Хроматин - Chromatin

Хроматин это комплекс ДНК и белок нашел в эукариотический клетки.[1] Его основная функция: упаковка длинные молекулы ДНК превращаются в более компактные и плотные структуры. Это предотвращает спутывание нитей, а также играет важную роль в укреплении ДНК во время деление клеток, предотвращая Повреждение ДНК, и регулирующий экспрессия гена и Репликация ДНК. В течение митоз и мейоз, хроматин способствует правильной сегрегации хромосомы в анафаза; характерные формы хромосом, видимые на этой стадии, являются результатом свертывания ДНК в сильно конденсированный хроматин.
Первичные белковые компоненты хроматина: гистоны, которые связываются с ДНК и функционируют как «якоря», вокруг которых наматываются нити. В целом, существует три уровня организации хроматина:
- ДНК обвивает гистоновые белки, образуя нуклеосомы и так называемый бусы на нитке структура (эухроматин ).
- Множественные гистоны обертываются 30-нанометр волокно, состоящее из массивов нуклеосом в их наиболее компактной форме (гетерохроматин ).[а]
- Высший уровень Суперспирализация ДНК 30-нм волокна дает метафаза хромосома (во время митоза и мейоза).
Однако многие организмы не следуют этой схеме организации. Например, сперматозоиды и птичий красные кровяные тельца имеют более плотно упакованный хроматин, чем большинство эукариотических клеток, и трипаносоматид простейшие вообще не конденсируют свой хроматин в видимые хромосомы. Прокариотический клетки имеют совершенно разные структуры для организации своей ДНК (эквивалент прокариотической хромосомы называется генофор и локализован в нуклеоид область, край).
Общая структура хроматиновой сети в дальнейшем зависит от стадии клеточный цикл. В течение межфазный, хроматин структурно рыхлый, чтобы обеспечить доступ к РНК и ДНК-полимеразы который расшифровывать и воспроизвести ДНК. Локальная структура хроматина в интерфазе зависит от конкретного гены присутствует в ДНК. Активно транскрибируемые («включенные») участки ДНК, содержащие гены, менее плотно уплотнены и тесно связаны с РНК-полимеразами в структуре, известной как эухроматин, в то время как области, содержащие неактивные гены («выключены»), обычно более конденсированы и связаны со структурными белками в гетерохроматин.[3] Эпигенетический модификация структурных белков хроматина через метилирование и ацетилирование также изменяет локальную структуру хроматина и, следовательно, экспрессию генов. Структура хроматиновых сетей в настоящее время плохо изучена и остается активной областью исследований в молекулярная биология.
Динамическая структура и иерархия хроматина

Хроматин претерпевает различные структурные изменения во время клеточный цикл. Гистон белки являются основными упаковщиками и организаторами хроматина и могут быть модифицированы различными посттрансляционными модификациями для изменения упаковки хроматина (гистоновая модификация ). Большинство модификаций происходит на хвостах гистонов. Последствия с точки зрения доступности и уплотнения хроматина зависят как от модифицированной аминокислоты, так и от типа модификации. Например, ацетилирование гистонов приводит к разрыхлению и повышенной доступности хроматина для репликации и транскрипции. Триметилирование лизина может привести либо к усилению транскрипционной активности (триметилирование гистон H3 лизин 4) или репрессия транскрипции и уплотнение хроматина (триметилирование гистона H3, лизин 9 или 27). Несколько исследований показали, что разные модификации могут происходить одновременно. Например, было предложено двухвалентный структура (с триметилированием лизина 4 и 27 на гистоне H3) участвует в раннем развитии млекопитающих.[4]
Белки группы поликомб играют роль в регуляции генов посредством модуляции структуры хроматина.[5]
Для получения дополнительной информации см. Модификации гистонов в регуляции хроматина и Контроль РНК-полимеразы структурой хроматина.
Структура ДНК

В природе ДНК может образовывать три структуры: А-, B-, и Z-ДНК. A- и B-ДНК очень похожи, образуя правые спирали, тогда как Z-ДНК представляет собой левую спираль с зигзагообразным фосфатным остовом. Считается, что Z-ДНК играет особую роль в структуре хроматина и транскрипции из-за свойств соединения между B- и Z-ДНК.
На стыке B- и Z-ДНК одна пара оснований отрывается от нормального связывания. Они играют двойную роль: точки распознавания многими белками и приемник торсионного напряжения от РНК-полимераза или связывание нуклеосом.
Нуклеосомы и бусины на нитке
- Основные статьи: Нуклеосома, Хроматосома и Гистон
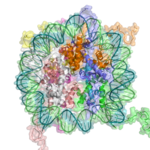
Основным повторяющимся элементом хроматина является нуклеосома, соединенная между собой участками линкер ДНК, гораздо более короткое расположение, чем чистая ДНК в растворе.
Помимо ядерных гистонов, линкер гистон H1 существует, который контактирует с выходом / входом нити ДНК на нуклеосоме. Частица ядра нуклеосомы вместе с гистоном H1 известна как хроматосома. Нуклеосомы, содержащие примерно от 20 до 60 пар оснований линкерной ДНК, могут образовывать, в нефизиологических условиях, примерно 10 нм бусы на нитке волокно.
Нуклеосомы связывают ДНК неспецифично, как того требует их функция в общей упаковке ДНК. Однако существуют большие предпочтения последовательностей ДНК, которые определяют расположение нуклеосом. Это связано в первую очередь с различными физическими свойствами различных последовательностей ДНК: например, аденин (A), и тимин (T) более предпочтительно вдавливаются во внутренние малые канавки. Это означает, что нуклеосомы могут связываться преимущественно в одном положении примерно через каждые 10 пар оснований (спиральное повторение ДНК) - где ДНК поворачивается, чтобы максимизировать количество оснований A и T, которые будут находиться во внутренней малой бороздке. (Видеть структура нуклеиновой кислоты.)
30-нанометровое хроматиновое волокно

Слева: 1 структура "соленоида" стартовой спирали.
Справа: 2 начала рыхлой спиральной структуры.
Примечание: гистоны на этой диаграмме не показаны - показана только ДНК.
С добавлением H1 бусины на нитке в свою очередь свертывается в спиральную структуру диаметром 30 нм, известную как 30 нм волокно или нить. Точная структура волокна хроматина в клетке подробно не известна.[6]
Этот уровень структуры хроматина считается формой гетерохроматин, который содержит в основном транскрипционно молчащие гены. Электронно-микроскопические исследования показали, что 30-нм волокно очень динамично, так что оно разворачивается в структуру 10-нм волокна в виде бусинок на нити при пересечении РНК-полимеразой, участвующей в транскрипции.

Линкерная ДНК желтого цвета и нуклеосомная ДНК розового цвета.
В существующих моделях принято считать, что нуклеосомы лежат перпендикулярно оси волокна, а линкерные гистоны расположены внутри. Стабильное 30-нм волокно основано на регулярном расположении нуклеосом вдоль ДНК. Линкерная ДНК относительно устойчива к изгибу и вращению. Это делает длину линкерной ДНК критичной для стабильности волокна, требуя, чтобы нуклеосомы были разделены на длины, которые позволяют вращение и складывание в требуемую ориентацию без чрезмерного напряжения для ДНК. С этой точки зрения, разная длина линкерной ДНК должна давать различные топологии сворачивания хроматинового волокна. Недавняя теоретическая работа, основанная на изображениях, полученных с помощью электронной микроскопии.[7]восстановленных волокон поддерживает эту точку зрения.[8]
Пространственная организация хроматина в ядре клетки
Пространственное расположение хроматина внутри ядра не является случайным - на определенных территориях можно найти специфические области хроматина. Территории, например, пластинка -ассоциированные домены (LAD) и топологически связывающие домены (ТАД), которые связаны вместе белковыми комплексами.[9] В настоящее время полимерные модели, такие как модель Strings & Binders Switch (SBS)[10] и модель динамического цикла (DL)[11] используются для описания сворачивания хроматина внутри ядра.
Структурная организация, зависящая от клеточного цикла


- Межфазный: Структура хроматина во время межфазный из митоз оптимизирован для обеспечения простого доступа к транскрипция и Ремонт ДНК факторов к ДНК, уплотняя ДНК в ядро. Структура варьируется в зависимости от необходимого доступа к ДНК. Гены которые требуют регулярного доступа со стороны РНК-полимераза требуют более рыхлой структуры, обеспечиваемой эухроматином.
- Метафаза: The метафаза структура хроматина сильно отличается от межфазный. Оптимизирован для физической силы[нужна цитата ] и управляемость, образующие классику хромосома структура замечена в кариотипы. Считается, что структура конденсированного хроматина представляет собой петли 30-нм волокна, ведущие к центральному каркасу белков. Однако он недостаточно охарактеризован. Хромосомные скаффолды играют важную роль в удерживании хроматина в компактных хромосомах. Петли структуры 30 нм далее конденсируются с каркасом в структуры более высокого порядка.[12] Хромосомные каркасы состоят из белков, в том числе конденсин, топоизомераза типа IIA и член семейства кинезинов 4 (KIF4).[13] Физическая сила хроматина жизненно важна на этой стадии деления, чтобы предотвратить повреждение ДНК сдвигом при разделении дочерних хромосом. Чтобы максимизировать силу, состав хроматина изменяется по мере приближения к центромере, прежде всего за счет альтернативных аналогов гистона H1. Во время митоза, хотя большая часть хроматина плотно уплотнена, есть небольшие участки, которые не так плотно уплотнены. Эти области часто соответствуют промоторным областям генов, которые были активны в этом типе клеток до образования хроматина. Отсутствие уплотнения этих регионов называется закладки, что является эпигенетический Этот механизм считается важным для передачи дочерним клеткам «памяти» о том, какие гены были активны до вступления в митоз.[14] Этот закладки необходим механизм, чтобы помочь передать эту память, потому что транскрипция прекращается во время митоз.
Хроматин и всплески транскрипции
Хроматин и его взаимодействие с ферментами были исследованы, и был сделан вывод, что он является важным фактором экспрессии генов. Винсент Г. Олфри, профессор Университета Рокфеллера, заявил, что синтез РНК связан с ацетилированием гистонов.[15] Аминокислота лизин, прикрепленная к концам гистонов, заряжена положительно. Ацетилирование этих хвостов сделает концы хроматина нейтральными, что обеспечит доступ ДНК.
Когда хроматин деконденсируется, ДНК открыта для проникновения молекулярных механизмов. Колебания между открытым и закрытым хроматином могут способствовать прерыванию транскрипции или транскрипционный разрыв. Вероятно, вовлечены и другие факторы, такие как ассоциация и диссоциация комплексов факторов транскрипции с хроматином. Этот феномен, в отличие от простых вероятностных моделей транскрипции, может объяснить высокую вариабельность экспрессии генов, происходящую между клетками в изогенных популяциях.[16]
Альтернативные организации хроматина
Во время многоклеточного спермиогенез, то сперматида Хроматин трансформируется в более разнесенную, расширенную, почти кристаллическую структуру. Этот процесс связан с прекращением транскрипция и включает ядерный белковый обмен. Гистоны в основном смещены и заменены протамины (маленький, аргинин -богатые белки).[17] Предполагается, что у дрожжей области, лишенные гистонов, становятся очень хрупкими после транскрипции; HMO1, an HMG-коробка белок, помогает в стабилизации хроматина без нуклеосом.[18][19]
Восстановление хроматина и ДНК
Упаковка эукариотической ДНК в хроматин представляет собой барьер для всех основанных на ДНК процессов, которые требуют привлечения ферментов к участкам их действия. Чтобы разрешить критический клеточный процесс репарации ДНК, хроматин должен быть реконструирован. У эукариот АТФ-зависимое ремоделирование хроматина комплексы и ферменты, модифицирующие гистоны являются двумя преобладающими факторами, используемыми для выполнения этого процесса ремоделирования.[20]
Релаксация хроматина происходит быстро в месте повреждения ДНК.[21] Инициирует этот процесс PARP1 белок, который начинает появляться при повреждении ДНК менее чем за секунду, с половиной максимального накопления в течение 1,6 секунды после повреждения.[22] Далее ремоделирующий хроматин Alc1 быстро прикрепляется к продукту PARP1 и достигает повреждения ДНК в течение 10 секунд после повреждения.[21] Примерно половина максимальной релаксации хроматина, предположительно из-за действия Alc1, происходит через 10 секунд.[21] Затем это позволяет задействовать фермент репарации ДНК. MRE11, чтобы начать восстановление ДНК, в течение 13 секунд.[22]
γH2AX, фосфорилированная форма H2AX также участвует в ранних стадиях, ведущих к деконденсации хроматина после возникновения повреждения ДНК. Вариант гистона H2AX составляет около 10% гистонов H2A в хроматине человека.[23] γH2AX (H2AX, фосфорилированный по серину 139) может быть обнаружен уже через 20 секунд после облучения клеток (с образованием двухцепочечного разрыва ДНК), а половина максимального накопления γH2AX происходит за одну минуту.[23] Размер хроматина с фосфорилированным γH2AX составляет около двух миллионов пар оснований в месте двухцепочечного разрыва ДНК.[23] γH2AX сам по себе не вызывает деконденсацию хроматина, но в течение 30 секунд после облучения RNF8 белок может быть обнаружен в ассоциации с γH2AX.[24] RNF8 опосредует обширную деконденсацию хроматина посредством его последующего взаимодействия с CHD4,[25] компонент ремоделирования нуклеосом и комплекса деацетилазы NuRD.
После релаксации после повреждения ДНК с последующей репарацией ДНК хроматин восстанавливается до состояния уплотнения, близкого к уровню до повреждения, примерно через 20 мин.[21]
Методы исследования хроматина
- ChIP-seq (Секвенирование иммунопреципитации хроматина), направленное против различных модификации гистонов, может использоваться для идентификации состояний хроматина по всему геному. Различные модификации были связаны с различными состояниями хроматина.
- DNase-seq (Секвенирование гиперчувствительных сайтов к ДНКазе I) использует чувствительность доступных участков генома к ДНКаза I фермент для картирования открытых или доступных областей в геноме.
- FAIRE-seq (Секвенирование выделения регуляторных элементов с помощью формальдегида) использует химические свойства связанной с белком ДНК в методе двухфазного разделения для извлечения областей, лишенных нуклеосом, из генома.[26]
- ATAC-seq (Анализ для секвенирования транспозиционного доступного хроматина) использует транспозазу Tn5 для интеграции (синтетических) транспозонов в доступные области генома, что, в свою очередь, подчеркивает локализацию нуклеосом и факторов транскрипции в геноме.
- ДНК-след это метод, направленный на идентификацию ДНК, связанной с белками. Он использует маркировку и фрагментацию в сочетании с гель-электрофорезом для идентификации областей генома, которые были связаны белками.[27]
- MNase-seq (Секвенирование микрококковой нуклеазы) использует микрококковая нуклеаза фермент для определения положения нуклеосом по всему геному.[28][29]
- Захват конформации хромосомы определяет пространственную организацию хроматина в ядре, предполагая геномные местоположения, которые физически взаимодействуют.
- Профилирование MACC (Профили доступности микрококковой нуклеазы) использует серию титрования гидролизатов хроматина с микрококковая нуклеаза для определения доступности хроматина, а также для картирования нуклеосом и негистоновых ДНК-связывающих белков как в открытых, так и в закрытых областях генома.[30]
Хроматин и узлы
Было загадкой, как деконденсированные интерфазные хромосомы остаются по существу незаузленными. Естественное ожидание состоит в том, что в присутствии топоизомераз ДНК типа II, которые допускают прохождения двухцепочечных участков ДНК друг через друга, все хромосомы должны достичь состояния топологического равновесия. Топологическое равновесие в сильно переполненных интерфазных хромосомах, образующих хромосомные территории, должно приводить к образованию сильно узловатых волокон хроматина. Однако методы Chromosome Conformation Capture (3C) показали, что разрушение контактов с геномным расстоянием в интерфазных хромосомах практически такое же, как в смятом состоянии глобулы, которое образуется, когда длинные полимеры конденсируются без образования каких-либо узлов. Чтобы удалить узлы из сильно переполненного хроматина, потребуется активный процесс, который должен не только обеспечивать энергию для вывода системы из состояния топологического равновесия, но и направлять опосредованные топоизомеразой проходы таким образом, чтобы узлы были эффективно развязаны, а не образованы. делая узлы еще более сложными. Было показано, что процесс экструзии петли хроматина идеально подходит для активного развязывания волокон хроматина в интерфазных хромосомах.[31]
Хроматин: альтернативные определения
Термин, введенный Вальтер Флемминг, имеет несколько значений:
- Простое и лаконичное определение: Хроматин - это макромолекулярный комплекс макромолекулы ДНК и макромолекул белка (и РНК). Белки упаковывают и упорядочивают ДНК и контролируют ее функции в ядре клетки.
- Оперативное определение биохимиков: Хроматин - это комплекс ДНК / белок / РНК, экстрагированный из лизированных межфазных ядер эукариот. Какое из множества веществ, присутствующих в ядре, будет составлять часть извлеченного материала, частично зависит от метода, который использует каждый исследователь. Кроме того, состав и свойства хроматина варьируются от одного типа клеток к другому, во время развития определенного типа клеток и на разных стадиях клеточного цикла.
- В ДНК + гистон = хроматин определение: Двойная спираль ДНК в ядре клетки упакована специальными белками, называемыми гистонами. Образованный комплекс белок / ДНК называется хроматином. Основная структурная единица хроматина - нуклеосома.
Первое определение позволяет определять «хроматины» в других сферах жизни, таких как бактерии и археи, с использованием любых ДНК-связывающих белков, которые конденсирует молекулу. Эти белки обычно относят к нуклеоид-ассоциированные белки (НПД); примеры включают AsnC / LrpC с HU. Кроме того, некоторые археи действительно производят нуклеосомы из белков, гомологичных эукариотическим гистонам.[32]
Нобелевские премии
Следующие ученые были отмечены за их вклад в исследования хроматина с Нобелевские премии:
| Год | ВОЗ | Награда |
|---|---|---|
| 1910 | Альбрехт Коссель (Гейдельбергский университет) | Нобелевская премия по физиологии и медицине за открытие пяти ядерных баз: аденин, цитозин, гуанин, тимин, и урацил. |
| 1933 | Томас Хант Морган (Калифорнийский технологический институт) | Нобелевская премия по физиологии и медицине за его открытия роли гена и хромосомы в наследственности, основанные на его исследованиях белоглазой мутации у плодовой мушки Дрозофила.[33] |
| 1962 | Фрэнсис Крик, Джеймс Уотсон и Морис Уилкинс (Лаборатория молекулярной биологии MRC, Гарвардский и Лондонский университет соответственно) | Нобелевская премия по физиологии и медицине за открытие двойной спиральной структуры ДНК и ее значение для передачи информации в живом материале. |
| 1982 | Аарон Клуг (Лаборатория молекулярной биологии MRC) | Нобелевская премия по химии «За развитие кристаллографической электронной микроскопии и структурное объяснение биологически важных комплексов нуклеиновая кислота-белок» |
| 1993 | Ричард Дж. Робертс и Филип А. Шарп | Нобелевская премия по физиологии "за независимые открытия расщепление генов, "в которых участки ДНК называются экзоны экспрессируют белки и прерываются участками ДНК, называемыми интроны, которые не экспрессируют белки. |
| 2006 | Роджер Корнберг (Стэндфордский Университет) | Нобелевская премия по химии за открытие механизма, с помощью которого ДНК транскрибируется в информационную РНК. |
Смотрите также
Примечания
Рекомендации
- ^ Понедельник, Танмой (июль 2010 г.). «Характеристика содержания РНК в хроматине». Genome Res. 20 (7): 899–907. Дои:10.1101 / гр.103473.109. ЧВК 2892091. PMID 20404130.
- ^ Хансен, Джеффри (март 2012 г.). «Структура митотической хромосомы человека: что случилось с 30-нм волокном?». Журнал EMBO. 31 (7): 1621–1623. Дои:10.1038 / emboj.2012.66. ЧВК 3321215. PMID 22415369.
- ^ Дама, Р. (Май 2005 г.). «Роль нуклеоид-ассоциированных белков в организации и уплотнении бактериального хроматина». Молекулярная микробиология. 56 (4): 858–870. Дои:10.1111 / j.1365-2958.2005.04598.x. PMID 15853876. S2CID 26965112.
- ^ Bernstein BE, Mikkelsen TS, Xie X, Kamal M, Huebert DJ, Cuff J, Fry B, Meissner A, Wernig M, Plath K, Jaenisch R, Wagschal A, Feil R, Schreiber SL, Lander ES (апрель 2006 г.). «Двухвалентная структура хроматина отмечает ключевые гены развития в эмбриональных стволовых клетках». Клетка. 125 (2): 315–26. Дои:10.1016 / j.cell.2006.02.041. ISSN 0092-8674. PMID 16630819. S2CID 9993008.
- ^ Портосо М., Кавалли Дж. (2008). «Роль РНКи и некодирующих РНК в опосредованном Polycomb контроле экспрессии генов и геномного программирования». РНК и регуляция экспрессии генов: скрытый уровень сложности. Caister Academic Press. ISBN 978-1-904455-25-7.
- ^ Аннунциато, Энтони Т. «Упаковка ДНК: нуклеосомы и хроматин». Scitable. Природное образование. Получено 2015-10-29.
- ^ Робинзон DJ; Fairall L; Huynh VA; Родс Д. (апрель 2006 г.). «Электромагнитные измерения определяют размеры« 30-нм »хроматинового волокна: свидетельство компактной встречно-гребенчатой структуры». Труды Национальной академии наук Соединенных Штатов Америки. 103 (17): 6506–11. Bibcode:2006ПНАС..103.6506Р. Дои:10.1073 / pnas.0601212103. ЧВК 1436021. PMID 16617109.
- ^ Wong H, Victor JM, Mozziconacci J (сентябрь 2007 г.). Чен П. (ред.). "Полноатомная модель хроматинового волокна, содержащего гистоны-линкеры, выявляет универсальную структуру, настраиваемую длиной нуклеосомного повтора". PLoS ONE. 2 (9): e877. Bibcode:2007PLoSO ... 2..877Вт. Дои:10.1371 / journal.pone.0000877. ЧВК 1963316. PMID 17849006.

- ^ Никодеми М., Помбо А. (июнь 2014 г.). «Модели строения хромосом» (PDF). Curr. Мнение. Cell Biol. 28: 90–5. Дои:10.1016 / j.ceb.2014.04.004. PMID 24804566.
- ^ Никодеми М., Паннинг Б., Приско А. (май 2008 г.). «Термодинамический переключатель для колокализации хромосом». Генетика. 179 (1): 717–21. arXiv:0809.4788. Дои:10.1534 / генетика.107.083154. ЧВК 2390650. PMID 18493085.
- ^ Бон М., Херманн Д.В. (2010). «Цикл, управляемый диффузией, обеспечивает последовательную основу для организации хроматина». PLOS ONE. 5 (8): e12218. Bibcode:2010PLoSO ... 512218B. Дои:10.1371 / journal.pone.0012218. ЧВК 2928267. PMID 20811620.
- ^ Лодиш, Харви Ф. (2016). Молекулярная клеточная биология (8-е изд.). Нью-Йорк: В. Х. Фриман и компания. п. 339. ISBN 978-1-4641-8339-3.
- ^ Poonperm, R; Таката, H; Хамано, Т; Мацуда, А; Учияма, S; Хираока, Й; Фукуи, К. (1 июля 2015 г.). «Хромосомный каркас представляет собой двухцепочечную сборку белков каркаса». Научные отчеты. 5: 11916. Дои:10.1038 / srep11916. ЧВК 4487240. PMID 26132639.
- ^ Xing H, Vanderford NL, Sarge KD (ноябрь 2008 г.). «Митотический комплекс TBP-PP2A закладывает гены, предотвращая действие конденсина». Nat. Cell Biol. 10 (11): 1318–23. Дои:10.1038 / ncb1790. ЧВК 2577711. PMID 18931662.
- ^ Олфри В.Г., Фолкнер Р., Мирский А.Е. (май 1964 г.). «Ацетилирование и метилирование гистонов и их возможная роль в регуляции синтеза РНК». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 51 (5): 786–94. Bibcode:1964ПНАС ... 51..786А. Дои:10.1073 / пнас.51.5.786. ЧВК 300163. PMID 14172992.
- ^ Каочар С., Ту БП (ноябрь 2012 г.). «Стражи хроматина: небольшие метаболиты вызывают большие изменения в экспрессии генов». Trends Biochem. Наука. 37 (11): 477–83. Дои:10.1016 / j.tibs.2012.07.008. ЧВК 3482309. PMID 22944281.
- ^ Де Врис М., Рамос Л., Хаусин З., Де Бур П. (май 2012 г.). «Инициирование ремоделирования хроматина во время спермиогенеза человека». Биол Открытый. 1 (5): 446–57. Дои:10.1242 / bio.2012844. ЧВК 3507207. PMID 23213436.
- ^ Муругесапиллай Д., МакКоли М.Дж., Хо Р., Нельсон Холте М.Х., Степанянц А., Махер Л.Дж., Исраэлофф Н.Э., Уильямс М.С. (август 2014 г.). «Соединение ДНК и образование петель посредством HMO1 обеспечивает механизм стабилизации хроматина, свободного от нуклеосом». Исследования нуклеиновых кислот. 42 (14): 8996–9004. Дои:10.1093 / нар / gku635. ЧВК 4132745. PMID 25063301.
- ^ Murugesapillai D, McCauley MJ, Maher LJ, Williams MC (февраль 2017 г.). «Одномолекулярные исследования белков, изгибающих архитектурную ДНК группы B с высокой подвижностью». Биофизические обзоры. 9 (1): 17–40. Дои:10.1007 / s12551-016-0236-4. ЧВК 5331113. PMID 28303166.
- ^ Лю Б., Ип Р.К., Чжоу З. (2012). «Ремоделирование хроматина, восстановление повреждений ДНК и старение». Curr. Геномика. 13 (7): 533–47. Дои:10.2174/138920212803251373. ЧВК 3468886. PMID 23633913.
- ^ а б c d Селлоу Х., Лебопен Т., Шапюи С., Смит Р., Хегеле А., Сингх Х.Р., Козловски М., Бультманн С., Ladurner AG, Тимински Г., Хуэт С. (2016). «Поли (АДФ-рибоза) -зависимый ремоделер хроматина Alc1 индуцирует локальную релаксацию хроматина при повреждении ДНК». Мол. Биол. Клетка. 27 (24): 3791–3799. Дои:10.1091 / mbc.E16-05-0269. ЧВК 5170603. PMID 27733626.
- ^ а б Haince JF, McDonald D, Rodrigue A, Déry U, Masson JY, Hendzel MJ, Poirier GG (2008). «PARP1-зависимая кинетика рекрутирования белков MRE11 и NBS1 на множественные участки повреждения ДНК». J. Biol. Chem. 283 (2): 1197–208. Дои:10.1074 / jbc.M706734200. PMID 18025084.
- ^ а б c Rogakou EP, Pilch DR, Orr AH, Иванова VS, Боннер WM (1998). «Двухцепочечные разрывы ДНК вызывают фосфорилирование гистона H2AX по серину 139». J. Biol. Chem. 273 (10): 5858–68. Дои:10.1074 / jbc.273.10.5858. PMID 9488723.
- ^ Mailand N, Bekker-Jensen S, Faustrup H, Melander F, Bartek J, Lukas C, Lukas J (2007). «RNF8 убиквитилирует гистоны на двухцепочечных разрывах ДНК и способствует сборке белков репарации». Клетка. 131 (5): 887–900. Дои:10.1016 / j.cell.2007.09.040. PMID 18001824. S2CID 14232192.
- ^ Luijsterburg MS, Acs K, Ackermann L, Wiegant WW, Bekker-Jensen S, Larsen DH, Khanna KK, van Attikum H, Mailand N, Dantuma NP (2012). «Новая некаталитическая роль убиквитинлигазы RNF8 в раскрытии структуры хроматина более высокого порядка». EMBO J. 31 (11): 2511–27. Дои:10.1038 / emboj.2012.104. ЧВК 3365417. PMID 22531782.
- ^ Giresi, Paul G .; Ким, Чжонхван; McDaniell, Ryan M .; Iyer, Vishwanath R .; Либ, Джейсон Д. (2007-06-01). «FAIRE (Изоляция регуляторных элементов с помощью формальдегида) изолирует активные регуляторные элементы из хроматина человека». Геномные исследования. 17 (6): 877–885. Дои:10.1101 / гр.5533506. ISSN 1088-9051. ЧВК 1891346. PMID 17179217.
- ^ Галас, Д. Дж .; Шмитц, А. (1978-09-01). «ДНК-следы: простой метод определения специфичности связывания белок-ДНК». Исследования нуклеиновых кислот. 5 (9): 3157–3170. Дои:10.1093 / nar / 5.9.3157. ISSN 0305-1048. ЧВК 342238. PMID 212715.
- ^ Цуй, Кайронг; Чжао, Кэджи (01.01.2012). Общегеномные подходы к определению занятости нуклеосом у многоклеточных животных с использованием MNase-Seq. Методы молекулярной биологии. 833. С. 413–419. Дои:10.1007/978-1-61779-477-3_24. ISBN 978-1-61779-476-6. ISSN 1940-6029. ЧВК 3541821. PMID 22183607.
- ^ Буэнростро, Джейсон Д .; Giresi, Paul G .; Заба, Лиза С .; Chang, Howard Y .; Гринлиф, Уильям Дж. (01.12.2013). «Транспозиция нативного хроматина для быстрого и чувствительного эпигеномного профилирования открытого хроматина, ДНК-связывающих белков и положения нуклеосом». Методы природы. 10 (12): 1213–1218. Дои:10.1038 / nmeth.2688. ISSN 1548-7105. ЧВК 3959825. PMID 24097267.
- ^ Мечковски Дж., Кук А., Боуман С. К., Мюллер Б., Алвер Б. Х., Кунду С., Дитон А. М., Урбан Дж. А., Ларшан Е., Парк П. Дж., Кингстон Р. Э., Толсторуков М. Ю. (2016-05-06). «Титрование MNase выявляет различия между занятостью нуклеосом и доступностью хроматина». Nature Communications. 7: 11485. Bibcode:2016НатКо ... 711485M. Дои:10.1038 / ncomms11485. ЧВК 4859066. PMID 27151365.
- ^ Рако Д., Бенедетти Ф, Гундарулис Д., Стасиак А. (2018). «Экструзия петли хроматина и развязывание хроматина». Полимеры. 10 (10): 1126–1137. Дои:10.3390 / polym10101126. ЧВК 6403842. PMID 30961051.
- ^ Luijsterburg, Martijn S .; Уайт, Малькольм Ф .; ван Дриэль, Роэль; Дама, Ремус Тх. (8 января 2009 г.). «Основные архитекторы хроматина: архитектурные белки у бактерий, архей и эукариот». Критические обзоры в биохимии и молекулярной биологии. 43 (6): 393–418. Дои:10.1080/10409230802528488. PMID 19037758. S2CID 85874882.
- ^ «Томас Хант Морган и его наследие». Nobelprize.org. 7 сен 2012
Дополнительные источники
- Купер, Джеффри М. 2000. Клетка, 2-е издание, Молекулярный подход. Глава 4.2 Хромосомы и хроматин.
- Корсес, В. Г. (1995). «Хроматиновые изоляторы. Держим усилители под контролем». Природа. 376 (6540): 462–463. Bibcode:1995Натура 376..462C. Дои:10.1038 / 376462a0. PMID 7637775. S2CID 26494996.
- Cremer, T. 1985. Von der Zellenlehre zur Chromosomentheorie: Naturwissenschaftliche Erkenntnis und Theorienwechsel in der frühen Zell- und Vererbungsforschung, Veröffentlichungen aus der Forschungsstelle fürgerefische Pathologie. Springer-Vlg., Берлин, Гейдельберг.
- Элгин, С.С.Р. (ред.). 1995. Структура хроматина и экспрессия генов, vol. 9. IRL Press, Оксфорд, Нью-Йорк, Токио.
- Герасимова, Т. И .; Корсес, В. Г. (1996). «Граничные и изолирующие элементы в хромосомах». Curr. Мнение. Genet. Dev. 6 (2): 185–192. Дои:10.1016 / s0959-437x (96) 80049-9. PMID 8722175.
- Герасимова, Т. И .; Корсес, В. Г. (1998). «Белки группы Polycomb и Trithorax опосредуют функцию изолятора хроматина». Клетка. 92 (4): 511–521. Дои:10.1016 / s0092-8674 (00) 80944-7. PMID 9491892. S2CID 8192263.
- Герасимова, Т. И .; Корсес, В. Г. (2001). «ХРОМАТИНОВЫЕ ИЗОЛЯТОРЫ И ГРАНИЦЫ: ВЛИЯНИЕ НА транскрипцию и ядерную организацию». Анну Рев Жене. 35: 193–208. Дои:10.1146 / annurev.genet.35.102401.090349. PMID 11700282. S2CID 22738830.
- Герасимова, Т. И .; Byrd, K .; Корсес, В. Г. (2000). «Хроматиновый изолятор определяет ядерную локализацию ДНК [В процессе цитирования]». Mol Cell. 6 (5): 1025–35. Дои:10.1016 / с1097-2765 (00) 00101-5. PMID 11106742.
- Ha, S.C .; Lowenhaupt, K .; Rich, A .; Kim, Y.G .; Ким, К. К. (2005). «Кристаллическая структура соединения между B-ДНК и Z-ДНК выявляет два выдавленных основания». Природа. 437 (7062): 1183–6. Bibcode:2005Натура 437.1183H. Дои:10.1038 / природа04088. PMID 16237447. S2CID 2539819.
- Поллард Т. и У. Эрншоу. 2002. Клеточная биология. Сондерс.
- Сомвебер, Х. 1987. Расположение хромосом в межфазных клеточных ядрах, стр. 223-234. В W. Hennig (ed.), Structure and Function of Eucaryotic Chromosomes, vol. 14. Шпрингер-Верлаг, Берлин, Гейдельберг.
- Синден, Р. Р. (2005). «Молекулярная биология: повороты ДНК». Природа. 437 (7062): 1097–8. Дои:10.1038 / 4371097a. PMID 16237426. S2CID 4409092.
- Ван Холде К.Э. 1989. Хроматин. Нью-Йорк: Springer-Verlag. ISBN 0-387-96694-3.
- Ван Холде К., Дж. Златанова, Г. Аренц, Э. Мудрианакис. 1995. Элементы структуры хроматина: гистоны, нуклеосомы и волокна, с. 1-26. В С.С.Р. Элгина (ред.), Структура хроматина и экспрессия генов. IRL Press в Oxford University Press, Оксфорд.
внешняя ссылка
- Хроматин, гистоны и катепсин; PMAP Карта протеолиза -анимация
- [Последние публикации и новости о хроматине]
- Протокол для in vitro Сборка хроматина
- ENCODE Thread Explorer Паттерны хроматина в сайтах связывания факторов транскрипции. Природа (журнал)