торцевые стенки, которые оказывают большое сопротивление потоку воды.[49] но, возможно, имел преимущество в изоляции воздушной эмболии, вызванной кавитацией или замерзанием. Сосуды впервые образовались во время сухого, низкого CO
2 периоды поздней перми, в хвощах, папоротниках и Selaginellales самостоятельно, а позже появился в среднем меловом периоде у гнетофитов и покрытосеменных растений.[44] Элементы емкости представляют собой открытые трубы без торцевых стенок, расположенные встык для работы, как если бы они были одним непрерывным сосудом.[49] Сосуды позволяли древесине той же площади поперечного сечения транспортировать гораздо больше воды, чем трахеиды. [44] Это позволило растениям заполнить больше стеблей структурным волокна а также открыли новую нишу для виноградных лоз, которые могли транспортировать воду, но не были такими толстыми, как дерево, на котором они росли.[44] Несмотря на эти преимущества, древесина на основе трахеид намного легче, а значит, дешевле в производстве, так как сосуды должны быть значительно усилены, чтобы избежать кавитации.[44] Когда растения развили такой уровень контроля над испарением воды и переносом воды, они действительно стали гомоатомный, способные извлекать воду из окружающей среды через органы, похожие на корни, вместо того, чтобы полагаться на пленку поверхностной влаги, что позволяет им расти до гораздо больших размеров.[45][44] но в результате возросшей независимости от окружающей среды большинство сосудистых растений утратило способность выживать при высыхании - потерять эту особенность, которую можно очень дорого обходить.[44] У ранних наземных растений поддержка в основном обеспечивалась тургорным давлением, особенно внешнего слоя клеток, известных как стеромные трахеиды, а не ксилемой, которая была слишком маленькой, слишком слабой и находилась в слишком центральном положении, чтобы обеспечить значительную структурную поддержку. .[44] Растения с вторичная ксилема которые появились к середине девона, такие как Тримерофиты и Прогимноспермные растения имели гораздо большие сосудистые сечения, дающие прочную древесную ткань.
Эндодерма
An энтодерма возможно, образовались в самых ранних корнях растений во время девона, но первые ископаемые свидетельства такой структуры относятся к каменноугольному периоду.[44] Энтодерма в корнях окружает ткань, транспортирующую воду, и регулирует ионный обмен между грунтовыми водами и тканями и предотвращает попадание нежелательных патогенов и т. Д. В систему транспорта воды. Энтодерма также может оказывать давление вверх, вытесняя воду из корней, когда транспирации недостаточно.
Эволюция морфологии растений
Уходит


Листья первичные фотосинтетический органы современного растения. Происхождение уходит почти наверняка был вызван падением концентрации атмосферных CO
2 в девонский период повышение эффективности улавливания углекислого газа для фотосинтез.[53][54]
Листья конечно эволюционировали не раз. По своему строению они делятся на два типа: микрофиллы, у которых отсутствует сложное жилкование и, возможно, возникли в виде колючих выростов, известных как энации, и мегафиллы, которые большие и имеют сложные жилкование которые могли возникнуть в результате модификации групп ветвей. Было высказано предположение, что эти структуры возникли независимо.[55] Мегафиллы, согласно теории телома Уолтера Циммермана,[56] произошли от растений, показавших трехмерную ветвящуюся архитектуру, в результате трех трансформаций:превышение, что привело к типичному для листьев боковому положению, выравнивание, который предполагал формирование планарной архитектуры, лямка или слияние, объединившие планарные ветви, что привело к формированию собственно лист пластинка. Все три этапа произошли несколько раз в эволюции сегодняшних листьев.[57]
Широко распространено мнение, что теория теломов хорошо подтверждается свидетельствами окаменелостей. Однако Вольфганг Хагеманн поставил это под сомнение по морфологическим и экологическим причинам и предложил альтернативную теорию.[58][59] В то время как согласно теории телома самые примитивные наземные растения имеют трехмерную систему ветвления радиально-симметричных осей (теломов), согласно альтернативе Хагеманна предлагается противоположное: самые примитивные наземные растения, давшие начало сосудистым растениям, были плоскими, таллоидными. листообразная, без топоров, чем-то похожа на проталлус печеночника или папоротника. Топоры, такие как стебли и корни, позже превратились в новые органы. Рольф Саттлер предложил всеобъемлющий процессно-ориентированный взгляд, который оставляет некоторое ограниченное пространство как для теории телома, так и для альтернативы Хагемана, и, кроме того, принимает во внимание весь континуум между дорсивентральными (плоскими) и радиальными (цилиндрическими) структурами, которые можно найти в ископаемых и живых организмах. наземные растения.[60][61] Эта точка зрения подтверждается исследованиями в области молекулярной генетики. Таким образом, Джеймс (2009)[62] пришел к выводу, что «в настоящее время широко признано, что ... радиальность [характеристика осей, таких как стебли] и дорсивентральность [характеристика листьев] - всего лишь крайности непрерывного спектра. Фактически, это просто время экспрессии гена KNOX! "
До эволюции уходит растения имели фотосинтетический аппарат на стеблях.Сегодняшние листья мегафилла, вероятно, стали обычным явлением примерно через 360 миллионов лет назад, примерно через 40 миллионов лет после того, как простые безлистные растения заселили землю в Ранний девон. Этот разброс был связан с падением атмосферного углекислый газ концентрации в Позднем Палеозой эра, связанная с ростом плотности устьица на поверхности листа.[53] Это привело бы к большему испарение ставки и газообмен, но особенно при высоких CO
2 концентрации, большие листья с меньшим количеством устьиц нагреваются до смертельной температуры при полном солнечном свете. Увеличение плотности устьиц позволило лучше охладить лист, что сделало возможным его распространение, но увеличилось. СО2 поглощение за счет снижения эффективности водопользования.[54][63]
Риниофиты кремня Rhynie состояли только из тонких топоров без орнаментов. Ранний и средний девон тримерофиты можно считать листовым. Эту группу сосудистых растений можно узнать по массам терминальных спорангиев, которые украшают концы осей, которые могут раздваиваться или тройничать.[6] Некоторые организмы, такие как Псилофитон, отверстие нации. Это небольшие колючие выросты стебля, лишенные собственного кровоснабжения.
В зостерофиллы были важны уже в позднем силуре, намного раньше, чем любые риниофиты сопоставимой сложности.[64] Эта группа, узнаваемая по их почковидным спорангиям, которые росли на коротких боковых ветвях рядом с главными осями, иногда разветвлялись в характерной H-образной форме.[6] Многие зостерофиллы несут на своей оси ярко выраженные шипы.[нужна цитата ] но ни один из них не имел сосудистых следов. Первое свидетельство наличия васкуляризованных образований встречается в ископаемом клубном мхе, известном как Барагванатия которые уже упоминались в летописи окаменелостей в позднем силуре.[65] В этом организме эти следы листа продолжаются в листе, образуя его среднюю жилку.[66] Одна теория, «теория энации», утверждает, что микрофилловые листья косолапости, образовавшиеся наростами протостелы, соединяются с существующими энациями.[6] Листья рода Rhynie Астероксилон, который сохранился в Ринском кремне почти на 20 миллионов лет позже, чем Барагванатия имели примитивное кровоснабжение - в виде следы листьев отходя от центральной протостелы к каждому отдельному «листу».[67] Астероксилон и Барагванатия широко рассматриваются как примитивные ликоподы,[6] группа все еще сохранившийся сегодня в лице перья, то спайкмосы и клубные мхи. Lycopods несут отличительные микрофиллы, определяемые как листья с одним сосудистым следом. Микрофиллы могут вырасти до некоторого размера, Lepidodendrales достигая более метра в длину, но почти все несут только один сосудистый пучок. Исключением является редкое ветвление у некоторых Селагинелла виды.
Чем знакомее листья, мегафиллы, как полагают, произошли четыре раза независимо от папоротников, хвощей, прогимноспернов и семенных растений.[68] Похоже, они возникли в результате изменения дихотомия ветви, которые сначала накладывались друг на друга (или «перекрывали») друг друга, становились уплощенными или выровненными и в конечном итоге образовывали «перепонки» и постепенно превращались в более листоподобные структуры.[66] Мегафиллы, автор: Циммермана теория телома, состоят из группы перепончатых ветвей[66] и, следовательно, «листовой промежуток», оставленный там, где сосудистый пучок листа выходит из пучка основной ветви, напоминает расщепление двух осей.[66] В каждой из четырех групп, образовавших мегафиллы, их листья впервые эволюционировали в период от позднего девона до раннего карбона, быстро диверсифицируясь, пока конструкции не обосновались в середине карбона.[68]
Прекращение дальнейшей диверсификации можно объяснить ограничениями развития,[68] но почему листья вообще так долго не развивались? Растения существовали на суше не менее 50 миллионов лет, прежде чем мегафиллы стали значительными. Однако из раннедевонского рода известны небольшие редкие мезофиллы. Эофиллофитон - так что развитие не могло быть препятствием для их появления.[69] Лучшее объяснение до сих пор включает наблюдения, что атмосферные CO
2 быстро снижалась в течение этого времени - упала примерно на 90% в течение девона.[70] Это потребовало увеличения плотности устьиц в 100 раз для поддержания скорости фотосинтеза. Когда устьица открываются, позволяя воде испаряться с листьев, они обладают охлаждающим эффектом в результате потери скрытая теплота испарения. Похоже, что низкая плотность устьиц в раннем девоне означала, что испарение и испарительное охлаждение были ограничены, и что листья могли бы перегреться, если бы вырастали до любого размера. Плотность устьиц не могла увеличиваться, поскольку примитивные стелы и ограниченная корневая система не могли обеспечивать воду достаточно быстро, чтобы соответствовать скорости транспирации.[54] Очевидно, что листья не всегда полезны, о чем свидетельствует частая вторичная потеря листьев, известный пример: кактусы и "венчик папоротник" Псилотум.
Вторичная эволюция также может скрыть истинное эволюционное происхождение некоторых листьев. Некоторые роды папоротников имеют сложные листья, которые прикрепляются к псевдостеле за счет разрастания сосудистого пучка, не оставляя промежутков между листьями.[66] Далее хвощ полевой (Equisetum ) листья несут только одну жилку и кажутся микрофилловыми; однако и летопись окаменелостей, и молекулярные свидетельства указывают на то, что их предки имели листья со сложным жилкованием, а нынешнее состояние - результат вторичного упрощения.[71]
Лиственный у деревьев есть еще один недостаток, связанный с наличием листьев. Распространенное мнение, что растения сбрасывают листья, когда дни становятся слишком короткими, ошибочно; вечнозеленые растения процветали в Полярный круг вовремя самый последний тепличная земля.[72] Общепринятая причина опадания листвы зимой - это справиться с погодой - сила ветра и вес снега гораздо удобнее переносить без листьев, чтобы увеличить площадь поверхности. Сезонная потеря листьев развивалась независимо несколько раз и проявляется в гинкгоалс, немного пинофит и некоторые покрытосеменные.[73] Потеря листьев также могла возникнуть в результате давления со стороны насекомых; Возможно, было дешевле полностью потерять листья зимой или в засушливый сезон, чем продолжать вкладывать ресурсы в их ремонт.[74]
Факторы, влияющие на архитектуру листьев
Различные физические и физиологические факторы, такие как свет интенсивность, влажность, температура, скорость ветра и др. повлияли на эволюцию формы и размера листьев. Высокие деревья редко имеют большие листья, потому что они повреждаются сильным ветром. Точно так же деревья, которые растут в умеренный или тайга у регионов есть заостренные листья,[нужна цитата ] предположительно для предотвращения образования льда на поверхности листа и уменьшения потери воды из-за транспирации. Травоядность, млекопитающими и насекомые, была движущей силой в эволюции листьев. Примером может служить то, что растения рода Новой Зеландии Ацифилла имеют шипы на пластинах, которые, вероятно, препятствовали вымершим Моа от поедания ими. Другие члены Ацифилла, которые не сосуществовали с моа, не имеют этих шипов.[75]
На генетическом уровне исследования развития показали, что репрессия генов KNOX необходима для инициации лист зачаток. Это вызвано ARP гены, которые кодируют факторы транскрипции. Репрессия генов KNOX в зачатках листьев, по-видимому, вполне консервативна, в то время как экспрессия генов KNOX в листьях дает сложные листья. В ARP функция, похоже, возникла рано сосудистое растение эволюция, потому что члены примитивной группы Ликофиты также имеют функционально подобный ген.[76] Другими участниками, которые играют консервативную роль в определении зачатков листьев, являются фитогормоны. ауксин, гибберелин и цитокинин.

Расположение листьев или филлотаксия на теле растения может максимально собирать свет и, как ожидается, будет генетически крепкий. Однако в кукуруза, мутация только в одном гене называется ABPHYL (Аномальная ФИЛлотаксия) достаточно, чтобы изменить филлотаксию листьев, подразумевая, что мутационная корректировка одного локуса на геном достаточно для создания разнообразия.[77]
После того, как первичные клетки листа созданы из клеток SAM, новый топоры для роста листьев, среди них абаксиально-адаксиальные (нижняя-верхняя поверхность) оси. Гены, участвующие в определении этого и других осей, кажутся более или менее консервативными у высших растений. Белки HD-ZIPIII семья была вовлечена в определение адаксиальной идентичности. Эти белки отклоняют некоторые клетки листа зачаток по умолчанию абаксиальный заявить и сделать их адаксиальный. У ранних растений с листьями листья, вероятно, имели только один тип поверхности - абаксиальную, нижнюю сторону современных листьев. Определение адаксиальной идентичности произошло примерно через 200 миллионов лет после того, как была установлена абаксиальная идентичность.[78]
То, как формируется большое разнообразие наблюдаемых морфологий листьев растений, является предметом интенсивных исследований. Возникли некоторые общие темы. Одним из наиболее важных является участие генов KNOX в генерации сложные листья, как в помидор (см. выше). Но это не универсально. Например, горох использует другой механизм для того же.[79][80] Мутации в генах, влияющих на лист кривизна также может изменить форму листа, изменив форму листа с плоской на морщинистую,[81] как форма капуста уходит. Также существуют разные морфоген градиенты в развивающемся листе, которые определяют ось листа и могут также влиять на форму листа. Еще один класс регуляторов развития листа - это микроРНК.[82][83]
Корни
 |
 |
| Считается, что корни (нижнее изображение) Lepidodendrales (Stigmaria) по своему развитию эквивалентны стеблям (вверху), что демонстрирует сходный внешний вид «шрамов от листьев» и «шрамов от корней» на этих образцах от разных видов. |
Корни важны для растений по двум основным причинам: во-первых, они обеспечивают связь с субстратом; важнее, они обеспечивают источник воды и питательные вещества из почвы. Корни позволяли растениям расти выше и быстрее.
Эволюция корней имела последствия в глобальном масштабе. Нарушая почву и способствуя ее подкислению (поглощая питательные вещества, такие как нитраты и фосфаты[84]), они позволили ему более глубоко выветриваться, вводя углеродные соединения глубже в почвы.[85] с огромными последствиями для климата.[86] Эти эффекты могли быть настолько глубокими, что привели к массовое вымирание.[87]
Хотя в позднесилурийском периоде в ископаемых почвах есть следы корневых отпечатков,[88] окаменелости тела показывают, что самые ранние растения лишены корней. У многих были распростертые ветви, которые тянулись по земле, с вертикальными топорами или слоевища здесь и там были разбросаны точки, а у некоторых даже были нефотосинтезирующие подземные ветви, у которых не было устьиц. Различие между корневой и специализированной ветвью связано с развитием.[требуется разъяснение ] различающиеся по схеме ветвления и наличием корневой чехлик.[11] Итак, в то время как силуро-девонские растения, такие как Риния и Хорнеофитон обладал физиологическим эквивалентом корней,[89][90] корни - определяемые как органы, отличные от стеблей - появились позже.[11] К сожалению, корни редко сохраняются в летописи окаменелостей, и наше понимание их эволюционного происхождения скудно.[11]
Ризоиды - небольшие структуры, выполняющие ту же роль, что и корни, обычно размером с клетку - вероятно, возникли очень рано, возможно, даже до того, как растения заселили землю; они признаны в Characeae, родственная группа водорослей для высаживания растений.[11] Тем не менее, ризоиды, вероятно, эволюционировали более одного раза; то ризины из лишайники, например, выполнять аналогичную роль. Даже некоторые животные (Ламеллибрахия ) имеют корневую структуру.[11] Ризоиды отчетливо видны в окаменелостях кремня Rhynie и присутствовали в большинстве самых ранних сосудистых растений, и на этом основании, кажется, были предвестниками истинных корней растений.[91]
Более продвинутые структуры обычны в кремни Рини, и многие другие окаменелости сопоставимого раннедевонского возраста несут структуры, которые выглядят и действуют как корни.[11] Риниофиты несли мелкие ризоиды, а тримерофиты и травянистые ликоподы кремня имели корнеобразную структуру, проникая в почву на несколько сантиметров.[92] Однако ни одна из этих окаменелостей не отражает всех черт, присущих современным корням.[11] за исключением Астероксилон, который недавно был признан несущим корни, которые развились независимо от корней существующих сосудистых растений.[93] Корни и корнеподобные структуры становились все более распространенными и глубже проникали во время Девонский, с деревьями ликопод, образующими корни длиной около 20 см в эйфельский и живетский периоды. К ним присоединились прогимноспермные растения, укоренившиеся на глубине около метра на последующем франском этапе.[92] Настоящие голосеменные растения и папоротники зигоптериды также образовывали неглубокие корневые системы во время фаменского века.[92]
Ризофоры ликопод обеспечивают несколько иной подход к укоренению. Они были эквивалентны стеблям, а органы, эквивалентные листьям, выполняли роль корешков.[11] Аналогичное строение наблюдается у современных ликоподов. Изотес, и это, по-видимому, свидетельствует о том, что корни развивались независимо, по крайней мере, дважды, у ликофитов и других растений,[11] предположение, подтвержденное исследованиями, показывающими, что корни инициируются и их рост стимулируется различными механизмами у ликофитов и эуфиллофитов.[94]
Сосудистая система необходима корневым растениям, поскольку корням, не связанным с фотосинтезом, необходим запас сахаров, а сосудистая система необходима для транспортировки воды и питательных веществ от корней к остальным частям растения.[10] Укорененные растения[который? ] немного более развиты, чем их силурийские предки, без специальной корневой системы; однако ясно видно, что на плоских осях есть наросты, похожие на ризоиды современных мохообразных.[95]
К среднему и позднему девону большинство групп растений независимо друг от друга развили корневую систему той или иной природы.[95] По мере того как корни становились больше, они могли поддерживать более крупные деревья, а почва выветривалась на большую глубину.[87] Это более глубокое выветривание повлияло не только на вышеупомянутую просадку CO
2, но также открыли новые места обитания для колонизации грибами и животными.[92]
Сегодня корни достигли физических пределов. Они проникают в грунт на глубину до 60 метров, чтобы найти уровень грунтовых вод.[96] Самые узкие корни имеют диаметр всего 40 мкм и не могут физически переносить воду, если бы они были уже.[11]Самые ранние извлеченные ископаемые корни, напротив, сузились с 3 мм до менее 700 мкм в диаметре; конечно, тафономия это окончательный контроль того, какую толщину можно увидеть.[11]
Форма дерева


Раннедевонский пейзаж был лишен растительности выше пояса. Большая высота давала конкурентное преимущество в сборе солнечного света для фотосинтеза, затмевая конкурентов и в распространении спор, поскольку споры (а позже и семена) могли разлетаться на большие расстояния, если они начинали выше. Для достижения больших высот требовалась эффективная сосудистая система. Достичь древообразование, растения должны были развиваться древесный ткани, которая обеспечивала поддержку и транспортировку воды, и, следовательно, была необходима для развития способности к вторичный рост. Стела растений, подвергающихся вторичному росту, окружена сосудистый камбий, кольцо меристематических клеток, которое производит больше ксилемы внутри и флоэмы снаружи. Поскольку клетки ксилемы включают мертвую одревесневшую ткань, последующие кольца ксилемы добавляются к уже имеющимся, образуя древесину. Окаменелости растений раннего девона показывают, что простая форма древесины впервые появилась по крайней мере 400 миллионов лет назад, в то время, когда все наземные растения были небольшими и травянистыми.[97] Поскольку древесина возникла задолго до кустарников и деревьев, вполне вероятно, что изначально она использовалась для водного транспорта, а позже использовалась только для механической поддержки.[98]
Первыми растениями, которые развили вторичный рост и приобрели древесный облик, были, по-видимому, папоротники, а еще в среднем девоне одним из видов, Wattieza, достиг высоты 8 м и стал древовидным.[99]
Другим кладам не потребовалось много времени, чтобы развить древовидный рост. Поздний девон Археоптерис, а предшественник к голосеменные которые произошли от тримерофитов,[100] достигал 30 м в высоту. Прогимноспермные растения были первыми растениями, которые развили настоящую древесину, выросшую из двустороннего камбия. Первое появление одного из них, Реллимия, находился в среднем девоне.[101] Считается, что истинное дерево эволюционировало только один раз, что привело к концепции клады «лигнофитов».[нужна цитата ]
Археоптерис леса вскоре пополнились древовидными ликоподами в виде Lepidodendrales, которая превышала 50 м в высоту и 2 м в ширину у основания. Эти древовидные ликоподы выросли, чтобы доминировать в лесах позднего девона и карбона, которые дали начало уголь депозиты.[102] Lepidodendrales отличаются от современных деревьев тем, что демонстрируют определенный рост: после накопления запаса питательных веществ на более низкой высоте растения «улетучиваются» как единый ствол до генетически определенной высоты, ветвятся на этом уровне, распространяют свои споры и умирают.[103] Они состояли из «дешевой» древесины, что позволяло им быстро расти, по крайней мере, половина их стеблей составляла полость, заполненную сердцевиной.[6] Их древесина также была образована односторонним сосудистым камбием - он не давал новой флоэмы, а это означало, что стволы не могли расширяться со временем.[требуется проверка ]
В хвощ Каламиты появился в Каменноугольный. В отличие от современного хвоща Equisetum, Каламиты имели односторонний сосудистый камбий, что позволяло им образовывать древесину и вырастать до высоты более 10 м, а также многократно ветвиться.
Хотя форма ранних деревьев была похожа на сегодняшнюю, Сперматофиты или семенные растения, группа, в которую входят все современные деревья, еще не получили развития. Доминирующими группами деревьев сегодня являются все семенные растения, голосеменные растения, в том числе хвойные, и покрытосеменные, содержащие все плодоносящие и цветущие деревья. Никаких свободоспоривых деревьев вроде Археоптерис существуют в сохранившейся флоре. Долгое время считалось, что покрытосеменные произошли из голосеменных, но недавние молекулярные данные свидетельствуют о том, что их живые представители образуют две отдельные группы.[104][105][106] Молекулярные данные еще не полностью согласованы с морфологическими данными,[107][108][109] но становится общепризнанным, что морфологическая поддержка парафилии не особенно сильна.[110]Это привело бы к заключению, что обе группы возникли внутри птеридоспермов, вероятно, еще в Пермский период.[110]
Покрытосеменные и их предки играли очень небольшую роль, пока в меловом периоде они не расширились. Они зародились как маленькие, любящие сырость организмы в подлеске и с середины[требуется проверка ]-Меловой, чтобы стать доминирующим членом не-северный леса сегодня.
Семена


Ранние наземные растения воспроизводились подобно папоротникам: споры прорастали в небольшие гаметофиты, которые давали яйца и / или сперму. Эти сперматозоиды будут плавать по влажной почве, чтобы найти женские органы (архегонии) на том же или другом гаметофите, где они сливаются с яйцеклеткой, чтобы произвести эмбрион, который прорастет в спорофит.[92]
Гетероспорические растения, как следует из названия, несут споры двух размеров - микроспоры и мегаспоры. Они прорастут с образованием микрогаметофитов и мегагаметофитов соответственно. Эта система проложила путь семяпочкам и семенам: в крайнем случае мегаспорангии могли нести только одну тетраду мегаспор, а для завершения перехода к истинным семяпочкам можно было прервать три мегаспоры в исходной тетраде, оставив одну мегаспору на каждую. мегаспорангий.
Переход к семяпочкам продолжался, и эта мегаспора была «закрыта» спорангием, пока он прорастал. Затем мегагаметофит содержался в водонепроницаемом покрове, который заключал семя. Пыльцевое зерно, которое содержало микрогаметофит, проросший из микроспоры, использовалось для распространения мужской гаметы, высвобождая ее предрасположенные к высыханию жгутиковые сперматозоиды, только когда они достигли рецептивного мегагаметофита.[6]
Ликоподы и сфенопсиды прошли путь к семенной привычке, даже не переступив порог. Известны мегаспоры ископаемых ликопод, достигающие 1 см в диаметре и окруженные вегетативной тканью (Lepidocarpon, Achlamydocarpon); они даже проросли в мегагаметофит. на месте. Однако они не были семяпочками, поскольку нуцеллус, внутренний слой, покрывающий споры, не полностью покрывает споры. Остается очень маленькая щель (микропиле), что означает, что мегаспорангий все еще подвергается воздействию атмосферы. Это имеет два последствия: во-первых, это означает, что она не полностью устойчива к высыханию, а во-вторых, сперматозоидам не нужно «зарываться», чтобы получить доступ к архегониям мегаспоры.[6]
Средний Девонский предшественник к семенным растениям из Бельгии был идентифицирован до самых ранних семенные растения примерно на 20 миллионов лет. Runcaria, мелкая и радиально-симметричная, представляет собой покровный мегаспорангий окружен купулой. В мегаспорангий несет неоткрытый дистальный удлинитель, выступающий над многодольчатым покров. Предполагается, что расширение было связано с анемофильным опыление. Runcaria проливает новый свет на последовательность приобретения персонажа, ведущую к семени. Runcaria обладает всеми качествами семенных растений, кроме твердого семенная оболочка и система для направления пыльцы к семяпочке.[111]
Первые сперматофиты (буквально: «семенные растения») - то есть первые растения, дающие настоящие семена - называются птеридоспермия: буквально «семенные папоротники», названные так потому, что их листва состояла из папоротниковых листьев, хотя они не были тесно связаны с папоротниками. Самые старые ископаемые свидетельства семенных растений относятся к позднему девонскому периоду и, по-видимому, произошли от более ранней группы, известной как прогимноспермные растения. Эти ранние семенные растения варьировались от деревьев до небольших беспорядочных кустов; как и большинство ранних прогимноспермов, это были древесные растения с папоротниковой листвой. Все они имели семяпочки, но не имели шишек, плодов или чего-то подобного. Хотя раннюю эволюцию семян сложно отследить, происхождение семенных папоротников можно проследить от простых тримерофитов до гомоспоровых. Аневрофиты.[6]
Эта модель семян характерна практически для всех голосеменных растений (буквально: «голые семена»), большинство из которых покрывают свои семена древесным конусом или мясистой оболочкой ( тис, например), но ни один из них не покрывает полностью свои семена. Покрытосеменные («сосудистые семена») - единственная группа, которая полностью окружает семя в плодолистнике.
Полностью закрытые семена открыли для растений новый путь: покой семян. Эмбрион, полностью изолированный от внешней атмосферы и, следовательно, защищенный от высыхания, мог пережить несколько лет засухи до прорастания. Было обнаружено, что семена голосеменных из позднего карбона содержат зародыши, что предполагает длительный промежуток между оплодотворением и прорастанием.[112] Этот период связан с вступлением в тепличная земля период, с соответствующим увеличением засушливости. Это говорит о том, что покой возник как реакция на более засушливые климатические условия, когда стало выгодно дождаться влажного периода перед прорастанием.[112] Этот эволюционный прорыв, похоже, открыл шлюзы: прежде негостеприимные места, такие как сухие горные склоны, теперь можно было терпеть, и вскоре они были покрыты деревьями.[112]
Семена давали своим носителям дополнительные преимущества: они увеличивали успешность оплодотворенных гаметофитов, а поскольку запас питательных веществ мог быть «упакован» вместе с зародышем, семена могли быстро прорастать в неблагоприятных условиях, достигая размера, в котором он мог сам себя позаботиться. быстрее.[92] Например, без эндосперма у сеянцев, растущих в засушливых средах, не было бы резервов, чтобы отрастить достаточно глубокие корни, чтобы достичь уровня грунтовых вод до того, как они истощатся от обезвоживания.[92] Точно так же семена, прорастающие в мрачном подлеске, требуют дополнительного запаса энергии, чтобы быстро вырасти достаточно высоко, чтобы улавливать достаточно света для самообеспечения.[92]Комбинация этих преимуществ дала семенным растениям экологическое преимущество перед доминирующим ранее родом. Археоптерис, тем самым увеличивая биоразнообразие ранних лесов.[92]
Несмотря на эти преимущества, оплодотворенные семяпочки часто не созревают в качестве семян.[113] Также во время покоя семян (часто связанного с непредсказуемыми и стрессовыми условиями) накапливаются повреждения ДНК.[114][115][116] Таким образом, повреждение ДНК, по-видимому, является основной проблемой для выживания семенных растений, так же как повреждение ДНК - это серьезная проблема. Главная проблема для жизни в общем.[117]
Цветы

Цветы - это видоизмененные листья, которыми обладают только покрытосеменные, которые относительно поздно появляются в летописи окаменелостей. Группа возникла и разнообразилась в раннем меловом периоде и впоследствии стала экологически значимой.[118] Структуры, похожие на цветы, впервые появляются в ископаемое регистрирует около 130 млн лет назад, в Меловой.[119] Однако в 2018 году ученые сообщили о находке окаменелости. цветок примерно 180 миллионов лет назад, на 50 миллионов лет раньше, чем предполагалось ранее.[120] Интерпретация была однако весьма спорной.[121]
Цветные и / или острые структуры окружают шишки таких растений, как саговники и Gnetales, делая строгое определение термина «цветок» неуловимым.[109]
Основная функция цветка - воспроизведение, который до эволюции цветка и покрытосеменные, была работа микроспорофиллов и мегаспорофиллов. Цветок можно считать мощным эволюционным инновации, потому что его присутствие позволило растительному миру получить доступ к новым средствам и механизмам воспроизводства.
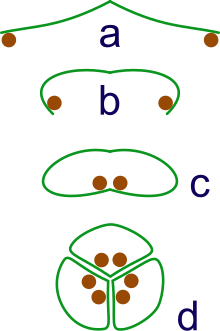
а: спорангии переносятся на кончиках листьев
b: Лист скручивается, чтобы защитить спорангии
c: листья скручиваются в свернутый рулон
d: объединение трех рулонов в синкарп
Долгое время считалось, что цветковые растения произошли изнутри голосеменные; согласно традиционной морфологической точке зрения, они тесно связаны с Gnetales. Однако, как отмечалось выше, недавние молекулярные данные расходятся с этой гипотезой.[105][106] и далее предполагает, что Gnetales более тесно связаны с некоторыми группами голосеменных, чем покрытосеменные,[104] и это сохранившийся голосеменные образуют отдельную кладу с покрытосеменными,[104][105][106] две клады расходятся 300 миллион лет назад.[122]
Отношения стволовые группы к покрытосеменные важен для определения эволюции цветов. Группы стержней дают представление о состоянии более ранних «вилок» на пути к текущему состоянию.Сходимость увеличивает риск неправильной идентификации стволовых групп. Поскольку защита мегагаметофит является эволюционно желательным, вероятно, многие отдельные группы независимо развили защитные оболочки. У цветов эта защита принимает форму карпель, эволюционировали из листа и выполняли защитную роль, защищая семяпочки. Эти семяпочки дополнительно защищены двустенной покров.
Проникновение этих защитных слоев требует чего-то большего, чем свободно плавающее микрогаметофит. Покрытосеменные имеют пыльцевые зерна, состоящие всего из трех клеток. Одна клетка отвечает за просверливание покровов и создание канала, по которому две сперматозоиды стекают вниз. У мегагаметофита всего семь клеток; из них один сливается со сперматозоидом, образуя ядро самой яйцеклетки, а другой соединяется с другим сперматозоидом и посвящает себя формированию богатого питательными веществами эндосперм. Остальные ячейки выполняют вспомогательные роли.[требуется разъяснение ] Этот процесс "двойное оплодотворение "является уникальным и общим для всех покрытосеменных растений.

В летописи окаменелостей есть три интригующие группы, которые имеют структуры, похожие на цветы. Первый - это Пермский период птеридосперм Глоссоптерис, на которых уже были загнутые листья, напоминающие плодолистики. В Мезозойский Caytonia еще больше похож на цветок, с закрытыми семяпочками, но только с одним покровом. Кроме того, детали их пыльцы и тычинок отличают их от настоящих цветковых растений.
В Bennettitales имел удивительно похожие на цветы органы, защищенные завитками прицветники которые, возможно, играли ту же роль, что и лепестки и чашелистики настоящих цветов; однако эти похожие на цветы структуры развивались независимо, поскольку Bennettitales более тесно связаны с саговники и гинкго чем покрытосеменным.[123]
Однако настоящих цветов нет ни в каких группах, кроме существующих сегодня. Место большинства морфологических и молекулярных анализов Амборелла, то нимфеи и Austrobaileyaceae в базальной кладе под названием «АНА». Эта клада, по-видимому, разошлась в раннем меловом периоде около 130 миллион лет назад - примерно в то же время, что и самые ранние ископаемые покрытосеменные,[124][125] и сразу после первая пыльца покрытосеменных, 136 миллионов лет назад.[110]В магнолииды вскоре после этого разошлись, и быстрое облучение произвело эвдикоты и однодольные 125 миллион лет назад.[110] К концу мелового периода 66 миллион лет назад, более 50% сегодняшних отрядов покрытосеменных эволюционировали, а клады составляют 70% глобальных видов.[126]Примерно в это же время цветущие деревья стали доминировать над хвойные породы.[6]:498
Характеристики базальных групп «ANA» позволяют предположить, что покрытосеменные произошли из темных, влажных, часто нарушаемых территорий.[127] Похоже, что покрытосеменные оставались ограниченными такими местообитаниями на протяжении всего мелового периода, занимая нишу небольших трав в начале сукцессионного ряда.[126] Возможно, это ограничивало их первоначальное значение, но давало им гибкость, объясняющую скорость их последующего разнообразия в других средах обитания.[127]
| Традиционный вид | |||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
| Современный вид | ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
Некоторые предполагают, что покрытосеменные произошли от неизвестного семенного папоротника, птеридофита, и рассматривают саговники как живые семенные папоротники с семеносными и стерильными листьями (Cycas revoluta).[108]
В августе 2017 года ученые представили подробное описание и 3D-изображение реконструкции, возможно, первого цветок который жил около 140 миллионов лет назад.[128][129]
Происхождение цветка

Семья Amborellaceae считается сестрой клады ко всем остальным живым цветущим растениям. Проект генома Amborella trichopoda была опубликована в декабре 2013 г.[130] Сравнивая его геном с геномами всех других живых цветковых растений, можно будет определить наиболее вероятные характеристики предка A. trichopoda и все другие цветковые растения, то есть предковые цветковые растения.[131]
Кажется, что на уровне органа лист может быть предком цветка или хотя бы некоторых цветочных органов. Когда некоторые важные гены, участвующие в развитии цветов, мутировавший на месте цветов возникают скопления листовидных структур. Таким образом, когда-то в истории программа развития, ведущая к формированию листа, должна была быть изменена, чтобы произвести цветок. Вероятно, также существует общая прочная структура, в которой было создано цветочное разнообразие. Примером этого является ген под названием ЛИФИ (LFY), который участвует в развитии цветов у Arabidopsis thaliana. В гомологи этого гена находятся в покрытосеменные столь же разнообразный, как помидор, львиный зев, горох, кукуруза и даже голосеменные. Выражение Arabidopsis thaliana LFY у далеких растений, таких как тополь и цитрусовые также приводит к цветению этих растений. В LFY ген регулирует экспрессию некоторых генов, принадлежащих к MADS-коробка семья. Эти гены, в свою очередь, действуют как прямые регуляторы развития цветов.[нужна цитата ]
Эволюция семейства MADS-box
Члены MADS-коробка Семейство факторов транскрипции играет очень важную и эволюционно законсервированную роль в развитии цветов. Согласно ABC Модель развития цветка, три зоны - A, B и C - образуются в развивающемся зачатке цветка под действием некоторых факторы транскрипции, которые являются членами MADS-коробка семья. Среди них функции генов B- и C-доменов были эволюционно более консервативными, чем гена A-домена. Многие из этих генов возникли благодаря дупликации генов предков членов этой семьи. Многие из них обладают избыточными функциями.
Эволюция MADS-коробка семья была широко изучена. Эти гены присутствуют даже в птеридофиты, но разброс и разнообразие во много раз выше покрытосеменные.[132] Похоже, что существует довольно много закономерностей в том, как развивалась эта семья. Рассмотрим эволюцию гена C-региона АГАМУСНЫЙ (AG). В сегодняшних цветах это выражается в тычинки, а карпель, которые являются репродуктивными органами. Его предок в голосеменные также имеет тот же образец выражения. Здесь это выражается в стробили, орган, производящий пыльца или семяпочки.[133] Точно так же B-гены ' (AP3 и PI) предки выражаются только в мужских органах в голосеменные. Их потомки у современных покрытосеменных также выражены только в тычинки, мужской репродуктивный орган. Таким образом, те же самые существующие компоненты были использованы растениями новым способом для создания первого цветка. Это повторяющийся образец в эволюция.
Факторы, влияющие на цветочное разнообразие
Цветочная структура растений сильно различается, как правило, из-за изменений MADS-коробка гены и характер их экспрессии. Например, травы обладают уникальной цветочной структурой. Плодолистики и тычинки окружены чешуевидными лодикулы и два прицветника, лемма и палеа, но генетические данные и морфология предполагают, что lodicules гомологичны эвдикот лепестки.[134] Палеа и лемма могут быть гомологами чашелистиков других групп или могут быть уникальными травяными структурами.[нужна цитата ]
Другой пример - это Linaria vulgaris, который имеет два вида симметрии цветов:радиальный и двусторонний. Эти симметрии обусловлены эпигенетический изменения только в одном гене, называемом ЦИКЛОИДЕЯ.[119]

Arabidopsis thaliana имеет ген под названием АГАМУСНЫЙ это играет важную роль в определении количества лепестки и чашелистики и другие органы образуются. Мутации в этом гене приводят к появлению цветочных меристема получение неопределенной судьбы и разрастание цветочных органов в махровый формы розы, гвоздики и утренняя слава. Эти фенотипы были выбраны садоводами из-за возросшего количества лепестки.[135] Несколько исследований различных растений, таких как петуния, помидор, Нетерпеливы, кукуруза и др. предположили, что огромное разнообразие цветов является результатом небольших изменений в гены контролируя их развитие.[136]
В Цветочный геномный проект подтвердил, что ABC Модель развития цветка не сохраняется во всех покрытосеменные. Иногда домены выражения меняются, как в случае многих однодольные, а также у некоторых базальных покрытосеменных, таких как Амборелла. Различные модели развития цветов, такие как Модель исчезающих границ, или Модель перекрывающихся границ которые предлагают нежесткие области выражения, могут объяснить эти архитектуры.[137] Есть вероятность, что от базальных до современных покрытосеменных, области цветочной архитектуры становились все более и более фиксированными в процессе эволюции.
Время цветения
Еще одна цветочная особенность, о которой говорили естественный отбор время цветения. Некоторые растения зацветают в начале своего жизненного цикла, другим требуется период яровизация до цветения. Этот результат основан на таких факторах, как температура, Интенсивность света, присутствие опылители и другие сигналы окружающей среды: гены, подобные КОНСТАНС (КО), Цветение Locus C (FLC) и ФРИГИДА регулируют интеграцию сигналов окружающей среды в путь развития цветов. Вариации в этих локусах были связаны с вариациями времени цветения между растениями. Например, Arabidopsis thaliana экотипы, растущие на морозе, умеренный регионы требуют длительной яровизации перед цветением, в то время как тропический разновидностей и наиболее распространенных лабораторных штаммов нет. Это изменение связано с мутациями в FLC и ФРИГИДА гены, что делает их нефункциональными.[138]
Многие из генов, участвующих в этом процессе, сохраняются на всех изученных растениях. Однако иногда, несмотря на генетическую консервацию, механизм действия оказывается другим. Например, рис растение короткого дня, а Arabidopsis thaliana растение долгого дня. Оба растения содержат белки. CO и ЦВЕТУЩИЙ ЛОКУС T (FT), но в Arabidopsis thaliana, CO усиливает FT производство, в то время как в рисе CO гомолог подавляет FT производства, что приводит к полностью противоположным эффектам на переработке.[139]
Теории эволюции цветов
В Теория антофитов было основано на наблюдении, что группа голосеменных Gnetales имеет подобный цветку яйцеклетка. Частично развился сосуды как найдено в покрытосеменные, а мегаспорангий покрыт тремя конвертами, как и яичник строение цветков покрытосеменных. Однако многие другие доказательства показывают, что Gnetales не имеют отношения к покрытосеменным.[123]
В В основном мужская теория имеет более генетическую основу. Сторонники этой теории указывают, что у голосеменных есть две очень похожие копии гена LFY, а у покрытосеменных только один. Молекулярные часы анализ показал, что другой LFY paralog был утерян у покрытосеменных примерно в то же время, когда окаменелости цветов стали многочисленными, что позволяет предположить, что это событие могло привести к эволюции цветов.[140] Согласно этой теории, потеря одного из LFY паралог привели к цветам, которые были более мужскими, с семяпочки выражается эктопически. Эти семяпочки изначально выполняли функцию привлечения опылители, но когда-нибудь позже, возможно, был интегрирован в основной цветок.
Механизмы и участники эволюции морфологии растений

Хотя факторы окружающей среды в значительной степени ответственны за эволюционные изменения, они действуют просто как агенты естественный отбор. Изменения по своей сути вызываются явлениями на генетическом уровне: мутации, хромосомные перестройки и эпигенетический изменения. Хотя общие типы мутации справедливо для живого мира, в растениях некоторые другие механизмы были задействованы как очень важные.
Удвоение генома является относительно обычным явлением в эволюции растений и приводит к полиплоидия, что, следовательно, является обычным явлением для растений. Подсчитано, что по крайней мере у половины (а возможно, у всех) растений удваивались геномы за свою историю. Удвоение генома влечет за собой дупликация гена, таким образом создавая функциональную избыточность в большинстве генов. Дублированные гены могут обрести новую функцию либо за счет изменения паттерна экспрессии, либо за счет изменения активности. Полиплоидия и дупликация генов считаются одними из самых мощных сил в эволюции формы растений; хотя не известно почему геном удвоение - такой частый процесс у растений. Одна из вероятных причин - производство большого количества вторичные метаболиты в клетках растений. Некоторые из них могут вмешиваться в нормальный процесс хромосомной сегрегации, вызывая дупликацию генома.

В последнее время было показано, что растения обладают значительными микроРНК семейства, которые сохраняются во многих линиях растений. По сравнению с животные, в то время как количество семейств miRNA растений меньше, чем у животных, размер каждой семьи намного больше. В miRNA гены также гораздо более распространены в геноме, чем у животных, где они более сгруппированы. Было высказано предположение, что эти семейства miRNA расширились за счет дупликаций хромосомных областей.[141] Многие гены miRNA участвуют в регуляции развитие растений было обнаружено, что они довольно консервативны между изученными растениями.
Одомашнивание таких растений, как кукуруза, рис, ячмень, пшеница и т. д. также была значительной движущей силой в их развитии. Исследования происхождения кукурузы показали, что это одомашненное производное дикого растения из Мексика называется Теосинте. Теозинте принадлежит к род Зеа, как кукуруза, но плодоносит очень мелко соцветие, 5–10 твердых початков и сильно разветвленный и раскидистый стебель.

При скрещивании определенного сорта теозинте и кукурузы получается плодовитое потомство, которое занимает промежуточное положение в фенотип между кукурузой и теосинте. QTL анализ также выявил некоторые локусы, которые при мутации у кукурузы дают теозинте-подобные стебли или теозинте-подобные початки. Молекулярные часы Анализ этих генов оценивает их происхождение примерно 9000 лет назад, что хорошо согласуется с другими записями об одомашнивании кукурузы. Считается, что небольшая группа фермеров должна была выбрать какой-то похожий на кукурузу естественный мутант теозинте около 9000 лет назад в Мексике и подвергнуть его непрерывному отбору, чтобы получить знакомое сегодня растение кукурузы.[142]
Съедобная цветная капуста - это одомашненная версия дикого растения. Brassica oleracea, не обладающий плотной недифференцированной соцветие Творог, которым обладает цветная капуста.
Цветная капуста обладает единственной мутацией в гене, называемом CAL, контролируя меристема дифференциация на соцветие. Это приводит к тому, что клетки цветочной меристемы приобретают недифференцированную идентичность и вместо того, чтобы расти в цветок, они превращаются в плотную массу клеток меристемы соцветий с задержкой развития.[143] Эта мутация была выбрана путем одомашнивания, по крайней мере, со времен Греческий империя.
Эволюция фотосинтетических путей

В C4 метаболический путь является ценным недавним эволюционным нововведением у растений, включающим сложный набор адаптивных изменений в физиология и экспрессия гена узоры.[144]
Фотосинтез представляет собой сложный химический путь, которому способствует ряд ферменты и коферменты. В фермент RuBisCO отвечает за "фиксацию" CO
2 - то есть он присоединяет его к молекуле на основе углерода с образованием сахара, который может использоваться растением, высвобождая молекулу кислорода. Однако этот фермент, как известно, неэффективен, и по мере повышения температуры окружающей среды он будет все больше связывать кислород вместо CO
2 в процессе, называемом фотодыхание. Это энергетически затратно, так как растение должно использовать энергию, чтобы превратить продукты фотодыхания обратно в форму, которая может реагировать с CO
2.
Концентрирующий углерод
C4 растения развили механизмы концентрации углерода, которые работают за счет увеличения концентрации CO
2 вокруг RuBisCO и исключая кислород, тем самым повышая эффективность фотосинтеза за счет уменьшения фотодыхания. Процесс концентрации CO
2 вокруг RuBisCO требует больше энергии, чем позволяет газам размытый, но при определенных условиях - например, теплые температуры (> 25 ° C), низкие CO
2 концентрации или высокой концентрации кислорода - окупается за счет уменьшения потерь сахаров из-за фотодыхания.
Один тип C4 метаболизм использует так называемый Анатомия Кранца. Это транспортирует CO
2 через внешний слой мезофилла, через ряд органических молекул, к центральным клеткам оболочки пучка, где CO
2 выпущен. В этом случае, CO
2 сконцентрирована недалеко от места работы RuBisCO. Потому что RuBisCO работает в среде с гораздо большим CO
2 чем в противном случае, он работает более эффективно.
Второй механизм, CAM фотосинтез, временно отделяет фотосинтез от действия RuBisCO. RuBisCO работает только в дневное время, когда устьица закрыты и CO
2 обеспечивается распадом химического малат. Больше CO
2 затем собирают из атмосферы, когда устьица открываются, в прохладные влажные ночи, что снижает потерю воды.
Эволюционный рекорд
Эти два пути с одинаковым влиянием на RuBisCO развивались несколько раз независимо - действительно, C4 одна возникла 62 раза на 18 разных заводах семьи. Ряд «преадаптаций», по-видимому, проложили путь для C4, что привело к его кластеризации в определенные клады: он чаще всего вводился в растения, у которых уже были такие особенности, как обширная ткань оболочки сосудистого пучка.[145] Многие потенциальные пути эволюции, приводящие к C4 фенотип возможны и были охарактеризованы с помощью Байесовский вывод,[144] подтверждая, что нефотосинтетические адаптации часто являются ступеньками эволюции для дальнейшей эволюции C4.
C4 конструкция используется подмножеством трав, в то время как CAM используется многими суккулентами и кактусы. C4 черта, похоже, возникла во время Олигоцен, около От 25 до 32 миллион лет назад;[146] однако они не стали экологически значимыми до Миоцен, С 6 до 7 миллион лет назад.[147] Примечательно, что некоторые окаменелости, превращенные в уголь, сохраняют ткань, организованную в анатомию Кранца, с неповрежденными клетками оболочки пучка,[148] позволяя присутствие C4 метаболизм предстоит определить. Изотопные маркеры используются для определения их распределения и значимости.3 растения предпочитают зажигалку из двух изотопы углерода в атмосфере, 12C, который более активно участвует в химических путях его фиксации. Потому что C4 метаболизм включает следующую химическую стадию, этот эффект усиливается. Растительный материал может быть проанализированы вывести соотношение более тяжелых 13C к 12C. Это соотношение обозначается δ13C. C3 растения в среднем примерно на 14 ‰ (частей на тысячу) легче атмосферного, в то время как C4 растения примерно на 28 ‰ легче. В δ13C CAM-растений зависит от процентного содержания углерода, зафиксированного ночью, по сравнению с тем, что зафиксировано днем, будучи ближе к C3 растения, если они фиксируют большую часть углерода днем и ближе к C4 растения, если они ночью фиксируют весь свой углерод.[149]
Первоначального ископаемого материала в достаточном количестве для анализа самой травы мало, но лошади являются хорошим показателем. В интересующий период они были широко распространены во всем мире и выращивались почти исключительно на травах. В изотопной палеонтологии есть старая фраза: «Вы то, что вы едите (плюс немного)» - это относится к тому факту, что организмы отражают изотопный состав всего, что они едят, плюс небольшой поправочный коэффициент. По всему земному шару имеются хорошие данные о конских зубах, а их δ13C запись показывает резкий отрицательный перегиб вокруг С 6 до 7 миллион лет назад, во время мессинизма, который интерпретируется как результат подъема C4 растения в мировом масштабе.[147]
Когда C4 Преимущество?
Хотя C4 повышает эффективность RuBisCO, концентрация углерода очень энергоемкая. Это означает, что C4 растения имеют преимущество только перед C3 организмов в определенных условиях: а именно, при высоких температурах и малом количестве осадков. C4 растения также нуждаются в большом количестве солнечного света, чтобы процветать.[150] Модели предполагают, что без лесных пожаров, удаляющих тени деревьев и кустарников, не было бы места для C4 растения.[151] Но лесные пожары происходили 400 миллионов лет назад - почему C4 так долго возникать, а потом столько раз появляться независимо? Каменноугольный период (~300 миллион лет назад) имели заведомо высокий уровень кислорода - почти достаточный, чтобы позволить случайное возгорание[152] - и очень низкий CO
2, но нет C4 должна быть найдена изотопная подпись. И, кажется, нет внезапного триггера для подъема миоцена.
В миоцене атмосфера и климат были относительно стабильными. Если что-нибудь, CO
2 постепенно увеличивался с 14–9 миллион лет назад до оседания до концентраций, аналогичных голоцену.[153] Это говорит о том, что он не играл ключевой роли в вызове C4 эволюция.[146] Сами травы (группа, которая дала бы начало наибольшему количеству встречаемости C4), вероятно, существовал около 60 миллионов лет или более, поэтому у него было достаточно времени, чтобы развить C4,[154][155] которые, в любом случае, присутствуют в различных группах и, таким образом, развивались независимо. Имеется сильный сигнал об изменении климата в Южной Азии;[146] усиление засушливости - следовательно, увеличение частоты и интенсивности пожаров - могло привести к повышению важности пастбищ.[156] Однако это трудно согласовать с североамериканскими рекордами.[146] Возможно, что сигнал полностью биологический, вызванный огнем (и слоном?) -[157] обусловленное ускорением эволюции травы - что, как за счет увеличения выветривания, так и за счет включения большего количества углерода в отложения, уменьшало атмосферный CO
2 уровни.[157] Наконец, есть свидетельства того, что начало C4 от С 9 до 7 миллион лет назад это смещенный сигнал, справедливый только для Северной Америки, откуда происходит большинство образцов; Новые данные свидетельствуют о том, что пастбища в Южной Америке превратились в доминирующее состояние как минимум 15 млн лет назад.
Эволюция регуляции транскрипции
Факторы транскрипции и сети регуляции транскрипции играют ключевую роль в развитии растений и ответах на стресс, а также в их эволюции. Во время посадки растений появилось много новых семейств факторов транскрипции, которые преимущественно связаны с сетями многоклеточного развития, репродукции и развития органов, внося вклад в более сложный морфогенез наземных растений.[158]
Эволюция вторичного метаболизма

Вторичные метаболиты существенно низкие молекулярный вес соединения, иногда имеющие сложную структуру, которые не являются необходимыми для нормальных процессов рост, развитие, или воспроизведение. Они участвуют в столь разнообразных процессах, как иммунитет, против травоядных, опылитель Привлечение, коммуникация между растениями, поддерживая симбиотический ассоциации с почвенной флорой или повышение скорости оплодотворение, и, следовательно, важны с точки зрения evo -dev. Вторичные метаболиты структурно и функционально разнообразны, и, по оценкам, сотни тысяч ферментов могут быть вовлечены в процесс их производства, причем около 15-25% генома кодирует эти ферменты, и каждый вид имеет свой уникальный арсенал вторичные метаболиты.[159] Многие из этих метаболитов, такие как салициловая кислота имеют медицинское значение для человека.
Цель производства такого количества вторичных метаболитов, при этом значительная часть метаболом посвящен этой деятельности неясно. Предполагается, что большинство этих химикатов помогают в создании иммунитета, и, как следствие, разнообразие этих метаболитов является результатом постоянного гонка вооружений между растениями и их паразитами. Некоторые доказательства подтверждают этот случай. Центральный вопрос связан с репродуктивными затратами на поддержание такого большого количества генов, предназначенных для производства вторичных метаболитов. Были предложены различные модели, исследующие этот аспект вопроса, но консенсус по поводу размера затрат еще не достигнут;[160] поскольку все еще трудно предсказать, увеличивает ли растение с большим количеством вторичных метаболитов свою выживаемость или репродуктивный успех по сравнению с другими растениями в его окрестностях.
Производство вторичных метаболитов, по-видимому, возникло довольно рано в ходе эволюции. Похоже, что у растений они распространились с помощью механизмов, включая дупликацию генов или эволюцию новых генов. Кроме того, исследования показали, что разнообразие некоторых из этих соединений может быть положительным. Хотя роль эволюции новых генов в эволюции вторичного метаболизма очевидна, есть несколько примеров, когда новые метаболиты образовывались в результате небольших изменений в реакции. Например, цианогликозиды Было высказано предположение, что они эволюционировали несколько раз в разных линиях растений. Есть несколько таких примеров конвергентная эволюция. Например, ферменты для синтеза лимонен - а терпен - более похожи между покрытосеменными и голосеменными, чем с собственными ферментами синтеза терпена. Это предполагает независимую эволюцию пути биосинтеза лимонена в этих двух ветвях.[161]
Эволюция взаимодействия растений и микробов

Происхождение микробов на Земле, восходящее к началу жизни более 3,5 миллиардов лет назад, указывает на то, что микробно-микробные взаимодействия постоянно развивались и диверсифицировались с течением времени, задолго до того, как растения начали колонизировать землю 450 миллионов лет назад. Таким образом, вполне вероятно, что межмикробные взаимодействия как внутри царства, так и между ними представляют собой сильные движущие силы установления растительно-ассоциированных микробные консорциумы на границе почва-корень. Тем не менее, остается неясным, в какой степени эти взаимодействия в ризосфере / филлосфере и в эндофитных компартментах растений (т. Е. Внутри хозяина) формируют микробные сообщества в природе и влияет ли адаптация микробов к средам обитания растений на стратегии взаимодействия микробов с конкретными местами обитания, которые влияют на Фитнес растений. Более того, вклад конкурентных и совместных взаимодействий микробов-микробов в общую структуру сообщества по-прежнему трудно оценить в природе из-за сильного шума окружающей среды.[162]
Коэволюция растений и грибковых паразитов
Дополнительным фактором, способствующим эволюционным изменениям некоторых растений, является сила, обусловленная совместной эволюцией с грибковыми паразитами. В среде с грибковым паразитом, который является обычным явлением в природе, растения должны адаптироваться, чтобы избежать вредного воздействия паразита.[165]
Всякий раз, когда паразитический гриб перекачивает ограниченные ресурсы от растения, возникает селективное давление на фенотип, который лучше предотвращает паразитарную атаку грибов. В то же время грибы, которые лучше приспособлены для обхода защитных сил растения, будут иметь более высокий уровень физической подготовки. Сочетание этих двух факторов приводит к бесконечному циклу эволюционных изменений в системе хозяин-патоген.[166]
Поскольку каждый вид во взаимоотношениях находится под влиянием постоянно меняющегося симбионта, эволюционные изменения обычно происходят более быстрыми темпами, чем если бы другой вид отсутствовал. Это верно для большинства случаев коэволюции. Это делает способность популяции быстро развиваться жизненно важной для ее выживания. Кроме того, если патогенные виды слишком успешны и угрожают выживанию и репродуктивному успеху растений-хозяев, патогенные грибы рискуют потерять свой источник питательных веществ для будущих поколений. Эти факторы создают динамику, которая формирует эволюционные изменения у обоих видов поколение за поколением.[166]
Гены, кодирующие защитные механизмы растений, должны постоянно меняться, чтобы не отставать от паразита, который постоянно пытается уклониться от защиты. Гены, кодирующие механизмы прикрепления, являются наиболее динамичными и напрямую связаны со способностью грибов уклоняться.[167] Чем больше изменений в этих генах, тем сильнее меняется механизм прикрепления. После селективных воздействий на результирующие фенотипы происходит эволюционное изменение, которое способствует уклонению от защиты хозяина.
Грибы не только эволюционируют, чтобы избежать защиты растений, но они также пытаются помешать растению задействовать механизмы, улучшающие свою защиту. Все, что грибы могут сделать, чтобы замедлить процесс эволюции растений-хозяев, улучшит приспособленность будущих поколений, потому что растение не сможет идти в ногу с эволюционными изменениями паразита. Одним из основных процессов, посредством которых растения быстро развиваются в ответ на окружающую среду, является половое размножение. Без полового размножения полезные черты не могли бы распространяться в популяции растений так быстро, что позволило бы грибам получить конкурентное преимущество. По этой причине половые репродуктивные органы растений становятся мишенями для поражения грибов. Исследования показали, что многие различные современные виды облигатных паразитических грибов растений разработали механизмы для отключения или иного воздействия на половое размножение растений. В случае успеха процесс полового размножения замедляется для растения, замедляя тем самым эволюционные изменения или, в крайних случаях, грибы могут сделать растение стерильным, создавая преимущество для патогенов. Точно неизвестно, как этот адаптивный признак развился у грибов, но ясно, что родство с растением привело к развитию этого процесса.[168]
Некоторые исследователи также изучают, как ряд факторов влияет на скорость эволюционных изменений и результаты изменений в различных средах. Например, как и в большинстве случаев эволюции, увеличение наследуемости в популяции дает возможность большей эволюционной реакции в присутствии давления отбора. По признакам, характерным для совместной эволюции растений и грибов, исследователи изучили, как вирулентность вторгающегося патогена влияет на коэволюцию. Исследования с участием Mycosphaerella graminicola последовательно показали, что вирулентность патогена не оказывает значительного влияния на эволюционный путь растения-хозяина.[169]
Могут быть и другие факторы, которые могут повлиять на процесс коэволюции. Например, в небольших популяциях отбор является относительно более слабой силой для популяции из-за генетический дрейф. Генетический дрейф увеличивает вероятность наличия фиксированных аллелей, что снижает генетическую изменчивость в популяции. Следовательно, если на территории есть только небольшая популяция растений, способных к совместному размножению, генетический дрейф может противодействовать эффектам отбора, ставящим растение в невыгодное положение по отношению к грибам, которые могут развиваться с нормальной скоростью. Различия в популяции хозяев и патогенов - главный фактор, определяющий успех эволюции по сравнению с другими видами. Чем больше генетическая изменчивость, тем быстрее вид может развиваться, чтобы противодействовать избеганию или защитным механизмам другого организма.[165]
Из-за процесса опыления растений эффективный размер популяции обычно больше, чем у грибов, потому что опылители могут связывать изолированные популяции так, как гриб не может. Это означает, что положительные черты, которые развиваются в несмежных, но близких областях, могут быть переданы в соседние области. Грибки должны развиваться индивидуально, чтобы уклоняться от защиты хозяина в каждой области. Очевидно, что это явное конкурентное преимущество для растений-хозяев. Половое размножение с широкой и высокой дисперсией населения приводит к быстрым эволюционным изменениям и более высокому репродуктивному успеху потомства.[170]
Условия окружающей среды и климата также играют роль в эволюционных результатах. Исследования на дубах и облигатном грибковом паразите на разных высотах ясно показывают это различие. Для одного и того же вида разные высотные положения имели резко разные темпы эволюции и изменения реакции на патогены из-за организма, а также в селективной среде из-за своего окружения.[171]
Коэволюция - это процесс, связанный с гипотеза красной королевы. И растение-хозяин, и паразитические грибы должны продолжать выживать, чтобы оставаться в своей экологической нише. Если один из двух видов во взаимосвязи развивается значительно быстрее, чем другой, более медленные виды окажутся в невыгодном конкурентном положении и рискуют потерять питательные вещества. Поскольку два вида в системе так тесно связаны, они вместе реагируют на факторы внешней среды, и каждый вид влияет на эволюционный результат другого. Другими словами, каждый вид оказывает избирательное давление на другой. Размер популяции также является важным фактором в результате, потому что различия в потоке генов и генетический дрейф могут вызвать эволюционные изменения, которые не соответствуют направлению отбора, ожидаемому силами, вызванными другим организмом. Коэволюция - важное явление, необходимое для понимания жизненно важных отношений между растениями и их грибковыми паразитами.
Смотрите также
- Растения
- Общая эволюция
- Изучение растений
- Взаимодействие с растениями
- Новые тенденции
использованная литература
- ^ Т. Кавальер Смит 2007, Эволюция и взаимоотношения водорослей основных ветвей древа жизни. из: Распутывание водорослей, Броди и Льюис. CRC Press
- ^ Шевчикова, Тереза; и другие. (2015). «Обновление эволюционных взаимоотношений водорослей посредством секвенирования пластидного генома». Научные отчеты. 5: 10134. Bibcode:2015НатСР ... 510134С. Дои:10.1038 / srep10134. ЧВК 4603697. PMID 26017773.
- ^ Теодор Коул и Хартмут Хильгер 2013 Филогения мохообразных В архиве 2015-11-23 в Wayback Machine
- ^ Теодор Коул и Хартмут Хильгер 2013 Филогения трахеофитов В архиве 2016-03-04 в Wayback Machine
- ^ Теодор Коул и Хартмут Хильгер 2015 Филогения покрытосеменных, систематика цветковых растений. Freie Universität Berlin
- ^ а б c d е ж г час я j k л м Стюарт, W.N .; Ротвелл, Г. (1993). Палеоботаника и эволюция растений (2-е изд.). Издательство Кембриджского университета. ISBN 978-0-521-23315-6.
- ^ а б Strother, Paul K .; Баттисон, Лейла; Brasier, Martin D .; Веллман, Чарльз Х. (2011). «Самые ранние неморские эукариоты на Земле». Природа. 473 (7348): 505–509. Bibcode:2011Натура.473..505S. Дои:10.1038 / природа09943. PMID 21490597. S2CID 4418860.
- ^ а б c Кнаут, Л. Пол; Кеннеди, Мартин Дж. (2009). «Позднедокембрийское озеленение Земли». Природа. 460 (7256): 728–732. Bibcode:2009Натура.460..728K. Дои:10.1038 / природа08213. PMID 19587681. S2CID 4398942.
- ^ Rothwell, G.W .; Scheckler, S.E .; Гиллеспи, В. Х. (1989). "Элькинсия ген. nov., позднедевонское голосеменное растение с куполообразными семяпочками ». Ботанический вестник. 150 (2): 170–189. Дои:10.1086/337763. S2CID 84303226.
- ^ а б П. Кенрик, П. Р. Крейн (1997) (1997). Происхождение и раннее разнообразие наземных растений. Кладистическое исследование. Пресса Смитсоновского института, Вашингтон и Лондон. Вашингтон: Смитсоновский институт. Нажмите. ISBN 978-1-56098-729-1.
- ^ а б c d е ж г час я j k л м п Raven, J.A .; Эдвардс, Д. (2001). «Корни: эволюционное происхождение и биогеохимическое значение». Журнал экспериментальной ботаники. 52 (90001): 381–401. Дои:10.1093 / jexbot / 52.suppl_1.381. PMID 11326045.
- ^ Дель-Бем, Л. Э. (2018). «Эволюция ксилоглюкана и земледелие зеленых растений». Новый Фитолог. 219 (4): 1150–1153. Дои:10.1111 / nph.15191. PMID 29851097.
- ^ Хекман, Д. С .; Гейзер, Д. М .; Eidell, B.R .; Stauffer, R.L .; Kardos, N.L .; Хеджес, С. Б. (август 2001 г.). «Молекулярные доказательства раннего заселения земли грибами и растениями». Наука. 293 (5532): 1129–1133. Дои:10.1126 / science.1061457. ISSN 0036-8075. PMID 11498589. S2CID 10127810.
- ^ Баттисон, Лейла; Бразье, Мартин Д. (август 2009 г.). «Исключительная сохранность ранних наземных сообществ в озерном фосфате один миллиард лет назад» (PDF). В Smith, Martin R .; О'Брайен, Лорна Дж .; Карон, Жан-Бернар (ред.). Абстрактный объем. Международная конференция по кембрийскому взрыву (Уолкотт 2009). Торонто, Онтарио, Канада: Консорциум Burgess Shale (опубликовано 31 июля 2009 г.). ISBN 978-0-9812885-1-2.
- ^ Battistuzzi, F. U .; Feijao, A .; Хеджес, С. Б. (2004). «Геномная шкала времени эволюции прокариот: понимание происхождения метаногенеза, фототрофии и колонизации земли». BMC Эволюционная биология. 4: 44. Дои:10.1186/1471-2148-4-44. ЧВК 533871. PMID 15535883.
- ^ Strother, P.K .; Аль-Хаджри, С .; Траверс, А. (1996). «Новые свидетельства наземных растений из нижнего среднего ордовика Саудовской Аравии». Геология. 24 (1): 55–59. Bibcode:1996Гео .... 24 ... 55S. Дои:10.1130 / 0091-7613 (1996) 024 <0055: NEFLPF> 2.3.CO; 2.
- ^ Rubinstein, C.V .; Gerrienne, P .; de la Puente, G. S .; Astini, R.A .; Стиманс, П. (2010). «Свидетельства раннего среднего ордовика для наземных растений в Аргентине (восточная Гондвана)». Новый Фитолог. 188 (2): 365–369. Дои:10.1111 / j.1469-8137.2010.03433.x. PMID 20731783.
- ^ Wellman, Charles H .; Остерлофф, Питер Л .; Мохиуддин, Узма (2003). «Фрагменты древнейших наземных растений» (PDF). Природа. 425 (6955): 282–285. Bibcode:2003 Натур. 425..282Вт. Дои:10.1038 / природа01884. PMID 13679913. S2CID 4383813.
- ^ Steemans, P .; Лепота К .; Marshallb, C.P .; Le Hérisséc, A .; Javauxa, E.J. (2010). «FTIR-характеристика химического состава силурийских миоспор (криптоспор и трилете спор) из Готланда, Швеция». Обзор палеоботаники и палинологии. 162 (4): 577–590. Дои:10.1016 / j.revpalbo.2010.07.006.
- ^ Kump, L.R .; Павлов, А .; Артур, М.А. (2005). «Массовый выброс сероводорода на поверхность океана и в атмосферу во время периодов океанической аноксии». Геология. 33 (5): 397. Bibcode:2005Гео .... 33..397K. Дои:10.1130 / G21295.1.
- ^ Баттерфилд, Н. Дж. (2009). «Кислород, животные и океаническая вентиляция: альтернативный взгляд». Геобиология. 7 (1): 1–7. Дои:10.1111 / j.1472-4669.2009.00188.x. PMID 19200141.
- ^ Steemans, P .; Herisse, L .; Мелвин, Дж .; Миллер, А .; Paris, F .; Verniers, J .; Веллман, Х. (апрель 2009 г.). «Происхождение и радиация древнейших сосудистых растений суши». Наука. 324 (5925): 353. Bibcode:2009Sci ... 324..353S. Дои:10.1126 / science.1169659. ISSN 0036-8075. PMID 19372423. S2CID 206518080.
- ^ Рубинштейн, Клаудиа; Вайда, Виви (2019). «Балтика - колыбель ранних наземных растений? Самая старая запись о спорах трилете и разнообразных скоплениях криптоспор; свидетельства из ордовикских последовательностей Швеции». GFF (Журнал геологического общества Швеции). 141 (3): 181–190. Дои:10.1080/11035897.2019.1636860.
- ^ а б c Gray, J .; Chaloner, W. G .; Вестолл, Т. С. (1985). «Летопись микрофоссилий ранних наземных растений: достижения в понимании ранней земной жизни, 1970–1984». Философские труды Королевского общества B. 309 (1138): 167–195. Bibcode:1985РСПТБ.309..167Г. Дои:10.1098 / рстб.1985.0077. JSTOR 2396358.
- ^ Томеску, А. М. Ф. (2006). «Водно-болотные угодья до трахеофитов: таллоидные наземные сообщества биоты ручья Раннесилурийского пролива (Вирджиния)». Болота сквозь время (PDF). Дои:10.1130/2006.2399(02). ISBN 9780813723990 http://www.humboldt.edu/biosci/docs/faculty/TomescuRothwell2006.pdf. Получено 2014-05-28. Отсутствует или пусто
| название =(Помогите) - ^ Томеску, А. М. Ф .; Honegger, R .; Ротвелл, Дж. У. (2008). «Самая ранняя летопись окаменелостей бактериально-цианобактериальных консорциумов: ранняя силурийская биота ручья Пассаж (440 млн лет назад, Вирджиния, США)». Геобиология. 6 (2): 120–124. Дои:10.1111 / j.1472-4669.2007.00143.x. PMID 18380874.
- ^ «Первые наземные растения и грибы изменили климат Земли, проложив путь к взрывной эволюции наземных животных, согласно новому исследованию генов». science.psu.edu. Получено 9 апреля 2018.
- ^ Glasspool, IJ; Эдвардс, D; Топор, L (2004). «Древесный уголь в силурии как свидетельство самого раннего лесного пожара». Геология. 32 (5): 381–383. Bibcode:2004Гео .... 32..381Г. Дои:10.1130 / G20363.1.
- ^ Bernstein, H; Байерс, GS; Michod, RE (1981). «Эволюция полового размножения: важность восстановления, дополнения и вариации ДНК». Американский натуралист. 117 (4): 537–549. Дои:10.1086/283734. S2CID 84568130.
- ^ Michod, RE; Гейли, TW (1992). «Маскировка мутаций и эволюция пола». Американский натуралист. 139 (4): 706–734. Дои:10.1086/285354. S2CID 85407883.
- ^ Сёвеньи, Петер; Рикка, Мариана; Хок, Жофия; Шоу, Джонатан А .; Симидзу, Кентаро К. и Вагнер, Андреас (2013). «Селекция на гаплоиде не более эффективна, чем на диплоидных стадиях жизни покрытосеменных и мха». Молекулярная биология и эволюция. 30 (8): 1929–39. Дои:10.1093 / molbev / mst095. PMID 23686659.
- ^ а б c d Бойс, К. К. (2008). "Как зеленый был Cooksonia? Важность размера в понимании ранней эволюции физиологии линии сосудистых растений ". Палеобиология. 34 (2): 179–194. Дои:10.1666 / 0094-8373 (2008) 034 [0179: HGWCTI] 2.0.CO; 2. ISSN 0094-8373.
- ^ Kerp, H .; Trewin, N.H .; Хасс, Х. (2004). «Новые гаметофиты из раннедевонского рини черта». Сделки Королевского общества Эдинбурга. 94 (4): 411–428. Дои:10,1017 / с026359330000078x.
- ^ Taylor, T.N .; Kerp, H .; Хасс, Х. (2005). «Биология жизненного цикла ранних наземных растений: расшифровка фазы гаметофита». Труды Национальной академии наук Соединенных Штатов Америки. 102 (16): 5892–5897. Bibcode:2005ПНАС..102.5892Т. Дои:10.1073 / pnas.0501985102. ЧВК 556298. PMID 15809414.
- ^ Холлоуэй, Джон Э. (1939). "Гаметофит, зародыш и молодое корневище Psilotum triquetrum (Шварц) ". Анналы ботаники. 3 (2): 313–336. Дои:10.1093 / oxfordjournals.aob.a085063.
- ^ Кидстон, Р. и Ланг, W.H. (1920), "На старых заводах из красного песчаника, показывающих структуру, из пласта Рини Черт, Абердиншир. Часть II. Дополнительные примечания к Риния гвинн-вогани, Кидстон и Ланг; с описанием Риния майор, n.sp. и Hornea lignieri, n.g., n.sp ", Сделки Королевского общества Эдинбурга, 52 (3): 603–627, Дои:10,1017 / с0080456800004488
- ^ Реми, В .; Taylor, T.N .; Hass, H .; Керп, Х. (1994). «Везикулярно-арбускулярная микориза возрастом 400 миллионов лет». Труды Национальной академии наук Соединенных Штатов Америки. 91 (25): 11841–11843. Bibcode:1994PNAS ... 9111841R. Дои:10.1073 / пнас.91.25.11841. ЧВК 45331. PMID 11607500.
- ^ Schüßler, A .; и другие. (2001). "Новый грибной тип, Гломеромикота: филогения и эволюция ». Mycol. Res. 105 (12): 1416. Дои:10.1017 / S0953756201005196.
- ^ а б Смит, Салли Э .; Прочтите, Дэвид Дж. (2008). Микоризный симбиоз (3-е изд.). Академическая пресса. ISBN 9780123705266.
- ^ Саймон, L .; Bousquet, J .; Lévesque, R.C .; Лалонд, М. (1993). «Происхождение и разнообразие эндомикоризных грибов и совпадение с сосудистыми наземными растениями». Природа. 363 (6424): 67–69. Bibcode:1993Натура.363 ... 67С. Дои:10.1038 / 363067a0. S2CID 4319766.
- ^ Саймон, L .; Bousquet, J .; Lévesque, R.C .; Лалонд, М. (1993). «Происхождение и разнообразие эндомикоризных грибов и совпадение с сосудистыми наземными растениями». Природа. 363 (6424): 67–69. Bibcode:1993Натура 363 ... 67С. Дои:10.1038 / 363067a0. S2CID 4319766.
- ^ Humphreys, C.P .; Franks, P.J .; Rees, M .; Bidartondo, M. I .; Лик, Дж. Р .; Бирлинг, Д. Дж. (2010). «Взаимный микоризоподобный симбиоз в древнейшей группе наземных растений». Nature Communications. 1 (8): 103. Bibcode:2010 НатКо ... 1E.103H. Дои:10.1038 / ncomms1105. PMID 21045821.
- ^ Брандретт, М. С. (2002). «Коэволюция корней и микориз наземных растений». Новый Фитолог. 154 (2): 275–304. Дои:10.1046 / j.1469-8137.2002.00397.x.
- ^ а б c d е ж г час я j k л м п о п q р Сперри, Дж. С. (2003). «Эволюция водного транспорта и структуры ксилемы». Международный журнал наук о растениях. 164 (3): S115 – S127. Дои:10.1086/368398. JSTOR 3691719. S2CID 15314720.
- ^ а б c d Ворон, Джон (1977). «Эволюция сосудистых наземных растений в связи с надклеточными транспортными процессами». Достижения в ботанических исследованиях. 1977: 153–219. Дои:10.1016 / S0065-2296 (08) 60361-4. ISBN 9780120059058.
- ^ Qiu, Y.L .; Li, L .; Ван, Б .; Chen, Z .; Knoop, V .; Groth-malonek, M .; Домбровская, О .; Lee, J .; Kent, L .; Rest, J .; и другие. (2006). «Самые глубокие расхождения в наземных растениях, выведенные из филогеномных данных». Труды Национальной академии наук. 103 (42): 15511–6. Bibcode:2006PNAS..10315511Q. Дои:10.1073 / pnas.0603335103. ЧВК 1622854. PMID 17030812.
- ^ Эдвардс, Д .; Дэвис, К.Л .; Акс, Л. (1992). "Сосудистая проводящая нить у раннего наземного растения Cooksonia". Природа. 357 (6380): 683–685. Bibcode:1992Натура.357..683E. Дои:10.1038 / 357683a0. S2CID 4264332.
- ^ Никлас, К. Дж .; Смоковитис В. (1983). "Доказательства проводящей нити в растениях раннего силурия (лландовериане): значение для эволюции наземных растений". Палеобиология. 9 (2): 126–137. Дои:10.1017 / S009483730000751X. JSTOR 2400461. S2CID 35550235.
- ^ а б c d е Никлас, К. Дж. (1985). «Эволюция диаметра трахеиды у ранних сосудистых растений и ее влияние на гидравлическую проводимость первичной ксилемы». Эволюция. 39 (5): 1110–1122. Дои:10.2307/2408738. JSTOR 2408738. PMID 28561493.
- ^ Никлас, К .; Пратт, Л. (1980). «Доказательства наличия лигнин-подобных компонентов в окаменелостях растений раннего силурия (лландовериан)». Наука. 209 (4454): 396–397. Bibcode:1980Sci ... 209..396N. Дои:10.1126 / science.209.4454.396. PMID 17747811. S2CID 46073056.
- ^ Pittermann J .; Sperry J.S .; Hacke U.G .; Wheeler J.K .; Sikkema E.H. (Декабрь 2005 г.). «Ямы Торус-Марго помогают хвойным растениям конкурировать с покрытосеменными». Научный журнал. 310 (5756): 1924. Дои:10.1126 / наука.1120479. PMID 16373568.
- ^ Почему рождественские елки не вымерли
- ^ а б Кенрик, Пол (2001). «Переворачивая новую страницу». Природа. 410 (6826): 309–310. Bibcode:2001Натура.410..309K. Дои:10.1038/35066649. PMID 11268183. S2CID 4335355.
- ^ а б c Бирлинг Д.; Osborne, C.P .; Чалонер, В. Г. (2001). "Эволюция формы листьев у наземных растений, связанная с атмосферным CO2 закат в позднепалеозойскую эру » (PDF). Природа. 410 (6826): 352–354. Bibcode:2001Натура.410..352Б. Дои:10.1038/35066546. PMID 11268207. S2CID 4386118.
- ^ Crane, P.R .; Кенрик, П. (1997). «Направленное развитие репродуктивных органов: источник морфологических инноваций у наземных растений». Систематика и эволюция растений. 206 (1): 161–174. Дои:10.1007 / BF00987946. S2CID 23626358.
- ^ Циммерманн, В. (1959). Die Phylogenie der Pflanzen (2-е изд.). Штутгарт: Густав Фишер Верлаг.
- ^ Piazza, P .; и другие. (2005). «Эволюция механизмов развития листа». Новый Фитол. 167 (3): 693–710. Дои:10.1111 / j.1469-8137.2005.01466.x. PMID 16101907.
- ^ Hagemann, W. 1976. Sind Farne Kormophyten? Eine Alternative zur Telomtheorie. Систематика и эволюция растений 124: 251–277.
- ^ Хагеманн, W (1999). «К организменной концепции наземных растений: маргинальная бластозона и развитие вегетативного тела избранных гаметофитов вайбовника печеночников и папоротников». Систематика и эволюция растений. 216 (1–2): 81–133. Дои:10.1007 / bf00985102. S2CID 21483458.
- ^ Саттлер, Р. (1992). «Морфология процессов: структурная динамика в развитии и эволюции». Канадский журнал ботаники. 70 (4): 708–714. Дои:10.1139 / b92-091.
- ^ Саттлер, Р. 1998. О происхождении симметрии, ветвления и филлотаксиса у наземных растений. В: R.V. Жан и Д. Барабе (ред.) Симметрия в растениях. World Scientific, Сингапур, стр. 775-793.
- ^ Джеймс, П.Дж. (2009). "'Дерево и лист: Другой ракурс ». Линней. 25: 17.
- ^ "Взгляд на СО2 теория ранней эволюции листа ». Архивировано из оригинал на 2011-06-29. Получено 2011-03-08.
- ^ Котык, МЭ; Бейсингер, Дж. Ф.; Gensel, PG; де Фрейтас, Т.А. (2002). «Морфологически сложные макрофоссилии растений из позднего силура арктической Канады». Американский журнал ботаники. 89 (6): 1004–1013. Дои:10.3732 / ajb.89.6.1004. PMID 21665700.
- ^ Рикардс, Р. Б. (2000). «Возраст самых ранних клубных мхов: силурийская флора Baragwanathia в Виктории, Австралия» (Абстрактные). Геологический журнал. 137 (2): 207–209. Bibcode:2000ГеоМ..137..207Р. Дои:10.1017 / S0016756800003800. Получено 2007-10-25.
- ^ а б c d е Каплан, Д. (2001). «Наука морфологии растений: определение, история и роль в современной биологии». Американский журнал ботаники. 88 (10): 1711–1741. Дои:10.2307/3558347. JSTOR 3558347. PMID 21669604.
- ^ Taylor, T.N .; Hass, H .; Kerp, H .; Krings, M .; Hanlin, R.T. (2005). «Перитециальные аскомицеты из Rhynie chert возрастом 400 миллионов лет: пример предкового полиморфизма». Микология. 97 (1): 269–285. Дои:10.3852 / mycologia.97.1.269. HDL:1808/16786. PMID 16389979.
- ^ а б c Boyce, C.K .; Knoll, A.H. (2002). «Эволюция потенциала развития и множественное независимое происхождение листьев у сосудистых растений палеозоя». Палеобиология. 28 (1): 70–100. Дои:10.1666 / 0094-8373 (2002) 028 <0070: EODPAT> 2.0.CO; 2. ISSN 0094-8373.
- ^ Hao, S .; Beck, C.B .; Деминг, В. (2003). «Структура самых ранних листьев: адаптации к высоким концентрациям атмосферного CO
2". Международный журнал наук о растениях. 164 (1): 71–75. Дои:10.1086/344557. S2CID 84741832. - ^ Berner, R.A .; Котавала, З. (2001). "Геокарб III: новая модель атмосферы. CO
2 за фанерозойское время ». Американский журнал науки. 301 (2): 182–204. Bibcode:2001AmJS..301..182B. CiteSeerX 10.1.1.393.582. Дои:10.2475 / ajs.301.2.182. Архивировано из оригинал (Абстрактные) на 2009-01-29. Получено 2008-04-07. - ^ Taylor, T.N .; Тейлор, Э. (1993). «Биология и эволюция ископаемых растений». Цитировать журнал требует
| журнал =(Помогите) - ^ Shellito, C.J .; Sloan, L.C. (2006). «Реконструкция потерянного эоценового рая: Часть I. Моделирование изменения глобального распределения цветков при начальном термальном максимуме эоцена». Глобальные и планетарные изменения. 50 (1–2): 1–17. Bibcode:2006GPC .... 50 .... 1S. Дои:10.1016 / j.gloplacha.2005.08.001.
- ^ Аертс, Р. (1995). «Преимущества вечнозеленого растения». Тенденции в экологии и эволюции. 10 (10): 402–407. Дои:10.1016 / S0169-5347 (00) 89156-9. PMID 21237084.
- ^ Labandeira, C.C .; Дилчер, Д.Л .; Дэвис, Д.Р .; Вагнер, Д. (1994). «Девяносто семь миллионов лет ассоциации покрытосеменных и насекомых: палеобиологическое понимание значения коэволюции». Труды Национальной академии наук Соединенных Штатов Америки. 91 (25): 12278–12282. Bibcode:1994PNAS ... 9112278L. Дои:10.1073 / пнас.91.25.12278. ЧВК 45420. PMID 11607501.
- ^ Коричневый V; и другие. (1991). «Травоядность и эволюция размера и формы листьев». Философские труды Королевского общества B. 333 (1267): 265–272. Дои:10.1098 / рстб.1991.0076.
- ^ Harrison C.J .; и другие. (2005). «Независимое задействование законсервированного механизма развития во время эволюции листа». Природа. 434 (7032): 509–514. Bibcode:2005Натура.434..509H. Дои:10.1038 / природа03410. PMID 15791256. S2CID 4335275.
- ^ Джексон Д .; Хак С. (1999). «Контроль филлотаксии кукурузы с помощью гена ABPHYL1». Развитие. 126 (2): 315–323. PMID 9847245.
- ^ Кронк К. (2001). «Эволюция и развитие растений в постгеномном контексте». Природа Обзоры Генетика. 2 (8): 607–619. Дои:10.1038/35084556. PMID 11483985. S2CID 22929939.
- ^ Таттерсолл; и другие. (2005). «Mutant crispa раскрывает множественные роли PHANTASTICA в развитии сложных листьев гороха». Растительная клетка. 17 (4): 1046–1060. Дои:10.1105 / tpc.104.029447. ЧВК 1087985. PMID 15749758.
- ^ Бхаратхан, Гита; Синха, Нилима Рой (Декабрь 2001 г.). «Регуляция сложного развития листьев». Физиология растений. 127 (4): 1533–1538. Дои:10.1104 / стр.010867. ЧВК 1540187. PMID 11743098.
- ^ Нат, У; Кроуфорд, Британская Колумбия; Карпентер, Р. и другие. (2003). «Генетический контроль кривизны поверхности». Наука. 299 (5611): 1404–1407. CiteSeerX 10.1.1.625.1791. Дои:10.1126 / science.1079354. PMID 12610308. S2CID 8059321.
- ^ Ли, Чао; Чжан, Баохун (февраль 2016 г.). «МикроРНК в контроле развития растений». Журнал клеточной физиологии. 231 (2): 303–13. Дои:10.1002 / jcp.25125. PMID 26248304. S2CID 23434501.
- ^ Меккья, Массачусетс; Дебернарди, JM; Родригес, RE; Шоммер, К; Палатник, JF (январь 2013 г.). «МикроРНК miR396 и RDR6 синергетически регулируют развитие листа». Механизмы развития. 130 (1): 2–13. Дои:10.1016 / j.mod.2012.07.005. PMID 22889666.
- ^ де Соуза, Рошели; Амброзини, Адриана; Пассалья, Лучиан М.П. (Декабрь 2015 г.). «Бактерии, способствующие росту растений, как инокулянты в сельскохозяйственных почвах». Генетика и молекулярная биология. 38 (4): 401–19. Дои:10.1590 / S1415-475738420150053. ЧВК 4763327. PMID 26537605.
- ^ Mora, C.I .; Driese, S.G .; Коларуссо, Л.А. (1996). «Средне-позднепалеозойский атмосферный CO
2 Уровни карбонатов и органических веществ в почве ». Наука. 271 (5252): 1105–1107. Bibcode:1996Научный ... 271.1105М. Дои:10.1126 / science.271.5252.1105. S2CID 128479221. - ^ Бернер, Р.А. (1994). «ГЕОКАРБ II: Пересмотренная модель атмосферного CO
2 за фанерозойское время ». Am. J. Sci. 294 (1): 56–91. Bibcode:1994AmJS..294 ... 56B. Дои:10.2475 / ajs.294.1.56. - ^ а б Algeo, T.J .; Berner, R.A .; Maynard, J.B .; Scheckler, S.E .; Архивы, G.S.A.T. (1995). "Океанические аноксические явления в позднем девоне и биотические кризисы:" корни "в эволюции сосудистых наземных растений?". GSA сегодня. 5 (3).
- ^ Реталлак, Дж. Дж. (1986). Райт, В. П. (ред.). Палеопочвы: их признание и интерпретация. Оксфорд: Блэквелл.
- ^ «Трахеофита - 2». Paleos - Жизнь сквозь глубокое время.
- ^ Ричард М. Бейтман, Питер Р. Крейн, Уильям А. Димикеле, Пол Р. Кенрик, Ник П. Роу, Томас Спек и Уильям Э. Стейн (1998). «РАННЯЯ ЭВОЛЮЦИЯ НАЗЕМНЫХ РАСТЕНИЙ: филогения, физиология и экология первичной земной радиации». Ежегодный обзор экологии и систематики. 29: 263–292. Дои:10.1146 / annurev.ecolsys.29.1.263. S2CID 44508826.CS1 maint: несколько имен: список авторов (ссылка на сайт)
- ^ Хетерингтон, Александр Дж; Долан, Лиам (2017). «Двусторонне симметричные оси с ризоидами составляли корневую структуру общего предка сосудистых растений». Философские труды Королевского общества B: биологические науки. 373 (1739): 20170042. Дои:10.1098 / rstb.2017.0042. ЧВК 5745339. PMID 29254968.
- ^ а б c d е ж г час я Algeo, T.J .; Шеклер, С. (1998). «Наземно-морские телесвязи в девоне: связи между эволюцией наземных растений, процессами выветривания и морскими бескислородными явлениями». Философские труды Королевского общества B. 353 (1365): 113–130. Дои:10.1098 / rstb.1998.0195. ЧВК 1692181.
- ^ Хетерингтон, Александр Дж .; Долан, Лиам (2018). «Поэтапное и независимое происхождение корней у наземных растений». Природа. 561 (7722): 235–238. Bibcode:2018Натура.561..235H. Дои:10.1038 / s41586-018-0445-z. ЧВК 6175059. PMID 30135586.
- ^ Кудерт, Йоан; Энн Диеварт; Гаэтан Дрок; Паскаль Ганте (2012). «Филогения ASL / LBD предполагает, что генетические механизмы инициации корня ниже по течению от ауксина различны у ликофитов и эвфиллофитов». Молекулярная биология и эволюция. 30 (3): 569–72. Дои:10.1093 / молбев / mss250. ISSN 0737-4038. PMID 23112232.
- ^ а б Kenrick, P .; Крейн, П.Р. (1997). «Происхождение и ранняя эволюция растений на суше». Природа. 389 (6646): 33–39. Bibcode:1997 Натур.389 ... 33К. Дои:10.1038/37918. S2CID 3866183.
- ^ Ален Пьер, Жан-Люк Ма, Корентен Клеман, Жан-Пьер Монторой, Кристиан Хартманн, Сантимейтри Гонкамди (июль 2016 г.). «Понимание глубоких корней и их функций в экосистемах: пропаганда более нетрадиционных исследований». Анналы ботаники. 118 (4): 621–635. Дои:10.1093 / aob / mcw130. ЧВК 5055635. PMID 27390351.CS1 maint: несколько имен: список авторов (ссылка на сайт)
- ^ Жерриенн, Филипп; Gensel, Patricia G .; Струллу-Дерриен, Кристина; Лардо, Юбер; Стиманс, Филипп; Престианни, Сирил (2011). «Простая порода древесины двух раннедевонских растений». Наука. 333 (6044): 837. Bibcode:2011Sci ... 333..837G. Дои:10.1126 / science.1208882. PMID 21836008. S2CID 23513139.
- ^ Кинвер, Марк (16 августа 2011 г.). "'Образцы ранней древесины меняют историю растений ». Новости BBC.
- ^ Stein, W.E .; Mannolini, F .; Hernick, L.V .; Посадка, Е .; Берри, К. (2007). «Гигантские кладоксилопсидовые деревья разрешают загадку самых ранних пней на Земле в Гильбоа». Природа. 446 (7138): 904–7. Bibcode:2007Натура.446..904С. Дои:10.1038 / природа05705. PMID 17443185. S2CID 2575688.
- ^ Retallack, G.J .; Catt, J.A .; Чалонер, W.G. (1985). «Ископаемые почвы как основание для интерпретации появления на суше крупных растений и животных [и обсуждение]». Философские труды Королевского общества B. 309 (1138): 105–142. Bibcode:1985РСПТБ.309..105Р. Дои:10.1098 / рстб.1985.0074. JSTOR 2396355.
- ^ Dannenhoffer, J.M .; Бонамо, П. (1989). "Rellimia thomsonii из Живетана в Нью-Йорке: вторичный рост в трех порядках ветвления ». Американский журнал ботаники. 76 (9): 1312–1325. Дои:10.2307/2444557. JSTOR 2444557.
- ^ Дэвис, П.; Кенрик, П. (2004). Ископаемые растения. Smithsonian Books, Вашингтон, округ Колумбия. ISBN 978-1-58834-156-3.
- ^ Донохью, М.Дж. (2005). «Ключевые инновации, конвергенция и успех: макроэволюционные уроки филогении растений» (Абстрактные). Палеобиология. 31 (2): 77–93. Дои:10.1666 / 0094-8373 (2005) 031 [0077: KICASM] 2.0.CO; 2. ISSN 0094-8373. Получено 2008-04-07.
- ^ а б c Bowe, L.M .; Пальто, G .; Депамфилис, C.W. (2000). «Филогения семенных растений на основе всех трех геномных компартментов: современные голосеменные растения монофилетичны, а ближайшие родственники Gnetales - хвойные». Труды Национальной академии наук. 97 (8): 4092–7. Bibcode:2000PNAS ... 97.4092B. Дои:10.1073 / pnas.97.8.4092. ЧВК 18159. PMID 10760278.
- ^ а б c Chaw, S.M .; Parkinson, C.L .; Cheng, Y .; Винсент, Т.М .; Палмер, Дж. Д. (2000). «Филогения семенных растений, выведенная из всех трех геномов растений: монофилия современных голосеменных растений и происхождение Gnetales из хвойных пород». Труды Национальной академии наук. 97 (8): 4086–91. Bibcode:2000PNAS ... 97.4086C. Дои:10.1073 / pnas.97.8.4086. ЧВК 18157. PMID 10760277.
- ^ а б c Soltis, D.E .; Soltis, P.S .; Занис, М.Дж. (2002). «Филогения семенных растений на основе данных восьми генов». Американский журнал ботаники. 89 (10): 1670–81. Дои:10.3732 / ajb.89.10.1670. PMID 21665594. Архивировано из оригинал (Абстрактные) на 2008-04-12. Получено 2008-04-08.
- ^ Friis, E.M .; Pedersen, K.R .; Крейн, П.Р. (2006). «Цветки покрытосеменных меловых: инновации и эволюция в репродукции растений». Палеогеография, палеоклиматология, палеоэкология. 232 (2–4): 251–293. Bibcode:2006ППП ... 232..251Ф. Дои:10.1016 / j.palaeo.2005.07.006.
- ^ а б Hilton, J .; Бейтман, Р. (2006). «Птеридоспермия - это основа филогении семенных растений». Журнал Ботанического общества Торри. 133 (1): 119–168. Дои:10.3159 / 1095-5674 (2006) 133 [119: PATBOS] 2.0.CO; 2. ISSN 1095-5674.
- ^ а б Bateman, R.M .; Hilton, J .; Рудалл, П.Дж. (2006). «Морфологический и молекулярно-филогенетический контекст покрытосеменных растений: контрастирование подходов« сверху вниз »и« снизу вверх », используемых для вывода вероятных характеристик первых цветков». Журнал экспериментальной ботаники. 57 (13): 3471–503. Дои:10.1093 / jxb / erl128. PMID 17056677.
- ^ а б c d Frohlich, M.W .; Чейз, М.В. (2007). «После десятка лет прогресса происхождение покрытосеменных растений все еще остается большой загадкой». Природа. 450 (7173): 1184–9. Bibcode:2007 Натур.450.1184F. Дои:10.1038 / природа06393. PMID 18097399. S2CID 4373983.
- ^ "Научный журнал". Рункария, предшественник семенных растений среднего девона. Американская ассоциация развития науки. 2011 г.. Получено 22 марта, 2011.
- ^ а б c Mapes, G .; Rothwell, G.W .; Хаворт, М. (1989). «Эволюция покоя семян». Природа. 337 (6208): 645–646. Bibcode:1989Натура.337..645М. Дои:10.1038 / 337645a0. S2CID 4358761.
- ^ Bawa, KS; Уэбб, CJ (1984). «Цветочные, фруктовые и семенные аборт в деревьях тропических лесов. Последствия для эволюции отцовских и материнских репродуктивных моделей». Американский журнал ботаники. 71 (5): 736–751. Дои:10.2307/2443371. JSTOR 2443371.
- ^ Чеа К.С., Осборн ди-джей (апрель 1978 г.). «Повреждения ДНК происходят с потерей жизнеспособности у зародышей стареющих семян ржи». Природа. 272 (5654): 593–9. Bibcode:1978Натура.272..593C. Дои:10.1038 / 272593a0. PMID 19213149. S2CID 4208828.
- ^ Коппен G, Вершаев L (2001). «Щелочной одноклеточный гель-электрофорез / анализ комет: способ изучения восстановления ДНК в корешковых клетках прорастающих Vicia faba». Folia Biol. (Прага). 47 (2): 50–4. PMID 11321247.
- ^ Брей К.М., Западная Европа (декабрь 2005 г.). «Механизмы репарации ДНК в растениях: важнейшие сенсоры и эффекторы для поддержания целостности генома». Новый Фитол. 168 (3): 511–28. Дои:10.1111 / j.1469-8137.2005.01548.x. PMID 16313635.
- ^ Бернштейн С., Бернштейн Х. (1991) Старение, секс и восстановление ДНК. Academic Press, Сан-Диего. ISBN 0120928604 ISBN 978-0120928606
- ^ Feild, T. S .; Brodribb, T. J .; Иглесиас, А .; Chatelet, D. S .; Baresch, A .; Upchurch, G.R .; Gomez, B .; Mohr, B.A.R .; Coiffard, C .; Kvacek, J .; Харамилло, К. (2011). «Ископаемые свидетельства эскалации мелового периода в эволюции жилок листа покрытосеменных». Труды Национальной академии наук. 108 (20): 8363–8366. Bibcode:2011PNAS..108.8363F. Дои:10.1073 / pnas.1014456108. ЧВК 3100944. PMID 21536892.
- ^ а б Lawton-Rauh, A .; Альварес-Буйлла, E.R .; Пуругганан, доктор медицины (2000). «Молекулярная эволюция развития цветов». Тенденции в экологии и эволюции. 15 (4): 144–149. Дои:10.1016 / S0169-5347 (99) 01816-9. PMID 10717683.
- ^ Китайская Академия Наук (18 декабря 2018 г.). «Цветы возникли на 50 миллионов лет раньше, чем считалось ранее». EurekAlert!. Получено 18 декабря 2018.
- ^ Койро, Марио; Дойл, Джеймс А .; Хилтон, Джейсон (2019-01-25). «Насколько глубок конфликт между молекулярными и ископаемыми данными о возрасте покрытосеменных растений?». Новый Фитолог. 223 (1): 83–99. Дои:10.1111 / nph.15708. PMID 30681148.
- ^ Nam, J .; Депамфилис, CW; Ма, Н; Ней, М. (2003). «Древность и эволюция семейства генов MADS-Box, контролирующих развитие цветков у растений». Мол. Биол. Evol. 20 (9): 1435–1447. Дои:10.1093 / molbev / msg152. PMID 12777513.
- ^ а б c Крепет, В. Л. (2000). «Прогресс в понимании истории покрытосеменных, успехов и взаимоотношений: отвратительно» загадочное явление Дарвина"". Труды Национальной академии наук. 97 (24): 12939–41. Дои:10.1073 / pnas.97.24.12939. ЧВК 34068. PMID 11087846.
- ^ Sun, G .; Ji, Q .; Дилчер, Д.Л .; Zheng, S .; Nixon, K.C .; Ван, X. (2002). "Archaefructaceae, новое семейство базальных покрытосеменных". Наука. 296 (5569): 899–904. Bibcode:2002Наука ... 296..899С. Дои:10.1126 / science.1069439. PMID 11988572. S2CID 1910388.
- ^ По факту, Archaefructus наверное настоящих цветов не родила: см.
- Friis, E.M .; Doyle, J.A .; Endress, P.K .; Ленг, К. (2003). «Archaefructus - предшественник покрытосеменных или специализированный ранний покрытосеменный?». Тенденции в растениеводстве. 8 (8): 369–373. Дои:10.1016 / S1360-1385 (03) 00161-4. PMID 12927969.
- ^ а б Wing, S.L .; Буше, Л. (1998). "Экологические аспекты радиации меловых цветковых растений". Ежегодный обзор наук о Земле и планетах. 26 (1): 379–421. Bibcode:1998AREPS..26..379Вт. Дои:10.1146 / annurev.earth.26.1.379.
- ^ а б Feild, T.S .; Аренс, Северная Каролина; Doyle, J.A .; Dawson, T.E .; Донохью, М.Дж. (2004). «Темный и тревожный: новый образ экологии ранних покрытосеменных» (Абстрактные). Палеобиология. 30 (1): 82–107. Дои:10.1666 / 0094-8373 (2004) 030 <0082: DADANI> 2.0.CO; 2. ISSN 0094-8373. Получено 2008-04-08.
- ^ Габботт, профессор Сара (1 августа 2017 г.). "Первый цветок выглядел так?". Новости BBC. Получено 1 августа 2017.
- ^ Sauquet, Эрве; и другие. (1 августа 2017 г.). «Родовой цветок покрытосеменных растений и его раннее разнообразие». Nature Communications. 8: 16047. Bibcode:2017НатКо ... 816047S. Дои:10.1038 / ncomms16047. ЧВК 5543309. PMID 28763051.
- ^ Проект генома амбореллы (20 декабря 2013 г.). "The Амборелла Геном и эволюция цветковых растений ». Наука. 342 (6165): 1241089. Дои:10.1126 / science.1241089. PMID 24357323. S2CID 202600898. Получено 1 февраля 2020.
- ^ Zuccolo, A .; и другие. (2011). "Физическая карта для Amborella trichopoda геном проливает свет на эволюцию структуры генома покрытосеменных ». Геномная биология. 12 (5): R48. Дои:10.1186 / gb-2011-12-5-r48. ЧВК 3219971. PMID 21619600.
- ^ Медарг Н.Г., Яновский М. (март 2001 г.). «Функция и эволюция семейства генов MADS-box растений». Природа Обзоры Генетика. 2 (3): 186–195. Дои:10.1038/35056041. PMID 11256070. S2CID 7399106.
- ^ Ягер; и другие. (2003). «Гены MADS-Box в гинкго билоба и эволюция семейства AGAMOUS». Мол. Биол. Evol. 20 (5): 842–854. Дои:10.1093 / molbev / msg089. PMID 12679535.
- ^ Трансляционная биология: от цветов арабидопсиса к архитектуре соцветий травы. Бет Э. Томпсон * и Сара Хейк, 2009 г., Физиология растений 149: 38–45.
- ^ Китахара К., Мацумото С. (2000). «Гены MADS-бокса розы MASAKO C1 и D1, гомологичные генам цветочной идентичности класса C». Растениеводство. 151 (2): 121–134. Дои:10.1016 / S0168-9452 (99) 00206-X. PMID 10808068.
- ^ Катер М; и другие. (1998). «Множественные агамовые гомологи из огурца и петунии различаются по своей способности влиять на судьбу репродуктивного органа». Растительная клетка. 10 (2): 171–182. Дои:10.1105 / tpc.10.2.171. ЧВК 143982. PMID 9490741.
- ^ Soltis D; и другие. (2007). «Цветочный геном: эволюционная история дупликации генов и смещения моделей экспрессии генов». Тенденции Plant Sci. 12 (8): 358–367. Дои:10.1016 / j.tplants.2007.06.012. PMID 17658290.
- ^ Путтерхилл; и другие. (2004). «Пора цвести: генетический контроль времени цветения». BioEssays. 26 (4): 353–363. Дои:10.1002 / bies.20021. PMID 15057934. S2CID 43335309. Архивировано из оригинал на 2012-12-16.
- ^ Бласкес; и другие. (2001). «Цветение вовремя: гены, регулирующие переход цветков». EMBO отчеты. 2 (12): 1078–1082. Дои:10.1093 / embo-reports / kve254. ЧВК 1084172. PMID 11743019.
- ^ Frohlich, Майкл В .; Паркер, Дэвид С. (2000). "В основном мужская теория эволюционного происхождения цветов: от генов до окаменелостей". Syst. Бот. 25 (2): 155–170. Дои:10.2307/2666635. JSTOR 2666635. S2CID 85612923.
- ^ Ли А., Мао Л. (2007). «Эволюция семейств генов микроРНК растений». Клеточные исследования. 17 (3): 212–218. Дои:10.1038 / sj.cr.7310113. PMID 17130846.
- ^ Добли Дж. Ф. (2004). «Генетика эволюции кукурузы». Анну. Преподобный Жене. 38 (1): 37–59. Дои:10.1146 / annurev.genet.38.072902.092425. PMID 15568971.
- ^ Пуруганнан; и другие. (2000). «Вариации и отбор цветочного гомеотического гена цветной капусты, сопровождающего эволюцию одомашненной Brassica olerace». Генетика. 155 (2): 855–862. ЧВК 1461124. PMID 10835404.
- ^ а б Уильямс Б.П., Джонстон И.Г., Ковшофф С., Хибберд Дж. М. (сентябрь 2013 г.). «Фенотипический ландшафтный вывод показывает несколько эволюционных путей к фотосинтезу C₄». eLife. 2: e00961. Дои:10.7554 / eLife.00961. ЧВК 3786385. PMID 24082995.
- ^ Кристин, П. -А .; Osborne, C.P .; Chatelet, D. S .; Columbus, J. T .; Besnard, G .; Hodkinson, T. R .; Гаррисон, Л. М .; Воронцова, М. С .; Эдвардс, Э. Дж. (2012). «Анатомические факторы и эволюция фотосинтеза C4 в травах». Труды Национальной академии наук. 110 (4): 1381–1386. Bibcode:2013PNAS..110.1381C. Дои:10.1073 / pnas.1216777110. ЧВК 3557070. PMID 23267116.
- ^ а б c d Osborne, C.P .; Бирлинг, Д.Дж. (2006). "Обзор. Зеленая революция природы: выдающийся эволюционный рост C4 растения » (PDF). Философские труды Королевского общества B. 361 (1465): 173–194. Дои:10.1098 / rstb.2005.1737. ЧВК 1626541. PMID 16553316. Получено 2008-02-11.[постоянная мертвая ссылка ]
- ^ а б Реталлак, Дж. Дж. (1 августа 1997 г.). «Неогеновая экспансия Североамериканских прерий». ПАЛАИ. 12 (4): 380–390. Bibcode:1997Палай..12..380р. Дои:10.2307/3515337. ISSN 0883-1351. JSTOR 3515337.
- ^ Thomasson, J.R .; Nelson, M.E .; Закжевский, Р.Дж. (1986). «Ископаемые травы (Gramineae: Chloridoideae) из миоцена с анатомией Кранца». Наука. 233 (4766): 876–878. Bibcode:1986Sci ... 233..876T. Дои:10.1126 / science.233.4766.876. PMID 17752216. S2CID 41457488.
- ^ О'Лири, Мэрион (май 1988). «Изотопы углерода в фотосинтезе». Бионаука. 38 (5): 328–336. Дои:10.2307/1310735. JSTOR 1310735.
- ^ Osborne, P .; Фреклтон, П. (февраль 2009 г.). «Экологические факторы отбора для фотосинтеза C4 в травах». Труды: Биологические науки.. 276 (1663): 1753–1760. Дои:10.1098 / rspb.2008.1762. ISSN 0962-8452. ЧВК 2674487. PMID 19324795.
- ^ Bond, W.J .; Woodward, F.I .; Мидгли, Г.Ф. (2005). «Глобальное распределение экосистем в мире без огня». Новый Фитолог. 165 (2): 525–538. Дои:10.1111 / j.1469-8137.2004.01252.x. PMID 15720663.
- ^ При содержании кислорода в воздухе выше 35% распространение огня невозможно остановить. Многие модели предсказывали более высокие значения, и их пришлось пересмотреть, потому что не произошло полного исчезновения растительной жизни.
- ^ Pagani, M .; Zachos, J.C .; Freeman, K.H .; Типпл, В .; Бохаты, С. (2005). «Заметное снижение концентрации углекислого газа в атмосфере в палеогене». Наука. 309 (5734): 600–603. Bibcode:2005Наука ... 309..600П. Дои:10.1126 / science.1110063. PMID 15961630. S2CID 20277445.
- ^ Piperno, D.R .; Сьюз, H.D. (2005). «Динозавры обедали на траве». Наука. 310 (5751): 1126–8. Дои:10.1126 / science.1121020. PMID 16293745. S2CID 83493897.
- ^ Прасад, V .; Stroemberg, C.A.E .; Alimohammadian, H .; Сахни, А. (2005). «Копролиты динозавров и ранняя эволюция трав и травоядных». Наука. 310 (5751): 1177–1180. Bibcode:2005Наука ... 310.1177П. Дои:10.1126 / science.1118806. PMID 16293759. S2CID 1816461.
- ^ Keeley, J.E .; Рундель, П. (2005). «Огонь и миоценовое расширение пастбищ C4». Письма об экологии. 8 (7): 683–690. Дои:10.1111 / j.1461-0248.2005.00767.x.
- ^ а б Retallack, G.J. (2001). «Кайнозойское расширение пастбищ и похолодание климата». Журнал геологии. 109 (4): 407–426. Bibcode:2001JG .... 109..407R. Дои:10.1086/320791. S2CID 15560105.
- ^ Джин, JP; Он, К; Тан, X; Ли, Z; Lv, L; Чжао, Y; Луо, Дж; Гао, Джи (июль 2015 г.). «Карта регуляторов транскрипции Arabidopsis выявляет отличительные функциональные и эволюционные особенности новых факторов транскрипции». Молекулярная биология и эволюция. 32 (7): 1767–1773. Дои:10.1093 / molbev / msv058. ЧВК 4476157. PMID 25750178.
- ^ Пичерский Э .; Банда Д. (2000). «Генетика и биохимия вторичных метаболитов растений: эволюционная перспектива». Тенденции Plant Sci. 5 (10): 439–445. Дои:10.1016 / S1360-1385 (00) 01741-6. PMID 11044721.
- ^ Нина Тайс; Мануэль Лердау (2003). «Эволюция функции вторичных метаболитов растений». Int. J. Plant Sci. 164 (S3): S93 – S102. Дои:10.1086/374190. S2CID 85705162.
- ^ Bohlmann J .; и другие. (1998). «Растительные терпеноид-синтазы: молекулярный и филогенетический анализ». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 95 (8): 4126–4133. Bibcode:1998PNAS ... 95.4126B. Дои:10.1073 / пнас.95.8.4126. ЧВК 22453. PMID 9539701.
- ^ а б Хассани, М.А., Дуран, П. и Хаккард, С. (2018) "Микробные взаимодействия внутри холобионта растения". Микробиом, 6(1): 58. Дои:10.1186 / s40168-018-0445-0.
 Материал был скопирован из этого источника, который доступен под Международная лицензия Creative Commons Attribution 4.0.
Материал был скопирован из этого источника, который доступен под Международная лицензия Creative Commons Attribution 4.0. - ^ Lucking, R., Huhndorf, S., Pfister, D.H., Plata, E.R., Lumbsch, H.T. (2009) «Грибы эволюционировали прямо на ходу». Микология, 101(6): 810–822. Дои:10.3852/09-016.
- ^ Хекман, Д.С., Гейзер, Д.М., Эйделл, Б.Р., Штауфер, Р.Л., Кардос, Н.Л. и Hedges, S.B. (2001) «Молекулярные доказательства раннего заселения земли грибами и растениями». Наука, 293(5532): 1129–1133. Дои:10.1126 / science.1061457.
- ^ а б Capelle, J .; Нима, К. (ноябрь 2005 г.). «Местная адаптация и структура популяции в микрогеографическом масштабе грибкового паразита на растении-хозяине». Журнал эволюционной биологии. 18 (6): 1445–1454. Дои:10.1111 / j.1420-9101.2005.00951.x. PMID 16313457. S2CID 12986653.
- ^ а б Burdon, J.J .; Тралл, П. (2009). «Совместная эволюция растений и их патогенов в естественной среде обитания». Наука. 324 (5928): 755–756. Bibcode:2009Sci ... 324..755B. Дои:10.1126 / science.1171663. ЧВК 2689373. PMID 19423818.
- ^ Sahai, A.S .; Маноча, М. (1992). «Хитиназы грибов и растений: их участие в морфогенезе и взаимодействии паразит-хозяин». Обзор микробиологии FEMS. 11 (4): 317–338. Дои:10.1111 / j.1574-6976.1993.tb00004.x.
- ^ Клей, Кит (1991). «Паразитарная кастрация растений грибами». Тенденции в экологии и эволюции. 6 (5): 162–166. Дои:10.1016/0169-5347(91)90058-6. PMID 21232447.
- ^ Zhan, J .; Mundt, C.C .; Hoffer, M.E .; Макдональд, Б.А. (2002). «Локальная адаптация и влияние генотипа хозяина на скорость эволюции патогена: экспериментальный тест в патосистеме растений». Журнал эволюционной биологии. 15 (4): 634–647. Дои:10.1046 / j.1420-9101.2002.00428.x. S2CID 83662474.
- ^ Delmotte, F .; Бучели, Э .; Шикофф, Я. (1999). «Структура популяции хозяев и паразитов в системе естественное растение-патоген». Наследственность. 82 (3): 300–308. Дои:10.1038 / sj.hdy.6884850. PMID 10336705.
- ^ Desprez-Loustau, M.-L .; Vitasse, Y .; Delzon, S .; Capdevielle, X .; Marcais, B .; Кремер, А. (2010). «Приспособлены ли популяции патогенов растений к встрече со своим хозяином? Тематическое исследование фенологической синхронности между дубом и облигатным грибковым паразитом вдоль высотного градиента» (PDF). Журнал эволюционной биологии. 23 (1): 87–97. Дои:10.1111 / j.1420-9101.2009.01881.x. PMID 19895655. S2CID 25006120.
внешние ссылки
| Субдисциплины | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Завод группы | |||||||||||
| |||||||||||
| |||||||||||
| Рост и привычка растений | |||||||||||
| Размножение | |||||||||||
| Таксономия растений | |||||||||||
| Практика | |||||||||||
| |||||||||||
| |||||||||||
