Аллель - Allele
An аллель (Великобритания: /ˈæлял/, /əˈлял/; нас: /əˈлял/; современное образование от греческого ἄλλος állos, "Другой")[1][2][3] одна из двух или более форм данного ген вариант.[4] Например. то Группа крови АВО контролируется Ген ABO который имеет шесть общих аллелей. Почти каждого живого человека фенотип для гена ABO - это некая комбинация этих шести аллелей.[5][6] Аллель - это одна из двух или более версий одного и того же гена в то же место на хромосома. Он также может относиться к различным вариациям последовательности для нескольких сотен пар оснований или более участка генома, который кодирует белок. Аллели могут быть разных размеров. При минимально возможном размере аллель может быть однонуклеотидный полиморфизм (SNP).[7] На более высоком уровне это может быть до нескольких тысяч пар оснований длинный.[8][9] Большинство аллелей приводят к небольшому или отсутствующему заметному изменению функции белка, кодируемого геном.
Однако иногда разные аллели могут приводить к разным наблюдаемым фенотипические признаки, например, разные пигментация. Ярким примером этого является Грегор Мендель открытие, что белый и фиолетовый цвета цветов в горох растения были результатом признаков «чистой линии», то есть одного гена с двумя аллелями.
Все многоклеточные организмы иметь два набора хромосомы в какой-то момент в их жизненный цикл; то есть они диплоид. В этом случае хромосомы могут быть парный. Каждая хромосома в паре содержит одни и те же гены в одинаковом порядке и в одном месте по длине хромосомы. За данный ген, если две хромосомы содержат один и тот же аллель, они и организм гомозиготный относительно этого гена. Если аллели разные, они и организм гетерозиготный относительно этого гена.
Этимология
Слово «аллель» - это краткая форма аллеломорф ("другая форма", слово придумано британскими генетиками Уильям Бейтсон и Эдит Ребекка Сондерс ),[10][11] который использовался в первые дни генетика описать вариантные формы ген определяется как разные фенотипы. Это происходит из Греческий префикс ἀλληλο-, аллело-, что означает «взаимный», «взаимный» или «друг друга», что само по себе связано с греческим прилагательным ἄλλος, аллос (родственные латинский Алиус), что означает «другое».
Аллели, приводящие к доминантным или рецессивным фенотипам
Во многих случаях генотипические взаимодействия между двумя аллелями в локусе можно описать как доминирующий или же рецессивный, по какому из двух гомозиготных фенотипов гетерозигота больше всего похож. Если гетерозигота неотличима от одной из гомозигот, выраженный аллель - это тот, который приводит к «доминантному» фенотипу,[12] а другой аллель называется «рецессивным». Степень и характер доминирования различаются по локусам. Впервые этот тип взаимодействия формально описал Грегор Мендель. Однако многие черты не поддаются этой простой классификации, и фенотипы моделируются совместное доминирование и полигенное наследование.
Период, термин "дикого типа "аллель" иногда используется для описания аллеля, который, как считается, вносит свой вклад в типичный фенотипический характер, наблюдаемый в "диких" популяциях организмов, таких как плодовые мухи (Drosophila melanogaster ). Такой аллель «дикого типа» исторически считался ведущим к доминантному (подавляющему - всегда выраженному), общему и нормальному фенотипу, в отличие от «мутант «аллели, которые приводят к рецессивным, редким и часто вредным фенотипам. Раньше считалось, что большинство людей были гомозиготными по аллелю« дикого типа »в большинстве локусов генов, и что любой альтернативный« мутантный »аллель был обнаружен в гомозиготной форме в небольшое меньшинство "затронутых" лиц, часто генетические заболевания, и чаще в гетерозиготной форме в "перевозчики "для мутантного аллеля. В настоящее время принято во внимание, что большинство или все локусы генов очень полиморфны, с множественными аллелями, частота которых варьируется от популяции к популяции, и что большая часть генетических вариаций скрыта в форме аллелей, которые не производят очевидные фенотипические различия.
Множественные аллели




Население или разновидность организмов обычно включает несколько аллелей в каждом локусе у разных индивидуумов. Аллельные вариации в локусе можно измерить как количество аллелей (полиморфизм ), или доля гетерозигот в популяции. А нулевой аллель представляет собой вариант гена, у которого отсутствует нормальная функция гена, потому что он либо не экспрессируется, либо экспрессируемый белок неактивен.
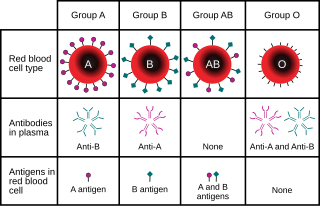
Например, в локусе гена для ABO группа крови углевод антигены в людях,[13] классическая генетика распознает три аллеля, IА, ЯB, и i, которые определяют совместимость переливание крови. У любого человека есть одно из шести возможных генотипы (ЯАяА, ЯАя, яBяB, ЯBя, яАяB, и ii) которые производят один из четырех возможных фенотипы: «Тип А» (производство IАяА гомозиготный и яАi гетерозиготные генотипы), «Тип B» (произведен IBяB гомозиготный и яBi гетерозиготные генотипы), "Тип AB", произведенный IАяB гетерозиготный генотип и "Тип О", полученный по гомозиготному генотипу ii. (Теперь известно, что каждый из аллелей A, B и O на самом деле является классом множества аллелей с разными последовательностями ДНК, которые продуцируют белки с идентичными свойствами: более 70 аллелей известны в локусе ABO.[14] Следовательно, человек с кровью «типа А» может быть гетерозиготой АО, гомозиготой АА или гетерозиготой АА с двумя разными аллелями «А».)
Частоты генотипов
Частоту аллелей в диплоидной популяции можно использовать для прогнозирования частот соответствующих генотипов (см. Принцип Харди – Вайнберга ). Для простой модели с двумя аллелями;


куда п - частота одного аллеля и q - частота альтернативного аллеля, сумма которой обязательно равна единице. Потом, п2 - доля популяции, гомозиготная по первому аллелю, 2pq - доля гетерозигот, а q2 - фракция, гомозиготная по альтернативному аллелю. Если первый аллель доминирует над вторым, то доля популяции, которая покажет доминантный фенотип, равна п2 + 2pq, а фракция с рецессивным фенотипом - q2.
С тремя аллелями:
- и


В случае множественных аллелей в диплоидном локусе количество возможных генотипов (G) с количеством аллелей (a) определяется выражением:

Аллельное доминирование при генетических нарушениях
Номер генетические нарушения возникают, когда человек наследует два рецессивных аллеля по единственному гену. Рецессивные генетические нарушения включают: альбинизм, кистозный фиброз, галактоземия, фенилкетонурия (ФКУ) и Болезнь Тея – Сакса. Другие нарушения также связаны с рецессивными аллелями, но поскольку локус гена расположен на X-хромосоме, у мужчин есть только одна копия (то есть они гемизиготный ), у самцов они чаще, чем у самок. Примеры включают красно-зеленый дальтонизм и синдром ломкой Х-хромосомы.
Другие расстройства, такие как болезнь Хантингтона, возникают, когда человек наследует только один доминантный аллель.
Epialleles
Пока наследственные черты обычно изучаются с точки зрения генетических аллелей, эпигенетический такие знаки, как Метилирование ДНК могут быть унаследованы в определенных областях генома у определенных видов, и этот процесс называется трансгенерационное эпигенетическое наследование. Период, термин эпиаллель используется для отличия этих наследственных признаков от традиционных аллелей, которые определяются нуклеотидная последовательность.[15] Особый класс эпиаллелей, метастабильные эпиаллелы, был обнаружен у мышей и людей, который характеризуется стохастическим (вероятностным) установлением эпигенетического состояния, которое может передаваться митотически.[16][17]
Смотрите также
Ссылки и примечания
- ^ "Аллель | Значение аллеля по лексике". Словари Lexico | английский. Получено 7 января 2020.
- ^ «аллель существительное - определение, изображения, произношение и примечания по использованию». Оксфордский словарь для продвинутых учащихся. Получено 29 октября 2017.
- ^ «Значение аллеля в Кембриджском словаре английского языка». Dictionary.cambridge.org. Получено 29 октября 2017.
- ^ «Что такое варианты, аллели и гаплотипы? | Генетическая изменчивость человека». Получено 16 ноября 2020.
- ^ Сельцам А., Халленслебен М., Коллманн А., Блашик Р. (октябрь 2003 г.). «Природа разнообразия и диверсификации в локусе ABO». Кровь. 102 (8): 3035–42. Дои:10.1182 / кровь-2003-03-0955. PMID 12829588.
- ^ Огасавара К., Баннаи М., Сайто Н., Ябе Р., Наката К., Такенака М., Фудзисава К., Утикава М., Исикава Ю., Джудзи Т., Токунага К. (июнь 1996 г.). «Обширный полиморфизм гена группы крови ABO: три основных линии аллелей для общих фенотипов ABO». Генетика человека. 97 (6): 777–83. Дои:10.1007 / BF02346189. PMID 8641696.
- ^ Smigielski, Elizabeth M .; Сироткин, Карл; Уорд, Минхонг; Шерри, Стивен Т. (1 января 2000 г.). «dbSNP: база данных однонуклеотидных полиморфизмов». Исследования нуклеиновых кислот. 28 (1): 352–355. Дои:10.1093 / nar / 28.1.352. ISSN 0305-1048. ЧВК 102496. PMID 10592272.
- ^ Элстон, Роберт; Сатагопан, Джая; Солнце, Шуин (2012). «Генетическая терминология». Статистическая генетика человека. Методы молекулярной биологии (Клифтон, Нью-Джерси). 850. С. 1–9. Дои:10.1007/978-1-61779-555-8_1. ISBN 978-1-61779-554-1. ISSN 1064-3745. ЧВК 4450815. PMID 22307690.
- ^ "Какой эффект имеют варианты в кодирующих областях?". EMBL-EBI Поезд онлайн. 2 мая 2019. Получено 14 ноября 2019.
- ^ Ремесло, Джуд (2013). «Гены и генетика: язык научных открытий». Гены и генетика. Оксфордский словарь английского языка. Получено 14 января 2016.
- ^ Бейтсон, В. и Сондерс, Э. Р. (1902) "Факты наследственности в свете открытия Менделя". Отчитывается перед комитетом по эволюции Королевского общества, Я. стр. 125–160
- ^ Hartl, Daniel L .; Элизабет У. Джонс (2005). Существенная генетика: перспектива геномики (4-е изд.). Издательство "Джонс и Бартлетт". п. 600. ISBN 978-0-7637-3527-2.
- ^ Виктор А. МакКусик; Кассандра Л. Книффин; Пол Дж. Конверс; Ада Хамош (10 ноября 2009 г.). «АВО Гликозилтрансфераза; АВО». Онлайн-менделевское наследование в человеке. Национальная медицинская библиотека. В архиве из оригинала 24 сентября 2008 г.. Получено 24 марта 2010.
- ^ Ип СП (январь 2002 г.). «Вариация последовательности в локусе ABO человека». Анналы генетики человека. 66 (1): 1–27. Дои:10.1017 / S0003480001008995. PMID 12014997.
- ^ Даксинджер, Лючия; Уайтлоу, Эмма (31 января 2012 г.). «Понимание трансгенерационного эпигенетического наследования через гаметы у млекопитающих». Природа Обзоры Генетика. 13 (3): 153–62. Дои:10.1038 / nrg3188. PMID 22290458.
- ^ Ракян, Вардхман К; Блевитт, Марни Э; Друкер, Рики; Прейс, Йост I; Уайтлоу, Эмма (июль 2002 г.). «Метастабильные эпиаллелы у млекопитающих». Тенденции в генетике. 18 (7): 348–351. Дои:10.1016 / S0168-9525 (02) 02709-9. PMID 12127774.
- ^ Уотерленд, РА; Долиной, ДК; Lin, JR; Смит, Калифорния; Ши, X; Тахилиани, К.Г. (сентябрь 2006 г.). «Материнские метиловые добавки увеличивают метилирование ДНК потомства в Axin Fused». Бытие. 44 (9): 401–6. Дои:10.1002 / dvg.20230. PMID 16868943.