Эндогенный ретровирус - Википедия - Endogenous retrovirus
Эндогенные ретровирусы (ERV) находятся эндогенные вирусные элементы в геном которые очень похожи и могут быть получены из ретровирусы. Их много в геномах челюстные позвоночные, и они составляют до 5–8% генома человека (нижняя оценка ~ 1%).[1][2] ERV являются подклассом типа ген называется транспозон, который может быть упакован и перемещен внутри генома, чтобы играть жизненно важную роль в экспрессия гена И в регулирование.[3][4] Они выделяются как ретротранспозоны, которые являются элементами класса I.[5] Исследователи предположили, что ретровирусы произошли от типа мобильного гена, называемого ретротранспозоном, который включает ERV; эти гены могут мутировать и вместо перемещения в другое место в геноме они могут стать экзогенными или патогенными. Это означает, что не все ERV могли возникнуть в результате вставки ретровируса, но что некоторые из них могли быть источником генетической информации в ретровирусах, на которые они похожи.[6] Когда интеграция вирусной ДНК происходит в зародышевой линии, это может дать начало ERV, который позже может закрепиться в генофонде популяции хозяина.[1][7]
Формирование
Цикл репликации ретровирус влечет за собой вставку («интеграцию») копии ДНК вирусного геном в ядерный геном хозяин клетка. Большинство ретровирусов заражают соматические клетки, но случайное заражение зародышевый клетки (клетки, производящие яйцеклетки и сперму) также могут возникать. В редких случаях интеграция ретровируса может происходить в клетке зародышевой линии, которая продолжает развиваться в жизнеспособный организм. Этот организм будет нести встроенный ретровирусный геном как неотъемлемую часть своего собственного генома - «эндогенный» ретровирус (ERV), который может быть унаследованный своим потомство как роман аллель. Многие ERV сохраняются в геноме своих хозяев миллионы лет. Однако большинство из них приобрели инактивирующий мутации во время хоста Репликация ДНК и больше не способны производить вирус. ERV также могут быть частично вырезаны из генома с помощью процесса, известного как рекомбинационная делеция, при котором рекомбинация между идентичными последовательностями, которые фланкируют вновь интегрированные ретровирусы, приводит к удалению внутренних, белок -кодирующие области вирусного генома.
Общий геном ретровируса состоит из трех генов, жизненно важных для вторжения, репликации, ускользания и распространения его вирусного генома. Эти три гена: gag (кодирует структурные белки ядра вируса), pol (кодирует обратная транскриптаза, интегрировать, и протеаза ) и env (кодирует для белки оболочки для внешнего вида вируса). Эти вирусные белки кодируются как полипротеины. Для осуществления своего жизненного цикла ретровирус в значительной степени полагается на механизмы клетки-хозяина. Протеаза разрушает пептидные связи вирусных полипротеинов, делая отдельные белки функциональными. Функция обратной транскриптазы заключается в синтезе вирусной ДНК из вирусной РНК в цитоплазме клетки-хозяина до того, как она попадет в ядро. Интеграция направляет интеграцию вирусной ДНК в геном хозяина.[7][8]
Со временем геном ERV не только приобретает точечные мутации, но также перетасовывается и рекомбинируется с другими ERV.[9] ERV с env сгнившая шерсть с большей вероятностью распространится.[10]
Роль в эволюции генома

Эндогенные ретровирусы могут играть активную роль в формировании геномов. Большинство исследований в этой области было сосредоточено на геномах людей и высших приматов, но другие позвоночные, такие как мыши и овцы, также были глубоко изучены.[11][12][13][14] Длинный терминальный повтор (LTR ) последовательности, фланкирующие геномы ERV, часто действуют как альтернативные промоутеры и усилители, часто способствуя транскриптом путем создания тканеспецифичных вариантов. Кроме того, ретровирусный белки сами были адаптированы для выполнения новых функций хозяина, особенно в репродукции и развитии. Рекомбинация между гомологичными ретровирусными последовательностями также способствовала перетасовке генов и генерации генетических вариаций. Более того, в случае потенциально антагонистических эффектов ретровирусных последовательностей, репрессорные гены эволюционировали совместно, чтобы бороться с ними.
Около 90% эндогенных ретровирусов являются одиночными LTR, у которых отсутствуют все открытые рамки для чтения (ORF).[15] Было показано, что одиночные LTR и LTR, связанные с полными ретровирусными последовательностями, действуют как транскрипционные элементы на генах хозяина. Их диапазон действия в основном заключается в установке в 5 ' UTR генов, кодирующих белок; однако известно, что они действуют на гены до 70–100kb прочь.[11][16][17][18] Большинство этих элементов вставлены в смысловом направлении в соответствующие им гены, но были доказательства того, что LTR действуют в антисмысловом направлении и как двунаправленный промотор для соседних генов.[19][20] В некоторых случаях LTR функционирует как главный промотор гена. Например, у человека AMY1C имеет полную последовательность ERV в своей промоторной области; связанный LTR обеспечивает специфическую экспрессию пищеварительной системы в слюне. фермент амилаза.[21] Кроме того, основной промоутер для желчная кислота-КоА: N-ацилтрансфераза аминокислоты (BAAT), который кодирует фермент, участвующий в метаболизме желчи, имеет LTR-происхождение.[17][22]
Включение одиночного ERV-9 LTR могло привести к появлению функциональной открытой рамки считывания, что привело к возрождению человека. ген GTPase, связанный с иммунитетом (КСИР).[23] Также было показано, что вставки ERV генерируют альтернативные сайты сплайсинга либо путем прямой интеграции в ген, как в случае рецептора человеческого лептинового гормона, либо за счет экспрессии расположенного выше LTR, как в случае белка, подобного фосфолипазе A-2.[24]
Однако большую часть времени LTR функционирует как один из многих альтернативных промоторов, часто обеспечивая тканеспецифическую экспрессию, связанную с воспроизводством и развитием. Фактически, 64% известных вариантов транскрипции, стимулированных LTR, являются выразил в репродуктивных тканях.[25] Например, ген CYP19 кодирует ароматаза P450, важный фермент для синтеза эстрогена, который обычно экспрессируется в головном мозге и репродуктивных органах большинства млекопитающих.[17] Однако у приматов вариант транскрипции, обеспечиваемый LTR, передает экспрессию плаценте и отвечает за контроль уровня эстрогена во время беременности.[17] Кроме того, белок, ингибирующий апоптоз нейронов (NAIP), обычно широко распространенный, имеет LTR семейства HERV-P, действующий как промотор, придающий экспрессию семенникам и простате.[26] Другие белки, такие как синтаза оксида азота 3 (NOS3), рецептор интерлейкина-2 B (IL2RB) и другой медиатор синтеза эстрогена, HSD17B1, также альтернативно регулируются LTR, которые обеспечивают экспрессию плаценты, но их конкретные функции еще не известны. .[22][27] Считается, что высокая степень репродуктивной экспрессии является последствием метода, которым они были эндогенизированы; однако это также может быть связано с отсутствием метилирования ДНК в тканях зародышевой линии.[22]
Наиболее хорошо охарактеризованный пример экспрессии плацентарного белка происходит не от альтернативно промотированного гена-хозяина, а от полной ко-опции ретровирусного белка. Ретровирусные слитные белки env, которые играют роль в проникновении вириона в клетку-хозяина, оказали важное влияние на развитие млекопитающих. плацента. У млекопитающих интактные белки env, называемые синцитины несут ответственность за формирование и функционирование синцитиотрофобласты.[13] Эти многоядерные клетки в основном отвечают за поддержание обмена питательными веществами и отделение плода от иммунной системы матери.[13] Было высказано предположение, что отбор и фиксация этих белков для этой функции сыграли решающую роль в эволюции живородство.[28]
Кроме того, вставка ERV и их соответствующих LTR может вызвать хромосомную перестройку из-за рекомбинации между вирусными последовательностями в межхромосомных локусах. Было показано, что эти перестройки вызывают дупликации и делеции генов, которые в значительной степени способствуют пластичности генома и резко изменяют динамику функции генов.[29] Более того, ретроэлементы в целом широко распространены в быстро развивающихся семействах генов, специфичных для млекопитающих, функция которых в значительной степени связана с реакцией на стресс и внешние раздражители.[17] В частности, как человеческие класс I и класс II MHC гены имеют высокую плотность элементов HERV по сравнению с другими семействами мультилокусных генов.[24] Было показано, что HERV вносят вклад в формирование сильно дублированных дупликонных блоков, которые составляют семейство генов HLA класса 1.[30] Более конкретно, HERV в основном занимают регионы внутри и между точками разрыва между этими блоками, указывая тем самым, что значительные события дупликации и делеций, обычно связанные с неравным кроссовером, способствовали их образованию.[31] Генерация этих блоков, унаследованных как иммуно-гаплотипы, действует как защитный полиморфизм против широкого спектра антигенов, которые, возможно, придавали людям преимущество перед другими приматами.[30]
Характеристика плаценты очень эволюционно отдельные органы между разными видами, как предполагалось, являются результатом коопция энхансеров ERV. Регуляторные мутации вместо мутаций в генах, кодирующих гормоны и факторы роста, поддерживают известную эволюцию морфологии плаценты, тем более что большинство генов гормонов и факторов роста экспрессируются в ответ на беременность, а не во время развития плаценты. Исследователи изучили регуляторный ландшафт развития плаценты у крыс и мышей, двух близкородственных видов. Это было сделано путем картирования всех регуляторных элементов крысы. трофобласт стволовые клетки (TSC) и сравнивая их с их ортологи в TSC мыши. TSC наблюдались, потому что они отражают исходные клетки, которые развиваются в плаценте плода. Несмотря на их ощутимое сходство, энхансерные и репрессированные области были в основном видоспецифичными. Однако большинство промоторных последовательностей у мыши и крысы было консервативным. В заключение своего исследования исследователи предположили, что ERV влияли на видоспецифичную эволюцию плаценты через посредничество роста плаценты, иммуносупрессия, и слияние клеток.[32]
Другим примером использования клеточных механизмов ERV является p53, а ген-супрессор опухоли (TSG). Повреждение ДНК и клеточный стресс индуцируют путь p53, что приводит к апоптоз. Используя иммунопреципитацию хроматина с секвенированием, тридцать процентов всех сайтов связывания p53 были локализованы в копиях нескольких семейств ERV, специфичных для приматов. Исследование показало, что это приносит пользу ретровирусам, потому что механизм p53 обеспечивает быструю индукцию транскрипции, которая приводит к выходу вирусной РНК из клетки-хозяина.[5]
Наконец, встраивание ERV или элементов ERV в генные области ДНК хозяина или сверхэкспрессия их транскрипционных вариантов имеет гораздо более высокий потенциал для получения вредных эффектов, чем положительные. Их появление в геноме создало коэволюционная динамика паразита и хозяина это увеличивало дупликацию и экспансию репрессорных генов. Самый наглядный пример этого - быстрое дублирование и распространение тандемных цинковый палец гены в геномах млекопитающих. Гены цинковых пальцев, особенно те, которые включают домен KRAB, существуют в большом количестве копий в геномах позвоночных, и их диапазон функций ограничен транскрипционными ролями.[33] Однако на млекопитающих было показано, что диверсификация этих генов происходит из-за множественных событий дупликации и фиксации в ответ на новые ретровирусные последовательности или их эндогенные копии, подавляющие их транскрипцию.[18]
Роль в болезни
Большинство ERV, которые встречаются в геномах позвоночных, являются древними, инактивированными мутациями и достигли генетического фиксация в их хозяевах. По этим причинам крайне маловероятно, что они окажут негативное влияние на своих хозяев, за исключением необычных обстоятельств. Тем не менее, это ясно из исследований на птицах и других млекопитающих, включая мышей, кошек и коалы, что более молодые (т.е. недавно интегрированные) ERV могут быть связаны с заболеванием. Количество активных ERV в геноме млекопитающих отрицательно связано с размером их тела, что предполагает вклад в Парадокс пето через патогенез рака.[34] Это побудило исследователей предположить роль ERV в нескольких формах рака человека и аутоиммунное заболевание, хотя убедительных доказательств нет.[35][36][37][38]
Неврологические расстройства
Предполагается, что у людей ERV вовлечены в рассеянный склероз (РС). Специфическая связь между РС и ERVWE1 Сообщалось о гене «синцитина», который происходит из вставки ERV, наряду с присутствием «MS-ассоциированного ретровируса» (MSRV) у пациентов с этим заболеванием.[39][40] ERV человека (HERV) также участвуют в ALS[41] и зависимость.[42][43][44]
В 2004 году сообщалось, что антитела к HERV чаще обнаруживались в сыворотка людей с шизофрения. Кроме того, спинномозговая жидкость людей с недавно начавшейся шизофренией содержали уровни ретровирусного маркера, обратная транскриптаза, в четыре раза выше, чем у контрольных испытуемых.[45] Исследователи продолжают изучать возможную связь между HERV и шизофренией с дополнительной возможностью инициирования инфекция, вызывающая шизофрению.[46]
Иммунитет
Было обнаружено, что ERV связаны с заболеванием не только через отношения, вызывающие болезнь, но и через иммунитет. Частота ERV в длинных концевых повторах (LTR), вероятно, коррелирует с вирусной адаптацией, чтобы использовать сигнальные пути иммунитета, которые способствуют транскрипции и репликации вируса. В исследовании, проведенном в 2016 году, изучалась польза древней вирусной ДНК, интегрированной в хозяина через сети регуляции генов, вызванные интерфероны, ветвь врожденного иммунитета.[47] Эти цитокины первыми реагируют на вирусную инфекцию, а также важны для иммунного надзора за злокачественными клетками.[48] Предполагается, что ERV действуют как цис-регуляторные элементы, но большая часть адаптивных последствий этого для определенных физиологических функций все еще неизвестна. Существуют данные, подтверждающие общую роль ERV в регуляции ответа на интерферон человека, в частности, на интерферон-гамма (IFNG). Например, было обнаружено, что гены, стимулированные интерфероном, в значительной степени обогащены ERV, связанными сигнальным преобразователем и активатором транскрипции (STAT1) и / или Фактор регуляции интерферона (IRF1) в CD14 + макрофаги.[1]
HERV также играют различные роли, формируя человеческий врожденный иммунитет ответ, причем одни последовательности активируют систему, а другие подавляют. Они также могут защищать от экзогенных ретровирусных инфекций: вирусоподобные транскрипты могут активировать рецепторы распознавания образов, и белки могут мешать активным ретровирусам. Показано, что белок gag из HERV-K (HML2) смешивается с Gag ВИЧ, в результате нарушая образование капсида ВИЧ.[49]
Генная регуляция
Другая предложенная идея заключалась в том, что ERV из одного семейства играют роль в рекрутировании нескольких генов в одну и ту же сеть регуляции. Было обнаружено, что элементы MER41 обеспечивают дополнительное избыточное регуляторное усиление генов, расположенных рядом с сайтами связывания STAT1.[1]
Роль в медицине
Эндогенный ретровирус свиней
Для людей, свинья эндогенные ретровирусы (PERV) представляют проблему при использовании тканей и органов свиньи при ксенотрансплантации, трансплантации живых клеток, тканей и органов из организма одного вида в организм другого вида. Хотя свиньи, как правило, являются наиболее подходящими донорами для лечения заболеваний органов человека по практическим, финансовым, безопасным и этическим причинам,[47] PERV ранее не могли быть удалены из свиней из-за его вирусной природы интеграции в геном хозяина и передачи потомству до 2017 года, когда лаборатория доктора Джорджа Черча удалила все 62 ретровируса из генома свиней.[50] Последствия межвидовой передачи остаются неизученными и имеют очень опасный потенциал.[51]
Исследователи указали, что инфицирование тканей человека PERV очень возможно, особенно у людей с ослабленным иммунитетом. Состояние с подавленным иммунитетом потенциально может позволить более быструю и стойкую репликацию вирусной ДНК, и впоследствии будет меньше проблем с адаптацией к передаче от человека к человеку. Хотя известные инфекционные патогены, присутствующие в донорском органе / ткани, могут быть устранены путем разведения стада, свободного от патогенов, у донора могут присутствовать неизвестные ретровирусы. Эти ретровирусы часто латентны и бессимптомны у донора, но могут стать активными у реципиента. Некоторые примеры эндогенных вирусов, которые могут инфицировать и размножаться в клетках человека, происходят от павианов (BaEV), кошек (RD114) и мышей.[47]
Существует три разных класса PERV: PERV-A, PERV-B и PERV-C. PERV-A и PERV-B являются политропный и может инфицировать клетки человека in vitro, тогда как PERV-C экотропный и не реплицируется на человеческих клетках. Основные различия между классами заключаются в домене связывания рецептора env белок и длинные концевые повторы (LTR), которые влияют на репликацию каждого класса. PERV-A и PERV-B отображают LTR, которые имеют повторы в U3 область, край. Однако PERV-A и PERV-C показывают повторяющиеся LTR. Исследователи обнаружили, что PERV в культуре активно адаптировались к повторяющейся структуре своих LTR, чтобы соответствовать наилучшим характеристикам репликации, которые могла выполнять клетка-хозяин. В конце своего исследования исследователи пришли к выводу, что повторяющийся PERV LTR произошел от повторяющегося LTR. Вероятно, это произошло из-за инсерционной мутации и было доказано с использованием данных о LTR и env/ Env. Считается, что генерация повторяющихся LTR может быть отражением процесса адаптации вируса, переходящего от экзогенного к эндогенному образу жизни.[52]
В ходе клинического исследования, проведенного в 1999 году, были отобраны 160 пациентов, которым вводили различные ткани живых свиней, и не было обнаружено никаких доказательств стойкой инфекции PERV у 97% пациентов, у которых для ПЦР было доступно достаточное количество ДНК для амплификации последовательностей PERV. Однако в этом исследовании говорится, что ретроспективные исследования ограничены выявлением истинной частоты инфекции или связанных клинических симптомов. Он предложил использовать тщательно контролируемые проспективные исследования, которые предоставят более полную и подробную оценку возможной межвидовой передачи PERV и сравнение PERV.[53]
Эндогенные ретровирусы человека
Эндогенные ретровирусы человека (HERV) составляют значительную часть человеческий геном, примерно 98 000 элементов и фрагментов ERV составляют 5–8%.[1] Согласно исследованию, опубликованному в 2005 году, не было обнаружено HERV, способных к репликации; все оказались дефектными, содержали серьезные делеции или бессмысленные мутации. Это потому, что большинство HERV - это просто следы исходных вирусов, впервые интегрированных миллионы лет назад. Анализ интеграции HERV продолжается в рамках Проект 100000 геномов.[54]
Эндогенные ретровирусы человека были обнаружены случайно в нескольких различных экспериментах. Геномные библиотеки человека были проверены в условиях низкой строгости с использованием зондов из ретровирусов животных, что позволило выделить и охарактеризовать несколько, хотя и дефектных, провирусов, которые представляли различные семейства. Другой эксперимент зависел от олигонуклеотидов, гомологичных сайтам связывания вирусных праймеров.[1]
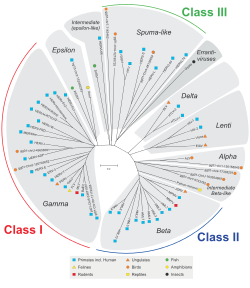
HERV классифицируются на основе их гомологии с ретровирусами животных. Семьи, принадлежащие к классу I, сходны по последовательности с млекопитающими. Гаммаретровирусы (тип C) и Эпсилонретровирусы (Тип E). Семьи, принадлежащие ко второму классу, гомологичны млекопитающим. Бетаретровирусы (Тип B) и Дельтаретровирусы (Тип D). Семьи, относящиеся к III классу, похожи на пенистые вирусы. Для всех классов, если гомологии хорошо сохраняются в кляп, pol, и env ген, они сгруппированы в надсемейство. Известно, что существует больше семейств класса I.[1][9] Сами семейства названы менее единообразно, со смесью наименований на основе экзогенного ретровируса, праймирующей тРНК (HERV-W, K) или некоторого соседнего гена (HERV-ADP), номера клонга (HERV-S71). или какой-либо аминокислотный мотив (HERV-FRD). Предлагаемая номенклатура призвана очистить иногда парафилетические стандарты.[55]
Есть два предположения о том, как HERV закрепились в геноме человека. Первый предполагает, что когда-то в процессе эволюции человека экзогенные предшественники HERV встраивались в клетки зародышевой линии, а затем реплицировались вместе с генами хозяина, используя и эксплуатируя клеточные механизмы хозяина. Из-за своей особой геномной структуры HERV подвергались множеству раундов амплификации и транспозиции, что привело к широкому распространению ретровирусной ДНК. Вторая гипотеза предполагает непрерывную эволюцию ретро-элементов от более простых по структуре предков.[1]
Тем не менее, одно семейство вирусов было активным с момента расхождения люди и шимпанзе. Эта семья, называемая HERV-K (HML2), составляет менее 1% элементов HERV, но является одним из наиболее изученных. Есть признаки того, что он был активен в последние несколько сотен тысяч лет, например, некоторые люди несут больше копий HML2, чем другие.[56] Традиционно оценка возраста HERV выполняется путем сравнения 5 'и 3' LTR HERV; однако этот метод актуален только для полноразмерных HERV. Недавний метод, называемый кросс-секционным датированием,[57] использует вариации в пределах одного LTR для оценки возраста вставок HERV. Этот метод более точен при оценке возраста HERV и может использоваться для любых вставок HERV. Поперечное датирование было использовано, чтобы предположить, что два члена HERV-K (HML2), HERV-K106 и HERV-K116, были активны в течение последних 800000 лет и что HERV-K106, возможно, заразил современных людей 150 000 лет назад.[58] Однако отсутствие известных инфекционных членов семейства HERV-K (HML2) и отсутствие элементов с полным кодирующим потенциалом в опубликованной последовательности генома человека позволяют некоторым предположить, что это семейство в настоящее время менее вероятно. В 2006 и 2007 годах исследователи, работавшие независимо во Франции и США, воссоздали функциональные версии HERV-K (HML2).[59][60]
MER41.AIM2 - это HERV, который регулирует транскрипцию AIM2 (отсутствует в меланоме 2), который кодирует датчик чужеродной цитозольной ДНК. Он действует как сайт связывания для AIM2, что означает, что он необходим для транскрипции AIM2. Исследователи продемонстрировали это, удалив MER41.AIM2 в клетках HeLa с использованием CRISPR / Cas9, что привело к неопределяемому уровню транскрипта AIM2 в модифицированных клетках HeLa. Контрольные клетки, которые все еще содержали ERV MER41.AIM2, наблюдались с нормальными количествами транскрипта AIM2. Что касается иммунитета, исследователи пришли к выводу, что MER41.AIM2 необходим для воспалительного ответа на инфекцию.[61]
Иммунологические исследования показали некоторые доказательства того, что Т-клетка иммунные ответы против HERV у ВИЧ-инфицированных.[62] Гипотеза о том, что ВИЧ индуцирует экспрессию HERV в ВИЧ-инфицированных клетках, привела к предположению, что вакцина, нацеленная на антигены HERV, может специфически уничтожать ВИЧ-инфицированные клетки. Потенциальное преимущество этого нового подхода заключается в том, что, используя антигены HERV в качестве суррогатных маркеров ВИЧ-инфицированных клеток, можно обойти трудности, связанные с прямым нацеливанием на заведомо разнообразные и быстро мутирующие антигены ВИЧ.[62]
Есть несколько классов эндогенных ретровирусов человека, которые все еще имеют неповрежденные открытые рамки считывания. Например, экспрессия HERV-K, биологически активного семейства HERV, продуцирует белки, обнаруженные в плаценте. Кроме того, экспрессия генов оболочки HERV-W (ERVW-1 )и HERV-FRD (ERVFRD-1 ) продуцирует синцитины, которые важны для образования слоя клеток синцитиотрофобласта во время плацентогенеза, индуцируя слияние клеток.[63] В Комитет по номенклатуре генов HUGO (HGNC) утверждает символы генов для транскрибированных человеческих ERV.[64]
Методы характеристики ERV
Секвенирование всего генома
Пример: свиной ERV (PERV) китайского происхождения. мини-свинья Изолят PERV-A-BM был полностью секвенирован вместе с различными породами и линиями клеток, чтобы понять его генетические вариации и эволюцию. Наблюдаемое количество нуклеотидных замен и среди различных последовательностей генома помогло исследователям определить приблизительный возраст, в котором PERV-A-BM был интегрирован в его геном-хозяин, который, как было установлено, имеет эволюционный возраст раньше, чем изоляты свиней европейского происхождения.[51]
Иммунопреципитация хроматина с секвенированием (ChIP-seq)
Этот метод используется для поиска гистоновых меток, указывающих на промоторы и энхансеры, которые являются сайтами связывания для белков ДНК, а также для репрессированных областей и триметилирования.[32] Было показано, что метилирование ДНК жизненно важно для поддержания сайленсинга ERV в соматических клетках мыши, в то время как гистоновые метки жизненно важны для той же цели в эмбриональные стволовые клетки (ЭСК) и ранний эмбриогенез.[5]
Приложения
Построение филогении
Поскольку большинство HERV не имеют функции, избирательно нейтральны и очень распространены в геномах приматов, они легко служат филогенетическими маркерами для анализа сцепления. Их можно использовать путем сравнения полиморфизмов сайтов интеграции или развивающихся провирусных нуклеотидных последовательностей ортологов. Чтобы оценить, когда произошла интеграция, исследователи использовали расстояния от каждого филогенетического дерева, чтобы определить скорость молекулярной эволюции в каждом конкретном локусе. Также полезно, что ERV богаты геномами многих видов (например, растений, насекомых, моллюсков, рыб, грызунов, домашних животных и домашнего скота), потому что их применение можно использовать для ответа на множество филогенетических вопросов.[7]
Определение возраста провируса и временные точки разделения видов
Это достигается путем сравнения различных HERV из разных эволюционных периодов. Например, это исследование проводилось для различных гоминоидов, от людей до обезьян и обезьян. Это сложно сделать с PERV из-за большого разнообразия.[52]
Дальнейшие исследования
Эпигенетическая изменчивость
Исследователи могли анализировать отдельные эпигеномы и транскриптомы изучить реактивацию бездействующих мобильных элементов посредством эпигенетического высвобождения и их потенциальную связь с заболеваниями человека, а также изучить особенности регуляторных сетей генов.[5]
Иммунологические проблемы ксенотрансплантации
Мало что известно об эффективном способе преодоления острейшее отторжение (HAR), который следует за активацией комплемента, инициированной ксенореактивными антителами, распознающими галактозил-альфа1-3галатозил (альфа-Gal) антигены на донорском эпителии.[47]
Факторы риска HERV в генной терапии
Поскольку ретровирусы способны рекомбинировать друг с другом и с другими эндогенными последовательностями ДНК, для генной терапии было бы полезно изучить потенциальные риски, которые могут вызвать HERV, если таковые имеются. Кроме того, этой способностью HERV рекомбинировать можно манипулировать для сайт-направленной интеграции путем включения последовательностей HERV в ретровирусные векторы.[1]
Экспрессия гена HERV
Исследователи считают, что РНК и белки, кодируемые генами HERV, следует продолжать изучать на предмет предполагаемой функции в физиологии клетки и при патологических состояниях. Это имело бы смысл изучить, чтобы глубже определить биологическое значение синтезированных белков.[1]
Смотрите также
- Вирус лейкоза саркомы птиц (ASLV)
- Эндогенный вирусный элемент
- ERV3
- HERV-FRD
- Горизонтальный перенос генов
- Ретровирус овец Jaagsiekte (JSRV)
- Коала ретровирус (КоРВ)
- Вирус опухоли молочной железы мыши (ММТВ)
- Вирус лейкемии мышей (MLV) и вирус, связанный с ксенотропным вирусом мышиного лейкоза (XMRV)
- Палеовирология
Рекомендации
- ^ а б c d е ж грамм час я j Белшоу Р., Перейра В., Кацуракис А., Талбот Г., Пейс Дж., Берт А., Тристем М. (апрель 2004 г.). «Длительная реинфекция генома человека эндогенными ретровирусами». Proc Natl Acad Sci USA. 101 (14): 4894–99. Bibcode:2004ПНАС..101.4894Б. Дои:10.1073 / pnas.0307800101. ЧВК 387345. PMID 15044706.
- ^ Нельсон П.Н., Хули П., Роден Д., Давари Эйтехади Х., Райланс П., Уоррен П. и др. (Октябрь 2004 г.). «Эндогенные ретровирусы человека: мобильные элементы с потенциалом?». Клиническая и экспериментальная иммунология. 138 (1): 1–9. Дои:10.1111 / j.1365-2249.2004.02592.x. ЧВК 1809191. PMID 15373898.
- ^ Ходосевич К., Лебедев Ю., Свердлов Е. (октябрь 2002 г.). «Эндогенные ретровирусы и эволюция человека». Сравнительная и функциональная геномика. 3 (6): 494–98. Дои:10.1002 / cfg.216. ЧВК 2448423. PMID 18629260.
- ^ Ким Ф.Дж., Баттини Дж.Л., Манель Н., Ситбон М. (январь 2004 г.). «Появление ретровирусов позвоночных и захват оболочки». Вирусология. 318 (1): 183–91. Дои:10.1016 / j.virol.2003.09.026. PMID 14972546.
- ^ а б c d Реболло Р., Романский MT, Магер DL (01.01.2012). «Мобильные элементы: богатый и естественный источник регуляторных последовательностей для генов хозяина». Ежегодный обзор генетики. 46 (1): 21–42. Дои:10.1146 / annurev-genet-110711-155621. PMID 22905872.
- ^ Коттон, Дж. (2001). «Ретровирусы из ретротранспозонов». Геномная биология. 2 (2): 6. Дои:10.1186 / gb-2001-2-2-reports0006.
Похоже, что переход от невирусного ретротранспозона к ретровирусу происходил независимо по крайней мере восемь раз, и источник гена оболочки, ответственного за инфекционную способность, теперь можно отнести к вирусу, по крайней мере, в четырех из этих случаев. Это говорит о том, что потенциально любой ретротранспозон LTR может стать вирусом за счет приобретения существующих вирусных генов.
- ^ а б c Джонсон В.Е., Гроб Дж. М. (август 1999 г.). «Построение филогении приматов из последовательностей древних ретровирусов». Труды Национальной академии наук Соединенных Штатов Америки. 96 (18): 10254–60. Bibcode:1999PNAS ... 9610254J. Дои:10.1073 / пнас.96.18.10254. ЧВК 17875. PMID 10468595.
- ^ Фудзивара Т., Мидзуучи К. (август 1988 г.). «Интеграция ретровирусной ДНК: структура интермедиата интеграции». Клетка. 54 (4): 497–504. Дои:10.1016/0092-8674(88)90071-2. PMID 3401925.
- ^ а б Vargiu, L; Rodriguez-Tomé, P; Спербер, GO; Кадедду, М; Гранди, N; Бликстад, В; Tramontano, E; Бломберг, Дж (22 января 2016 г.). «Классификация и характеристика эндогенных ретровирусов человека; часто встречаются мозаичные формы». Ретровирология. 13: 7. Дои:10.1186 / s12977-015-0232-у. ЧВК 4724089. PMID 26800882.
- ^ Magiorkinis, G; Гиффорд, RJ; Кацуракис, А; Де Рантер, Дж; Белшоу, Р. (8 мая 2012 г.). «Эндогенные ретровирусы без Env - суперраспространители генома». Труды Национальной академии наук Соединенных Штатов Америки. 109 (19): 7385–90. Дои:10.1073 / pnas.1200913109. ЧВК 3358877. PMID 22529376.
- ^ а б Ли Дж., Акаги К., Ху Й., Триветт А.Л., Хлынялюк С.Дж., Свинг Д.А., Вольфовский Н., Морган Т.С., Голубева Ю., Стивенс Р.М., Смит Д.Е., Саймер Д.Е. (март 2012 г.). «Эндогенные ретровирусы мышей могут вызывать преждевременное прекращение транскрипции на расстоянии». Genome Res. 22 (5): 870–84. Дои:10.1101 / гр.130740.111. ЧВК 3337433. PMID 22367191.
- ^ Спенсер Т.Э., Пальмарини М. (2012). «Эндогенные ретровирусы овец: модельная система для понимания физиологической адаптации к эволюционирующему геному жвачных животных». Джей Репрод Дев. 58 (1): 33–37. Дои:10.1262 / jrd.2011-026. PMID 22450282.
- ^ а б c Блэк С.Г., Арно Ф, Пальмарини М, Спенсер Т.Е. (2010). «Эндогенные ретровирусы в дифференцировке трофобластов и развитии плаценты». Американский журнал репродуктивной иммунологии. 64 (4): 255–64. Дои:10.1111 / j.1600-0897.2010.00860.x. ЧВК 4198168. PMID 20528833.
- ^ Райан Ф.П. (декабрь 2004 г.). «Эндогенные ретровирусы человека в состоянии здоровья и болезни: симбиотическая перспектива». Журнал Королевского медицинского общества. 97 (12): 560–65. Дои:10.1258 / jrsm.97.12.560. ЧВК 1079666. PMID 15574851.
- ^ Ishak, Charles A .; Де Карвалью, Даниэль Д. (2020). «Реактивация эндогенных ретроэлементов в развитии и терапии рака». Ежегодный обзор биологии рака. 4: 159–176. Дои:10.1146 / annurev-Cancebio-030419-033525.
- ^ Пи В., Чжу X, Ву М., Ван И, Фулзеле С., Эроглу А., Лин Дж., Туан Д. (июль 2010 г.). «Дальнодействующая функция межгенного ретротранспозона». PNAS. 107 (29): 12992–97. Bibcode:2010PNAS..10712992P. Дои:10.1073 / pnas.1004139107. ЧВК 2919959. PMID 20615953.
- ^ а б c d е van de Lagemaat LN, Landry JR, Mager DL, Medstrand P (октябрь 2003 г.). «Мобильные элементы у млекопитающих способствуют регуляторной изменчивости и диверсификации генов со специализированными функциями». Тенденции Genet. 19 (10): 530–36. Дои:10.1016 / j.tig.2003.08.004. PMID 14550626.
- ^ а б Ковальская Е., Буздин А., Гогвадзе Е., Виноградова Т., Свердлов Е. (2006). «Функциональные сайты начала транскрипции эндогенного ретровируса LTR человека расположены между областями R и U5». Вирусология. 346 (2): 373–78. Дои:10.1016 / j.virol.2005.11.007. PMID 16337666.
- ^ Dunn CA, Romanish MT, Gutierrez LE, van de Lagemaat LN, Mager DL (2006). «Транскрипция двух генов человека с двунаправленного промотора эндогенного ретровируса». Ген. 366 (2): 335–42. Дои:10.1016 / j.gene.2005.09.003. PMID 16288839.
- ^ Гогвадзе Э., Стукачева Э., Буздин А, Свердлов Э. (2009). «Специфическая для человека модуляция транскрипционной активности, обеспечиваемая эндогенными ретровирусными вставками». J Virol. 83 (12): 6098–105. Дои:10.1128 / JVI.00123-09. ЧВК 2687385. PMID 19339349.
- ^ Тинг К.Н., Розенберг М.П., Сноу К.М., Самуэльсон Л.С., Мейслер М.Х. (1992). «Эндогенные ретровирусные последовательности необходимы для тканеспецифической экспрессии гена амилазы слюны человека». Genes Dev. 6 (8): 1457–65. Дои:10.1101 / gad.6.8.1457. PMID 1379564.
- ^ а б c Коэн CJ, Лок WM, Mager DL (2009). «Эндогенные ретровирусные LTR как промоторы генов человека: критическая оценка». Ген. 448 (2): 105–14. Дои:10.1016 / j.gene.2009.06.020. PMID 19577618.
- ^ Бекпен С., Маркес-Бонет Т., Алкан С., Антоначчи Ф., Леогранде М.Б., Вентура М., Кидд Дж. М., Сисвара П., Ховард Дж. К., Эйхлер Э. Э. (2009). «Смерть и воскрешение человеческого гена IRGM». PLOS Genet. 5 (3): e1000403. Дои:10.1371 / journal.pgen.1000403. ЧВК 2644816. PMID 19266026.
- ^ а б Джерн П., Гроб Дж. М. (2008). «Влияние ретровирусов на функцию генома хозяина». Анну Рев Жене. 42: 709–32. Дои:10.1146 / annurev.genet.42.110807.091501. PMID 18694346.
- ^ Оливер KR, Грин WK (2011). «Мобильная ДНК и гипотеза TE-Thrust: подтверждающие данные от приматов». ДНК мафии. 2 (1): 8. Дои:10.1186/1759-8753-2-8. ЧВК 3123540. PMID 21627776.
- ^ Romanish MT, Lock WM, van de Lagemaat LN, Dunn CA, Mager DL (2007). «Повторное привлечение ретротранспозонов LTR в качестве промоторов антиапоптотическим локусом NAIP во время эволюции млекопитающих». PLOS Genet. 3 (1): e10. Дои:10.1371 / journal.pgen.0030010. ЧВК 1781489. PMID 17222062.
- ^ Ха JW, Ха HS, Ким Д.С., Ким HS (2008). «Ограниченная плацентой экспрессия NOS3, производного от LTR». Плацента. 29 (7): 602–08. Дои:10.1016 / j.placenta.2008.04.002. PMID 18474398.
- ^ Вильярреал LP, Вильярреал LP (февраль 1997 г.). «О вирусах, сексе и материнстве». Дж. Вирол. 71 (2): 859–65. Дои:10.1128 / JVI.71.2.859-865.1997. ЧВК 191132. PMID 8995601.
- ^ Хьюз Дж. Ф., Гроб Дж. М. (декабрь 2001 г.). «Доказательства геномных перестроек, опосредованных человеческими эндогенными ретровирусами во время эволюции приматов». Природа Генетика. 29 (4): 487–89. Дои:10,1038 / ng775. PMID 11704760. S2CID 20262212.
- ^ а б Докинз Р., Лилаюват С., Гаудиери С., Тай Г., Хуэй Дж., Каттли С. и др. (Февраль 1999 г.). «Геномика главного комплекса гистосовместимости: гаплотипы, дупликация, ретровирусы и болезни». Иммунологические обзоры. 167: 275–304. Дои:10.1111 / j.1600-065X.1999.tb01399.x. PMID 10319268.
- ^ Доксиадис Г.Г., де Гроот Н., Бонтроп Р.Э. (июль 2008 г.). «Влияние эндогенных интронных ретровирусов на разнообразие и стабильность класса II главного комплекса гистосовместимости». Журнал вирусологии. 82 (13): 6667–77. Дои:10.1128 / JVI.00097-08. ЧВК 2447082. PMID 18448532.
- ^ а б Чуонг Е.Б., Руми М.А., Соарес М.Дж., Бейкер Дж.С. (март 2013 г.). «Эндогенные ретровирусы функционируют как видоспецифические энхансерные элементы в плаценте». Природа Генетика. 45 (3): 325–29. Дои:10.1038 / ng.2553. ЧВК 3789077. PMID 23396136.
- ^ Томас JH, Шнайдер S (2011). «Коэволюция ретроэлементов и тандемных генов цинковых пальцев». Genome Res. 21 (11): 1800–12. Дои:10.1101 / гр.121749.111. ЧВК 3205565. PMID 21784874.
- ^ Кацуракис А., Магиоркинис Г., Лим А.Г., Гупта С., Белшоу Р., Гиффорд Р. (июль 2014 г.). «Больший размер тела млекопитающего приводит к снижению ретровирусной активности». Патогены PLOS. 10 (7): e1004214. Дои:10.1371 / journal.ppat.1004214. ЧВК 4102558. PMID 25033295.
- ^ Баннерт Н., Курт Р. (октябрь 2004 г.). «Ретроэлементы и геном человека: новые взгляды на старые отношения». Труды Национальной академии наук Соединенных Штатов Америки. 101 Дополнение 2 (Дополнение 2): 14572–79. Bibcode:2004PNAS..10114572B. Дои:10.1073 / pnas.0404838101. ЧВК 521986. PMID 15310846.
- ^ Нельсон П.Н., Карнеги П.Р., Мартин Дж., Давари Эйтехади Х., Хули П., Роден Д. и др. (Февраль 2003 г.). «Демистифицировано. Эндогенные ретровирусы человека». Молекулярная патология. 56 (1): 11–18. Дои:10.1136 / mp.56.1.11. ЧВК 1187282. PMID 12560456.
- ^ Сингх С.К. (июнь 2007 г.). «Эндогенные ретровирусы: подозреваемые в мире болезней». Будущая микробиология. 2 (3): 269–75. Дои:10.2217/17460913.2.3.269. PMID 17661701.
- ^ Магиоркинис Г., Белшоу Р., Кацуракис А. (сентябрь 2013 г.). "'Туда и обратно »: пересмотр патофизиологической роли эндогенных ретровирусов человека в постгеномную эру». Философские труды Лондонского королевского общества. Серия B, Биологические науки. 368 (1626): 20120504. Дои:10.1098 / rstb.2012.0504. ЧВК 3758188. PMID 23938753.
- ^ Мамели Дж., Астон В., Арру Дж., Маркони С., Ловато Л., Серра С. и др. (Январь 2007 г.). «Мононуклеарные клетки мозга и периферической крови пациентов с рассеянным склерозом (РС) гиперэкспрессируют РС-ассоциированный ретровирус / эндогенный ретровирус HERV-W, но не герпесвирус 6 человека». Журнал общей вирусологии. 88 (Pt 1): 264–74. Дои:10.1099 / vir.0.81890-0. PMID 17170460.
- ^ Серра К., Мамели Дж., Арру Дж., Сотгиу С., Росати Дж., Долей А. (декабрь 2003 г.). «Модуляция in vitro ретровируса, ассоциированного с рассеянным склерозом (РС), цитокинами: последствия для патогенеза РС». Журнал нейровирологии. 9 (6): 637–43. Дои:10.1080/714044485. PMID 14602576.
- ^ «Реактивированный вирус может способствовать развитию БАС». 2016-01-23.
- ^ Карамитрос Т., Херст Т., Марчи Э., Карамичали Э., Георгопулу Ю., Ментис А. и др. (Октябрь 2018 г.). «RASGRF2 связан со злоупотреблением наркотиками внутривенно и модулирует транскрипцию в модели клеточной линии». Труды Национальной академии наук Соединенных Штатов Америки. 115 (41): 10434–39. Дои:10.1073 / pnas.1811940115. ЧВК 6187174. PMID 30249655.
- ^ Роб Пичета. «Зависимость может быть вызвана древним ретровирусом, - говорится в исследовании». CNN. Получено 2019-10-13.
- ^ «Древний вирус может способствовать развитию зависимости у современных людей». Экономист. 2018-10-02. ISSN 0013-0613. Получено 2019-10-13.
- ^ Йолкен Р (июнь 2004 г.). «Вирусы и шизофрения: внимание к вирусу простого герпеса». Герпес. 11 (Дополнение 2): 83A – 88A. PMID 15319094.
- ^ Фокс Д (2010). "Вирус безумия". Обнаружить. Получено 2011-02-17.
- ^ а б c d Пейшенс С., Такеучи Ю., Вайс Р.А. (март 1997 г.). «Заражение клеток человека эндогенным ретровирусом свиней». Природа Медицина. 3 (3): 282–86. Дои:10,1038 / нм0397-282. PMID 9055854.
- ^ Platanias LC (май 2005 г.). «Механизмы передачи сигналов, опосредованной интерфероном типа I и типа II». Обзоры природы. Иммунология. 5 (5): 375–86. Дои:10.1038 / nri1604. PMID 15864272.
- ^ Grandi, N; Трамонтано, Э (2018). «Эндогенные ретровирусы человека - это древние приобретенные элементы, которые все еще формируют врожденные иммунные реакции». Границы иммунологии. 9: 2039. Дои:10.3389 / fimmu.2018.02039. ЧВК 6139349. PMID 30250470.
- ^ Niu D, Wei HJ, Lin L, George H, Wang T, Lee IH, Zhao HY, Wang Y, Kan Y, Shrock E, Lesha E, Wang G, Luo Y, Qing Y, Jiao D, Zhao H, Zhou X , Ван С., Вэй Х, Гуэль М., Чёрч Г.М., Ян Л. (сентябрь 2017 г.). «Инактивация эндогенного ретровируса свиней у свиней с использованием CRISPR-Cas9». Наука. 357 (6357): 1303–07. Bibcode:2017Научный ... 357.1303N. Дои:10.1126 / science.aan4187. ЧВК 5813284. PMID 28798043.
- ^ а б Тан Х. Б., Оуян К., Рао ГБ, Ма Л., Чжун Х., Бай А. и др. (01.01.2016). «Характеристика полных последовательностей генома свиного эндогенного ретровируса, выделенного из китайских мини-свиней, выявляет время эволюции раньше, чем изолятов европейских мини-свиней». Трансплантация. 48 (1): 222–28. Дои:10.1016 / j.transproceed.2015.12.005. PMID 26915872.
- ^ а б Tönjes RR, Niebert M (ноябрь 2003 г.). «Относительный возраст последовательностей провирусных эндогенных ретровирусов свиней в Sus scrofa на основе гипотезы молекулярных часов». Журнал вирусологии. 77 (22): 12363–68. Дои:10.1128 / JVI.77.22.12363-12368.2003. ЧВК 254287. PMID 14581574.
- ^ Парадис К., Лэнгфорд Дж., Лонг З., Хенеин В., Сандстром П., Свитцер В.М. и др. (Август 1999 г.). «Поиск межвидовой передачи эндогенного ретровируса свиней у пациентов, получавших лечение живой тканью свиньи. Исследовательская группа XEN 111». Наука. 285 (5431): 1236–41. Дои:10.1126 / science.285.5431.1236. PMID 10455044.
- ^ "Genomics England› Комплексный домен GeCIP для патогенов и мобильных элементов ". Получено 2019-10-13.
- ^ Гиффорд, RJ; Бломберг, Дж; Гроб, JM; Вентилятор, H; Heidmann, T; Майер, Дж; Стоу, Дж; Тристем, М; Джонсон, WE (28 августа 2018 г.). «Номенклатура локусов эндогенного ретровируса (ERV)». Ретровирология. 15 (1): 59. Дои:10.1186 / s12977-018-0442-1. ЧВК 6114882. PMID 30153831.
- ^ Белшоу Р., Доусон А.Л., Вулвен-Аллен Дж., Реддинг Дж., Берт А., Тристем М. (октябрь 2005 г.). «Общегеномный скрининг выявляет высокие уровни инсерционного полиморфизма в семействе эндогенных ретровирусов человека HERV-K (HML2): последствия для современной деятельности». Дж. Вирол. 79 (19): 12507–14. Дои:10.1128 / JVI.79.19.12507-12514.2005. ЧВК 1211540. PMID 16160178.
- ^ Jha AR, Pillai SK, York VA, Sharp ER, Storm EC, Wachter DJ, Martin JN, Deeks SG, Rosenberg MG, Nixon DF, Garrison KE (август 2009 г.). «Поперечное датирование новых гаплотипов HERV-K 113 и HERV-K 115 указывает на то, что эти провирусы возникли в Африке до появления Homo sapiens». Мол Биол Эвол. 26 (11): 2617–26. Дои:10.1093 / молбев / msp180. ЧВК 2760466. PMID 19666991.
- ^ Jha AR, Nixon DF, Rosenberg MG, Martin JN, Deeks SG, Hudson RR, Garrison KE, Pillai SK (май 2011 г.). «Эндогенный ретровирус человека K106 (HERV-K106) был заразным после появления людей современного анатомического типа». PLOS ONE. 6 (5): e20234. Bibcode:2011PLoSO ... 620234J. Дои:10.1371 / journal.pone.0020234. ЧВК 3102101. PMID 21633511.
- ^ Биениаш, Пол; Ли, Ён Нам (январь 2007 г.). «Восстановление инфекционного эндогенного ретровируса человека». PLOS Pathog. 3 (1): e10. Дои:10.1371 / journal.ppat.0030010. ЧВК 1781480. PMID 17257061.
- ^ Деваннье М., Харпер Ф, Ришо А., Летцельтер С., Рибет Д., Пьерон Г., Хайдманн Т. (декабрь 2006 г.). «Идентификация инфекционного предшественника многокопийных эндогенных ретроэлементов HERV-K человека». Genome Res. 16 (12): 1548–56. Дои:10.1101 / гр.5565706. ЧВК 1665638. PMID 17077319. Сложить резюме – Новости ScienceNOW Daily.
- ^ Чыонг Е.Б., Элде NC, Фешотт С. (март 2016 г.). «Регуляторная эволюция врожденного иммунитета посредством кооптации эндогенных ретровирусов». Наука. 351 (6277): 1083–87. Bibcode:2016Научный ... 351.1083C. Дои:10.1126 / science.aad5497. ЧВК 4887275. PMID 26941318.
- ^ а б Гаррисон К.Э., Джонс Р.Б., Мейкледжон Д.А., Анвар Н., Ндхлову Л.К., Чепмен Дж. М., Эриксон А. Л., Агравал А., Споттс Г., Хехт Ф. М., Ракофф-Нахум С., Ленц Дж., Островски М. А., Никсон Д. Ф. (ноябрь 2007 г.). «Т-клеточные ответы на эндогенные ретровирусы человека при инфекции ВИЧ-1». PLOS Pathog. 3 (11): e165. Дои:10.1371 / journal.ppat.0030165. ЧВК 2065876. PMID 17997601.
- ^ Морозов В.А., Дао Тхи В.Л., Деннер Дж. (2013). «Трансмембранный белок эндогенного ретровируса человека - K (HERV-K) модулирует высвобождение цитокинов и экспрессию генов». PLOS ONE. 8 (8): e70399. Bibcode:2013PLoSO ... 870399M. Дои:10.1371 / journal.pone.0070399. ЧВК 3737193. PMID 23950929.
- ^ Майер Дж., Бломберг Дж., Сил Р.Л. (май 2011 г.). «Пересмотренная номенклатура транскрибированных эндогенных ретровирусных локусов человека». Мобильная ДНК. 2 (1): 7. Дои:10.1186/1759-8753-2-7. ЧВК 3113919. PMID 21542922.
дальнейшее чтение
- Лёвер Р., Лёвер Дж., Курт Р. (май 1996 г.). «Вирусы в каждом из нас: характеристики и биологическое значение последовательностей эндогенных ретровирусов человека». Proc Natl Acad Sci USA. 93 (11): 5177–84. Bibcode:1996PNAS ... 93.5177L. Дои:10.1073 / пнас.93.11.5177. ЧВК 39218. PMID 8643549.
- Молес Дж. П., Тесьер А., Гильху Дж. Дж. (Июль 2005 г.). «Новая эндогенная ретровирусная последовательность экспрессируется в коже пациентов с псориазом». Br J Dermatol. 153 (1): 83–89. Дои:10.1111 / j.1365-2133.2005.06555.x. PMID 16029331.
- Зайфарт В., Франк О., Цайльфельдер Ю., Списс Б., Гринвуд А. Д., Хельманн Р., Лейб-Мёш К. (январь 2005 г.). «Комплексный анализ транскрипционной активности эндогенного ретровируса человека в тканях человека с помощью ретровирус-специфичного микрочипа». Дж. Вирол. 79 (1): 341–52. Дои:10.1128 / JVI.79.1.341-352.2005. ЧВК 538696. PMID 15596828.
- Кнерр И., Байндер Э, Рашер В. (февраль 2002 г.). «Синцитин, новый человеческий эндогенный ретровирусный ген в человеческой плаценте: доказательства его дисрегуляции при преэклампсии и HELLP-синдроме». Am J Obstet Gynecol. 186 (2): 210–13. Дои:10.1067 / моб.2002.119636. PMID 11854637.
- Гиффорд Р., Тристем М. (май 2003 г.). «Эволюция, распространение и разнообразие эндогенных ретровирусов» (PDF). Гены вирусов. 26 (3): 291–315. Дои:10.1023 / А: 1024455415443. PMID 12876457. Архивировано из оригинал (PDF) на 2013-02-15. Получено 2008-01-16.
внешняя ссылка
- Эндогенные + ретровирусы в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)
- HERVd - база данных эндогенных ретровирусов человека