Дублирование гена - Gene duplication
Дублирование гена (или же хромосомная дупликация или амплификация гена) является основным механизмом, посредством которого генерируется новый генетический материал во время молекулярная эволюция. Его можно определить как любое дублирование области ДНК который содержит ген. Дупликации генов могут возникать в результате нескольких типов ошибок в Репликация ДНК и ремонт машины, а также через случайный захват эгоистичными генетическими элементами. Общие источники дупликации генов включают: эктопическая рекомбинация, ретротранспозиция мероприятие, анеуплоидия, полиплоидия, и проскальзывание репликации.[1]
Механизмы дублирования
Эктопическая рекомбинация
Дублирование возникает в результате события, называемого неравный кроссинговер это происходит во время мейоза между смещенными гомологичными хромосомами. Вероятность того, что это произойдет, зависит от степени совместного использования повторяющихся элементов между двумя хромосомами. Продуктами этой рекомбинации являются дупликация в месте обмена и реципрокная делеция. Эктопическая рекомбинация обычно опосредуется сходством последовательностей в точках дублирования, которые образуют прямые повторы. Повторяющиеся генетические элементы, такие как сменный элементы предлагают один источник повторяющейся ДНК, которая может облегчить рекомбинацию, и они часто обнаруживаются в точках разрыва дупликации у растений и млекопитающих.[2]
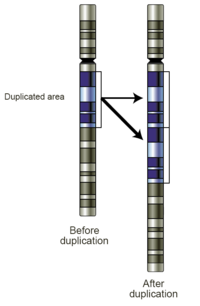
Проскальзывание репликации
Проскальзывание репликации Ошибка репликации ДНК, которая может приводить к дублированию коротких генетических последовательностей. Во время репликации ДНК-полимераза начинает копировать ДНК. В какой-то момент в процессе репликации полимераза отделяется от ДНК, и репликация останавливается. Когда полимераза снова присоединяется к цепи ДНК, она выравнивает реплицирующуюся цепь в неправильном положении и случайно копирует один и тот же участок более одного раза. Проскальзыванию репликации также часто способствуют повторяющиеся последовательности, но требуется лишь несколько оснований для сходства.
Ретротранспозиция
Ретротранспозоны, в основном L1, иногда может действовать на мРНК клетки. Транскрипты обратно транскрибируются в ДНК и вставляются в случайное место в геноме, создавая ретрогены. Результирующая последовательность обычно не имеет интронов и часто содержит поли, последовательности, которые также интегрированы в геном. Многие ретрогены обнаруживают изменения в регуляции генов по сравнению с последовательностями их родительских генов, что иногда приводит к новым функциям.
Анеуплоидия
Анеуплоидия возникает, когда нерасхождение одной хромосомы приводит к ненормальному количеству хромосом. Анеуплоидия часто опасна и у млекопитающих регулярно приводит к самопроизвольным абортам (выкидышам). Некоторые анеуплоидные особи жизнеспособны, например трисомия 21 у человека, которая приводит к Синдром Дауна. Анеуплоидия часто изменяет дозировку генов вредным для организма образом; поэтому маловероятно, что он будет распространяться среди населения.
Дублирование всего генома
Дублирование всего генома, или полиплоидия, является продуктом нерасхождение во время мейоза, что приводит к появлению дополнительных копий всего генома. Полиплоидия является обычным явлением у растений, но исторически также встречалась и у животных, с двумя раундами дупликации всего генома в линии позвоночных, ведущих к человеку.[3] После дупликации всего генома многие наборы дополнительных генов в конечном итоге теряются, возвращаясь в одноэлементное состояние. Однако сохранение многих генов, прежде всего Hox-гены, привело к адаптивным инновациям.
Полиплоидия также является хорошо известным источником видообразования, поскольку потомство, которое имеет другое количество хромосом по сравнению с родительскими видами, часто неспособно скрещиваться с неполиплоидными организмами. Считается, что дупликации всего генома менее вредны, чем анеуплоидия, поскольку относительная доза отдельных генов должна быть одинаковой.
Как эволюционное событие

Скорость дупликации гена
Сравнение геномов показывает, что дупликации генов обычны у большинства исследованных видов. На это указывает переменное количество копий (вариация количества копий) в геноме человека.[4][5] или плодовые мушки.[6] Однако было трудно измерить скорость, с которой происходит такое дублирование. Недавние исследования дали первую прямую оценку скорости дупликации генов у C. elegans, первый многоклеточный эукариот, для которого такие оценки стали доступны. Скорость дупликации гена в C. elegans порядка 10−7 дупликаций / генов / поколений, то есть в популяции из 10 миллионов червей у одного будет дупликация гена на поколение. Эта частота на два порядка больше, чем частота спонтанных точечных мутаций на нуклеотидный сайт у этого вида.[7] В более ранних (непрямых) исследованиях сообщалось о показателях локус-специфической дупликации у бактерий, Дрозофила, а люди - от 10−3 до 10−7/ ген / поколение.[8][9][10]
Неофункционализация
Дупликации генов - важный источник генетической новизны, которая может привести к эволюционным инновациям. Дупликация создает генетическую избыточность, при которой вторая копия гена часто свободна от селективное давление -то есть, мутации из него не оказывают вредного воздействия на организм-хозяин. Если одна копия гена подвергается мутации, которая влияет на ее исходную функцию, вторая копия может служить «запасной частью» и продолжать правильно функционировать. Таким образом, повторяющиеся гены накапливают мутации быстрее, чем функциональные гены-единственные копии, на протяжении поколений организмов, и для одной из двух копий возможно развитие новой и иной функции. Некоторые примеры такой неофункционализации - очевидная мутация дублированного пищеварительного гена в семействе ледяная рыба в ген антифриза и дублирование, ведущее к новому гену змеиного яда[11] и синтез 1-бета-гидрокситестостерона у свиней.[12]
Считается, что дупликация генов играет важную роль в эволюция; Такую позицию придерживаются представители научного сообщества более 100 лет.[13] Сусуму Оно был одним из самых известных разработчиков этой теории в своей классической книге Эволюция путем дупликации генов (1970).[14] Оно утверждал, что дупликация генов - самая важная эволюционная сила с момента появления универсальный общий предок.[15]Главный дупликация генома события могут быть довольно обычными. Считается, что весь дрожжи геном претерпела дублирование около 100 миллионов лет назад.[16] Растения являются наиболее плодовитыми дупликаторами генома. Например, пшеница гексаплоид (разновидность полиплоид ), что означает, что у него шесть копий своего генома.
Субфункционализация
Другая возможная судьба повторяющихся генов заключается в том, что обе копии в равной степени могут накапливать дегенеративные мутации, если любые дефекты дополняются другой копией. Это приводит к нейтральной модели «субфункционализации» или DDC (дублирование-дегенерация-комплементация),[17][18] в котором функциональность исходного гена распределена между двумя копиями. Ни один из генов не может быть потерян, так как оба теперь выполняют важные неизбыточные функции, но в конечном итоге ни один из них не может достичь новых функций.
Субфункционализация может происходить через нейтральные процессы, в которых мутации накапливаются без вредных или полезных эффектов. Однако в некоторых случаях может происходить субфункционализация с явными адаптивными преимуществами. Если наследственный ген плейотропный и выполняет две функции, часто ни одна из этих двух функций не может быть изменена, не затрагивая другую функцию. Таким образом, разделение наследственных функций на два отдельных гена может позволить адаптивную специализацию подфункций, тем самым обеспечивая адаптивное преимущество.[19]
Потеря
Часто возникающие геномные вариации приводят к неврологическим расстройствам, зависящим от дозировки гена, таким как Синдром Ретта и Болезнь Пелицея-Мерцбахера.[20] Такие вредные мутации, вероятно, будут потеряны в популяции и не будут сохранены или не будут развиваться в новых функциях. Однако многие дублирования на самом деле не являются вредными или полезными, и эти нейтральные последовательности могут быть потеряны или могут распространяться по популяции посредством случайных колебаний через генетический дрейф.
Выявление дупликаций в секвенированных геномах
Критерии и сканирование одного генома
Два гена, которые существуют после события дублирования гена, называются паралоги и обычно код для белки с аналогичной функцией и / или структурой. Напротив, ортологичный гены, присутствующие у разных видов, каждый из которых происходит от одной и той же наследственной последовательности. (Видеть Гомология последовательностей в генетике ).
В биологических исследованиях важно (но часто трудно) различать паралогов и ортологов. Эксперименты с функцией генов человека часто можно проводить на других разновидность если гомолог человеческого гена можно найти в геноме этого вида, но только если гомолог является ортологом. Если они паралоги и возникли в результате дублирования гена, их функции, вероятно, будут слишком разными. Одна или несколько копий дублированных генов, составляющих семейство генов, могут быть затронуты вставкой сменные элементы что вызывает значительные различия между ними в их последовательности и, наконец, может стать причиной дивергентная эволюция. Это также может повлиять на вероятность и скорость преобразование гена между гомологами дубликатов генов из-за меньшего сходства или отсутствия сходства в их последовательностях.
Паралоги могут быть идентифицированы в отдельных геномах путем сравнения последовательностей всех аннотированных генных моделей друг с другом. Такое сравнение может быть выполнено с транслированными аминокислотными последовательностями (например, BLASTp, tBLASTx) для выявления древних дупликаций или с нуклеотидными последовательностями ДНК (например, BLASTn, мегабластные) для выявления более поздних дупликаций. В большинстве исследований по выявлению дупликаций генов требуются взаимные наилучшие совпадения или нечеткие взаимные наилучшие совпадения, где каждый паралог должен быть единственным лучшим совпадением другого при сравнении последовательностей.[21]
Большинство дупликаций генов существуют как низкая копия повторяется (LCR), довольно часто повторяющиеся последовательности, такие как мобильные элементы. В основном они встречаются в перицентрономический, субтеломерный и межстраничный участки хромосомы. Многие LCR из-за своего размера (> 1 КБ), сходства и ориентации очень чувствительны к дупликациям и делециям.
Геномные микрочипы обнаруживают дупликации
Такие технологии, как геномные микрочипы, также называемый сравнением массивов геномный гибридизация (массив CGH), используются для обнаружения хромосомных аномалий, таких как микродупликации, с высокой пропускной способностью из образцов геномной ДНК. В частности, ДНК микрочип технология может одновременно контролировать выражение уровней тысяч генов при различных методах лечения или экспериментальных условиях, что значительно облегчает эволюционные исследования генная регуляция после дупликации гена или видообразование.[22][23]
Секвенирование нового поколения
Дупликации генов также можно идентифицировать с помощью платформ секвенирования следующего поколения. Простейший способ выявления дупликаций в данных геномного повторного секвенирования - использование считывания парных концов секвенирования. Тандемные дупликации указываются секвенированием пар чтения, которые отображаются в аномальной ориентации. Благодаря комбинации увеличенного покрытия последовательностей и аномальной ориентации картирования можно идентифицировать дупликации в данных геномного секвенирования.
Как усиление
Дублирование генов не обязательно означает длительное изменение генома вида. Фактически, такие изменения часто не длятся дольше исходного организма-хозяина. С точки зрения молекулярная генетика, амплификация гена это один из многих способов, которыми ген возможно чрезмерно выраженный. Генетическая амплификация может происходить искусственно, например, с использованием полимеразной цепной реакции техника для усиления коротких прядей ДНК in vitro с помощью ферменты, или это может происходить естественным путем, как описано выше. Если это естественное дублирование, оно все еще может иметь место в Соматическая клетка, а не зародышевый клетка (которая необходима для длительного эволюционного изменения).
Роль в раке
Дублирование онкогены являются частой причиной многих типов рак. В таких случаях генетическая дупликация происходит в соматической клетке и затрагивает только геном самих раковых клеток, а не весь организм, не говоря уже о любом последующем потомстве.
| Тип рака | Ассоциированный ген усиления | Распространенность усиление по типу рака (процент) |
|---|---|---|
| Рак молочной железы | МОЙ С | 20%[24] |
| ERBB2 (HER2 ) | 20%[24] | |
| CCND1 (Циклин D1 ) | 15–20%[24] | |
| FGFR1 | 12%[24] | |
| FGFR2 | 12%[24] | |
| Рак шейки матки | МОЙ С | 25–50%[24] |
| ERBB2 | 20%[24] | |
| Колоректальный рак | HRAS | 30%[24] |
| KRAS | 20%[24] | |
| MYB | 15–20%[24] | |
| Рак пищевода | МОЙ С | 40%[24] |
| CCND1 | 25%[24] | |
| MDM2 | 13%[24] | |
| Рак желудка | CCNE (Циклин E ) | 15%[24] |
| KRAS | 10%[24] | |
| ВСТРЕТИЛИСЬ | 10%[24] | |
| Глиобластома | ERBB1 (EGFR ) | 33–50%[24] |
| CDK4 | 15%[24] | |
| Рак головы и шеи | CCND1 | 50%[24] |
| ERBB1 | 10%[24] | |
| МОЙ С | 7–10%[24] | |
| Гепатоцеллюлярный рак | CCND1 | 13%[24] |
| Нейробластома | MYCN | 20–25%[24] |
| Рак яичников | МОЙ С | 20–30%[24] |
| ERBB2 | 15–30%[24] | |
| AKT2 | 12%[24] | |
| Саркома | MDM2 | 10–30%[24] |
| CDK4 | 10%[24] | |
| Мелкоклеточный рак легкого | МОЙ С | 15–20%[24] |
Смотрите также
- Сравнительная геномика
- Человеческий геном
- Инпараноид
- Молекулярная эволюция
- Псевдоген
- Дупликация тандемного экзона
- Неравный переход
Рекомендации
- ^ Чжан Дж (2003). «Эволюция путем дупликации генов: обновление» (PDF). Тенденции в экологии и эволюции. 18 (6): 292–8. Дои:10.1016 / S0169-5347 (03) 00033-8.
- ^ «Определение дупликации гена». медицинский словарь medterms. MedicineNet. 2012-03-19.
- ^ Дехал П., Буре Дж. Л. (октябрь 2005 г.). «Два раунда дупликации всего генома у предкового позвоночного». PLoS Биология. 3 (10): e314. Дои:10.1371 / journal.pbio.0030314. ЧВК 1197285. PMID 16128622.
- ^ Себат Дж., Лакшми Б., Троге Дж., Александр Дж., Янг Дж., Лундин П. и др. (Июль 2004 г.). «Крупномасштабный полиморфизм числа копий в геноме человека». Наука. 305 (5683): 525–8. Bibcode:2004Наука ... 305..525С. Дои:10.1126 / science.1098918. PMID 15273396.
- ^ Яфрат А.Дж., Феук Л., Ривера М.Н., Листевник М.Л., Донахью П.К., Ци Й. и др. (Сентябрь 2004 г.). «Обнаружение крупномасштабных вариаций в геноме человека». Природа Генетика. 36 (9): 949–51. Дои:10,1038 / ng1416. PMID 15286789.
- ^ Эмерсон Дж. Дж., Кардосо-Морейра М., Боревиц Дж. О., Лонг М. (июнь 2008 г.). «Естественный отбор формирует полногеномные модели полиморфизма числа копий у Drosophila melanogaster». Наука. 320 (5883): 1629–31. Bibcode:2008Sci ... 320.1629E. Дои:10.1126 / science.1158078. PMID 18535209.
- ^ Липински К.Дж., Фарслоу Дж. К., Фицпатрик К.А., Линч М., Катю В., Бергторссон Ю. (февраль 2011 г.). «Высокий уровень спонтанной дупликации генов у Caenorhabditis elegans». Текущая биология. 21 (4): 306–10. Дои:10.1016 / j.cub.2011.01.026. ЧВК 3056611. PMID 21295484.
- ^ Андерсон П., Рот Дж. (Май 1981 г.). «Спонтанные тандемные генетические дупликации у Salmonella typhimurium возникают из-за неравной рекомбинации между цистронами рРНК (рРНК)». Труды Национальной академии наук Соединенных Штатов Америки. 78 (5): 3113–7. Bibcode:1981PNAS ... 78.3113A. Дои:10.1073 / pnas.78.5.3113. ЧВК 319510. PMID 6789329.
- ^ Ватанабэ Ю., Такахаши А., Ито М., Такано-Симидзу Т. (март 2009 г.). «Молекулярный спектр спонтанных мутаций de novo в мужских и женских половых клетках Drosophila melanogaster». Генетика. 181 (3): 1035–43. Дои:10.1534 / генетика.108.093385. ЧВК 2651040. PMID 19114461.
- ^ Тернер Д. Д., Миретти М., Раджан Д., Фиглер Х., Картер Н. П., Блейни М. Л. и др. (Январь 2008 г.). «Частота мейотических делеций и дупликаций de novo в зародышевой линии, вызывающих несколько геномных нарушений». Природа Генетика. 40 (1): 90–5. Дои:10,1038 / нг.2007,40. ЧВК 2669897. PMID 18059269.
- ^ Линч VJ (январь 2007 г.). «Изобретая арсенал: адаптивная эволюция и неофункционализация генов фосфолипазы A2 змеиного яда». BMC Эволюционная биология. 7: 2. Дои:10.1186/1471-2148-7-2. ЧВК 1783844. PMID 17233905.
- ^ Conant GC, Wolfe KH (декабрь 2008 г.). «Превращение хобби в работу: как дублированные гены находят новые функции». Обзоры природы. Генетика. 9 (12): 938–50. Дои:10.1038 / nrg2482. PMID 19015656.
- ^ Тейлор Дж. С., Раес Дж. (2004). «Дублирование и дивергенция: эволюция новых генов и старых идей». Ежегодный обзор генетики. 38: 615–43. Дои:10.1146 / annurev.genet.38.072902.092831. PMID 15568988.
- ^ Оно, С. (1970). Эволюция путем дупликации генов. Springer-Verlag. ISBN 978-0-04-575015-3.
- ^ Оно, С. (1967). Половые хромосомы и сцепленные с полом гены. Springer-Verlag. ISBN 978-91-554-5776-1.
- ^ Келлис М., Биррен Б.В., Лендер Е.С. (апрель 2004 г.). «Доказательство и эволюционный анализ дупликации древнего генома дрожжей Saccharomyces cerevisiae». Природа. 428 (6983): 617–24. Bibcode:2004Натура.428..617K. Дои:10.1038 / природа02424. PMID 15004568.
- ^ Force A, Линч M, Пикетт FB, Amores A, Yan YL, Postlethwait J (апрель 1999 г.). «Сохранение повторяющихся генов за счет дополнительных дегенеративных мутаций». Генетика. 151 (4): 1531–45. ЧВК 1460548. PMID 10101175.
- ^ Штольцфус А (август 1999 г.). «О возможности конструктивной нейтральной эволюции». Журнал молекулярной эволюции. 49 (2): 169–81. Bibcode:1999JMolE..49..169S. CiteSeerX 10.1.1.466.5042. Дои:10.1007 / PL00006540. PMID 10441669.
- ^ Des Marais DL, Rausher MD (август 2008 г.). «Избежать адаптивного конфликта после дупликации в гене пути антоциана». Природа. 454 (7205): 762–5. Bibcode:2008Натура.454..762D. Дои:10.1038 / природа07092. PMID 18594508.
- ^ Ли Дж. А., Лупски-младший (октябрь 2006 г.). «Геномные перестройки и изменения числа копий генов как причина расстройств нервной системы». Нейрон. 52 (1): 103–21. Дои:10.1016 / j.neuron.2006.09.027. PMID 17015230.
- ^ Хан М.В., Хан М.В., Хан С.Г. (ноябрь 2007 г.). «Эволюция семейства генов в 12 геномах дрозофилы». PLoS Genetics. 3 (11): e197. Дои:10.1371 / journal.pgen.0030197. ЧВК 2065885. PMID 17997610.
- ^ Мао Р., Певснер Дж. (2005). «Использование геномных микрочипов для изучения хромосомных аномалий при умственной отсталости». Обзоры исследований в области умственной отсталости и пороков развития. 11 (4): 279–85. Дои:10.1002 / mrdd.20082. PMID 16240409.
- ^ Гу X, Чжан З., Хуан В. (январь 2005 г.). «Быстрая эволюция экспрессии и регуляторных расхождений после дупликации дрожжевого гена». Труды Национальной академии наук Соединенных Штатов Америки. 102 (3): 707–12. Bibcode:2005ПНАС..102..707Г. Дои:10.1073 / pnas.0409186102. ЧВК 545572. PMID 15647348.
- ^ а б c d е ж грамм час я j k л м п о п q р s т ты v ш Икс у z аа ab ac Кинзлер К.В., Фогельштейн Б. (2002). Генетическая основа рака человека. Макгроу-Хилл. п. 116. ISBN 978-0-07-137050-9.