Мейоз - Википедия - Meiosis
Было высказано предположение, что Возобновление мейоза быть слился в эту статью. (Обсуждать) Предлагается с сентября 2020 года. |


Мейоз (/маɪˈoʊsɪs/ (![]() Слушать); из Греческий μείωσις, мейоз, что означает «уменьшение») - это особый вид деление клеток из стволовые клетки в воспроизводящий половым путем организмы, используемые для производства гаметы, Такие как сперма или же яйцеклетки. Он включает в себя два раунда деления, которые в конечном итоге приводят к четырем ячейкам только с одной копией каждой отцовской и материнской. хромосома (гаплоидный ). Кроме того, перед делением генетический материал от отцовской и материнской копий каждой хромосомы пересек, создавая новые комбинации кода на каждой хромосоме.[1] Позже, во время оплодотворение, гаплоидные клетки, полученные в результате мейоза у мужчины и женщины, будут сливаться, чтобы снова создать клетку с двумя копиями каждой хромосомы, зигота.
Слушать); из Греческий μείωσις, мейоз, что означает «уменьшение») - это особый вид деление клеток из стволовые клетки в воспроизводящий половым путем организмы, используемые для производства гаметы, Такие как сперма или же яйцеклетки. Он включает в себя два раунда деления, которые в конечном итоге приводят к четырем ячейкам только с одной копией каждой отцовской и материнской. хромосома (гаплоидный ). Кроме того, перед делением генетический материал от отцовской и материнской копий каждой хромосомы пересек, создавая новые комбинации кода на каждой хромосоме.[1] Позже, во время оплодотворение, гаплоидные клетки, полученные в результате мейоза у мужчины и женщины, будут сливаться, чтобы снова создать клетку с двумя копиями каждой хромосомы, зигота.
Ошибки в мейозе, приводящие к анеуплоидия (аномальное количество хромосом) являются ведущей известной причиной выкидыш и наиболее частая генетическая причина нарушение развития.[2]
В мейозе Репликация ДНК за которым следуют два раунда деления клеток с образованием четырех дочерних клеток, в каждой из которых вдвое меньше хромосомы как исходная родительская ячейка.[1] Эти два подразделения мейоза известны как мейоз I и мейоз II. До начала мейоза, во время Фаза S из клеточный цикл, ДНК каждой хромосомы реплицируется так, что она состоит из двух идентичных сестринские хроматиды, которые остаются скрепленными за счет сцепления сестринских хроматид. Эта S-фаза может упоминаться как «премейотическая S-фаза» или «мейотическая S-фаза». Сразу после репликации ДНК мейотические клетки вступают в длительную грамм2 -подобная стадия, известная как мейотическая профаза. В течение этого времени, гомологичные хромосомы соединяться друг с другом и проходить генетическая рекомбинация, запрограммированный процесс, в котором ДНК может быть разрезана, а затем восстановлена, что позволяет им обмениваться некоторыми из своих генетическая информация. Подмножество событий рекомбинации приводит к кроссоверы, которые создают физические ссылки, известные как хиазмы (единственное число: хиазма, от греческой буквы Чи (X) ) между гомологичными хромосомами. У большинства организмов эти связи могут помочь направить каждую пару гомологичных хромосом к разделять друг от друга во время Мейоза I, в результате чего два гаплоидный клетки, которые имеют половину количества хромосом по сравнению с родительской клеткой.
Во время мейоза II сцепление между сестринскими хроматидами ослабляется, и они отделяются друг от друга, как во время митоз. В некоторых случаях все четыре мейотических продукта образуют гаметы Такие как сперма, споры или же пыльца. У самок животных три из четырех продуктов мейоза обычно выводятся экструзия в полярные тела, и только одна клетка развивается, чтобы произвести яйцеклетка. Поскольку количество хромосом уменьшается вдвое во время мейоза, гаметы могут сливаться (т.е. оплодотворение ) образовать диплоид зигота который содержит две копии каждой хромосомы, по одной от каждого родителя. Таким образом, чередующиеся циклы мейоза и оплодотворения позволяют половое размножение, с последовательными поколениями, сохраняющими одинаковое количество хромосом. Например, диплоид humancells содержат 23 пары хромосом, включая 1 пару половых хромосом (всего 46), половину материнского происхождения и половину отцовского происхождения. Мейоз производит гаплоидный гаметы (яйцеклетки или сперматозоиды), содержащие один набор из 23 хромосом. Когда две гаметы (яйцеклетка и сперматозоид) сливаются, образующаяся зигота снова становится диплоидной, причем каждая мать и отец вносят по 23 хромосомы. Тот же образец, но не одинаковое количество хромосом, встречается у всех организмов, которые используют мейоз.
Мейоз встречается у всех одноклеточных и размножающихся половым путем. многоклеточный организмов (которые все эукариоты ), включая животные, растения и грибы.[3][4][5] Это важный процесс для оогенез и сперматогенез.
Обзор
Хотя процесс мейоза связан с более общим процессом деления клеток митоз, он отличается двумя важными аспектами:
| рекомбинация | мейоз | тасует гены между двумя хромосомами в каждой паре (по одному от каждого родителя), производящие рекомбинантные хромосомы с уникальными генетическими комбинациями в каждой гамете | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| митоз | возникает только при необходимости восстановления повреждений ДНК; обычно встречается между идентичными сестринскими хроматидами и не приводит к генетическим изменениям | ||||||||||
| число хромосом (плоидность) | мейоз | производит четыре генетически уникальные клетки, каждая с половина количество хромосом как у родителя | |||||||||
| митоз | производит две генетически идентичные клетки, каждая с тот же номер хромосом как у родителя | ||||||||||
Мейоз начинается с диплоидной клетки, которая содержит по две копии каждой хромосомы, называемой гомологи. Сначала клетка подвергается Репликация ДНК, так что теперь каждый гомолог состоит из двух идентичных сестринских хроматид. Затем каждый набор гомологов соединяется друг с другом и обменивается генетической информацией посредством гомологичная рекомбинация часто приводит к физическим связям (кроссоверам) между гомологами. При первом делении мейоза гомологи разделяются на дочерние клетки шпиндельный аппарат. Затем клетки переходят ко второму делению без промежуточного цикла репликации ДНК. Сестринские хроматиды разделяются на дочерние клетки с образованием в общей сложности четырех гаплоидных клеток. Самки животных используют небольшую вариацию этого паттерна и производят одну большую яйцеклетку и два маленьких полярных тела. Из-за рекомбинации индивидуальная хроматида может состоять из новой комбинации материнской и отцовской генетической информации, в результате чего потомство генетически отличается от любого из родителей. Кроме того, отдельная гамета может включать в себя набор материнских, отцовских и рекомбинантных хроматид. Это генетическое разнообразие, возникающее в результате полового размножения, способствует изменчивости признаков, по которым естественный отбор может действовать.
Мейоз использует многие из тех же механизмов, что и митоз, тип деления клеток, используемый эукариоты разделить одну клетку на две идентичные дочерние клетки. У некоторых растений, грибов и протисты мейоз приводит к образованию споры: гаплоидные клетки, которые могут вегетативно делиться без оплодотворения. Некоторые эукариоты, например бделлоидные коловратки, не обладают способностью осуществлять мейоз и приобрели способность воспроизводить партеногенез.
Мейоз не возникает в археи или же бактерии, которые обычно размножаются бесполым путем двойное деление. Однако «сексуальный» процесс, известный как горизонтальный перенос генов включает перенос ДНК от одной бактерии или Археон к другому и рекомбинация этих молекул ДНК разного родительского происхождения.
История
Мейоз был открыт и впервые описан в морской еж яйца в 1876 г. немецким биологом Оскар Хертвиг. Он был снова описан в 1883 году на уровне хромосомы бельгийского зоолога Эдуард Ван Бенеден, в Аскариды яйца аскариды. Однако значение мейоза для воспроизводства и наследования было описано только в 1890 году немецким биологом. Август Вейсманн, которые отметили, что для преобразования одной диплоидной клетки в четыре гаплоидных клетки необходимы два деления, если необходимо сохранить количество хромосом. В 1911 г. Американец генетик Томас Хант Морган обнаружены кроссоверы в мейозе у плодовой мушки Drosophila melanogaster, что помогло установить, что генетические признаки передаются по хромосомам.
Термин «мейоз» происходит от греческого слова μείωσις, что означает «уменьшение». Он был введен в биологию Дж. Б. Фармер и J.E.S. Мур в 1905 году, используя идиосинкразическую интерпретацию «майоза»:
Мы предлагаем применять термины Майоз или Майотическая фаза для охвата всей серии ядерных изменений, включенных в два подразделения, которые были обозначены как Гетеротип и Гомотип. Флемминг.[6]
Написание было изменено на «мейоз» Кёрнике (1905) и Пантелем и Де Синети (1906), чтобы следовать обычным соглашениям для транслитерация греческого.[7]
Фазы
Мейоз делится на мейоз I и мейоз II, которые далее подразделяются на кариокинез I и цитокинез I и кариокинез II и цитокинез II соответственно. Подготовительные шаги, ведущие к мейозу, идентичны по структуре и названию интерфазе митотического клеточного цикла.[8] Межфазный делится на три этапа:
- Рост 1 (G1) фаза: В этой очень активной фазе клетка синтезирует широкий спектр белков, включая ферменты и структурные белки, необходимые для роста. В G1каждая из хромосом состоит из одной линейной молекулы ДНК.
- Фаза синтеза (S): Генетический материал воспроизводится; каждая из хромосом клетки дублируется, чтобы стать двумя идентичными сестринские хроматиды прикреплен к центромере. Эта репликация не меняет плоидность ячейки, поскольку номер центромеры остается прежним. Идентичные сестринские хроматиды еще не сконденсировались в плотно упакованные хромосомы, видимые в световой микроскоп. Это происходит во время профазы I мейоза.
- Рост 2 (G2) фаза: ГРАММ2 фаза, которая наблюдалась перед митозом, отсутствует в мейозе. Профаза мейоза наиболее близко соответствует G2 фаза митотического клеточного цикла.
За интерфазой следует мейоз I, затем мейоз II. Мейоз I разделяет реплицированные гомологичные хромосомы, каждая из которых по-прежнему состоит из двух сестринских хроматид, на две дочерние клетки, таким образом уменьшая число хромосом вдвое. Во время мейоза II сестринские хроматиды отделяются и образующиеся дочерние хромосомы разделяются на четыре дочерние клетки. У диплоидных организмов дочерние клетки в результате мейоза являются гаплоидными и содержат только одну копию каждой хромосомы. У некоторых видов клетки входят в фазу покоя, известную как интеркинез между мейозом I и мейозом II.
Каждый мейоз I и II делится на профаза, метафаза, анафаза, и телофаза стадии, сходные по назначению с аналогичными им субфазами в митотическом клеточном цикле. Следовательно, мейоз включает стадии мейоза I (профаза I, метафаза I, анафаза I, телофаза I) и мейоза II (профаза II, метафаза II, анафаза II, телофаза II).

Во время мейоза специфические гены более высоки. записано.[9][10] В дополнение к сильной мейотической стадии экспрессии мРНК, существуют также повсеместные средства контроля трансляции (например, избирательное использование предварительно сформированной мРНК), регулирующие конечную мейотическую стадию специфической экспрессии белков генов во время мейоза.[11] Таким образом, и транскрипционный, и трансляционный контроль определяют широкую реструктуризацию мейотических клеток, необходимую для осуществления мейоза.
Мейоз I
Мейоз I сегрегирует гомологичные хромосомы, которые соединяются как тетрады (2n, 4c), производя две гаплоидные клетки (n хромосом, 23 у человека), каждая из которых содержит пары хроматид (1n, 2c). Поскольку плоидность снижается от диплоидной до гаплоидной, мейоз I называют редукционное деление. Мейоз II - это эквациональное деление аналогично митозу, при котором сестринские хроматиды разделяются с образованием четырех гаплоидных дочерних клеток (1n, 1c).[12]


Профаза I
Профаза I на сегодняшний день является самой продолжительной фазой мейоза (длится 13 из 14 дней у мышей).[13]). Во время профазы I пара гомологичных материнских и отцовских хромосом, синапс, и обмениваться генетической информацией ( гомологичная рекомбинация ), образуя по крайней мере один кроссовер на хромосому.[14] Эти пересечения становятся видимыми как хиазмы (множественное число; единственное число хиазма ).[15] Этот процесс способствует стабильному спариванию между гомологичными хромосомами и, следовательно, обеспечивает точное разделение хромосом при первом делении мейоза. Парные и реплицированные хромосомы называются бивалентами (две хромосомы) или тетрадами (четыре хроматиды ), по одной хромосоме от каждого родителя. Профаза I делится на ряд подстадий, названных в соответствии с внешним видом хромосом.
Лептотен
Первым этапом профазы I является лептотена этап, также известный как лептонема, от греческих слов, означающих «тонкие нити».[16]:27 На этой стадии профазы I отдельные хромосомы - каждая из которых состоит из двух реплицированных сестринских хроматид - становятся «индивидуализированными», образуя видимые нити внутри ядра.[16]:27[17]:353 Каждая хромосома образует линейный массив петель, опосредованных когезин, а боковые элементы синаптонемный комплекс собрать, образуя «осевой элемент», от которого отходят петли.[18] На этой стадии рекомбинация инициируется ферментом SPO11 который создает запрограммированные двухрядные разрывы (около 300 в мейозе у мышей).[19] Этот процесс генерирует однонитевые нити ДНК, покрытые RAD51 и DMC1 которые вторгаются в гомологичные хромосомы, образуя межосевые мосты и приводя к спариванию / ко-выравниванию гомологов (на расстоянии ~ 400 нм у мышей).[18][20]
Зиготена
Лептотене следует зиготена этап, также известный как зигонема, от греческих слов, означающих «парные нити»,[16]:27 который у некоторых организмов также называют стадией букета из-за того, как теломеры группируются на одном конце ядра.[21] На этой стадии гомологичные хромосомы становятся намного более близкими (~ 100 нм) и стабильно спаренными (процесс, называемый синапсисом), опосредованный установкой поперечных и центральных элементов синаптонемный комплекс.[18] Считается, что синапсис происходит по типу «молнии», начиная с узла рекомбинации. Парные хромосомы называются двухвалентными или тетрадными хромосомами.
Пахитена
В пахитены сцена (/ˈпækɪтяп/ ПАК-i-подросток ), также известный как пахинема, от греческих слов, означающих «толстые нити».[16]:27 это стадия, на которой все аутосомные хромосомы синапсируются. На этом этапе гомологичная рекомбинация, включая хромосомный кроссовер (кроссинговер), завершается за счет репарации двухцепочечных разрывов, образованных в лептотене.[18] Большинство поломок ремонтируется без образования пересечений, что приводит к преобразование гена.[22] Однако подмножество разрывов (по крайней мере, по одному на хромосому) формирует кроссоверы между несестринскими (гомологичными) хромосомами, что приводит к обмену генетической информацией.[23] Половые хромосомы, однако, не полностью идентичны и обмениваются информацией только в небольшой области гомологии, называемой псевдоавтосомальная область.[24] Обмен информацией между гомологичными хроматидами приводит к рекомбинации информации; каждая хромосома имеет полный набор информации, которая у нее была раньше, и в результате этого процесса нет пробелов. Поскольку хромосомы не могут быть различимы в синаптонемном комплексе, фактический акт кроссинговера не воспринимается через обычный световой микроскоп, а хиазмы не видны до следующей стадии.
Диплотен
Вовремя диплотена этап, также известный как Дипломема, от греческих слов, означающих "две нити",[16]:30 то синаптонемный комплекс разбирает и гомологичные хромосомы немного отделяются друг от друга. Однако гомологичные хромосомы каждого бивалента остаются прочно связанными в хиазмах, областях, где произошел кроссинговер. Хиазмы остаются на хромосомах до тех пор, пока они не разрываются при переходе в анафазу I, чтобы позволить гомологичным хромосомам перемещаться к противоположным полюсам клетки.
У человеческого плода оогенез все развивающиеся ооциты развиваются до этой стадии и задерживаются в профазе I еще до рождения.[25] Это приостановленное состояние называется диктиотеновая стадия или диктиате. Это длится до возобновления мейоза, чтобы подготовить ооцит к овуляции, которая происходит в период полового созревания или даже позже.
Диакинез
Хромосомы конденсируются дальше во время диакинез стадия, от греческих слов, означающих «продвижение».[16]:30 Это первая точка мейоза, где фактически видны четыре части тетрад. Участки кроссинговера переплетаются друг с другом, эффективно перекрываясь, делая хиазмы четко видимыми. Помимо этого наблюдения, остальная часть сцены очень похожа на прометафаза митоза; то ядрышки исчезнуть, ядерная мембрана распадается на пузырьки, и мейотическое веретено начинает формироваться.
Формирование мейотического веретена
В отличие от митотических клеток, ооциты человека и мыши не имеют центросомы для производства мейотического веретена. У мышей примерно 80 центров организации микротрубочек (MTOC) образуют сферу в ооплазме и начинают зародышеобразование микротрубочек, которые доходят до хромосом, прикрепляясь к хромосомам в кинетохора. Со временем MTOC сливаются, пока не образуются два полюса, образуя шпиндель бочкообразной формы.[26] В человеческих ооцитах зарождение микротрубочек веретена начинается на хромосомах, образуя звездочку, которая в конечном итоге расширяется, чтобы окружить хромосомы.[27] Затем хромосомы скользят по микротрубочкам к экватору веретена, и в этот момент кинетохоры хромосомы образуют прикрепления на конце к микротрубочкам.[28]
Метафаза I
Гомологические пары движутся вместе по метафазной пластине: As кинетохорные микротрубочки от обоих полюсов веретена присоединяются к своим соответствующим кинетохорам, парные гомологичные хромосомы выравниваются вдоль экваториальной плоскости, которая делит веретено пополам, из-за непрерывных уравновешивающих сил, оказываемых на биваленты микротрубочками, исходящими от двух кинетохор гомологичных хромосом. Эта насадка называется биполярной. Физическая основа независимый ассортимент хромосом - это случайная ориентация каждого бивалента вдоль метафазной пластинки относительно ориентации других бивалентов вдоль той же экваториальной линии.[15] Белковый комплекс когезин удерживает сестринские хроматиды вместе с момента их репликации до анафазы. В митозе сила вытягивания микротрубочек кинетохор в противоположных направлениях создает напряжение. Клетка ощущает это напряжение и не продвигается в анафазе до тех пор, пока все хромосомы не станут должным образом двунаправленными. В мейозе для установления напряжения обычно требуется по крайней мере один кроссовер на пару хромосом в дополнение к когезину между сестринскими хроматидами (см. Расщепление хромосом ).
Анафаза I
Микротрубочки кинетохор укорачиваются, притягивая гомологичные хромосомы (каждая из которых состоит из пары сестринских хроматид) к противоположным полюсам. Некинетохорные микротрубочки удлиняются, раздвигая центросомы дальше друг от друга. Клетка удлиняется, готовясь к делению по центру.[15] В отличие от митоза, разрушается только когезин хромосомных плеч, в то время как когезин, окружающий центромеру, остается защищенным белком, названным Shugoshin (по-японски «дух-хранитель»), что препятствует разделению сестринских хроматид.[29] Это позволяет сестринским хроматидам оставаться вместе, в то время как гомологи разделены.
Телофаза I
Первое мейотическое деление эффективно заканчивается, когда хромосомы достигают полюсов. В каждой дочерней клетке теперь половина хромосом, но каждая хромосома состоит из пары хроматид. Микротрубочки, составляющие сеть веретена, исчезают, и каждый набор гаплоидов окружает новая ядерная мембрана. Хромосомы раскручиваются обратно в хроматин. Цитокинез, защемление клеточной мембраны в клетках животных или формирование клеточной стенки в клетках растений, завершается образованием двух дочерних клеток. Однако цитокинез не завершается полностью, что приводит к образованию «цитоплазматических мостов», которые позволяют разделять цитоплазму между дочерними клетками до конца мейоза II.[30] Сестринские хроматиды остаются прикрепленными во время телофазы I.
Клетки могут войти в период покоя, известный как интеркинез или интерфаза II. На этом этапе репликации ДНК не происходит.
Мейоз II
Мейоз II - это второе мейотическое деление, обычно включающее эквациональную сегрегацию или разделение сестринских хроматид. Механически этот процесс аналогичен митозу, хотя его генетические результаты принципиально отличаются. Конечным результатом является производство четырех гаплоидных клеток (n хромосом, 23 у человека) из двух гаплоидных клеток (с n хромосомами, каждая из которых состоит из двух сестринских хроматид), образующихся в мейозе I. Четыре основных этапа мейоза II: профаза II. , метафаза II, анафаза II и телофаза II.
В профаза II, мы видим исчезновение ядрышек и ядерная оболочка опять же, а также укорачивание и утолщение хроматид. Центросомы перемещаются в полярные регионы и организуют волокна веретена для второго деления мейоза.
В метафаза IIцентромеры содержат две кинетохоры, которые прикрепляются к волокнам веретена от центросом на противоположных полюсах. Новая экваториальная метафазная пластинка повернута на 90 градусов по сравнению с мейозом I, перпендикулярно предыдущей пластине.[31]
Далее следует анафаза II, в котором оставшийся центромерный cohesin, больше не защищенный Shugoshin, расщепляется, позволяя сестринским хроматидам сегрегировать. Сестринские хроматиды по соглашению теперь называются сестринскими хромосомами, поскольку они движутся к противоположным полюсам.[29]
Процесс заканчивается телофаза II, который похож на телофазу I, и отмечен деконденсацией и удлинением хромосом и разборкой веретена. Ядерные оболочки реформируются и отщепляются, или образование клеточной пластинки в конечном итоге дает в общей сложности четыре дочерних клетки, каждая с гаплоидным набором хромосом.
Мейоз завершен, и в его результате появляются четыре новых дочерних клетки.
Происхождение и функция
В происхождение и функция мейоза в настоящее время недостаточно изучены с научной точки зрения и могут дать фундаментальное понимание эволюция полового размножения в эукариоты. В настоящее время среди биологов нет единого мнения по вопросу о том, как секс у эукариот возник в эволюция, какая основная функция половое размножение служит, и почему она поддерживается, учитывая основные двукратная стоимость секса. Ясно, что он появился более 1,2 миллиарда лет назад, и что почти все виды, которые являются потомками первоначальных видов, воспроизводящих половым путем, по-прежнему являются половыми репродукторами, включая растения, грибы, и животные.
Мейоз является ключевым событием полового цикла у эукариот. Это этап жизненный цикл когда клетка дает начало двум гаплоидным клеткам (гаметы ) каждый имеет вдвое меньше хромосомы. Две такие гаплоидные гаметы, происходящие от разных особей. организмы, заплавляются в процессе оплодотворение, завершая таким образом половой цикл.
Мейоз распространен среди эукариот. Это происходит в одноклеточных организмах, таких как дрожжи, а также в многоклеточных организмах, таких как люди. Эукариоты возникли из прокариоты более 2,2 миллиарда лет назад[32] и самые ранние эукариоты, вероятно, были одноклеточными организмами. Чтобы понять пол у эукариот, необходимо понять (1) как возник мейоз у одноклеточных эукариот и (2) функция мейоза.
Новые комбинации ДНК, созданные во время мейоза, являются значительным источником генетическая вариация наряду с мутацией, что приводит к новым комбинациям аллели, что может быть полезно. Мейоз вызывает генетическое разнообразие гамет двумя способами: (1) Закон независимого ассортимента. Независимая ориентация гомологичных пар хромосом вдоль метафазной пластинки во время метафазы I и ориентация сестринских хроматид в метафазе II, это последующее разделение гомологов и сестринских хроматид во время анафазы I и II, это позволяет случайное и независимое распределение хромосом по каждой из них. дочерняя клетка (и, в конечном итоге, гаметы);[33] и (2) Пересекая. Физический обмен гомологичными хромосомными областями путем гомологичной рекомбинации во время профазы I приводит к новым комбинациям генетической информации в хромосомах.[34]
Вхождение
В жизненных циклах


Мейоз возникает у эукариот жизненные циклы с участием половое размножение, состоящий из постоянного циклического процесса мейоза и оплодотворения. Это происходит вместе с обычным митотический деление клеток. В многоклеточных организмах существует промежуточный этап между диплоидным и гаплоидным переходом, на котором организм растет. На определенных этапах жизненного цикла стволовые клетки производят гаметы. Соматические клетки составляют тело организма и не участвуют в производстве гамет.
Чередование событий мейоза и оплодотворения вызывает серию переходов назад и вперед между чередующимися гаплоидными и диплоидными состояниями. Фаза жизненного цикла организма может происходить либо во время диплоидного состояния (дипломатический жизненный цикл), во время гаплоидного состояния (гаплонический жизненный цикл) или оба (гаплодипонтический жизненный цикл, в котором есть две отдельные фазы организма, одна во время гаплоидного состояния, а другая во время диплоидного состояния). В этом смысле существует три типа жизненных циклов, в которых используется половое воспроизводство, которые различаются расположением фазы (фаз) организма.[нужна цитата ]
в дипломатический жизненный цикл (с предгаметическим мейозом), частью которого являются люди, организм является диплоидным, вырос из диплоидной клетки, называемой зигота. Диплоидные стволовые клетки зародышевой линии организма подвергаются мейозу с образованием гаплоидных гамет ( сперматозоиды для мужчин и яйцеклетка для самок), которые удобряют, образуя зиготу. Диплоидная зигота подвергается многократному клеточному делению митоз врастать в организм.
в гаплонтический жизненный цикл (при постзиготическом мейозе) организм гаплоидный, порожденный пролиферацией и дифференцировкой одной гаплоидной клетки, называемой гамета. Два организма противоположного пола вносят свои гаплоидные гаметы в диплоидную зиготу. Зигота немедленно подвергается мейозу, образуя четыре гаплоидных клетки. Эти клетки подвергаются митоз создать свой организм. Много грибы и много простейшие использовать гаплонтический жизненный цикл.[нужна цитата ]
Наконец, в гаплодипонтический жизненный цикл (при споровом или промежуточном мейозе) в живом организме чередуются гаплоидные и диплоидные состояния. Следовательно, этот цикл также известен как смена поколений. Клетки зародышевой линии диплоидного организма подвергаются мейозу с образованием спор. Споры размножаются митозом, превращаясь в гаплоидный организм. Затем гамета гаплоидного организма объединяется с гаметой другого гаплоидного организма, создавая зиготу. Зигота подвергается повторным митозам и дифференцировке, чтобы снова стать диплоидным организмом. Гаплодипонтический жизненный цикл можно рассматривать как слияние диплонтического и гаплонтического жизненных циклов.[35][нужна цитата ]
У растений и животных
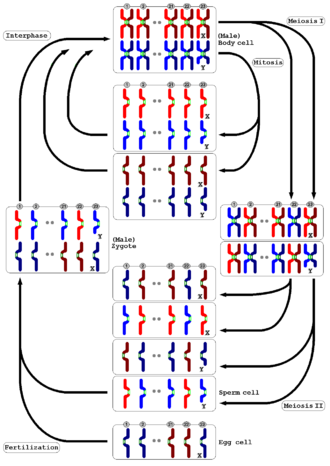
Мейоз встречается у всех животных и растений. Конечный результат, производство гамет с половиной количества хромосом по сравнению с родительской клеткой, такой же, но подробный процесс отличается. У животных мейоз непосредственно производит гаметы. У наземных растений и некоторых водорослей есть смена поколений такой, что мейоз в диплоиде спорофит поколение производит гаплоидные споры. Эти споры размножаются митозом, превращаясь в гаплоид. гаметофит поколение, которое затем дает начало гаметам напрямую (то есть без дальнейшего мейоза). И у животных, и у растений заключительный этап - это слияние гамет, восстанавливающее исходное количество хромосом.[36]
У млекопитающих
У женщин мейоз происходит в клетках, известных как ооциты (единственное число: ооцит). Каждый первичный ооцит делится дважды в мейозе, в каждом случае неравномерно. Первое деление дает дочернюю клетку и полярное тело гораздо меньшего размера, которое может или не может подвергаться второму делению. В мейозе II деление дочерней клетки дает второе полярное тельце и одну гаплоидную клетку, которая увеличивается в размерах и становится яйцеклетка. Следовательно, у самок каждый первичный ооцит, претерпевающий мейоз, дает одну зрелую яйцеклетку и одно или два полярных тельца.
Обратите внимание, что во время мейоза у самок бывают паузы. Созревающие ооциты задерживаются в профазе I мейоза I и бездействуют в защитной оболочке соматических клеток, называемой фолликул. В начале каждого менструальный цикл, ФСГ секреция передней доли гипофиза стимулирует созревание нескольких фолликулов в процессе, известном как фолликулогенез. Во время этого процесса созревающие ооциты возобновляют мейоз и продолжают до метафазы II мейоза II, где они снова задерживаются непосредственно перед овуляцией. Если эти ооциты оплодотворены спермой, они возобновят и завершат мейоз. Во время фолликулогенеза у человека обычно один фолликул становится доминирующим, в то время как другие подвергаются атрезия. Процесс мейоза у самок происходит во время оогенез, и отличается от типичного мейоза тем, что имеет длительный период мейотической остановки, известный как диктовать стадии и испытывает недостаток в помощи центросомы.[37][38]
У мужчин мейоз возникает во время сперматогенез в семенные канальцы из яички. Мейоз во время сперматогенеза специфичен для типа клеток, называемых сперматоциты, который позже созреет, чтобы стать сперматозоиды. Мейоз примордиальных половых клеток происходит в период полового созревания, значительно позже, чем у женщин. Ткани мужских семенников подавляют мейоз за счет разложения ретиноевой кислоты, которая, как предполагается, является стимулятором мейоза. Это преодолевается в период полового созревания, когда клетки семенных канальцев, называемые клетками Сертоли, начинают вырабатывать собственную ретиноевую кислоту. Чувствительность к ретиноевой кислоте также регулируется белками, называемыми нано и DAZL.[39][40] Исследования генетической потери функции ферментов, генерирующих ретиноевую кислоту, показали, что ретиноевая кислота необходима постнатально для стимуляции дифференцировки сперматогониев, которая через несколько дней проявляется в сперматоцитах, подвергающихся мейозу, однако ретиноевая кислота не требуется в то время, когда начинается мейоз.[41]
В самки млекопитающих, мейоз начинается сразу после миграции примордиальных половых клеток в яичник у эмбриона. Некоторые исследования предполагают, что ретиноевая кислота, полученная из примитивной почки (мезонефрос), стимулирует мейоз в эмбриональных яичниковых оогониях, а ткани эмбриональных мужских семенников подавляют мейоз за счет разложения ретиноевой кислоты.[42] Однако исследования генетической потери функции ферментов, генерирующих ретиноевую кислоту, показали, что ретиноевая кислота не требуется для инициации мейоза у самок, который происходит во время эмбриогенеза.[43] или мужской мейоз, который возникает постнатально.[41]
Вариации
Нерасхождение
Нормальное разделение хромосом в мейозе I или сестринских хроматид в мейозе II называется дизъюнкция. Когда сегрегация ненормальна, это называется нерасхождение. Это приводит к образованию гамет, которые имеют либо слишком много, либо слишком мало определенной хромосомы, и является обычным механизмом для трисомия или же моносомия. Нерасхождение может происходить в мейозе I или мейозе II, фазах клеточного воспроизводства или во время митоз.
Большинство моносомных и трисомных человеческих эмбрионов нежизнеспособны, но некоторые анеуплоидии могут переноситься, например, трисомия по наименьшей хромосоме, хромосоме 21. Фенотипы этих анеуплоидий варьируются от тяжелых нарушений развития до бессимптомных. Медицинские условия включают, но не ограничиваются:
- Синдром Дауна - трисомия хромосомы 21
- Синдром Патау - трисомия хромосомы 13
- Синдром Эдвардса - трисомия 18 хромосомы
- Синдром Клайнфельтера - дополнительные Х-хромосомы у мужчин - например, XXY, XXXY, XXXXY и т. Д.
- Синдром Тернера - отсутствие одной Х-хромосомы у женщин - т.е. X0
- Синдром тройного Х - лишняя Х-хромосома у женщин
- Синдром Джейкобса - лишняя Y-хромосома у мужчин.
Вероятность нерасхождения ооцитов человека увеличивается с увеличением возраста матери.[44] предположительно из-за потери когезин через некоторое время.[45]
Другой
Наряду с вариациями мейоза, связанными с моментом возникновения мейоза в жизненных циклах, что приводит к постзиготическому, предгаметическому и промежуточному мейозу (см. Выше), количество ядерных делений в мейозе также варьируется. У большинства эукариот мейоз состоит из двух частей (хотя иногда ахиазматический ), но очень редкая форма - однораздельный мейоз - встречается у некоторых жгутиконосцев (парабазалиды и оксимонады ) из кишечника лесного таракана Криптоцерк.[46]
Сравнение с митозом
Для понимания мейоза полезно сравнение с митозом. В таблице ниже показаны различия между мейозом и митозом.[47]
| Мейоз | Митоз | |
|---|---|---|
| Конечный результат | Обычно четыре клетки, каждая с половиной хромосом в каждой из них. | Две клетки, имеющие такое же количество хромосом, что и родительская |
| Функция | Производство гамет (половых клеток) у эукариот, размножающихся половым путем, с диплонтовым жизненным циклом | Клеточное размножение, рост, ремонт, бесполое размножение |
| Где это происходит? | Практически все эукариоты (животные, растения, грибы и протисты );[48][46] В гонадах, до гамет (в диплонтических жизненных циклах); После зиготы (в гаплонтике); Перед спорами (при гаплодипонтике) | Все пролиферирующие клетки у всех эукариот |
| Шаги | Профаза I, Метафаза I, Анафаза I, Телофаза I, Профаза II, Метафаза II, Анафаза II, Телофаза II | Профаза, Прометафаза, Метафаза, Анафаза, Телофаза |
| Генетически такой же, как родитель? | Нет | да |
| Переход происходит? | Да, обычно встречается между каждой парой гомологичных хромосом | Очень редко |
| Спаривание гомологичных хромосом? | да | Нет |
| Цитокинез | Встречается в телофазе I и телофазе II. | Occurs in Telophase |
| Centromeres split | Does not occur in Anaphase I, but occurs in Anaphase II | Occurs in Anaphase |
Molecular regulation
Эта секция нуждается в расширении. Вы можете помочь добавляя к этому. (Август 2020 г.) |
How a cell proceeds to meiotic division in meiotic cell division is not well known. Maturation promoting factor (MPF) seemingly have role in frog Oocyte meiosis. In the fungus С. Помбе. there is a role of MeiRNA binding protein for entry to meiotic cell division [49]
It has been suggested that Yeast CEP1 gene product, that binds centromeric region CDE1, may play a role in chromosome pairing during meiosis-I [50]
Meiotic recombination is mediated through double stranded break, which is catalyzed by spo11 protein. Also Mre11, sae2 and exo1 play role in breakage and recombination. After the breakage happen, recombination take place which is typically homologous. The recombination may go through either a double Holliday junction (dHJ) pathway or synthesis-dependent strand annealing (SDSA). (The second one gives to noncrossover product) [51]
Seemingly there are checkpoints for meiotic cell division too. In S. pombe, Rad proteins, S. pombe Mek1 (with FHA kinase domain), Cdc25, Cdc2 and unknown factor is thought to form a checkpoint [52]
In vertebrate oogenesis, maintained by cytostatic factor (CSF) has role in switching into meiosis-II.[50]
Смотрите также
Рекомендации
- ^ а б Freeman, Scott (2011). Биологические науки (6-е изд.). Hoboken, NY: Pearson. п. 210.
- ^ Hassold T, Hunt P (April 2001). "To err (meiotically) is human: the genesis of human aneuploidy". Природа Обзоры Генетика. 2 (4): 280–91. Дои:10.1038/35066065. PMID 11283700. S2CID 22264575.
- ^ Letunic I, Bork P (2006). "Interactive Tree of Life". В архиве из оригинала 29 января 2018 г.. Получено 23 июля 2011.
- ^ Bernstein H, Bernstein C (2010). "Evolutionary origin of recombination during meiosis". Бионаука. 60 (7): 498–505. Дои:10.1525/bio.2010.60.7.5. S2CID 86663600.
- ^ Lodé T (June 2011). "Sex is not a solution for reproduction: the libertine bubble theory". BioEssays. 33 (6): 419–22. Дои:10.1002/bies.201000125. PMID 21472739.
- ^ J.B. Farmer and J.E.S. Мур, Quarterly Journal of Microscopic Science 48:489 (1905) as quoted in the Оксфордский словарь английского языка, Third Edition, June 2001, s.v.
- ^ Battaglia E. (1985). Meiosis and mitosis: a terminological criticism. Энн Бот (Rome) 43: 101–140. связь.
- ^ "Mitosis". 2012-10-27. Архивировано из оригинал на 2012-10-27. Получено 2018-02-09.
- ^ Zhou A, Pawlowski WP (August 2014). "Regulation of meiotic gene expression in plants". Границы науки о растениях. 5: 413. Дои:10.3389/fpls.2014.00413. ЧВК 4142721. PMID 25202317.
- ^ Jung, Min; Wells, Daniel; Rusch, Jannette; Ahmad, Suhaira; Marchini, Jonathan; Myers, Simon R; Conrad, Donald F (2019-06-25). "Unified single-cell analysis of testis gene regulation and pathology in five mouse strains". eLife. 8: e43966. Дои:10.7554/eLife.43966. ISSN 2050-084X. ЧВК 6615865. PMID 31237565.
- ^ Брар Г.А., Яссур М., Фридман Н., Регев А., Инголия Н.Т., Вайсман Дж.С. (февраль 2012 г.). «Изображение с высоким разрешением мейотической программы дрожжей, выявленное с помощью профилирования рибосом». Наука. 335 (6068): 552–7. Bibcode:2012Sci ... 335..552B. Дои:10.1126 / science.1215110. ЧВК 3414261. PMID 22194413.
- ^ Фримен 2005, pp. 244–45
- ^ Cohen, P. E.; Pollack, S. E.; Pollard, J. W. (2006-06-01). "Genetic Analysis of Chromosome Pairing, Recombination, and Cell Cycle Control during First Meiotic Prophase in Mammals". Эндокринные обзоры. 27 (4): 398–426. Дои:10.1210/er.2005-0017. ISSN 0163-769X. PMID 16543383.
- ^ Hunter, Neil (2015-10-28). "Meiotic Recombination: The Essence of Heredity". Перспективы Колд-Спринг-Харбор в биологии. 7 (12): a016618. Дои:10.1101/cshperspect.a016618. ISSN 1943-0264. ЧВК 4665078. PMID 26511629.
- ^ а б c Фримен 2005, стр. 249–250
- ^ а б c d е ж Snustad DP, Simmons MJ (December 2008). Principles of Genetics (5-е изд.). Вайли. ISBN 978-0-470-38825-9.
- ^ Krebs JE, Goldstein ES, Kilpatrick ST (November 2009). Lewin's Genes X (10-е изд.). Jones & Barlett Learning. ISBN 978-0-7637-6632-0.
- ^ а б c d Zickler, Denise; Kleckner, Nancy (June 2015). "Recombination, Pairing, and Synapsis of Homologs during Meiosis". Перспективы Колд-Спринг-Харбор в биологии. 7 (6): a016626. Дои:10.1101/cshperspect.a016626. ISSN 1943-0264. ЧВК 4448610. PMID 25986558.
- ^ Baudat, Frédéric; de Massy, Bernard (July 2007). "Regulating double-stranded DNA break repair towards crossover or non-crossover during mammalian meiosis". Хромосомные исследования. 15 (5): 565–577. Дои:10.1007/s10577-007-1140-3. ISSN 0967-3849. PMID 17674146. S2CID 26696085.
- ^ "Stages of Meiosis and Sexual Reproduction | Learn Science at Scitable". www.nature.com. Получено 2020-10-18.
- ^ Link, Jana; Jantsch, Verena (2019-03-15). "Meiotic chromosomes in motion: a perspective from Mus musculus and Caenorhabditis elegans". Хромосома. 128 (3): 317–330. Дои:10.1007/s00412-019-00698-5. ISSN 0009-5915. ЧВК 6823321. PMID 30877366.
- ^ Чен, Цзянь-Минь; Купер, Дэвид Н .; Чужанова, Надя; Ферек, Клод; Patrinos, George P. (October 2007). "Gene conversion: mechanisms, evolution and human disease". Природа Обзоры Генетика. 8 (10): 762–775. Дои:10.1038/nrg2193. ISSN 1471-0056. PMID 17846636. S2CID 205484180.
- ^ Bolcun-Filas, Ewelina; Handel, Mary Ann (2018-07-01). "Meiosis: the chromosomal foundation of reproduction". Биология размножения. 99 (1): 112–126. Дои:10.1093/biolre/ioy021. ISSN 0006-3363. PMID 29385397. S2CID 38589675.
- ^ Hinch, Anjali G.; Altemose, Nicolas; Noor, Nudrat; Доннелли, Питер; Myers, Simon R. (July 2014). "Recombination in the human Pseudoautosomal region PAR1". PLOS Genetics. 10 (7): e1004503. Дои:10.1371/journal.pgen.1004503. ISSN 1553-7404. ЧВК 4102438. PMID 25033397.
- ^ 1950-, Nussbaum, Robert L. (2015-05-21). Thompson & Thompson genetics in medicine. McInnes, Roderick R.,, Willard, Huntington F.,, Hamosh, Ada,, Preceded by: Nussbaum, Robert L., 1950- (Eighth ed.). Филадельфия, Пенсильвания. п. 19. ISBN 978-1437706963. OCLC 908336124.CS1 maint: числовые имена: список авторов (связь)
- ^ Schuh, Melina; Ellenberg, Jan (2007-08-10). "Self-Organization of MTOCs Replaces Centrosome Function during Acentrosomal Spindle Assembly in Live Mouse Oocytes". Клетка. 130 (3): 484–498. Дои:10.1016/j.cell.2007.06.025. ISSN 0092-8674. PMID 17693257. S2CID 5219323.
- ^ Holubcova, Zuzana; Blayney, Martyn; Elder, Kay; Schuh, Melina (2015-06-05). "Error-prone chromosome-mediated spindle assembly favors chromosome segregation defects in human oocytes". Наука. 348 (6239): 1143–1147. Bibcode:2015Sci...348.1143H. Дои:10.1126/science.aaa9529. ISSN 0036-8075. ЧВК 4477045. PMID 26045437.
- ^ Kitajima, Tomoya S.; Ohsugi, Miho; Ellenberg, Jan (2011-08-19). "Complete Kinetochore Tracking Reveals Error-Prone Homologous Chromosome Biorientation in Mammalian Oocytes". Клетка. 146 (4): 568–581. Дои:10.1016/j.cell.2011.07.031. ISSN 0092-8674. PMID 21854982. S2CID 5637615.
- ^ а б Pierce, Benjamin (2009). «Chromosomes and Cell Reproduction». Genetics: A Conceptual Approach, Third Edition. W.H. FREEMAN AND CO. ISBN 9780716779285 P. 32
- ^ Haglund, Kaisa; Nezis, Ioannis P.; Stenmark, Harald (2011-01-01). "Structure and functions of stable intercellular bridges formed by incomplete cytokinesis during development". Коммуникативная и интегративная биология. 4 (1): 1–9. Дои:10.4161/cib.13550. ЧВК 3073259. PMID 21509167.
- ^ "BioCoach Activity: Concept 11: Meiosis II: Metaphase II". The Biology Place. Пирсон. В архиве from the original on 2018-02-28. Получено 2018-02-10.
- ^ Retallack GJ, Krull ES, Thackeray GD, Parkinson D (2013-09-01). "Problematic urn-shaped fossils from a Paleoproterozoic (2.2 Ga) paleosol in South Africa". Докембрийские исследования. 235: 71–87. Bibcode:2013PreR..235...71R. Дои:10.1016/j.precamres.2013.05.015.
- ^ Monaghan F, Corcos A (1984-01-01). "On the origins of the Mendelian laws". Журнал наследственности. 75 (1): 67–9. Дои:10.1093/oxfordjournals.jhered.a109868. PMID 6368675.
- ^ Saleem, Muhammad (2001). "Inherited Differences in Crossing Over and Gene Conversion Frequencies Between Wild Strains of Sordaria fimicola From "Evolution Canyon"". Генетика. 159. В архиве из оригинала от 03.03.2017. Получено 2017-03-02.
- ^ South GR, Whittick A (2009-07-08). An Introduction to Phycology. Джон Вили и сыновья. ISBN 9781444314205.
- ^ Bidlack, James E. (2011). Introductory Plant Biology. New York, NY: McGraw HIll. pp. 214–29.
- ^ Brunet S, Verlhac MH (2010). "Positioning to get out of meiosis: the asymmetry of division". Обновление репродукции человека. 17 (1): 68–75. Дои:10.1093/humupd/dmq044. PMID 20833637. S2CID 13988521.
- ^ Rosenbusch B (November 2006). "The contradictory information on the distribution of non-disjunction and pre-division in female gametes". Репродукция человека. 21 (11): 2739–42. Дои:10.1093/humrep/del122. PMID 16982661.
- ^ Lin Y, Gill ME, Koubova J, Page DC (December 2008). "Germ cell-intrinsic and -extrinsic factors govern meiotic initiation in mouse embryos". Наука. 322 (5908): 1685–7. Bibcode:2008Sci...322.1685L. Дои:10.1126/science.1166340. PMID 19074348. S2CID 11261341.
- ^ Suzuki A, Saga Y (February 2008). "Nanos2 suppresses meiosis and promotes male germ cell differentiation". Гены и развитие. 22 (4): 430–5. Дои:10.1101/gad.1612708. ЧВК 2238665. PMID 18281459.
- ^ а б Teletin M, Vernet N, Yu J, Klopfenstein M, Jones JW, Féret B, Kane MA, Ghyselinck NB, Mark M (January 2019). "Two functionally redundant sources of retinoic acid secure spermatogonia differentiation in the seminiferous epithelium". Разработка. 146 (1): dev170225. Дои:10.1242/dev.170225. ЧВК 6340151. PMID 30487180.
- ^ Bowles J, Knight D, Smith C, Wilhelm D, Richman J, Mamiya S, Yashiro K, Chawengsaksophak K, Wilson MJ, Rossant J, Hamada H, Koopman P (April 2006). "Retinoid signaling determines germ cell fate in mice". Наука. 312 (5773): 596–600. Bibcode:2006Sci...312..596B. Дои:10.1126/science.1125691. PMID 16574820. S2CID 2514848.
- ^ Kumar S, Chatzi C, Brade T, Cunningham TJ, Zhao X, Duester G (January 2011). "Sex-specific timing of meiotic initiation is regulated by Cyp26b1 independent of retinoic acid signalling". Nature Communications. 2 (1): 151. Bibcode:2011NatCo...2..151K. Дои:10.1038/ncomms1136. ЧВК 3034736. PMID 21224842.
- ^ Hassold T, Jacobs P, Kline J, Stein Z, Warburton D (July 1980). "Effect of maternal age on autosomal trisomies". Анналы генетики человека. 44 (1): 29–36. Дои:10.1111/j.1469-1809.1980.tb00943.x. PMID 7198887. S2CID 24469567.
- ^ Tsutsumi M, Fujiwara R, Nishizawa H, Ito M, Kogo H, Inagaki H, Ohye T, Kato T, Fujii T, Kurahashi H (May 2014). "Age-related decrease of meiotic cohesins in human oocytes". PLOS ONE. 9 (5): e96710. Bibcode:2014PLoSO...996710T. Дои:10.1371/journal.pone.0096710. ЧВК 4013030. PMID 24806359.
- ^ а б Raikov, I. B. (1995). "Meiosis in protists: recent advances and persisting problems". Europ J Protistol. 31: 1–7. Дои:10.1016/s0932-4739(11)80349-4.
- ^ "How Cells Divide". PBS. Служба общественного вещания. В архиве из оригинала 1 ноября 2012 г.. Получено 6 декабря 2012.
- ^ Heywood P, Magee PT (March 1976). "Meiosis in protists. Some structural and physiological aspects of meiosis in algae, fungi, and protozoa". Бактериологические обзоры. 40 (1): 190–240. Дои:10.1128/mmbr.40.1.190-240.1976. ЧВК 413949. PMID 773364.
- ^ Molecular Regulation of the Mitosis/Meiosis Decision in Multicellular Organisms, by Judith Kimble, Cold Spring Harbor perspectives in biology, 3(8), a002683. https://doi.org/10.1101/cshperspect.a002683 , NCBI-PMC
- ^ а б Regulatory mechanisms in meiosis, by Honigberg, M.McCarroll, Esposito; https://doi.org/10.1016/0955-0674(93)90106-Z, Sciencedirect
- ^ Lam, I., & Keeney, S. (2014). Mechanism and regulation of meiotic recombination initiation. Cold Spring Harbor perspectives in biology, 7(1), a016634. https://doi.org/10.1101/cshperspect.a016634
- ^ Regulation of meiotic progression by the meiosis-specific checkpoint kinase Mek1 in fission yeast, by Livia Pérez-Hidalgo, Sergio Moreno, Pedro A. San-Segundo, Journal of Cell Science 2003 116: 259-271; doi: 10.1242/jcs.00232, https://jcs.biologists.org/content/116/2/259#:~:text=Checkpoints%20are%20regulatory%20mechanisms%20that,at%20the%20first%20meiotic%20division.
Цитированные тексты
- Freeman, Scott (2005). Биологические науки (3-е изд.). Река Аппер Сэдл, штат Нью-Джерси: Pearson Prentice Hall.
внешняя ссылка
- Meiosis Flash Animation
- Animations from the U. of Arizona Biology Dept.
- Meiosis at Kimball's Biology Pages
- Khan Academy, video lecture
- CCO The Cell-Cycle Ontology
- Stages of Meiosis animation
- *"Abby Dernburg Seminar: Chromosome Dynamics During Meiosis"
| Биологические термины | |
|---|---|
| Сексуальный reproduction | |
| Сексуальность | |
| |