Центромера - Википедия - Centromere

В центромера это специализированная последовательность ДНК хромосома что связывает пару сестринские хроматиды (диада).[1] В течение митоз, волокна веретена прикрепить к центромере через кинетохора.[2] Центромеры сначала считались генетические локусы которые направляют поведение хромосомы.
Физическая роль центромеры заключается в том, чтобы действовать как место сборки кинетохоры - очень сложная мультипротеиновая структура, отвечающая за реальные события сегрегация хромосом - т.е. обязательная микротрубочки и передача сигналов аппарату клеточного цикла, когда все хромосомы правильно прикреплены к шпиндель, так что это безопасно для деление клеток перейти к завершению и для ввода ячеек анафаза.[3]
Вообще говоря, существует два типа центромер. «Точечные центромеры» связываются со специфическими белки которые признают особые ДНК последовательности с высоким КПД.[4] Любой фрагмент ДНК с точечной последовательностью ДНК центромеры на нем обычно образует центромеру, если он присутствует у соответствующих видов. Лучше всего охарактеризованы точечные центромеры у почкующихся дрожжей, Saccharomyces cerevisiae. «Региональные центромеры» - это термин, придуманный для описания большинства центромер, которые обычно образуются на участках предпочтительной последовательности ДНК, но могут также образовываться на других последовательностях ДНК.[4] Сигналом к формированию региональной центромеры, по-видимому, является эпигенетический. Большинство организмов, начиная от делящихся дрожжей Schizosaccharomyces pombe людям имеют региональные центромеры.
Что касается структуры митотической хромосомы, центромеры представляют собой суженную область хромосомы (часто называемую первичным сужением), где два идентичных сестринские хроматиды находятся в наиболее тесном контакте. Когда клетки входят в митоз, сестринские хроматиды (две копии каждой молекулы хромосомной ДНК, полученные в результате Репликация ДНК в форме хроматина) связаны по своей длине действием когезин сложный. Сейчас считается, что этот комплекс в основном высвобождается из плеч хромосом во время профазы, так что к тому времени, когда хромосомы выстраиваются в линию в средней плоскости митотического веретена (также известной как метафазная пластинка), последнее место, где они связаны друг с другом находятся в хроматине внутри и вокруг центромеры.[5]
Позиция
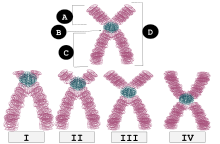
| я | Телоцентрический | Центромеры расположены очень близко к верхушке, p руки едва видны, если видны вообще. |
| II | Акроцентрический | Плечи q по-прежнему намного длиннее, чем плечи p, но плечи p длиннее, чем у телоцентрических. |
| III | Субметацентрический | Плечи p и q очень близки по длине, но не равны. |
| IV | Метацентрический | Плечи p и q равны по длине. |
B: Центромера
C: Длинная рука (q рука)
D: Сестринские хроматиды
Каждая хромосома имеет два плеча, помеченных п (короче из двух) и q (чем дольше). Многие помнят, что короткая рука «p» названа в честь французского слова «petit», означающего «маленький», хотя это объяснение оказалось апокрифическим.[6] Они могут быть соединены метацентрическим, субметацентрическим, акроцентрическим или телецентрическим образом.[7][8]
| Категоризация хромосом по относительной длине плеч[8] | ||||||
| Положение центромеры | Соотношение длины рук | Знак | Описание | |||
| Медиальный Sensu stricto | 1.0 – 1.6 | M | Метацентрический | |||
| Медиальная область | 1.7 | м | Метацентрический | |||
| Submedial | 3.0 | см | Субметацентрический | |||
| Субтерминал | 3.1 – 6.9 | ул | Субтелоцентрический | |||
| Терминальный регион | 7.0 | т | Акроцентрический | |||
| Терминал Sensu stricto | ∞ | Т | Телоцентрический | |||
| Примечания | – | Метацентрический: M+м | Ателоцентрический: M+м+см+ул+т | |||
Метацентрический
Это Х-образные хромосомы с центромерой посередине, так что два плеча хромосом почти равны.
Хромосома является метацентрической, если ее два плеча примерно равны по длине. У нормального человека кариотип, пять хромосом считаются метацентрическими: хромосомы 1, 3, 16, 19 и 20. В некоторых случаях метацентрическая хромосома образуется в результате сбалансированной транслокации: слияния двух акроцентрический хромосомы с образованием одной метацентрической хромосомы.[9][10]
Субметацентрический
Если длины плеч неравны, хромосома считается субметацентрической. Они имеют Г-образную форму.[11]
Акроцентрический
Если p (короткое) плечо настолько короткое, что его трудно наблюдать, но оно все еще присутствует, тогда хромосома акроцентрическая ("акро- "в акроцентрическом" относится к греческому слову "пик"). человеческий геном включает пять акроцентрических хромосом: 13, 14, 15, 21, 22.[12] В Y-хромосома также акроцентрический.[12]
В акроцентрической хромосоме p-плечо содержит генетический материал, включая повторяющиеся последовательности, такие как ядрышковые организующие области, и может быть перемещено без значительного вреда, как в сбалансированном Робертсоновская транслокация. В домашняя лошадь геном включает одну метацентрическую хромосому, которая гомологичный двум акроцентрическим хромосомам в сородич но неприрученный Лошадь Пржевальского.[13] Это может отражать либо фиксацию сбалансированной робертсоновской транслокации у домашних лошадей, либо, наоборот, фиксацию деления одной метацентрической хромосомы на две акроцентрические хромосомы у лошадей Пржевальского. Похожая ситуация существует между геномами человека и великой обезьяны, с сокращением двух акроцентрических хромосом у человекообразных обезьян до одной метацентрической хромосомы у человека (см. анеуплоидия и хромосома 2 человека ).[11]
Поразительно, что вредные транслокации в контексте заболевания, особенно несбалансированные транслокации при раке крови, чаще затрагивают акроцентрические хромосомы, чем неакроцентрические хромосомы.[14] Хотя причина неизвестна, вероятно, это связано с физическим расположением акроцентрических хромосом в ядро. Акроцентрические хромосомы обычно расположены внутри и вокруг ядрышко, поэтому в центре ядра, где хромосомы имеют тенденцию быть менее плотно упакованными, чем хромосомы на периферии ядра.[15] Соответственно, менее плотно упакованные хромосомные области также более склонны к хромосомным транслокациям при раке.[14]
Телоцентрический
Центрера телоцентрической хромосомы расположена на конце хромосомы. Следовательно, у телоцентрической хромосомы только одно плечо. Теломеры могут отходить от обоих концов хромосомы, их форма похожа на букву «i» в анафазе. Например, стандартный домовая мышь кариотип имеет только телецентрические хромосомы.[16][17] У людей нет телецентрических хромосом.
Субтелоцентрический
Если центромера хромосомы расположена ближе к ее концу, чем к ее центру, ее можно назвать субтелоцентрической.[18][19]
Число центромеры
Ацентрический
Если в хромосоме отсутствует центромера, говорят: ацентрический. В макронуклеус из инфузории например, содержит сотни ацентрических хромосом.[20] События разрушения хромосом также могут генерировать ацентрические хромосомы или ацентрические фрагменты.
Дицентрик
А дицентрическая хромосома - аномальная хромосома с двумя центромерами. Он образуется в результате слияния двух сегментов хромосомы, каждый с центромерой, что приводит к потере ацентрических фрагментов (без центромеры) и образованию дицентрических фрагментов.[21] Образование дицентрических хромосом связывают с генетическими процессами, такими как Робертсоновская транслокация[12] и парацентрическая инверсия.[22] Дицентрические хромосомы играют важную роль в митотической стабильности хромосом и формировании псевдодицентрических хромосом.[23]
Моноцентрический
В моноцентрический хромосома - это хромосома, которая имеет только одну центромеру в хромосоме и образует узкую перетяжку.
Моноцентрические центромеры являются наиболее распространенной структурой ДНК растений и животных с большим числом повторений.[24]
Голоцентрический
В отличие от моноцентрических хромосом в холоцентрических хромосомах вся длина хромосомы действует как центромера. В холоцентрических хромосомах нет одного первичного сужения, но центромера имеет много CenH3 локусы распространяются по всей хромосоме.[25] Примеры центромеры этого типа можно найти повсюду в царствах растений и животных.[26] наиболее известным примером является нематода Caenorhabditis elegans.
Полицентрический
Хромосомы человека
| Хромосома | Центромера позиция (Мбит / с ) | Категория | Хромосома Размер (Мбит / с) | Центромера размер (Мбайт) |
|---|---|---|---|---|
| 1 | 125.0 | метацентрический | 247.2 | 7.4 |
| 2 | 93.3 | субметацентрический | 242.8 | 6.3 |
| 3 | 91.0 | метацентрический | 199.4 | 6.0 |
| 4 | 50.4 | субметацентрический | 191.3 | — |
| 5 | 48.4 | субметацентрический | 180.8 | — |
| 6 | 61.0 | субметацентрический | 170.9 | — |
| 7 | 59.9 | субметацентрический | 158.8 | — |
| 8 | 45.6 | субметацентрический | 146.3 | — |
| 9 | 49.0 | субметацентрический | 140.4 | — |
| 10 | 40.2 | субметацентрический | 135.4 | — |
| 11 | 53.7 | субметацентрический | 134.5 | — |
| 12 | 35.8 | субметацентрический | 132.3 | — |
| 13 | 17.9 | акроцентрический | 114.1 | — |
| 14 | 17.6 | акроцентрический | 106.3 | — |
| 15 | 19.0 | акроцентрический | 100.3 | — |
| 16 | 36.6 | метацентрический | 88.8 | — |
| 17 | 24.0 | субметацентрический | 78.7 | — |
| 18 | 17.2 | субметацентрический | 76.1 | — |
| 19 | 26.5 | метацентрический | 63.8 | — |
| 20 | 27.5 | метацентрический | 62.4 | — |
| 21 | 13.2 | акроцентрический | 46.9 | — |
| 22 | 14.7 | акроцентрический | 49.5 | — |
| Икс | 60.6 | субметацентрический | 154.9 | — |
| Y | 12.5 | акроцентрический | 57.7 | — |
Последовательность
Есть два типа центромер.[27] В региональных центромерах ДНК последовательности вносят вклад в функцию, но не определяют ее. Региональные центромеры содержат большое количество ДНК и часто упаковываются в гетерохроматин. В большинстве эукариоты последовательность ДНК центромеры состоит из больших массивов повторяющейся ДНК (например, спутниковая ДНК ), где последовательность в отдельных повторяющихся элементах аналогична, но не идентична. У человека первичная центромерная повторяющаяся единица называется α-сателлитом (или альфоидом), хотя в этой области обнаруживается ряд других типов последовательностей.[28]
Точечные центромеры меньше и компактнее. Последовательности ДНК необходимы и достаточны для определения идентичности и функции центромеры у организмов с точечными центромерами. У почкующихся дрожжей область центромеры относительно мала (около 125 п.н. ДНК) и содержит две высококонсервативные последовательности ДНК, которые служат сайтами связывания для основных кинетохора белки.[28]
Наследование
Поскольку центромерная последовательность ДНК не является ключевым детерминантом центромерной идентичности в многоклеточные животные считается, что эпигенетическая наследственность играет важную роль в определении центромеры.[29] Дочерние хромосомы будут собирать центромеры в том же месте, что и родительская хромосома, независимо от последовательности. Было высказано предположение, что вариант гистона H3 CENP-A (Центромерный протеин А) - эпигенетическая метка центромеры.[30] Возникает вопрос, должен ли существовать какой-то оригинальный способ спецификации центромеры, даже если он впоследствии будет размножен эпигенетически. Если центромера наследуется эпигенетически от одного поколения к другому, проблема отодвигается назад к происхождению первых многоклеточных животных.
Структура
Центромерная ДНК обычно находится в гетерохроматин государство, что важно для набора когезин комплекс, который обеспечивает сцепление сестринских хроматид после репликации ДНК, а также координирует разделение сестринских хроматид во время анафазы. В этом хроматине нормальный гистон H3 заменен центромер-специфическим вариантом CENP-A у человека.[31] Считается, что присутствие CENP-A важно для сборки кинетохоры на центромере. Было показано, что CENP-C локализуется почти исключительно в этих областях CENP-A-ассоциированного хроматина. В клетках человека гистоны наиболее обогащены H4K20me 3 и H3K9me3[32] которые являются известными гетерохроматическими модификациями. У Drosophila островки ретроэлементов являются основными компонентами центромер.[33]
В дрожжах Schizosaccharomyces pombe (и, вероятно, у других эукариот) образование центромерного гетерохроматина связано с РНКи.[34] У нематод, таких как Caenorhabditis elegans, у некоторых растений и отрядов насекомых Lepidoptera и Hemiptera хромосомы являются «голоцентрическими», что указывает на отсутствие первичного участка прикрепления микротрубочек или первичного сужения, а «диффузная» кинетохора собирается по всей длине хромосомы.
Центромерные аберрации
В редких случаях неоцентромеры могут образовываться на новых участках хромосомы в результате репозиции центромеры. Это явление наиболее хорошо известно из клинических исследований на людях, и в настоящее время известно более 90 неоцентромеров человека, идентифицированных на 20 различных хромосомах.[35][36] Образование неоцентромеры должно быть связано с инактивацией предыдущей центромеры, поскольку хромосомы с двумя функциональными центромерами (Дицентрическая хромосома ) приведет к поломке хромосом во время митоза. В некоторых необычных случаях неоцентромеры человека спонтанно образуются на фрагментированных хромосомах. Некоторые из этих новых позиций изначально были эухроматическими и вообще лишены альфа-сателлитной ДНК. Неоцентромеры Отсутствие повторяющейся структуры, наблюдаемой в нормальных центромерах, что позволяет предположить, что образование центромер в основном контролируется эпигенетически.[37][38] Со временем неоцентромера может накапливать повторяющиеся элементы и превращаться в то, что известно как эволюционная новая центромера. Есть несколько хорошо известных примеров в хромосомах приматов, где положение центромеры отличается от центромеры человека той же самой хромосомы и считается эволюционно новыми центромерами.[37] Было высказано предположение, что репозиционирование центромер и формирование новых эволюционных центромер является механизмом видообразование.[39]
Центромерные белки также являются аутоантигенной мишенью для некоторых антиядерные антитела, Такие как антицентромерные антитела.
Дисфункция и болезнь
Известно, что неправильная регуляция центромер способствует неправильной сегрегации хромосом, что сильно связано с раком и абортом. Примечательно, что сверхэкспрессия многих центромерных генов связана со злокачественными фенотипами рака. Сверхэкспрессия этих центромерных генов может увеличивать геномную нестабильность при раке. Повышенная геномная нестабильность, с одной стороны, связана со злокачественными фенотипами; с другой стороны, он делает опухолевые клетки более уязвимыми для специфических адъювантных методов лечения, таких как определенные виды химиотерапии и лучевая терапия.[40] Нестабильность повторяющейся ДНК центромеры была недавно показана при раке и старении.[41]
Этимология и произношение
Слово центромера (/ˈsɛптрəˌмɪər/[42][43]) использует комбинирование форм из центро- и -простой, давая «центральную часть», описывающую положение центромеры в центре хромосомы.
Смотрите также
Рекомендации
- ^ Альбертс, Брюс; Брей, Деннис; Хопкин, Карен; Джонсон, Александр; Льюис, Джулиан; Рафф, Мартин; Робертс, Кейт; Уолтер, Питер (2014). Эссенциальная клеточная биология (4-е изд.). Нью-Йорк, Нью-Йорк: Наука о гирляндах. п. 183. ISBN 978-0-8153-4454-4.
- ^ Поллард, Т. Д. (2007). Клеточная биология. Филадельфия: Сондерс. С. 200–203. ISBN 978-1-4160-2255-8.
- ^ Поллард, Т.Д. (2007). Клеточная биология. Филадельфия: Сондерс. С. 227–230. ISBN 978-1-4160-2255-8.
- ^ а б Pluta, A .; ЯВЛЯЮСЬ. Маккей; ЯВЛЯЮСЬ. Айнштейн; I.G. Гольдберг; ТУАЛЕТ. Эрншоу (1995). «Центромера: концентратор хромосомной активности». Наука. 270 (5242): 1591–1594. Bibcode:1995Научный ... 270.1591P. Дои:10.1126 / science.270.5242.1591. PMID 7502067.
- ^ «Сцепление сестринских хроматид». Домашний справочник по генетике. Национальная медицинская библиотека США. 15 мая 2011 г.
- ^ «p + q = решено, являясь правдивым рассказом о том, как хромосома получила свое имя». 2011-05-03.
- ^ Николай Уроки генетики (2013-10-12), Какие существуют разные типы хромосом?, получено 2017-05-28
- ^ а б Леван А., Фредга К., Сандберг А. А. (1964): Номенклатура центромерного положения на хромосомах. Эредитас, Лунд, 52: 201.
- ^ «Хромосомы, хромосомные аномалии».
- ^ *Гилберт Ф (1999). «Гены и хромосомы болезней: карты болезней генома человека. Хромосома 16». Genet Test. 3 (2): 243–54. Дои:10.1089 / gte.1999.3.243. PMID 10464676.
- ^ а б Нуссбаум, Роберт Л .; Макиннес, Родерик Р .; Томпсон, Маргарет Уилсон; Томпсон, Джеймс Скотт; Уиллард, Хантингтон Ф. (2001). Томпсон и Томпсон Генетика в медицине. ISBN 0721669026.
- ^ а б c Томпсон и Томпсон ГЕНЕТИКА В МЕДИЦИНЕ, 7-е издание. п. 62.
- ^ Myka, J.L .; Lear, T.L .; Houck, M.L .; Ryder, O.A .; Бейли, Э. (2003). "Анализ FISH, сравнивающий организацию генома у домашней лошади (Equus caballus) к монгольской дикой лошади (Э. Пржевальский)". Цитогенетические и геномные исследования. 102 (1–4): 222–5. Дои:10.1159/000075753. PMID 14970707.
- ^ а б Lin, C.Y .; Шукла, А .; Grady, J.P .; Fink, J.L .; Dray, E .; Duijf, P.H.G. (2018), «Точки разрыва транслокации преимущественно встречаются в эухроматине и акроцентрических хромосомах», Раки (Базель), 10 (1): E13, Дои:10.3390 / раки10010013, ЧВК 5789363, PMID 29316705
- ^ Bolzer, A .; и другие. (2005), «Трехмерные карты всех хромосом в ядрах мужских фибробластов человека и розетках прометафаз». PLOS Биология, 3 (5): e157, Дои:10.1371 / journal.pbio.0030157, ЧВК 1084335, PMID 15839726
- ^ Сильвер, Ли М. (1995). «Кариотипы, хромосомы и транслокации». Генетика мышей: концепции и приложения. Оксфорд: Издательство Оксфордского университета. С. 83–92. ISBN 978-0-19-507554-0.
- ^ Chinwalla, Asif T .; Повар, Лиза Л .; Delehaunty, Kimberly D .; Fewell, Ginger A .; Fulton, Lucinda A .; Фултон, Роберт С .; Могилы, Тина А .; Hillier, Ladeana W .; и другие. (2002). «Первоначальное секвенирование и сравнительный анализ генома мыши». Природа. 420 (6915): 520–62. Bibcode:2002 Натур. 420..520Вт. Дои:10.1038 / природа01262. PMID 12466850.
- ^ "определение субтелоцентрической хромосомы". groups.molbiosci.northwestern.edu. Получено 2017-10-29.
- ^ Маргулис, Линн; Мэтьюз, Клиффорд; Хазелтон, Аарон (01.01.2000). Эволюция окружающей среды: влияние происхождения и эволюции жизни на планете Земля. MIT Press. ISBN 9780262631976.
- ^ Певснер, Джонатан (2015-08-17). Биоинформатика и функциональная геномика. Джон Вили и сыновья. ISBN 9781118581766.
- ^ Нуссбаум, Роберт; Макиннес, Родерик; Уиллард, Хантингтон; Хамош, Ада (2007). Томпсон и Томпсон Генетика в медицине. Филадельфия (Пенсильвания): Сондерс. п. 72. ISBN 978-1-4160-3080-5.
- ^ Хартвелл, Лиланд; Худ, Лирой; Гольдберг, Майкл; Рейнольдс, Энн; Ли, Сильвер (2011). Генетика от генов к геномам, 4e. Нью-Йорк: Макгроу-Хилл. ISBN 9780073525266.
- ^ Линч, Салли; и другие. (1995). «Особенности синдрома Кабуки у монозиготных мальчиков-близнецов с псевдодицентрической хромосомой 13». J. Med. Genet. 32 (32:227–230): 227–230. Дои:10.1136 / jmg.32.3.227. ЧВК 1050324. PMID 7783176.
- ^ Barra, V .; Фачинетти, Д. (2018). «Темная сторона центромер: типы, причины и последствия структурных аномалий, затрагивающих центромерную ДНК». Nature Communications. 9 (1): 4340. Bibcode:2018НатКо ... 9,4340B. Дои:10.1038 / s41467-018-06545-у. ЧВК 6194107. PMID 30337534.
- ^ Нойман, Павел; Навратилова, Алиса; Шредер-Райтер, Элизабет; Коближкова, Андреа; Стейнбауэрова, Вероника; Хохолова, Ева; Новак, Петр; Ваннер, Герхард; Макас, Иржи (2012). «Растягивая правила: моноцентрические хромосомы с множеством центромерных доменов». PLOS Genetics. 8 (6): e1002777. Дои:10.1371 / journal.pgen.1002777. ЧВК 3380829. PMID 22737088.
- ^ Дернбург, А. Ф. (2001). «Здесь, там и везде: функция кинетохор на голоцентрических хромосомах». Журнал клеточной биологии. 153 (6): F33–8. Дои:10.1083 / jcb.153.6.F33. ЧВК 2192025. PMID 11402076.
- ^ Pluta, A. F .; MacKay, A.M .; Ainsztein, A. M .; Гольдберг, И. Г .; Эрншоу, В. К. (1995). «Центромера: центр хромосомной активности». Наука. 270 (5242): 1591–4. Bibcode:1995Научный ... 270.1591P. Дои:10.1126 / science.270.5242.1591. PMID 7502067.
- ^ а б Mehta, G.D .; Agarwal, M .; Гош, С. К. (2010). «Центромеры идентичности: вызов, который предстоит решить». Мол. Genet. Геномика. 284 (2): 75–94. Дои:10.1007 / s00438-010-0553-4. PMID 20585957.
- ^ Далал, Ямини (2009). «Эпигенетическая спецификация центромер». Биохимия и клеточная биология. 87 (1): 273–82. Дои:10.1139 / O08-135. PMID 19234541.
- ^ Бернад, Рафаэль; Санчес, Патрисия; Лосада, Ана (2009). «Эпигенетическая спецификация центромер по CENP-A». Экспериментальные исследования клеток. 315 (19): 3233–41. Дои:10.1016 / j.yexcr.2009.07.023. PMID 19660450.
- ^ Chueh, A.C .; Вонг, LH; Wong, N; Чу, К. Х. (2004). «Переменное и иерархическое распределение по размерам кластеров CENP-A, обогащенных L1-ретроэлементом, в функциональном неоцентромере человека». Молекулярная генетика человека. 14 (1): 85–93. Дои:10.1093 / hmg / ddi008. PMID 15537667.
- ^ Розенфельд, Джеффри А; Ван, Жибин; Шонес, Дастин Э; Чжао, Кэджи; Десалл, Роб; Чжан, Майкл Кью (2009). «Определение модификаций обогащенных гистонов в негенных частях генома человека». BMC Genomics. 10: 143. Дои:10.1186/1471-2164-10-143. ЧВК 2667539. PMID 19335899.
- ^ Чанг, C-H; Чаван, А; Палладино, Дж; Wei, X; Мартинс, NMC; Сантинелло, B; и другие. (2019). «Острова ретроэлементов являются основными компонентами центромер дрозофилы». ПЛОС Биол. 17 (5): e3000241. Дои:10.1371 / journal.pbio.3000241. ЧВК 6516634. PMID 31086362.
- ^ Volpe, T. A .; Киднер, К; Холл, ИМ; Teng, G; Grewal, SI; Мартиенссен, РА (2002). "Регулирование гетерохроматического молчания и метилирования гистона H3 лизина-9 с помощью РНКи". Наука. 297 (5588): 1833–7. Bibcode:2002Научный ... 297.1833V. Дои:10.1126 / science.1074973. PMID 12193640. S2CID 2613813.
- ^ Маршалл, Оуэн Дж .; Chueh, Anderly C .; Вонг, Ли Х .; Чу, К. Энди (2008). «Неоцентромеры: новый взгляд на структуру центромер, развитие заболеваний и эволюцию кариотипа». Американский журнал генетики человека. 82 (2): 261–82. Дои:10.1016 / j.ajhg.2007.11.009. ЧВК 2427194. PMID 18252209.
- ^ Уорбертон, Питер Э. (2004). «Хромосомная динамика образования неоцентромеров человека». Хромосомные исследования. 12 (6): 617–26. Дои:10.1023 / B: CHRO.0000036585.44138.4b. PMID 15289667.
- ^ а б Рокки, М; Archidiacono, N; Шемпп, Вт; Капоцци, О; Станьон, Р. (январь 2012 г.). «Репозиционирование центромеры у млекопитающих». Наследственность. 108 (1): 59–67. Дои:10.1038 / hdy.2011.101. ISSN 0018-067X. ЧВК 3238114. PMID 22045381.
- ^ Толомео, Дорон; Капоцци, Оронцо; Станьон, Роско Р .; Арчидиаконо, Николетта; Д’Аддаббо, Пьетро; Catacchio, Claudia R .; Пургато, Стефания; Перини, Джованни; Шемпп, Вернер; Хаддлстон, Джон; Малиг, Майка (03.02.2017). «Эпигенетическое происхождение эволюционных новых центромер». Научные отчеты. 7 (1): 41980. Дои:10.1038 / srep41980. ISSN 2045-2322. ЧВК 5290474. PMID 28155877.
- ^ Браун, Джудит Д .; О'Нил, Рэйчел Дж. (Сентябрь 2010 г.). "Хромосомы, конфликт и эпигенетика: новый взгляд на хромосомную спецификацию". Ежегодный обзор геномики и генетики человека. 11 (1): 291–316. Дои:10.1146 / annurev-genom-082509-141554. ISSN 1527-8204.
- ^ Zhang, W .; Mao, JH .; Zhu, W .; Jain, A.K .; Liu, L .; Brown, J.B .; Карпен, Г. (2016). «Неправильная экспрессия генов центромеры и кинетохор предсказывает выживаемость больных раком и реакцию на лучевую терапию и химиотерапию». Nature Communications. 7: 12619. Bibcode:2016 НатКо ... 712619Z. Дои:10.1038 / ncomms12619. ЧВК 5013662. PMID 27577169.
- ^ Джунта, S; Фунабики, H (21 февраля 2017 г.). «Целостность повторов ДНК центромеры человека защищена CENP-A, CENP-C и CENP-T». Труды Национальной академии наук Соединенных Штатов Америки. 114 (8): 1928–1933. Дои:10.1073 / pnas.1615133114. ЧВК 5338446. PMID 28167779.
- ^ «Центромера». Словарь Merriam-Webster.
- ^ «Центромера». Dictionary.com Несокращенный. Случайный дом.
дальнейшее чтение
- Mehta, G.D .; Agarwal, M .; Гош, С. К. (2010). «Центромеры идентичности: вызов, который предстоит решить». Мол. Genet. Геномика. 284 (2): 75–94. Дои:10.1007 / s00438-010-0553-4. PMID 20585957.
- Лодиш, Харви; Берк, Арнольд; Kaiser, Chris A .; Кригер, Монти; Скотт, Мэтью П .; Бретчер, Энтони; Плоег, Хиддл; Мацудаира, Пол (2008). Молекулярная клеточная биология (6-е изд.). Нью-Йорк: W.H. Фримен. ISBN 978-0-7167-7601-7.
- Нагаки, Киётака; Ченг, Чжукуань; Оуян, Шу; Талберт, Пол Б; Ким, Мэри; Джонс, Кристин М; Хеникофф, Стивен; Буэлл, С. Робин; Цзян, Цзимин (2004). «Секвенирование центромеры риса раскрывает активные гены». Природа Генетика. 36 (2): 138–45. Дои:10.1038 / ng1289. PMID 14716315. Сложить резюме – Science Daily (13 января 2004 г.).