Хромосома - Википедия - Chromosome
Эта статья может быть слишком техническим для большинства читателей, чтобы понять. (Апрель 2017 г.) (Узнайте, как и когда удалить этот шаблон сообщения) |
| Часть серия на |
| Генетика |
|---|
 |
| Ключевые компоненты |
| История и темы |
| Исследование |
| Персонализированная медицина |
| Персонализированная медицина |

А хромосома длинный ДНК молекула с частью или полностью генетический материал организма. Наиболее эукариотический хромосомы включают упаковочные белки, называемые гистоны чему, при поддержке белки-шапероны, привязать к и конденсировать молекула ДНК для поддержания ее целостности.[1][2] Эти хромосомы имеют сложную трехмерную структуру, которая играет важную роль в транскрипционная регуляция.[3]
Хромосомы обычно видны под оптический микроскоп только во время метафаза из деление клеток (где все хромосомы выровнены по центру клетки в их уплотненной форме).[4] Прежде чем это произойдет, каждая хромосома дублируется (S фаза ), и обе копии соединены центромера, в результате получается Х-образная структура (на фото выше), если центромера располагается экваториально или двуплечий, если центромера расположена дистально. Объединенные копии теперь называются сестринские хроматиды. Во время метафазы Х-образная структура называется метафазной хромосомой, которая сильно уплотнена и поэтому ее легче всего различить и изучить.[5] В клетках животных хромосомы достигают наивысшего уровня уплотнения в анафаза в течение расщепление хромосом.[6]
Хромосомный рекомбинация в течение мейоз и последующие половое размножение играть важную роль в генетическое разнообразие. Если этими структурами манипулировать неправильно в результате процессов, известных как хромосомная нестабильность и транслокация, клетка может подвергнуться митотическая катастрофа. Обычно это заставляет ячейку инициировать апоптоз приводит к собственной гибели, но иногда мутации в клетке препятствуют этому процессу и, таким образом, вызывают прогрессирование рака.
Некоторые используют термин хромосома в более широком смысле, чтобы обозначить индивидуализированные части хроматин в клетках, видимых или невидимых под световой микроскопией. Другие используют это понятие в более узком смысле, чтобы относиться к индивидуализированным частям хроматина во время деления клеток, видимым под световой микроскопией из-за высокой конденсации.
Этимология
Слово хромосома (/ˈkрoʊмəˌsoʊм,-ˌzoʊм/[7][8]) происходит из Греческий χρῶμα (цветность, "цвет") и σῶμα (сома, «тело»), характеризуя их сильное окрашивание особым красители.[9] Термин был придуман немецким анатомом. Генрих Вильгельм Вальдейер,[10] ссылаясь на термин хроматин, который был введен Вальтер Флемминг, первооткрыватель деление клеток.
Некоторые из ранних кариологических терминов устарели.[11][12] Например, Хроматин (Flemming 1880) и Хромосома (Waldeyer 1888) приписывают цвет неокрашенному состоянию.[13]
История открытия


Немецкие ученые Schleiden,[5] Вирхов и Bütschli были одними из первых ученых, которые распознали структуры, теперь известные как хромосомы.[14]
В серии экспериментов, начавшейся в середине 1880-х гг., Теодор Бовери внесли решающий вклад в выяснение того, что хромосомы являются векторов наследственности, с двумя понятиями, которые стали известны как «хромосомная непрерывность» и «хромосомная индивидуальность».[15]
Вильгельм Ру предположил, что каждая хромосома несет разные генетическая конфигурация, и Бовери смог проверить и подтвердить эту гипотезу. Благодаря повторному открытию в начале 1900-х гг. Грегор Мендель В более ранней работе Бовери смог указать на связь между правилами наследования и поведением хромосом. Бовери оказал влияние на два поколения американских цитологов: Эдмунд Бичер Уилсон, Нетти Стивенс, Уолтер Саттон и Теофил Живописец все находились под влиянием Бовери (на самом деле с ним работали Уилсон, Стивенс и Пейнтер).[16]
В его знаменитом учебнике Клетка в развитии и наследственности, Уилсон соединил независимые работы Бовери и Саттона (около 1902 г.), назвав хромосомную теорию наследования Хромосомная теория Бовери – Саттона (имена иногда меняют местами).[17] Эрнст Майр отмечает, что эту теорию горячо оспаривали некоторые известные генетики: Уильям Бейтсон, Вильгельм Йоханссен, Ричард Гольдшмидт и T.H. Морган, все с довольно догматичным складом ума. В конце концов, полное доказательство было получено на основе карт хромосом в собственной лаборатории Моргана.[18]
Число хромосом человека было опубликовано в 1923 г. Теофил Художник. Посмотрев в микроскоп, он насчитал 24 пары, что означало 48 хромосом. Его ошибка была скопирована другими, и только в 1956 году истинное число, 46, было определено цитогенетиком из Индонезии. Джо Хин Тжио.[19]
Прокариоты
В прокариоты - бактерии и археи - обычно есть один круговая хромосома, но существует множество вариаций.[20] Хромосомы большинства бактерий, которые некоторые авторы предпочитают называть генофоры, может варьироваться от 130 000 пар оснований в эндосимбиотический бактерии Candidatus Hodgkinia cicadicola[21] и Кандидат Тремблая принцепс,[22] до более чем 14000000 пар оснований в почвенной бактерии Сорангиум целлюлозум.[23] Спирохеты из род Borrelia являются заметным исключением из этого правила, с такими бактериями, как Borrelia burgdorferi, причина Болезнь Лайма, содержащий один линейный хромосома.[24]
Структура в последовательностях
Прокариотические хромосомы имеют менее последовательную структуру, чем эукариоты. Бактерии обычно имеют одну точку ( начало репликации ), с которого начинается репликация, тогда как некоторые археи содержат несколько источников репликации.[25] Гены прокариот часто организованы в опероны, и обычно не содержат интроны, в отличие от эукариот.
Упаковка ДНК
Прокариоты не обладают ядрами. Вместо этого их ДНК организована в структуру, называемую нуклеоид.[26][27] Нуклеоид представляет собой отдельную структуру и занимает определенную область бактериальной клетки. Эта структура, однако, динамична, поддерживается и модифицируется под действием ряда гистоноподобных белков, которые связаны с бактериальной хромосомой.[28] В археи ДНК в хромосомах еще более организована, ДНК упакована в структуры, подобные эукариотическим нуклеосомам.[29][30]
Некоторые бактерии также содержат плазмиды или другой внехромосомная ДНК. Это круглые конструкции в цитоплазма которые содержат клеточную ДНК и играют роль в горизонтальный перенос генов.[5] У прокариот (см. нуклеоиды ) и вирусы,[31] ДНК часто плотно упакована и организована; в случае археи, по гомологии с эукариотическими гистонами, а в случае бактерий - по гистоновый белки.
Бактериальные хромосомы имеют тенденцию быть привязанными к плазматическая мембрана бактерий. В приложениях молекулярной биологии это позволяет изолировать его от плазмидной ДНК центрифугированием лизированных бактерий и осаждением мембран (и прикрепленной ДНК).
Прокариотические хромосомы и плазмиды, как и эукариотическая ДНК, обычно суперскрученный. ДНК должна быть сначала переведена в расслабленное состояние для доступа транскрипция, регулирование и репликация.
Эукариоты
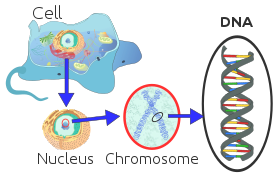
Каждая эукариотическая хромосома состоит из длинной линейной молекулы ДНК, связанной с белками, образуя компактный комплекс белков и ДНК, называемый хроматин. Хроматин содержит подавляющее большинство ДНК организма, но небольшое количество наследуется по материнской линии, можно найти в митохондрии. Он присутствует в большинстве клетки, за некоторыми исключениями, например, красные кровяные тельца.
Гистоны отвечают за первую и самую основную единицу организации хромосомы, нуклеосома.
Эукариоты (клетки с ядрами, такими как ядра растений, грибов и животных) обладают множеством больших линейных хромосом, содержащихся в ядре клетки. В каждой хромосоме по одному центромера, с одним или двумя плечами, выступающими из центромеры, хотя в большинстве случаев эти руки не видны как таковые. Кроме того, у большинства эукариот есть небольшой круговой митохондриальный геном, а некоторые эукариоты могут иметь дополнительные маленькие круглые или линейные цитоплазматический хромосомы.
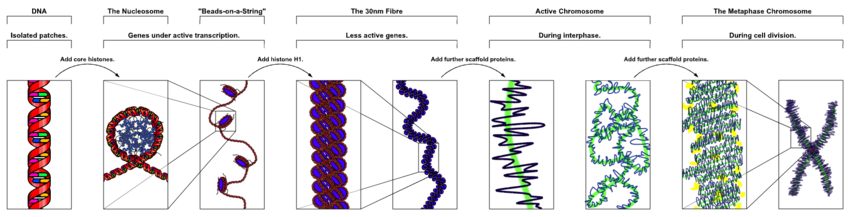
В ядерных хромосомах эукариоты, неконденсированная ДНК существует в полуупорядоченной структуре, где она обернута вокруг гистоны (структурные белки ), образуя композитный материал, называемый хроматин.
Интерфазный хроматин
Упаковка ДНК в нуклеосомы приводит к образованию 10-нанометрового волокна, которое может дополнительно конденсировать до 30-нанометрового волокна.[32] Большая часть эухроматина в интерфазных ядрах, по-видимому, имеет форму 30-нм волокон.[32] Структура хроматина - это более деконденсированное состояние, то есть конформация 10 нм допускает транскрипцию.[32]
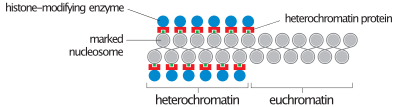
В течение межфазный (период клеточный цикл где клетка не делится), два типа хроматин можно выделить:
- Эухроматин, который состоит из активной ДНК, например, выраженной в виде белка.
- Гетерохроматин, который состоит в основном из неактивной ДНК. Похоже, он служит структурным целям на хромосомных стадиях. Гетерохроматин можно разделить на два типа:
- Конститутивный гетерохроматин, который никогда не выражается. Он расположен вокруг центромеры и обычно содержит повторяющиеся последовательности.
- Факультативный гетерохроматин, что иногда выражается.
Метафазный хроматин и деление


На ранних этапах митоз или же мейоз (деление клетки) двойная спираль хроматина становится все более конденсированной. Они перестают функционировать как доступный генетический материал (транскрипция остановок) и стать компактной транспортабельной формой. Считается, что петли 30-нм хроматиновых волокон дополнительно складываются сами по себе, образуя компактные метафазные хромосомы митотических клеток. Таким образом, ДНК конденсируется примерно в 10 000 раз.[32]
Хромосомный каркас, состоящий из белков, таких как конденсин, TOP2A и KIF4,[33] играют важную роль в удерживании хроматина в компактной хромосоме. Петли структуры 30 нм далее конденсируются с каркасом в структуры более высокого порядка.[34]
Эта очень компактная форма делает отдельные хромосомы видимыми, и они образуют классическую четырехлепестковую структуру, сестринскую пару. хроматиды прикреплены друг к другу на центромера. Более короткие руки называются р руки (от французского petit, малые), а более длинные руки называются q руки (q следует п в латинском алфавите; q-g "гранде"; в качестве альтернативы иногда говорят, что q - это сокращение от очередь что означает хвост по-французски[35]). Это единственный естественный контекст, в котором отдельные хромосомы видны оптическим микроскоп.
Митотические метафазные хромосомы лучше всего описываются линейно организованным продольно сжатым массивом последовательных петель хроматина.[36]
Во время митоза микротрубочки растут из центросом, расположенных на противоположных концах клетки, а также прикрепляются к центромере в специализированных структурах, называемых кинетохоры, по одному на каждой сестре хроматида. Особая последовательность оснований ДНК в области кинетохор обеспечивает, наряду со специальными белками, более длительное прикрепление в этой области. Затем микротрубочки раздвигают хроматиды в сторону центросом, так что каждая дочерняя клетка наследует один набор хроматид. После разделения клеток хроматиды разворачиваются, и ДНК снова может быть расшифрована. Несмотря на их внешний вид, хромосомы структурно сильно конденсированы, что позволяет этим гигантским структурам ДНК содержаться в ядре клетки.
Хромосомы человека
Хромосомы у человека можно разделить на два типа: аутосомы (хромосома (ы) тела) и аллосома (половая хромосома (s)). Определенные генетические особенности связаны с полом человека и передаются через половые хромосомы. Аутосомы содержат остальную генетическую наследственную информацию. Все действуют одинаково во время деления клеток. Клетки человека имеют 23 пары хромосом (22 пары аутосом и одна пара половых хромосом), то есть всего 46 на клетку. В дополнение к этому, человеческие клетки имеют многие сотни копий митохондриальный геном. Последовательность действий из человеческий геном предоставил много информации о каждой из хромосом. Ниже представлена таблица, в которой собрана статистика по хромосомам, основанная на Институт Сэнгера информация о геноме человека в База данных аннотаций генома позвоночных (VEGA).[37] Количество генов является приблизительным, так как оно частично основано на генные предсказания. Общая длина хромосомы также является оценкой, основанной на предполагаемом размере несеквенированных гетерохроматин регионы.

| Хромосома | Гены[38] | Общий пар оснований | % баз | Последовательные пары оснований[39] | % секвенированных пар оснований |
|---|---|---|---|---|---|
| 1 | 2000 | 247,199,719 | 8.0 | 224,999,719 | 91.02% |
| 2 | 1300 | 242,751,149 | 7.9 | 237,712,649 | 97.92% |
| 3 | 1000 | 199,446,827 | 6.5 | 194,704,827 | 97.62% |
| 4 | 1000 | 191,263,063 | 6.2 | 187,297,063 | 97.93% |
| 5 | 900 | 180,837,866 | 5.9 | 177,702,766 | 98.27% |
| 6 | 1000 | 170,896,993 | 5.5 | 167,273,993 | 97.88% |
| 7 | 900 | 158,821,424 | 5.2 | 154,952,424 | 97.56% |
| 8 | 700 | 146,274,826 | 4.7 | 142,612,826 | 97.50% |
| 9 | 800 | 140,442,298 | 4.6 | 120,312,298 | 85.67% |
| 10 | 700 | 135,374,737 | 4.4 | 131,624,737 | 97.23% |
| 11 | 1300 | 134,452,384 | 4.4 | 131,130,853 | 97.53% |
| 12 | 1100 | 132,289,534 | 4.3 | 130,303,534 | 98.50% |
| 13 | 300 | 114,127,980 | 3.7 | 95,559,980 | 83.73% |
| 14 | 800 | 106,360,585 | 3.5 | 88,290,585 | 83.01% |
| 15 | 600 | 100,338,915 | 3.3 | 81,341,915 | 81.07% |
| 16 | 800 | 88,822,254 | 2.9 | 78,884,754 | 88.81% |
| 17 | 1200 | 78,654,742 | 2.6 | 77,800,220 | 98.91% |
| 18 | 200 | 76,117,153 | 2.5 | 74,656,155 | 98.08% |
| 19 | 1500 | 63,806,651 | 2.1 | 55,785,651 | 87.43% |
| 20 | 500 | 62,435,965 | 2.0 | 59,505,254 | 95.31% |
| 21 | 200 | 46,944,323 | 1.5 | 34,171,998 | 72.79% |
| 22 | 500 | 49,528,953 | 1.6 | 34,893,953 | 70.45% |
| X (половая хромосома) | 800 | 154,913,754 | 5.0 | 151,058,754 | 97.51% |
| Y (половая хромосома) | 200[40] | 57,741,652 | 1.9 | 25,121,652 | 43.51% |
| Общий | 21,000 | 3,079,843,747 | 100.0 | 2,857,698,560 | 92.79% |
Количество в различных организмах
У эукариот
В этих таблицах указано общее количество хромосом (включая половые хромосомы) в ядре клетки. Например, большинство эукариоты находятся диплоид, подобно люди у которых 22 разных типа аутосомы, каждая представлена в виде двух гомологичных пар и двух половые хромосомы. Всего получается 46 хромосом. Другие организмы имеют более двух копий своих типов хромосом, например хлеб пшеничный, который гексаплоид и имеет шесть копий семи разных типов хромосом - всего 42 хромосомы.
|
|
|
Нормальные члены определенного эукариот разновидность у всех одинаковое количество ядерных хромосом (см. таблицу). Другие эукариотические хромосомы, то есть митохондриальные и плазмидоподобные маленькие хромосомы, гораздо более разнообразны по количеству, и могут быть тысячи копий на клетку.

Бесполое размножение виды имеют один набор хромосом, одинаковый во всех клетках тела. Однако бесполые виды могут быть гаплоидными или диплоидными.
Половое размножение виды имеют соматические клетки (клетки тела), которые диплоид [2n], имеющий два набора хромосом (23 пары у людей с одним набором из 23 хромосом от каждого родителя), один набор от матери и один от отца. Гаметы, репродуктивные клетки, являются гаплоидный [n]: У них один набор хромосом. Гаметы производятся мейоз диплоида линия зародыша клетка. Во время мейоза совпадающие хромосомы отца и матери могут обмениваться небольшими частями самих себя (кроссовер ) и, таким образом, создают новые хромосомы, которые не наследуются исключительно от одного из родителей. Когда мужская и женская гамета сливаются (оплодотворение ) образуется новый диплоидный организм.
Некоторые виды животных и растений полиплоид [Xn]: у них более двух наборов гомологичные хромосомы. Важные в сельском хозяйстве растения, такие как табак или пшеница, часто полиплоидны по сравнению с их предковыми видами. Пшеница имеет гаплоидное число из семи хромосом, которое все еще встречается в некоторых сорта а также диких предков. Более распространенные типы макаронных изделий и мягкой пшеницы являются полиплоидными и имеют 28 (тетраплоид) и 42 (гексаплоид) хромосомы по сравнению с 14 (диплоидными) хромосомами у дикой пшеницы.[66]
У прокариот
Прокариот разновидность обычно имеют по одной копии каждой основной хромосомы, но большинство клеток могут легко выжить с несколькими копиями.[67] Например, Бухнера, а симбионт из тля имеет несколько копий своей хромосомы, от 10 до 400 копий на клетку.[68] Однако у некоторых крупных бактерий, таких как Epulopiscium fishelsoni может присутствовать до 100 000 копий хромосомы.[69] Плазмиды и маленькие плазмидоподобные хромосомы, как и у эукариот, сильно различаются по количеству копий. Количество плазмид в клетке почти полностью определяется скоростью деления плазмиды - быстрое деление вызывает большое количество копий.
Кариотип

В целом кариотип это характерный хромосомный набор эукариот разновидность.[70] Подготовка и исследование кариотипов является частью цитогенетика.
Хотя репликация и транскрипция из ДНК очень стандартизирован в эукариоты, то же самое нельзя сказать об их кариотипах, которые часто сильно различаются. Между видами могут быть различия в количестве хромосом и в детальной организации, а в некоторых случаях - внутри видов. Часто бывает:
- 1. различия между двумя полами
- 2. разница между зародышевый путь и сома (между гаметы и остальное тело)
- 3. различия между членами популяции из-за сбалансированный генетический полиморфизм
- 4. географическое разнообразие между скачки
- 5. мозаика или другие ненормальные люди.
Кроме того, вариации кариотипа могут происходить во время развития от оплодотворенной яйцеклетки.
Методика определения кариотипа обычно называется кариотипирование. Клетки могут быть заблокированы частично через деление (в метафазе) in vitro (в реакционном флаконе) с колхицин. Затем эти клетки окрашивают, фотографируют и помещают в кариограмма, с расположенным набором хромосом, аутосомами в порядке длины и половыми хромосомами (здесь X / Y) в конце.
Как и у многих видов, размножающихся половым путем, у людей есть особые гоносомы (половые хромосомы, в отличие от аутосомы ). Это XX у женщин и XY у мужчин.
История и методы анализа
Изучение кариотипа человека заняло много лет, чтобы решить самый главный вопрос: Сколько хромосом у нормального диплоид клетка человека содержит? В 1912 г. Ханс фон Винивартер сообщили о 47 хромосомах в сперматогония и 48 в оогония, завершая XX / XO механизм определения пола.[71] Художник в 1922 году не было уверенности, 46 или 48 диплоидных чисел людей, поначалу отдавая предпочтение 46.[72] Позже он пересмотрел свое мнение с 46 на 48, и он правильно настаивал на том, что у людей есть XX / XY система.[73]
Для окончательного решения проблемы потребовались новые методы:
- Использование клеток в культуре
- Арест митоз в метафаза решением колхицин
- Предварительная обработка клеток в гипотонический раствор 0,075 M KCl, который их набухает и распространяет хромосомы
- Сдавить препарат на слайде, вынуждая хромосомы в единую плоскость
- Разрезание микрофотографии и преобразование результата в бесспорную кариограмму.
Только в 1954 году диплоидное число человека было подтверждено как 46.[74][75] Учитывая методы Winiwarter и Painter, их результаты были весьма замечательными.[76] Шимпанзе, самые близкие родственники современного человека, имеют 48 хромосом, как и другие большие обезьяны: у человека две хромосомы слились в хромосома 2.
Аберрации
Хромосомные аберрации - это нарушения нормального хромосомного содержимого клетки и основная причина генетических заболеваний у людей, таких как Синдром Дауна, хотя большинство аберраций практически не влияют. Некоторые хромосомные аномалии не вызывают заболевания у носителей, например: транслокации, или же хромосомные инверсии, хотя они могут повысить вероятность вынашивания ребенка с хромосомным заболеванием. Аномальное количество хромосом или хромосомных наборов, называемое анеуплоидия может привести к летальному исходу или вызвать генетические нарушения.[77] Генетическое консультирование предлагается семьям, которые могут иметь хромосомную перестройку.
Приобретение или потеря ДНК хромосом может привести к различным генетические нарушения. Примеры людей включают:
- Кри дю чат, что вызвано удаление части короткого плеча хромосомы 5. «Cri du chat» по-французски означает «кошачий крик»; состояние было названо так потому, что пораженные дети издают пронзительные крики, похожие на кошачьи. Больные имеют широко расставленные глаза, небольшую голову и челюсть, умеренные или серьезные проблемы с психическим здоровьем и очень низкий рост.
- Синдром Дауна, наиболее распространенная трисомия, обычно вызванная дополнительной копией хромосомы 21 (трисомия 21 ). Характеристики включают снижение мышечного тонуса, более коренастое телосложение, асимметричный череп, раскосые глаза и нарушение развития от легкой до средней степени.[78]
- Синдром Эдвардса, или трисомия-18, вторая по распространенности трисомия.[79] Симптомы включают задержку моторики, нарушение развития и многочисленные врожденные аномалии, вызывающие серьезные проблемы со здоровьем. Девяносто процентов заболевших умирают в младенчестве. У них характерные сжатые руки и пальцы внахлест.
- Изодицентрик 15, также называемый idic (15), частичной тетрасомией 15q или инвертированной дупликацией 15 (inv dup 15).
- Синдром Якобсена, что бывает очень редко. Это также называется терминальным нарушением делеции 11q.[80] Пострадавшие имеют нормальный интеллект или легкие отклонения в развитии с плохими навыками выразительной речи. У большинства есть нарушение свертываемости крови, называемое Синдром Пари-Труссо.
- Синдром Клайнфельтера (XXY). Мужчины с синдромом Клайнфельтера обычно бесплодны, обычно выше и имеют более длинные руки и ноги, чем их сверстники. Мальчики с синдромом часто застенчивы и тихи, у них чаще встречается задержка речи и дислексия. Без лечения тестостероном у некоторых может развиться гинекомастия в период полового созревания.
- Синдром Патау, также называемый D-синдромом или трисомией-13. Симптомы несколько схожи с симптомами трисомии-18, но без характерной сложенной руки.
- Маленькая дополнительная маркерная хромосома. Это означает, что есть лишняя аномальная хромосома. Характеристики зависят от происхождения дополнительного генетического материала. Синдром кошачьего глаза и синдром изодицентрической хромосомы 15 (или Idic15) оба вызваны дополнительным маркером хромосомы, как и Синдром Паллистера-Киллиана.
- Синдром Triple-X (ХХХ). XXX девушки, как правило, высокие и худые и чаще страдают дислексией.
- Синдром Тернера (X вместо XX или XY). При синдроме Тернера женские половые признаки присутствуют, но недостаточно развиты. Женщины с синдромом Тернера часто имеют низкий рост, низкую линию роста волос, аномальные черты глаз и развитие костей, а также вид «прогнутой» груди.
- Синдром Вольфа – Хиршхорна, который вызван частичным удалением короткого плеча хромосомы 4. Он характеризуется задержкой роста, задержкой развития моторики, чертами лица «греческого шлема» и легкими или серьезными проблемами психического здоровья.
- XYY синдром. Мальчики XYY обычно выше своих братьев и сестер. Как и у XXY мальчиков и XXX девочек, у них чаще возникают трудности с обучением.
Анеуплоидия спермы
Воздействие на мужчин определенного образа жизни, окружающей среды и / или профессиональных опасностей может увеличить риск анеуплоидных сперматозоидов.[81] В частности, риск анеуплоидии увеличивается при курении табака,[82][83] и профессиональное воздействие бензола,[84] инсектициды,[85][86] и перфторированные соединения.[87] Повышенная анеуплоидия часто связана с повышенным повреждением ДНК в сперматозоидах.
Смотрите также
- Анеуплоидия
- Хромомер
- Расщепление хромосом
- Cohesin
- Конденсин
- ДНК
- Генетическое удаление
- Эпигенетика
- Для получения информации о хромосомах в генетические алгоритмы, видеть хромосома (генетический алгоритм)
- Генетическая генеалогия
- Хромосома ламповой щетки
- Список количества хромосом различных организмов
- Locus (объясняет номенклатуру расположения генов)
- Материнское влияние на определение пола
- Не дизъюнкция
- Система определения пола
- Политенная хромосома
- Протамин
- Неохромосома
- Паразитарная хромосома
Примечания и ссылки
- ^ Хаммонд CM, Strømme CB, Huang H, Patel DJ, Groth A (март 2017 г.). «Гистоновые шаперонные сети, формирующие функцию хроматина». Обзоры природы. Молекулярная клеточная биология. 18 (3): 141–158. Дои:10.1038 / nrm.2016.159. ЧВК 5319910. PMID 28053344.
- ^ Уилсон, Джон (2002). Молекулярная биология клетки: проблемный подход. Нью-Йорк: Наука о гирляндах. ISBN 978-0-8153-3577-1.
- ^ Бонев, Боян; Кавалли, Джакомо (14 октября 2016 г.). «Организация и функции трехмерного генома». Природа Обзоры Генетика. 17 (11): 661–678. Дои:10.1038 / nrg.2016.112. HDL:2027.42/151884. PMID 27739532. S2CID 31259189.
- ^ Альбертс Б., Брей Д., Хопкин К., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П. (2014). Эссенциальная клеточная биология (Четвертое изд.). Нью-Йорк, Нью-Йорк, США: Garland Science. С. 621–626. ISBN 978-0-8153-4454-4.
- ^ а б c Шлейден, М. Дж. (1847). Микроскопические исследования соответствия строения и роста животных и растений..
- ^ Антонин В, Нойман Х (июнь 2016 г.). «Конденсация и деконденсация хромосом во время митоза». Текущее мнение в области клеточной биологии. 40: 15–22. Дои:10.1016 / j.ceb.2016.01.013. PMID 26895139.
- ^ Джонс, Дэниел (2003) [1917], Питер Роуч; Джеймс Хартманн; Джейн Сеттер (ред.), Словарь английского произношения, Кембридж: Издательство Кембриджского университета, ISBN 978-3-12-539683-8
- ^ «Хромосома». Словарь Merriam-Webster.
- ^ Кокс, Х. Дж. (1925). Биологические пятна - Справочник по природе и использованию красителей, используемых в биологической лаборатории. Комиссия по стандартизации биологических пятен.
- ^ Вальдейер-Харц (1888 г.). "Über Karyokinese und ihre Beziehungen zu den Befruchtungsvorgängen". Archiv für Mikroskopische Anatomie und Entwicklungsmechanik. 32: 27.
- ^ Гарбари Ф., Бедини Дж., Перуцци Л. (2012). «Хромосомные числа итальянской флоры. От основания Caryologia до наших дней». Кариология - Международный журнал цитологии, цитосистематики и цитогенетики. 65 (1): 65–66. Дои:10.1080/00087114.2012.678090. S2CID 83748967.
- ^ Перуцци Л., Гарбари Ф, Бедини Г. (2012). «Новые тенденции в цитогенетике и цитоэмбриологии растений: памяти Эмилио Батталья». Биосистемы растений. 146 (3): 674–675. Дои:10.1080/11263504.2012.712553 (неактивно с 16 декабря 2020 г.).CS1 maint: DOI неактивен по состоянию на декабрь 2020 г. (связь)
- ^ Батталья, Эмилио (2009). «Карионема, альтернатива хромосоме и новая кариологическая номенклатура» (PDF). Caryologia - Международный журнал цитологии, цитосистематики. 62 (4): 1–80. Получено 6 ноября 2017.
- ^ Фокин С.И. (2013). "Отто Бютчли (1848–1920) Где мы преклоним колени?" (PDF). Протистология. 8 (1): 22–35.
- ^ Мадеспахер, Флориан (2008). «Теодор Бовери и естественный эксперимент». Текущая биология. 18 (7): R279 – R286. Дои:10.1016 / j.cub.2008.02.061. PMID 18397731. S2CID 15479331.
- ^ Карлсон, Элоф А. (2004). Наследие Менделя: происхождение классической генетики (PDF). Колд-Спринг-Харбор, Нью-Йорк: Лаборатория Колд-Спринг-Харбор. п. 88. ISBN 978-087969675-7.
- ^ Уилсон, Э. (1925). Клетка в развитии и наследственности, Ред. 3. Макмиллан, Нью-Йорк. п. 923.
- ^ Майр, Э. (1982). Рост биологической мысли. Гарвард. п. 749. ISBN 9780674364462
- ^ Мэтьюз, Роберт. «Причудливый случай хромосомы, которой никогда не было» (PDF). Архивировано из оригинал (PDF) 15 декабря 2013 г.. Получено 13 июля 2013.[самостоятельно опубликованный источник? ]
- ^ Танбихлер М., Шапиро Л. (ноябрь 2006 г.). «Хромосомная организация и сегрегация у бактерий». Журнал структурной биологии. 156 (2): 292–303. Дои:10.1016 / j.jsb.2006.05.007. PMID 16860572.
- ^ Ван Левен Дж. Т., Майстер Р. К., Саймон К., Маккатчеон Дж. П. (сентябрь 2014 г.). «Симпатрическое видообразование у бактериального эндосимбионта приводит к образованию двух геномов с функциональностью одного». Клетка. 158 (6): 1270–1280. Дои:10.1016 / j.cell.2014.07.047. PMID 25175626. S2CID 11839535.
- ^ McCutcheon JP, von Dohlen CD (август 2011 г.). «Взаимозависимый метаболический пэчворк во вложенном симбиозе мучнистых червецов». Текущая биология. 21 (16): 1366–72. Дои:10.1016 / j.cub.2011.06.051. ЧВК 3169327. PMID 21835622.
- ^ Хань К., Ли Цзыф, Пэн Р., Чжу Л.П., Чжоу Т., Ван Л.Г., Ли С.Г., Чжан ХБ, Ху В., Ву Чж, Цинь Н., Ли Ю.З. (2013). «Необычайное расширение генома Sorangium cellulosum из щелочной среды». Научные отчеты. 3: 2101. Bibcode:2013НатСР ... 3Э2101Н. Дои:10.1038 / srep02101. ЧВК 3696898. PMID 23812535.
- ^ Хиннебуш Дж., Тилли К. (декабрь 1993 г.). «Линейные плазмиды и хромосомы у бактерий». Молекулярная микробиология. 10 (5): 917–22. Дои:10.1111 / j.1365-2958.1993.tb00963.x. PMID 7934868. S2CID 23852021.
- ^ Кельман Л.М., Кельман З. (сентябрь 2004 г.). «Множественные источники репликации в архее». Тенденции в микробиологии. 12 (9): 399–401. Дои:10.1016 / j.tim.2004.07.001. PMID 15337158.
- ^ Танбихлер М., Ван С.К., Шапиро Л. (октябрь 2005 г.). «Бактериальный нуклеоид: высокоорганизованная и динамичная структура». Журнал клеточной биохимии. 96 (3): 506–21. Дои:10.1002 / jcb.20519. PMID 15988757. S2CID 25355087.
- ^ Ле ТБ, Имакаев М.В., Мирный Л.А., Лауб М.Т. (ноябрь 2013 г.). «Картирование пространственной организации бактериальной хромосомы с высоким разрешением». Наука. 342 (6159): 731–4. Bibcode:2013Наука ... 342..731Л. Дои:10.1126 / science.1242059. ЧВК 3927313. PMID 24158908.
- ^ Sandman K, Pereira SL, Reeve JN (декабрь 1998 г.). «Разнообразие прокариотических хромосомных белков и происхождение нуклеосом». Клеточные и молекулярные науки о жизни. 54 (12): 1350–64. Дои:10.1007 / с000180050259. PMID 9893710. S2CID 21101836.
- ^ Дрема К., Рив Дж. Н. (март 2000 г.). «Структура и функциональные отношения архейных и эукариальных гистонов и нуклеосом». Архив микробиологии. 173 (3): 165–9. Дои:10.1007 / s002039900122. PMID 10763747. S2CID 28946064.
- ^ Перейра С.Л., Грейлинг Р.А., Лурз Р., Рив Дж. Н. (ноябрь 1997 г.). «Архейские нуклеосомы». Труды Национальной академии наук Соединенных Штатов Америки. 94 (23): 12633–7. Bibcode:1997ПНАС ... 9412633П. Дои:10.1073 / пнас.94.23.12633. ЧВК 25063. PMID 9356501.
- ^ Джонсон Дж. Э., Чиу В. (апрель 2000 г.). «Структуры вирусов и вирусоподобных частиц». Текущее мнение в структурной биологии. 10 (2): 229–35. Дои:10.1016 / S0959-440X (00) 00073-7. PMID 10753814.
- ^ а б c d Купер, Г. (2019). Клетка (8-е изд.). Oxford University Press. ISBN 978-1605357072.
- ^ Пунперм, Равин; Таката, Хидеаки; Хамано, Тору; Мацуда, Ацуши; Учияма, Сусуму; Хираока, Ясуши; Фукуи, Киити (1 июля 2015 г.). «Хромосомный каркас представляет собой двухцепочечную сборку белков каркаса». Научные отчеты. 5 (1): 11916. Bibcode:2015НатСР ... 511916П. Дои:10.1038 / srep11916. ЧВК 4487240. PMID 26132639.
- ^ Lodish, U.H .; Lodish, H .; Берк, А .; Kaiser, C.A .; Kaiser, C .; Kaiser, U.C.A .; Krieger, M .; Scott, M.P .; Bretscher, A .; Ploegh, H .; другие (2008). Молекулярная клеточная биология. В. Х. Фриман. ISBN 978-0-7167-7601-7.
- ^ "Картирование хромосом: идиограммы " Природное образование - 13 августа 2013 г.
- ^ Наумова Н., Имакаев М., Фуденберг Г., Жан Й., Ладжуа Б.Р., Мирный Л.А., Деккер Дж. (Ноябрь 2013 г.). «Организация митотической хромосомы». Наука. 342 (6161): 948–53. Bibcode:2013Наука ... 342..948N. Дои:10.1126 / science.1236083. ЧВК 4040465. PMID 24200812.
- ^ Vega.sanger.ad.uk, все данные в этой таблице были получены из этой базы данных, 11 ноября 2008 г.
- ^ "Обозреватель ансамбльного генома 71: Homo sapiens - Сводка хромосом - Хромосома 1: 1–1 000 000". apr2013.archive.ensembl.org. Получено 11 апреля 2016.
- ^ Последовательные проценты основаны на доле эухроматина, поскольку Проект генома человека цели требовали определения только эухроматический часть генома. Теломеры, центромеры, и другие гетерохроматический регионы остались неопределенными, как и небольшое количество неклонируемых пробелов. Видеть https://www.ncbi.nlm.nih.gov/genome/seq/ для получения дополнительной информации о проекте "Геном человека".
- ^ Гены и болезнь. Бетесда, Мэриленд: Национальный центр биотехнологической информации. 1998 г.
- ^ Армстронг SJ, Джонс GH (январь 2003 г.). «Мейотическая цитология и поведение хромосом у Arabidopsis thaliana дикого типа». Журнал экспериментальной ботаники. 54 (380): 1–10. Дои:10.1093 / jxb / 54.380.1. PMID 12456750.
- ^ Гилл Б.С., Кимбер Г. (апрель 1974 г.). "Гимза С-полосный кариотип ржи". Труды Национальной академии наук Соединенных Штатов Америки. 71 (4): 1247–9. Bibcode:1974PNAS ... 71.1247G. Дои:10.1073 / пнас.71.4.1247. ЧВК 388202. PMID 4133848.
- ^ а б c Дубковски Дж., Ло М.К., Чжун Дж., Бранштайтер Р., Десаи А., Килиан А., Кляйнхофс А., Дворак Дж. (Июнь 1996 г.). «Генетическая карта диплоидной пшеницы Triticum monococcum L. и ее сравнение с картами Hordeum vulgare L». Генетика. 143 (2): 983–99. ЧВК 1207354. PMID 8725244.
- ^ Като А., Лэмб Дж. С., Берчлер Дж. А. (сентябрь 2004 г.). «Хромосомная окраска с использованием повторяющихся последовательностей ДНК в качестве зондов для идентификации соматических хромосом у кукурузы». Труды Национальной академии наук Соединенных Штатов Америки. 101 (37): 13554–9. Bibcode:2004ПНАС..10113554К. Дои:10.1073 / pnas.0403659101. ЧВК 518793. PMID 15342909.
- ^ Кентон А., Пароконный А.С., Глеба Ю.Ю., Беннетт М.Д. (август 1993 г.). «Характеристика генома Nicotiana tabacum L. методом молекулярной цитогенетики». Молекулярная и общая генетика. 240 (2): 159–69. Дои:10.1007 / BF00277053. PMID 8355650. S2CID 6953185.
- ^ Leitch IJ, Soltis DE, Солтис PS, Беннетт MD (январь 2005 г.). «Эволюция количества ДНК у наземных растений (эмбриофиты)». Анналы ботаники. 95 (1): 207–17. Дои:10.1093 / aob / mci014. ЧВК 4246719. PMID 15596468.
- ^ Ambarish, C.N .; Шридхар, К. (2014). "Цитологические и кариологические наблюдения двух эндемичных гигантских пилюль-многоножек. Артросфаэра (Pocock 1895) (Diplopoda: Sphaerotheriida) Западных Гат Индии ". Кариология. 67 (1): 49–56. Дои:10.1080/00087114.2014.891700.
- ^ Виттури Р., Коломба М.С., Пирроне А.М., Мандриоли М. (2002). "rDNA (18S–28S and 5S) colocalization and linkage between ribosomal genes and (TTAGGG)(n) telomeric sequence in the earthworm, Octodrilus complanatus (Annelida: Oligochaeta: Lumbricidae), revealed by single- and double-color FISH". The Journal of Heredity. 93 (4): 279–82. Дои:10.1093/jhered/93.4.279. PMID 12407215.
- ^ Nie W, Wang J, O'Brien PC, Fu B, Ying T, Ferguson-Smith MA, Yang F (2002). "The genome phylogeny of domestic cat, red panda and five mustelid species revealed by comparative chromosome painting and G-banding". Chromosome Research. 10 (3): 209–22. Дои:10.1023/A:1015292005631. PMID 12067210. S2CID 9660694.
- ^ а б Romanenko SA, Perelman PL, Serdukova NA, Trifonov VA, Biltueva LS, Wang J, Li T, Nie W, O'Brien PC, Volobouev VT, Stanyon R, Ferguson-Smith MA, Yang F, Graphodatsky AS (December 2006). "Reciprocal chromosome painting between three laboratory rodent species". Mammalian Genome. 17 (12): 1183–92. Дои:10.1007/s00335-006-0081-z. PMID 17143584. S2CID 41546146.
- ^ а б Painter TS (March 1928). "A Comparison of the Chromosomes of the Rat and Mouse with Reference to the Question of Chromosome Homology in Mammals". Генетика. 13 (2): 180–9. ЧВК 1200977. PMID 17246549.
- ^ Hayes H, Rogel-Gaillard C, Zijlstra C, De Haan NA, Urien C, Bourgeaux N, Bertaud M, Bosma AA (2002). "Establishment of an R-banded rabbit karyotype nomenclature by FISH localization of 23 chromosome-specific genes on both G- and R-banded chromosomes". Cytogenetic and Genome Research. 98 (2–3): 199–205. Дои:10.1159/000069807. PMID 12698004. S2CID 29849096.
- ^ "The Genetics of the Popular Aquarium Pet – Guppy Fish". Получено 6 декабря 2009.
- ^ а б De Grouchy J (August 1987). "Chromosome phylogenies of man, great apes, and Old World monkeys". Genetica. 73 (1–2): 37–52. Дои:10.1007/bf00057436. PMID 3333352. S2CID 1098866.
- ^ Robinson TJ, Yang F, Harrison WR (2002). "Chromosome painting refines the history of genome evolution in hares and rabbits (order Lagomorpha)". Cytogenetic and Genome Research. 96 (1–4): 223–7. Дои:10.1159/000063034. PMID 12438803. S2CID 19327437.
- ^ Chapman JA, Flux JE (1990), "section 4.W4", Rabbits, Hares and Pikas. Status Survey and Conservation Action Plan, pp. 61–94, ISBN 9782831700199
- ^ Vitturi R, Libertini A, Sineo L, Sparacio I, Lannino A, Gregorini A, Colomba M (2005). "Cytogenetics of the land snails Cantareus aspersus and C. mazzullii (Mollusca: Gastropoda: Pulmonata)". Micron. 36 (4): 351–7. Дои:10.1016/j.micron.2004.12.010. PMID 15857774.
- ^ Yasukochi Y, Ashakumary LA, Baba K, Yoshido A, Sahara K (July 2006). "A second-generation integrated map of the silkworm reveals synteny and conserved gene order between lepidopteran insects". Генетика. 173 (3): 1319–28. Дои:10.1534/genetics.106.055541. ЧВК 1526672. PMID 16547103.
- ^ Houck ML, Kumamoto AT, Gallagher DS, Benirschke K (2001). "Comparative cytogenetics of the African elephant (Loxodonta africana) and Asiatic elephant (Elephas maximus)". Cytogenetics and Cell Genetics. 93 (3–4): 249–52. Дои:10.1159/000056992. PMID 11528120. S2CID 23529399.
- ^ Semba U, Umeda Y, Shibuya Y, Okabe H, Tanase S, Yamamoto T (October 2004). "Primary structures of guinea pig high- and low-molecular-weight kininogens". International Immunopharmacology. 4 (10–11): 1391–400. Дои:10.1016/j.intimp.2004.06.003. PMID 15313436.
- ^ Wayne RK, Ostrander EA (March 1999). "Origin, genetic diversity, and genome structure of the domestic dog". BioEssays. 21 (3): 247–57. Дои:10.1002/(SICI)1521-1878(199903)21:3<247::AID-BIES9>3.0.CO;2-Z. PMID 10333734.
- ^ Ciudad J, Cid E, Velasco A, Lara JM, Aijón J, Orfao A (May 2002). "Flow cytometry measurement of the DNA contents of G0/G1 diploid cells from three different teleost fish species". Cytometry. 48 (1): 20–5. Дои:10.1002/cyto.10100. PMID 12116377.
- ^ Burt DW (2002). "Origin and evolution of avian microchromosomes". Cytogenetic and Genome Research. 96 (1–4): 97–112. Дои:10.1159/000063018. PMID 12438785. S2CID 26017998.
- ^ Itoh M, Ikeuchi T, Shimba H, Mori M, Sasaki M, Makino S (1969). "A Comparative Karyotype Study in Fourteen Species of Birds". The Japanese Journal of Genetics. 44 (3): 163–170. Дои:10.1266/jjg.44.163.
- ^ Smith J, Burt DW (August 1998). "Parameters of the chicken genome (Gallus gallus)". Animal Genetics. 29 (4): 290–4. Дои:10.1046/j.1365-2052.1998.00334.x. PMID 9745667.
- ^ Sakamura, Tetsu (1918). "Kurze Mitteilung über die Chromosomenzahlen und die Verwandtschaftsverhältnisse der Triticum-Arten". Shokubutsugaku Zasshi. 32 (379): 150–3. Дои:10.15281/jplantres1887.32.379_150.
- ^ Charlebois R.L. (ed) 1999. Organization of the prokaryote genome. ASM Press, Washington DC.
- ^ Komaki K, Ishikawa H (March 2000). "Genomic copy number of intracellular bacterial symbionts of aphids varies in response to developmental stage and morph of their host". Insect Biochemistry and Molecular Biology. 30 (3): 253–8. Дои:10.1016/S0965-1748(99)00125-3. PMID 10732993.
- ^ Mendell JE, Clements KD, Choat JH, Angert ER (May 2008). "Extreme polyploidy in a large bacterium". Proceedings of the National Academy of Sciences of the United States of America. 105 (18): 6730–4. Bibcode:2008PNAS..105.6730M. Дои:10.1073/pnas.0707522105. ЧВК 2373351. PMID 18445653.
- ^ White, M. J. D. (1973). The chromosomes (6-е изд.). London: Chapman and Hall, distributed by Halsted Press, New York. п. 28. ISBN 978-0-412-11930-9.
- ^ von Winiwarter H (1912). "Études sur la spermatogenèse humaine". Archives de Biologie. 27 (93): 147–9.
- ^ Painter TS (1922). "The spermatogenesis of man". Anat. Res. 23: 129.
- ^ Painter, Theophilus S. (April 1923). "Studies in mammalian spermatogenesis. II. The spermatogenesis of man". Journal of Experimental Zoology. 37 (3): 291–336. Дои:10.1002/jez.1400370303.
- ^ Tjio JH, Levan A (1956). "The chromosome number of man". Hereditas. 42 (1–2): 723–4. Дои:10.1111/j.1601-5223.1956.tb03010.x. HDL:10261/15776. PMID 345813.
- ^ Ford CE, Hamerton JL (November 1956). "The chromosomes of man". Природа. 178 (4541): 1020–3. Bibcode:1956Natur.178.1020F. Дои:10.1038/1781020a0. PMID 13378517. S2CID 4155320.
- ^ Hsu T.C. (1979) Human and mammalian cytogenetics: a historical perspective. Springer-Verlag, N.Y. ISBN 9780387903644 п. 10: "It's amazing that he [Painter] even came close!"
- ^ Santaguida S, Amon A (August 2015). "Short- and long-term effects of chromosome mis-segregation and aneuploidy" (PDF). Nature Reviews. Molecular Cell Biology. 16 (8): 473–85. Дои:10.1038/nrm4025. HDL:1721.1/117201. PMID 26204159. S2CID 205495880.
- ^ Miller KR (2000). "Chapter 9-3". Биология (5th ed.). Upper Saddle River, New Jersey: Prentice Hall. стр.194 –5. ISBN 978-0-13-436265-6.
- ^ "What is Trisomy 18?". Trisomy 18 Foundation. Получено 4 февраля 2017.
- ^ European Chromosome 11 Network[неудачная проверка ]
- ^ Templado C, Uroz L, Estop A (October 2013). "New insights on the origin and relevance of aneuploidy in human spermatozoa". Molecular Human Reproduction. 19 (10): 634–43. Дои:10.1093/molehr/gat039. PMID 23720770.
- ^ Shi Q, Ko E, Barclay L, Hoang T, Rademaker A, Martin R (August 2001). "Cigarette smoking and aneuploidy in human sperm". Molecular Reproduction and Development. 59 (4): 417–21. Дои:10.1002/mrd.1048. PMID 11468778. S2CID 35230655.
- ^ Rubes J, Lowe X, Moore D, Perreault S, Slott V, Evenson D, Selevan SG, Wyrobek AJ (October 1998). "Smoking cigarettes is associated with increased sperm disomy in teenage men". Fertility and Sterility. 70 (4): 715–23. Дои:10.1016/S0015-0282(98)00261-1. PMID 9797104.
- ^ Xing C, Marchetti F, Li G, Weldon RH, Kurtovich E, Young S, Schmid TE, Zhang L, Rappaport S, Waidyanatha S, Wyrobek AJ, Eskenazi B (June 2010). "Benzene exposure near the U.S. permissible limit is associated with sperm aneuploidy". Environmental Health Perspectives. 118 (6): 833–9. Дои:10.1289/ehp.0901531. ЧВК 2898861. PMID 20418200.
- ^ Xia Y, Bian Q, Xu L, Cheng S, Song L, Liu J, Wu W, Wang S, Wang X (October 2004). "Genotoxic effects on human spermatozoa among pesticide factory workers exposed to fenvalerate". Токсикология. 203 (1–3): 49–60. Дои:10.1016/j.tox.2004.05.018. PMID 15363581.
- ^ Xia Y, Cheng S, Bian Q, Xu L, Collins MD, Chang HC, Song L, Liu J, Wang S, Wang X (May 2005). "Genotoxic effects on spermatozoa of carbaryl-exposed workers". Toxicological Sciences. 85 (1): 615–23. Дои:10.1093/toxsci/kfi066. PMID 15615886.
- ^ Governini L, Guerranti C, De Leo V, Boschi L, Luddi A, Gori M, Orvieto R, Piomboni P (November 2015). "Chromosomal aneuploidies and DNA fragmentation of human spermatozoa from patients exposed to perfluorinated compounds". Andrologia. 47 (9): 1012–9. Дои:10.1111/and.12371. PMID 25382683. S2CID 13484513.
внешняя ссылка
- An Introduction to DNA and Chromosomes из HOPES: Huntington's Outreach Project for Education at Stanford
- Chromosome Abnormalities at AtlasGeneticsOncology
- On-line exhibition on chromosomes and genome (SIB)
- What Can Our Chromosomes Tell Us?, from the University of Utah's Genetic Science Learning Center
- Try making a karyotype yourself, from the University of Utah's Genetic Science Learning Center
- Kimballs Chromosome pages
- Chromosome News from Genome News Network
- Eurochromnet, European network for Rare Chromosome Disorders on the Internet
- Ensembl.org, Ансамбль project, presenting chromosomes, their гены и syntenic loci graphically via the web
- Genographic Project
- Home reference on Chromosomes from the U.S. National Library of Medicine
- Visualisation of human chromosomes and comparison to other species
- Unique – The Rare Chromosome Disorder Support Group Support for people with rare chromosome disorders