Schizosaccharomyces pombe - Schizosaccharomyces pombe
| Schizosaccharomyces pombe | |
|---|---|
 | |
| Научная классификация | |
| Королевство: | |
| Разделение: | |
| Учебный класс: | |
| Заказ: | |
| Семья: | |
| Род: | |
| Разновидность: | С. Помбе |
| Биномиальное имя | |
| Schizosaccharomyces pombe Линднер (1893) | |
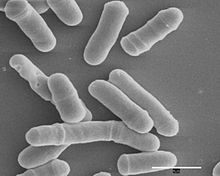
Schizosaccharomyces pombe, также называемый "делящиеся дрожжи", это разновидность из дрожжи используется в традиционном пивоварении и как модельный организм в молекулярный и клеточная биология. Это одноклеточный эукариот, чей клетки имеют стержневидную форму. Ячейки обычно имеют размер от 3 до 4 микрометры в диаметре и от 7 до 14 мкм в длину. Его геном, который составляет приблизительно 14,1 миллиона пар оснований, по оценкам содержит 4970 кодирующих белок гены и не менее 450 некодирующие РНК.[1]
Эти клетки сохраняют свою форму за счет роста исключительно через кончики клеток и деления медиальной части. деление производить две дочерние клетки одинакового размера, что делает их мощным инструментом в клеточный цикл исследование.
Делящиеся дрожжи были выделены в 1893 г. Пол Линднер из Восточной Африки пшенное пиво. Название вида помба это суахили слово для пива. Впервые он был разработан как экспериментальная модель в 1950-х годах: Урс Леупольд для изучения генетики,[2][3] и по Мердок Митчисон для изучения клеточного цикла.[4][5][6]
Пол Медсестра, исследователь делящихся дрожжей, успешно объединил независимые школы генетики делящихся дрожжей и исследований клеточного цикла. Вместе с Ли Хартвелл и Тим Хант, Медсестра выиграла 2001 Нобелевская премия по физиологии и медицине за их работу по регуляции клеточного цикла.
Последовательность С. Помбе геном был опубликован в 2002 году консорциумом во главе с Институт Сэнгера, став шестой моделью эукариотического организма, чей геном был полностью последовательный. Исследователи S. pombe поддерживаются PomBase МОД (База данных модельных организмов ). Это полностью раскрыло возможности этого организма, и было идентифицировано множество генов, ортологичных человеческим генам - на сегодняшний день 70%,[7][8] включая многие гены, участвующие в заболеваниях человека.[9] В 2006 г. субклеточная локализация почти всех белков в С. Помбе был опубликован с использованием зеленый флуоресцентный белок как молекулярный тег.[10]
Schizosaccharomyces pombe также стал важным организмом в изучении клеточных реакций на Повреждение ДНК и процесс Репликация ДНК.
Около 160 природных штаммов С. Помбе были изолированы. Они были собраны из разных мест, включая Европу, Северную и Южную Америку и Азию. Большинство этих штаммов было собрано с культурных фруктов, таких как яблоки и виноград, или из различных Алкогольные напитки, например бразильский Cachaça. С. Помбе также известно, что он присутствует в ферментированном чае, чайный гриб.[11] В настоящее время неясно, С. Помбе является основным ферментером или загрязнителем такого пива. Естественная экология Шизосахаромицеты дрожжи мало изучены.
История
Schizosaccharomyces pombe был впервые обнаружен в 1893 году, когда группа, работающая в лаборатории ассоциации пивоваренных заводов в Германии, изучала осадок, обнаруженный в пиве из проса, импортированном из Восточной Африки, который придавал ему кислый вкус. Термин «шизо», означающий «расщепление» или «расщепление», ранее использовался для описания других Шизосахаромицеты. Добавление слова pombe было связано с его изоляцией от восточноафриканского пива, поскольку pombe означает «пиво» на суахили. Стандарт С. Помбе Штаммы были выделены Урсом Леупольдом в 1946 и 1947 годах из культуры, полученной им из коллекции дрожжей в г. Делфт, Нидерланды. Он был сдан на хранение А. Остервальдеру под именем С. Помбе var. liquefaciens, после того как он изолировал его в 1924 году от французского вина (скорее всего, прогорклого) на Федеральной экспериментальной станции виноградарства и садоводства в г. Wädenswil, Швейцария. Культура, использованная Урсом Лейпольдом, содержала (помимо других) клетки с типами спаривания h90 (штамм 968), h- (штамм 972) и h + (штамм 975). После этого были предприняты две большие попытки изолировать С. Помбе из фруктов, нектара или ферментации: одно от Florenzano et al.[12] на виноградниках западной Сицилии, а другой - Gomes et al. (2002) в четырех регионах юго-востока Бразилии.[13]
Экология
Делящиеся дрожжи С. Помбе принадлежит к divisio Ascomycota, который представляет собой самую большую и разнообразную группу грибов. Свободноживущие аскомицеты обычно встречаются в экссудатах деревьев, на корнях растений и в окружающей почве, на спелых и гниющих плодах, а также в ассоциации с насекомыми-переносчиками, переносящими их между субстратами. Многие из этих ассоциаций являются симбиотическими или сапрофитными, хотя многочисленные аскомицеты (и их родственники-базидиомицеты) представляют собой важные патогены растений, поражающие мириады видов растений, включая товарные культуры. Среди родов аскомицетных дрожжей делящиеся дрожжи Шизосахаромицеты является уникальным из-за отложения α- (1,3) -глюкана или псевдонигерана в клеточной стенке в дополнение к более известным β-глюканам и фактическому отсутствию хитина. Виды этого рода также различаются по составу маннана, который показывает концевые сахара d-галактозы в боковых цепях их маннанов. С. Помбе пройти аэробная ферментация при наличии лишнего сахара.[14] С. Помбе могут разлагать L-яблочную кислоту, одну из доминирующих органических кислот в вине, что делает их разнообразными среди других Сахаромицеты штаммы.
Сравнение с бутонизированными дрожжами (Saccharomyces cerevisiae)
Виды дрожжей Schizosaccharomyces pombe и Saccharomyces cerevisiae оба широко изучены; эти два вида разошлись примерно за 300-600 миллионов лет до настоящего времени,[15] и являются важными инструментами в молекулярной и клеточной биологии. Вот некоторые из технических различий между этими двумя видами:
- С. cerevisiae насчитывает около 5600 открытые рамки для чтения; С. Помбе имеет примерно 5070 открытых рамок чтения.
- Несмотря на одинаковое количество генов, С. cerevisiae всего около 250 интроны, пока С. Помбе насчитывает почти 5000 человек.
- С. cerevisiae имеет 16 хромосомы, С. Помбе имеет 3.
- С. cerevisiae часто диплоид пока С. Помбе обычно гаплоидный.
- С. Помбе имеет укрытие -подобно теломер сложный пока С. cerevisiae не.[16]
- С. cerevisiae находится в фазе G1 клеточный цикл в течение длительного периода (как следствие, переход G1-S жестко контролируется), а С. Помбе остается в фазе G2 клеточного цикла в течение длительного периода (как следствие, переход G2-M находится под жестким контролем).
- Оба вида имеют общие гены с высшими эукариотами, которые они не разделяют друг с другом. С. Помбе имеет RNAi машины гены, подобные таковым у позвоночных, в то время как это отсутствует в С. cerevisiae. С. cerevisiae также значительно упростил гетерохроматин по сравнению с С. Помбе.[17] Наоборот, С. cerevisiae хорошо развит пероксисомы, пока С. Помбе не.
- С. cerevisiae имеет небольшую точку центромера 125 п.н., и точки начала репликации с определенными последовательностями примерно того же размера. Напротив, С. Помбе имеет большие повторяющиеся центромеры (40–100 kb), более похожие на центромеры млекопитающих, и вырожденные точки начала репликации размером по крайней мере 1 kb.
Пути S. pombe и клеточные процессы
С. Помбе генные продукты (белки и РНК) участвуют во многих клеточных процессах, общих для всей жизни. В делящиеся дрожжи GO slim предоставляет категоричный обзор высокого уровня биологической роли всех генных продуктов S. pombe.[7]
Жизненный цикл

Делящиеся дрожжи - это одноклеточный гриб с простым, полностью охарактеризованным геномом и быстрой скоростью роста. Он издавна используется в пивоварении, выпечке и молекулярной генетике. С. Помбе представляет собой палочковидную клетку диаметром около 3 мкм, которая полностью растет за счет удлинения на концах. После митоза деление происходит за счет образования перегородки или клеточной пластинки, которая расщепляет клетку в ее средней точке.
Центральными событиями размножения клеток являются удвоение хромосом, которое происходит в S (синтетической) фазе, за которой следует сегрегация хромосом и ядерное деление (митоз) и деление клеток (цитокинез), которые в совокупности называются M (митотической) фазой. G1 - это промежуток между фазами M и S, а G2 - промежуток между фазами S и M. У делящихся дрожжей фаза G2 особенно увеличена, и цитокинез (сегрегация дочерних клеток) не происходит до тех пор, пока не будет запущена новая S (синтетическая) фаза.
Делящиеся дрожжи управляют митозом с помощью механизмов, аналогичных таковым у многоклеточных животных. Обычно он размножается в гаплоидном состоянии. При голодании клетки противоположных типов спаривания (P и M) сливаются, образуя диплоидную зиготу, которая сразу же входит в мейоз и генерирует четыре гаплоидные споры. Когда условия улучшаются, эти споры прорастают с образованием пролиферирующих гаплоидных клеток.[18]

Общие особенности клеточного цикла.

Конкретный клеточный цикл делящихся дрожжей.

Этапы разделения Шизосахаромицеты в световой и темнопольной микроскопии


Цитокинез

Здесь показаны общие особенности цитокинеза. Перед анафазой определяется место деления клеток. Веретено анафазы (зеленое на рисунке) затем позиционируется так, чтобы сегрегированные хромосомы находились на противоположных сторонах заданной плоскости расщепления.
Контроль размера

У делящихся дрожжей, рост которых управляет прохождением через G2 / M, мутация wee1 вызывает вступление в митоз аномально маленького размера, что приводит к более короткому G2. G1 удлиняется, что свидетельствует о том, что прогрессирование через Start (начало клеточного цикла) реагирует на рост, когда контроль G2 / M теряется. Более того, клетки в плохих питательных условиях растут медленно, поэтому требуется больше времени, чтобы удвоиться в размере и разделиться. Низкие уровни питательных веществ также сбрасывают порог роста, так что клетка проходит через клеточный цикл с меньшим размером. При воздействии стрессовых условий [тепло (40 ° C) или окислитель перекись водорода] С. Помбе клетки подвергаются старение что измеряется увеличением времени деления клеток и повышенной вероятностью гибели клеток.[19] Наконец, мутантные делящиеся дрожжевые клетки wee1 меньше, чем клетки дикого типа, но им требуется столько же времени, чтобы пройти клеточный цикл. Это возможно, потому что маленькие дрожжевые клетки растут медленнее, то есть их общая добавленная масса в единицу времени меньше, чем у нормальных клеток.
Считается, что пространственный градиент координирует размер клеток и митотический вход у делящихся дрожжей.[20][21][22]В Пом1 протеинкиназа (зеленая) локализована в коре клетки с наибольшей концентрацией на концах клеток. Регуляторы клеточного цикла Cdr2, Cdr1 и Wee1 присутствуют в корковых узлах в середине клетки (синие и красные точки). а, В маленьких ячейках Пом1 градиент достигает большинства корковых узлов (синие точки). Пом1 ингибирует Cdr2, предотвращая ингибирование Cdr2 и Cdr1 Wee1 и позволяя Wee1 фосфорилировать Cdk1, таким образом инактивируя активность циклин-зависимой киназы (CDK) и предотвращая вступление в митоз. б, В длинных ячейках Пом1 градиент не достигает корковых узлов (красные точки), и поэтому Cdr2 и Cdr1 остаются активными в узлах. Cdr2 и Cdr1 ингибируют Wee1, предотвращая фосфорилирование Cdk1 и тем самым приводя к активации CDK и митотическому входу. (Эта упрощенная диаграмма не включает несколько других регуляторов деятельности CDK.)
Переключение типа сопряжения
Делящиеся дрожжи переключают тип спаривания с помощью события рекомбинации, связанной с репликацией, которое происходит во время S-фазы клеточного цикла. Делящиеся дрожжи используют внутреннюю асимметрию процесса репликации ДНК для переключения типа спаривания; это была первая система, в которой было показано, что направление репликации необходимо для изменения типа клеток. Исследования системы переключения типа спаривания привели к открытию и характеристике сайт-специфичного сайта терминации репликации RTS1, сайт-специфичного сайта паузы репликации MPS1 и нового типа хромосомного отпечатка, маркирующего одну из сестринских хроматид при спаривании. -тип locus mat1. Кроме того, работа с молчащей донорской областью привела к большим успехам в понимании образования и поддержания гетерохроматина.[23]
Ответы на повреждение ДНК
Schizosaccharomyces pombe - это факультативный половой микроорганизм, который может спариваться при ограничении питательных веществ.[24] Контакт с С. Помбе в перекись водорода, агент, который вызывает окислительный стресс приводящий к окислительному Повреждение ДНК, сильно индуцирует вязка и образование мейотических спор.[25] Это открытие предполагает, что мейоз, и особенно мейотическая рекомбинация, может быть адаптацией для восстановления повреждений ДНК.[нужна цитата ] Эта точка зрения подтверждается открытием того факта, что одноосновные повреждения типа dU: dG в ДНК С. Помбе стимулируют мейотическую рекомбинацию.[26] Эта рекомбинация требует урацил-ДНК гликозилаза, фермент, который удаляет урацил из основной цепи ДНК и инициирует эксцизионную репарацию оснований. На основании этого открытия было предложено, что эксцизионная репарация основания урацила, базисного сайта или однонитевого разрыва является достаточной для инициирования рекомбинация в S. pombe.[26] Другие эксперименты с С. Помбе указали, что неправильная обработка промежуточных продуктов репликации ДНК, т.е. Фрагменты Окадзаки, вызывает повреждения ДНК, такие как одноцепочечные разрывы или разрывы, и это стимулирует мейотическую рекомбинацию.[27]
Как модельная система
Делящиеся дрожжи стали заметной модельной системой для изучения основных принципов работы клетки, которую можно использовать для понимания более сложных организмов, таких как млекопитающие и, в частности, люди.[28][29] Этот одноклеточный эукариот непатогенен, его легко выращивать и манипулировать им в лаборатории.[30][31] Делящиеся дрожжи содержат одно из наименьших количеств генов из известной последовательности генома эукариота и всего три хромосомы в его геноме.[32] Многие из генов, ответственных за деление клеток и клеточную организацию в клетках делящихся дрожжей, также обнаружены в геноме человека.[30][31][33] Регуляция и деление клеточного цикла имеют решающее значение для роста и развития любой клетки. Консервативные гены делящихся дрожжей были тщательно изучены и стали причиной многих недавних биомедицинских разработок.[34][35] Делящиеся дрожжи также представляют собой практическую модельную систему для наблюдения за делением клеток, поскольку делящиеся дрожжи представляют собой одноклеточные эукариоты цилиндрической формы, которые делятся и воспроизводятся посредством медиального деления.[30] Это легко увидеть с помощью микроскопии. Делящиеся дрожжи также имеют чрезвычайно короткое время генерации, от 2 до 4 часов, что также делает их простой модельной системой для наблюдения и выращивания в лаборатории.[31] Простота геномной структуры делящихся дрожжей и их сходство с геномом млекопитающих, простота манипуляций и возможность использования для анализа лекарств - вот почему делящиеся дрожжи вносят большой вклад в исследования биомедицины и клеточной биологии, а также в модельную систему для генетического анализа.[31][24][29][36][37]
Геном
Schizosaccharomyces pombe часто используется для изучения деления и роста клеток из-за консервативных областей генома, которые также наблюдаются у человека, включая: белки гетерохроматина, крупные источники репликации, большие центромеры, консервативные клеточные контрольные точки, функцию теломер, сплайсинг генов и многие другие клеточные процессы.[32][38][39] С. Помбе'Геном s был полностью секвенирован в 2002 году, это шестой геном эукариот, который будет секвенирован в рамках проекта «Геном». По оценкам, 4979 генов были обнаружены в трех хромосомах, содержащих около 14 МБ ДНК. Эта ДНК содержится в 3 разных хромосомах в ядре с промежутками в центромерном (40kb) и теломерном (260kb) областях.[32] После первоначального секвенирования генома делящихся дрожжей были секвенированы другие ранее не секвенированные области генов. Структурный и функциональный анализ этих участков генов можно найти в крупномасштабных базах данных по делящимся дрожжам, таким как PomBase.[40]
Было обнаружено, что 43 процента генов в проекте «Геном» содержат интроны в 4739 генах. Делящиеся дрожжи не имеют такого количества дублированных генов, как почкующиеся дрожжи, они содержат только 5%, что делает делящиеся дрожжи отличной моделью генома для наблюдения и дает исследователям возможность создавать более функциональные исследовательские подходы. С. Помбе'Наличие большого количества интронов дает возможности для увеличения диапазона типов белков, полученных в результате альтернативного сплайсинга, и генов, которые кодируют сопоставимые гены у человека.[32]81% из трех центромер у делящихся дрожжей секвенированы. Длина трех центромер составила 34, 65 и 110 т.п.н. Это в 300–100 раз длиннее центромеры почкующихся дрожжей. Чрезвычайно высокий уровень консервативности (97%) также наблюдается в области 1780 п.н. в областях DGS центромеры. Это удлинение центромер и ее консервативные последовательности делают делящиеся дрожжи практической модельной системой, которую можно использовать для наблюдения за делением клеток и у людей из-за их сходства.[32][41][42]
PomBase[7][43] сообщает, что более 69% генов, кодирующих белок, имеют человеческие ортологи и более 500 из них связано с болезнью человека . Это делает С. Помбе отличная система для изучения человеческих генов и путей распространения болезней, особенно клеточного цикла и систем контрольных точек ДНК.[42][44][45][46]
Генетическое разнообразие
Изучение биоразнообразия и эволюции делящихся дрожжей проводилось на 161 штамме Schizosaccharomyces pombe, собранном из 20 стран.[47] Моделирование скорости эволюции показало, что все штаммы произошли от общего предка, жившего примерно 2300 лет назад. Исследование также выявило набор из 57 штаммов делящихся дрожжей, каждый из которых отличается ≥1900 SNP;[47] и все обнаруженные 57 штаммов делящихся дрожжей были прототрофными (способны расти на той же минимальной среде, что и контрольный штамм).[47] Ряд исследований генома S. pombe поддерживает идею о том, что генетическое разнообразие штаммов делящихся дрожжей немного меньше, чем у почкующихся дрожжей.[47] Действительно, только ограниченные вариации S. pombe встречаются при пролиферации в различных средах. Кроме того, количество сегрегированных фенотипических вариаций у делящихся дрожжей меньше, чем у S. cerevisiae.[48] Поскольку большинство штаммов делящихся дрожжей были выделены из сваренных напитков, это распространение не имеет экологического или исторического контекста.
Анализ клеточного цикла
Репликация ДНК у дрожжей все больше изучается многими исследователями. Дальнейшее понимание репликации ДНК, экспрессии генов и консервативных механизмов у дрожжей может предоставить исследователям информацию о том, как эти системы действуют в клетках млекопитающих в целом и в клетках человека в частности.[39][49][50][51] Другие стадии, такие как клеточный рост и старение, также наблюдаются у дрожжей, чтобы понять эти механизмы в более сложных системах.[33][52][53][54]
С. Помбе ячейки стационарной фазы подвергаются хронологическому старение из-за производства активные формы кислорода это причина Повреждения ДНК. Большинство таких повреждений обычно можно исправить с помощью ДНК. базовая эксцизионная пластика и эксцизионная репарация нуклеотидов.[55] Дефекты этих процессов восстановления приводят к снижению выживаемости.
Цитокинез - один из компонентов деления клеток, который часто наблюдается у делящихся дрожжей. Хорошо законсервированные компоненты цитокинеза наблюдаются у делящихся дрожжей и позволяют нам рассматривать различные геномные сценарии и точно определять мутации.[45][56][57] Цитокинез - это постоянный шаг, который очень важен для благополучия клетки.[58] В частности, образование сократительного кольца тщательно изучается исследователями, использующими С. Помбе как модельная система. Сократительное кольцо высоко консервативно как в цитокинезе делящихся дрожжей, так и у человека.[45] Мутации в цитокинезе могут привести ко многим сбоям в работе клетки, включая гибель клеток и развитие раковых клеток.[45] Это сложный процесс деления клеток человека, но в С. Помбе более простые эксперименты могут дать результаты, которые затем могут быть применены для исследований в модельных системах более высокого порядка, таких как люди.
Одна из мер предосторожности, которую принимает клетка для обеспечения точного деления клетки, - это контрольная точка клеточного цикла.[59][60] Эти контрольные точки обеспечивают устранение любых мутагенов.[61] Это часто осуществляется с помощью ретрансляционных сигналов, которые стимулируют убиквитинирование мишеней и задерживают цитокинез.[32] Без таких митотических контрольных точек мутагены создаются и реплицируются, что приводит к множеству клеточных проблем, включая гибель клеток или туморогенез, наблюдаемые в раковых клетках. Пол Нерс, Лиланд Хартвелл и Тим Хант были удостоены Нобелевской премии по физиологии и медицине в 2001 году. Они обнаружили ключевые консервативные контрольные точки, которые имеют решающее значение для правильного деления клетки. Эти результаты были связаны с раком и больными клетками и являются важным открытием для биомедицины.[62]
Исследователи, использующие делящиеся дрожжи в качестве модельной системы, также изучают динамику и ответы органелл, а также возможные корреляции между дрожжевыми клетками и клетками млекопитающих.[63][64] Заболевания митохондрий и различные системы органелл, такие как аппарат Гольджи и эндоплазматический ретикулум, можно дополнительно понять, наблюдая за динамикой хромосом делящихся дрожжей, уровнями экспрессии и регуляции белков.[46][50][65][66][67][68]
Биомедицинский инструмент
Однако есть ограничения при использовании делящихся дрожжей в качестве модельной системы: их множественная лекарственная устойчивость. «МЛУ-ответ включает сверхэкспрессию двух типов насосов оттока лекарств, семейства АТФ-связывающих кассет (ABC) ... и суперсемейства основных фасилитаторов».[34] Пол Нерс и некоторые его коллеги недавно создали С. Помбе штаммы, чувствительные к химическим ингибиторам, и обычные зонды, чтобы увидеть, можно ли использовать делящиеся дрожжи в качестве модельной системы химических исследований лекарственных препаратов.[34]
Например, доксорубицин, очень распространенный химиотерапевтический антибиотик, имеет множество побочных эффектов. Исследователи ищут способы лучше понять, как работает доксорубицин, наблюдая за генами, связанными с устойчивостью, используя делящиеся дрожжи в качестве модельной системы. Была обнаружена связь между побочными эффектами доксорубицина и хромосомным метаболизмом и мембранным транспортом. Метаболические модели для нацеливания лекарств сейчас используются в биотехнологии, и в будущем ожидается дальнейший прогресс с использованием системы моделей делящихся дрожжей.[35]
Экспериментальные подходы
Делящиеся дрожжи легко доступны, их легко выращивать и манипулировать ими для получения мутантов, а также их можно поддерживать в гаплоидном или диплоидном состоянии. С. Помбе Обычно это гаплоидная клетка, но в стрессовых условиях, обычно при дефиците азота, две клетки будут конъюгировать с образованием диплоида, который позже образует четыре споры в тетрадной аске.[31] Этот процесс легко увидеть и наблюдать под любым микроскопом и позволяет нам взглянуть на мейоз в более простой модельной системе, чтобы увидеть, как работает это явление.
Таким образом, практически любой генетический эксперимент или метод может быть применен к этой модельной системе, например: тетрадное рассечение, анализ мутагенов, трансформации и методы микроскопии, такие как FRAP и FRET. Новые модели, такие как Tug-Of-War (gTOW), также используются для анализа устойчивости дрожжей и наблюдения за экспрессией генов. Создание генов «нокаута» и «нокаута» довольно просто, а при секвенировании генома делящихся дрожжей эта задача очень доступна и хорошо известна.[69][70]
Смотрите также
Рекомендации
- ^ Вильгельм Б.Т., Маргерат С., Ватт С., Шуберт Ф., Вуд В., Гудхед И. и др. (Июнь 2008 г.). «Динамический репертуар эукариотического транскриптома, исследуемый при разрешении одного нуклеотида». Природа. 453 (7199): 1239–43. Bibcode:2008 Натур.453.1239W. Дои:10.1038 / природа07002. PMID 18488015. S2CID 205213499.
- ^ Леупольд У (1950). "Die Vererbung von Homothallie und Heterothallie bei" Schizosaccharomyces pombe". CR Trav Lab Carlsberg Ser Physiol. 24: 381–480.
- ^ Леупольд У. (1993) Истоки Schizosaccharomyces pombe генетика. В: Hall MN, Linder P. eds. Первые годы генетики дрожжей. Нью-Йорк. Лабораторный пресс Колд-Спринг-Харбор. С. 125–128.
- ^ Митчисон Дж. М. (октябрь 1957 г.). «Рост одиночных клеток. I. Schizosaccharomyces pombe». Экспериментальные исследования клеток. 13 (2): 244–62. Дои:10.1016/0014-4827(57)90005-8. PMID 13480293.
- ^ Митчисон Дж. М. (апрель 1990 г.). «Делящиеся дрожжи Schizosaccharomyces pombe». BioEssays. 12 (4): 189–91. Дои:10.1002 / bies.950120409. PMID 2185750.
- ^ Fantes PA, Hoffman CS (июнь 2016 г.). "Краткая история исследования Schizosaccharomyces pombe: взгляд за последние 70 лет". Генетика. 203 (2): 621–9. Дои:10.1534 / genetics.116.189407. ЧВК 4896181. PMID 27270696.
- ^ а б c Wood V, Harris MA, McDowall MD, Rutherford K, Vaughan BW, Staines DM, et al. (Январь 2012 г.). «PomBase: обширный онлайн-ресурс по делящимся дрожжам». Исследования нуклеиновых кислот. 40 (Проблема с базой данных): D695-9. Дои:10.1093 / nar / gkr853. ЧВК 3245111. PMID 22039153.
- ^ «ПомБаза».
- ^ «ПомБаза».
- ^ Мацуяма А., Араи Р., Яширода Й., Шираи А., Камата А., Секидо С. и др. (Июль 2006 г.). «Клонирование ORFeome и глобальный анализ локализации белка в делящихся дрожжах Schizosaccharomyces pombe». Природа Биотехнологии. 24 (7): 841–7. Дои:10.1038 / nbt1222. PMID 16823372. S2CID 10397608.
- ^ Теох А.Л., Херд Дж., Кокс Дж. (Сентябрь 2004 г.). «Экология дрожжей ферментации чайного гриба». Международный журнал пищевой микробиологии. 95 (2): 119–26. Дои:10.1016 / j.ijfoodmicro.2003.12.020. PMID 15282124.
- ^ Флоренцано Г., Баллони В., Матерасси Р. (1977). "Contributo alla ecologia dei lieviti Schizosaccharomyces sulle uve". Vitis. 16: 38–44.
- ^ Гомес Е.Б., Байлис Дж.М., Форсбург С.Л. (2002). «Делящиеся дрожжи вступают в новую радостную эру». Геномная биология. 3 (6): ОТЧЕТЫ4017. Дои:10.1186 / gb-2002-3-6-reports4017. ЧВК 139370. PMID 12093372.
- ^ Lin Z, Li WH (апрель 2011 г.). «Эволюция аэробной ферментации у Schizosaccharomyces pombe была связана с регуляторным репрограммированием, но не реорганизацией нуклеосом». Молекулярная биология и эволюция. 28 (4): 1407–13. Дои:10.1093 / molbev / msq324. ЧВК 3058771. PMID 21127171.
- ^ Douzery EJ, Snell EA, Bapteste E, Delsuc F, Philippe H (октябрь 2004 г.). «Время эволюции эукариот: согласовывают ли расслабленные молекулярные часы белки и окаменелости?». Труды Национальной академии наук Соединенных Штатов Америки. 101 (43): 15386–91. Bibcode:2004PNAS..10115386D. Дои:10.1073 / pnas.0403984101. ЧВК 524432. PMID 15494441.
- ^ Прайс К.М., Больц К.А., Чайкен М.Ф., Стюарт Дж. А., Бейльштейн М.А., Шиппен Д.Е. (август 2010 г.). «Эволюция функции CST в поддержании теломер». Клеточный цикл. 9 (16): 3157–65. Дои:10.4161 / cc.9.16.12547. ЧВК 3041159. PMID 20697207.
- ^ Грюнштейн, Майкл и Сьюзан Гассер. «Эпигенетика в Saccharomyces cerevisiae». Эпигенетика. 1. Cold Spring Harbor Press, 2007.
- ^ Морган, Дэвид О. (2007). Принципы контроля клеточного цикла. Лондон: New Science Press. ISBN 978-0-19-920610-0. OCLC 70173205.
- ^ Коэльо М., Дерели А., Хезе А., Кюн С., Малиновска Л., ДеСантис М.Э. и др. (Октябрь 2013). «Делящиеся дрожжи не стареют при благоприятных условиях, но стареют после стресса». Текущая биология. 23 (19): 1844–52. Дои:10.1016 / j.cub.2013.07.084. ЧВК 4620659. PMID 24035542.
- ^ Мозли, Джеймс Б.; Майе, Аделина; Паолетти, Энн; Медсестра, Пол (2009). «Пространственный градиент координирует размер клетки и митотический вход у делящихся дрожжей». Природа. 459 (7248): 857–860. Bibcode:2009Натура.459..857М. Дои:10.1038 / природа08074. ISSN 1476-4687. PMID 19474789. S2CID 4330336.
- ^ Мартин, Софи Дж .; Бертело-Грожан, Мартина (11.06.2009). «Полярные градиенты киназы Pom1 семейства DYRK связывают длину клетки с клеточным циклом». Природа. 459 (7248): 852–856. Bibcode:2009Натура.459..852М. Дои:10.1038 / природа08054. ISSN 1476-4687. PMID 19474792. S2CID 4412402.
- ^ Савин К.Е. (июнь 2009 г.). «Клеточный цикл: деление клеток уменьшено до размеров». Природа. 459 (7248): 782–3. Bibcode:2009Натура.459..782S. Дои:10.1038 / 459782a. PMID 19516326. S2CID 4402226.
- ^ Клар, Амар Дж. (2007-12-01).«Уроки, извлеченные из исследований переключения типа спаривания и молчания делящихся дрожжей». Ежегодный обзор генетики. 41 (1): 213–236. Дои:10.1146 / annurev.genet.39.073103.094316. ISSN 0066-4197. PMID 17614787.
- ^ а б Дэйви Дж. (Декабрь 1998 г.). «Сплав делящихся дрожжей». Дрожжи. 14 (16): 1529–66. Дои:10.1002 / (SICI) 1097-0061 (199812) 14:16 <1529 :: AID-YEA357> 3.0.CO; 2-0. PMID 9885154.
- ^ Бернштейн С., Джонс В. (апрель 1989 г.). «Половое размножение как ответ на повреждение H2O2 у Schizosaccharomyces pombe». Журнал бактериологии. 171 (4): 1893–7. Дои:10.1128 / jb.171.4.1893-1897.1989. ЧВК 209837. PMID 2703462.
- ^ а б Пауклин С., Беркерт Дж. С., Мартин Дж., Осман Ф., Веллер С., Боултон С. Дж. И др. (Май 2009 г.). «Альтернативная индукция мейотической рекомбинации из одноосновных повреждений ДНК дезаминаз». Генетика. 182 (1): 41–54. Дои:10.1534 / genetics.109.101683. ЧВК 2674839. PMID 19237686.
- ^ Фарах Дж. А., Кроми Дж., Дэвис Л., Штайнер В. В., Смит Г. Р. (декабрь 2005 г.). «Активация альтернативного, rec12 (spo11) -независимого пути мейотической рекомбинации делящихся дрожжей в отсутствие эндонуклеазы лоскута ДНК». Генетика. 171 (4): 1499–511. Дои:10.1534 / генетика.105.046821. ЧВК 1456079. PMID 16118186.
- ^ Форсбург SL (июнь 2005 г.). «Дрожжи Saccharomyces cerevisiae и Schizosaccharomyces pombe: модели для исследования клеточной биологии». Бюллетень гравитационной и космической биологии. 18 (2): 3–9. PMID 16038088.
- ^ а б Форсбург С.Л., Райн Н. (февраль 2006 г.). «Основные методы получения делящихся дрожжей». Дрожжи. 23 (3): 173–83. Дои:10.1002 / да.1347. ЧВК 5074380. PMID 16498704.
- ^ а б c Виксон Дж (2002). «Рекомендуемый организм: Schizosaccharomyces pombe, делящиеся дрожжи». Сравнительная и функциональная геномика. 3 (2): 194–204. Дои:10.1002 / cfg.92. ЧВК 2447254. PMID 18628834.
- ^ а б c d е Форсбург SL. «ПомбеНет». Получено 7 февраля 2016.
- ^ а б c d е ж Вуд В., Гвиллиам Р., Раджандрим М.А., Лайн М., Лайн Р., Стюарт А. и др. (Февраль 2002 г.). «Последовательность генома Schizosaccharomyces pombe». Природа. 415 (6874): 871–80. Bibcode:2002Натурал.415..871Вт. Дои:10.1038 / природа724. PMID 11859360.
- ^ а б Das M, Wiley DJ, Medina S, Vincent HA, Larrea M, Oriolo A, Verde F (июнь 2007 г.). «Регулирование диаметра клеток, локализации For3p и симметрии клеток с помощью делящихся дрожжей Rho-GAP Rga4p». Молекулярная биология клетки. 18 (6): 2090–101. Дои:10.1091 / mbc.E06-09-0883. ЧВК 1877093. PMID 17377067.
- ^ а б c Кавасима С.А., Такемото А., медсестра П., Капур Т.М. (июль 2012 г.). «Анализ механизмов множественной лекарственной устойчивости делящихся дрожжей для разработки генетически управляемой модельной системы для химической биологии». Химия и биология. 19 (7): 893–901. Дои:10.1016 / j.chembiol.2012.06.008. ЧВК 3589755. PMID 22840777.
- ^ а б Тай З., Энг Р.Дж., Саджики К., Лим К.К., Тан М.Ю., Янагида М., Чен Э.С. (24 января 2013 г.). «Клеточная устойчивость, обеспечиваемая генетическими перекрестными помехами, лежит в основе устойчивости к химиотерапевтическому препарату доксорубицину у делящихся дрожжей». PLOS ONE. 8 (1): e55041. Bibcode:2013PLoSO ... 855041T. Дои:10.1371 / journal.pone.0055041. ЧВК 3554685. PMID 23365689.
- ^ Forsburg SL (сентябрь 1999 г.). «Лучшие дрожжи?». Тенденции в генетике. 15 (9): 340–4. Дои:10.1016 / s0168-9525 (99) 01798-9. PMID 10461200.
- ^ Хоффман К.С., Вуд V, Фантес ПА (октябрь 2015 г.). "Древние дрожжи для молодых генетиков: учебник по модельной системе Schizosaccharomyces pombe". Генетика. 201 (2): 403–23. Дои:10.1534 / genetics.115.181503. ЧВК 4596657. PMID 26447128.
- ^ Sabatinos SA, Mastro TL, Green MD, Forsburg SL (январь 2013 г.). "Реакция делящихся дрожжей на повреждение ДНК у млекопитающих на аналоги нуклеозидов". Генетика. 193 (1): 143–57. Дои:10.1534 / генетика.112.145730. ЧВК 3527242. PMID 23150603.
- ^ а б Хаяно М., Кано И, Мацумото С., Ренар-Гийе С., Сирахиге К., Масаи Х (январь 2012 г.). «Rif1 является глобальным регулятором времени запуска репликации в делящихся дрожжах». Гены и развитие. 26 (2): 137–50. Дои:10.1101 / gad.178491.111. ЧВК 3273838. PMID 22279046.
- ^ "PomBase Series Edition". EMBL-Европейский институт биоинформатики. Wellcome Trust Genome Campus Place Издатель: Хинкстон, Кембридж.
- ^ Буррак Л.С., Берман Дж. (Июль 2012 г.). «Неоцентромеры и эпигенетически наследуемые особенности центромер». Хромосомные исследования. 20 (5): 607–19. Дои:10.1007 / s10577-012-9296-х. ЧВК 3409321. PMID 22723125.
- ^ а б Стимпсон К.М., Матени Дж. Э., Салливан Б. А. (июль 2012 г.). «Дицентрические хромосомы: уникальные модели для изучения функции и инактивации центромер». Хромосомные исследования. 20 (5): 595–605. Дои:10.1007 / s10577-012-9302-3. ЧВК 3557915. PMID 22801777.
- ^ McDowall MD, Harris MA, Lock A, Rutherford K, Staines DM, Bähler J, et al. (Январь 2015 г.). «PomBase 2015: обновления базы данных по делящимся дрожжам». Исследования нуклеиновых кислот. 43 (Выпуск базы данных): D656-61. Дои:10.1093 / нар / gku1040. ЧВК 4383888. PMID 25361970.
- ^ Кадура С., Сазер С. (июль 2005 г.). "Митотические ошибки в SAC: как контрольная точка сборки веретена (SAC) защищает от неправильной сегрегации хромосом". Подвижность клеток и цитоскелет. 61 (3): 145–60. Дои:10,1002 / см. 2007. PMID 15887295.
- ^ а б c d Ли Ай Джей, Коффман В.К., Ву Дж.К. (октябрь 2012 г.). «Сборка сократительного кольца в цитокинезе делящихся дрожжей: последние достижения и новые перспективы». Цитоскелет. 69 (10): 751–63. Дои:10.1002 / см. 21052. ЧВК 5322539. PMID 22887981.
- ^ а б Ринальди Т., Даллабона С., Ферреро И., Фронтали Л., Болотин-Фукухара М. (декабрь 2010 г.). «Митохондриальные заболевания и роль дрожжевых моделей». FEMS дрожжевые исследования. 10 (8): 1006–22. Дои:10.1111 / j.1567-1364.2010.00685.x. PMID 20946356.
- ^ а б c d Дэниел С. Джеффарес и др. (2015). Геномное и фенотипическое разнообразие Schizosaccharomyces pombe Nature Genetics 47, 235–241 doi: 10,1038 / ng.3215
- ^ Brown et al. (2011) Географически разнообразная коллекция изолятов Schizosaccharomyces pombe демонстрирует ограниченное фенотипическое разнообразие, но обширное кариотипическое разнообразие. G3 (Bethesda) 7: 615-26 DOI: 10.1534 / g3.111.001123.
- ^ Мохардин Л., Васкес Э., Антекера Ф. (ноябрь 2013 г.). «Спецификация происхождения репликации ДНК и состава геномных оснований у делящихся дрожжей». Журнал молекулярной биологии. 425 (23): 4706–13. Дои:10.1016 / j.jmb.2013.09.023. HDL:10261/104754. PMID 24095860.
- ^ а б Forsburg SL (апрель 2002 г.). «Только соединение: связывание репликации мейотической ДНК с динамикой хромосом». Молекулярная клетка. 9 (4): 703–11. Дои:10.1016 / S1097-2765 (02) 00508-7. PMID 11983163.
- ^ Мория Х., Чино А., Капуй О., Чикаш-Надь А., Новак Б. (декабрь 2011 г.). «Пределы сверхэкспрессии регуляторов клеточного цикла делящихся дрожжей in vivo и in silico». Молекулярная системная биология. 7 (1): 556. Дои:10.1038 / msb.2011.91. ЧВК 3737731. PMID 22146300.
- ^ Das M, Wiley DJ, Chen X, Shah K, Verde F (август 2009 г.). «Консервативная киназа NDR Orb6 контролирует рост поляризованных клеток с помощью пространственной регуляции малой GTPase Cdc42». Текущая биология. 19 (15): 1314–9. Дои:10.1016 / j.cub.2009.06.057. PMID 19646873. S2CID 12744756.
- ^ Мозли Дж. Б. (октябрь 2013 г.). «Клеточное старение: симметрия ускользает от старения». Текущая биология. 23 (19): R871-3. Дои:10.1016 / j.cub.2013.08.013. ЧВК 4276399. PMID 24112980.
- ^ Купер С (2013). «Schizosaccharomyces pombe растет экспоненциально в течение цикла деления без точек изменения скорости» (PDF). FEMS дрожжи Res. 13 (7): 650–8. Дои:10.1111/1567-1364.12072. PMID 23981297.
- ^ Сено Т., Кавано С., Икеда С. (март 2017 г.). «Эксцизионная репарация оснований ДНК и эксцизионная репарация нуклеотидов синергетически способствуют выживанию стационарных клеток делящихся дрожжей Schizosaccharomyces pombe». Cell Biology International. 41 (3): 276–286. Дои:10.1002 / cbin.10722. PMID 28032397.
- ^ Cadou A, Couturier A, Le Goff C, Xie L, Paulson JR, Le Goff X (март 2013 г.). «Киназа Kin1 и кальциневринфосфатаза взаимодействуют, чтобы связать сборку актинового кольца и синтез перегородки у делящихся дрожжей». Биология клетки. 105 (3): 129–48. Дои:10.1111 / boc.201200042. PMID 23294323.
- ^ Balazs A, Batta G, Miklos I., Acs-Szabo L, Vazquez de Aldana CR, Sipiczki M (март 2012 г.). «Консервативные регуляторы процесса разделения клеток у Schizosaccharomyces». Грибковая генетика и биология. 49 (3): 235–49. Дои:10.1016 / j.fgb.2012.01.003. HDL:10261/51389. PMID 22300943.
- ^ Ринкон С.А., Паолетти А. (октябрь 2012 г.). «Mid1 / аниллин и пространственная регуляция цитокинеза у делящихся дрожжей». Цитоскелет. 69 (10): 764–77. Дои:10.1002 / см. 21056. PMID 22888038.
- ^ Дас М, Хирон С, Верде Ф (2010). «Зависимая от микротрубочек пространственная организация митохондрий у делящихся дрожжей». Микротрубочки: In vivo. Методы клеточной биологии. 97. С. 203–21. Дои:10.1016 / S0091-679X (10) 97012-X. ISBN 9780123813497. PMID 20719273.
- ^ Фрейзер HB (2013). «Транскрипция, регулируемая клеточным циклом, связана со временем репликации ДНК у дрожжей и человека». Геномная биология. 14 (10): R111. arXiv:1308.1985. Дои:10.1186 / gb-2013-14-10-r111. ЧВК 3983658. PMID 24098959.
- ^ Ли П.К., Грин MD, Форсбург С.Л. (2013). «Мутации, нарушающие метилирование гистонов, по-разному влияют на время репликации в центромере S. pombe». PLOS ONE. 8 (5): e61464. Bibcode:2013PLoSO ... 861464L. Дои:10.1371 / journal.pone.0061464. ЧВК 3641051. PMID 23658693.
- ^ "Сэр Пол Медсестра - Биографический". Официальный сайт Нобелевской премии. 2001. Получено 7 февраля 2016.
- ^ Чжао Дж., Лендаль У., Нистер М. (март 2013 г.). «Регулирование митохондриальной динамики: конвергенции и расхождения между дрожжами и позвоночными». Клеточные и молекулярные науки о жизни. 70 (6): 951–76. Дои:10.1007 / s00018-012-1066-6. ЧВК 3578726. PMID 22806564.
- ^ Абеловская Л (2011). «Митохондрии как протеановые органеллы: мембранные процессы, влияющие на форму митохондрий у дрожжей». Общая физиология и биофизика. 30 Спецификация № (5): S13-24. Дои:10.4149 / gpb_2011_SI1_13. PMID 21869447.
- ^ Чино А., Маканаэ К., Мория Х. (3 сентября 2013 г.). «Взаимосвязь между числом копий гена регулятора клеточного цикла и уровнями экспрессии белка у Schizosaccharomyces pombe». PLOS ONE. 8 (9): e73319. Bibcode:2013PLoSO ... 873319C. Дои:10.1371 / journal.pone.0073319. ЧВК 3760898. PMID 24019917.
- ^ Райчаудхури С., Янг Б. П., Эспеншад П. Дж., Лёвен С. (август 2012 г.). «Регулирование липидного обмена: сказка о двух дрожжах». Текущее мнение в области клеточной биологии. 24 (4): 502–8. Дои:10.1016 / j.ceb.2012.05.006. ЧВК 4339016. PMID 22694927.
- ^ Бабу М., Власблом Дж., Пу С., Го Х, Грэм С., Бин Б. Д. и др. (Сентябрь 2012 г.). «Пейзаж взаимодействия мембранно-белковых комплексов у Saccharomyces cerevisiae». Природа. 489 (7417): 585–9. Bibcode:2012Натура 489..585Б. Дои:10.1038 / природа11354. PMID 22940862. S2CID 4344457.
- ^ Суда Й., Накано А. (апрель 2012 г.). «Дрожжевой аппарат Гольджи». Трафик. 13 (4): 505–10. Дои:10.1111 / j.1600-0854.2011.01316.x. PMID 22132734.
- ^ «Инициатива Trans-NIH.pombe». 2002. Цитировать журнал требует
| журнал =(помощь) - ^ Грин, М. Д. Сабатинос, С. А. Форсбург, С. Л. (2009). Методы микроскопии для изучения репликации ДНК у делящихся дрожжей Journal. Методы молекулярной биологии. 521. С. 463–82. Дои:10.1007/978-1-60327-815-7_26. ISBN 978-1-60327-814-0. PMID 19563123.CS1 maint: несколько имен: список авторов (связь)