РНК-полимераза - RNA polymerase
| ДНК-направленная РНК-полимераза | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 РНК-полимераза гетеро27мер, человек | |||||||||
| Идентификаторы | |||||||||
| Номер ЕС | 2.7.7.6 | ||||||||
| Количество CAS | 9014-24-8 | ||||||||
| Базы данных | |||||||||
| IntEnz | Просмотр IntEnz | ||||||||
| БРЕНДА | BRENDA запись | ||||||||
| ExPASy | Просмотр NiceZyme | ||||||||
| КЕГГ | Запись в KEGG | ||||||||
| MetaCyc | метаболический путь | ||||||||
| ПРИАМ | профиль | ||||||||
| PDB структуры | RCSB PDB PDBe PDBsum | ||||||||
| Генная онтология | AmiGO / QuickGO | ||||||||
| |||||||||

В молекулярная биология, РНК-полимераза (сокращенно RNAP или РНКапол, и официально ДНК-направленная РНК-полимераза), является фермент который синтезирует РНК из ДНК шаблон.
Использование фермента геликаза, RNAP локально открывает двухцепочечную ДНК, так что одна цепь экспонированного нуклеотиды может использоваться в качестве шаблона для синтеза РНК, процесса, называемого транскрипция. А фактор транскрипции и связанная с ней транскрипция посреднический комплекс должен быть прикреплен к Сайт связывания ДНК называется промоутер регион до того, как RNAP сможет инициировать раскручивание ДНК в этом положении. РНКП не только инициирует транскрипцию РНК, но и направляет нуклеотиды в нужное положение, облегчает прикрепление и удлинение, имеет встроенные возможности корректуры и замены, а также возможность распознавания прерывания. В эукариоты, RNAP может строить цепи длиной до 2,4 миллиона нуклеотидов.
RNAP производит РНК, которая функционально предназначена либо для белка кодирование, т.е. информационная РНК (мРНК); или некодирование (так называемые «гены РНК»). Существует по крайней мере четыре функциональных типа генов РНК:
- переносить РНК (тРНК) - переносит специфические аминокислоты к росту полипептид цепочки на рибосомальном участке синтез белка в течение перевод;
- рибосомная РНК (рРНК) - встраивается в рибосомы;
- микро РНК (miRNA) - регулирует активность генов; и,
- каталитическая РНК (рибозим ) - функционирует как ферментативно активная молекула РНК.
РНК-полимераза необходима для жизни и встречается у всех живых существ. организмы и много вирусы. В зависимости от организма РНК-полимераза может быть белковый комплекс (многосубъединичная RNAP) или состоит только из одной субъединицы (односубъединичная RNAP, ssRNAP), каждая из которых представляет независимую линию. Первый находится в бактерии, археи, и эукариоты одинаково, имея схожую основную структуру и механизм.[1] Последний находится в фаги а также эукариотические хлоропласты и митохондрии, и относится к современным ДНК-полимеразы.[2] Эукариотические и архейные РНКП имеют больше субъединиц, чем бактериальные, и контролируются по-разному.
Бактерии и археи имеют только одну РНК-полимеразу. У эукариот есть несколько типов ядерных РНКП, каждый из которых отвечает за синтез определенного подмножества РНК:
- РНК-полимераза I синтезирует пре-рРНК 45S (35S дюйм дрожжи ), который созревает и сформирует основные участки РНК рибосомы.
- РНК-полимераза II синтезирует предшественники мРНК и большинство мРНК и микроРНК.
- РНК-полимераза III синтезирует тРНК, рРНК 5S и другие малые РНК найдено в ядро и цитозоль.
- РНК-полимераза IV и V найденные в растениях менее изучены; Они делают миРНК. В дополнение к ssRNAP, хлоропласты также кодируют и используют бактериоподобную RNAP.
Структура


2006 год Нобелевская премия по химии был присужден Роджер Д. Корнберг для создания подробных молекулярных изображений РНК-полимеразы на различных этапах процесса транскрипции.[3]
В большинстве прокариоты, один вид РНК-полимеразы транскрибирует все типы РНК. «Ядро» РНК-полимеразы из Кишечная палочка состоит из пяти субъединиц: две альфа (α) субъединицы 36кДа, субъединица бета (β) размером 150 кДа, первичная субъединица бета (β ') размером 155 кДа и малая субъединица омега (ω). Фактор сигма (σ) связывается с ядром, образуя холофермент. После начала транскрипции фактор может развязаться и позволить ферменту продолжить свою работу.[4][5] Комплекс ядерной РНК-полимеразы образует структуру «клешня краба» или «зажим-челюсть» с внутренним каналом, проходящим по всей длине.[6] РНК-полимеразы эукариот и архей имеют схожую структуру ядра и работают аналогичным образом, хотя имеют много дополнительных субъединиц.[7]
Все RNAP содержат металл кофакторы, особенно цинк и магний катионы, которые помогают в процессе транскрипции.[8][9]
Функция
Контроль за процессом ген транскрипция влияет на паттерны экспрессия гена и, таким образом, позволяет ячейка адаптироваться к изменяющейся среде, выполнять особые роли в организме и поддерживать основные метаболические процессы, необходимые для выживания. Поэтому неудивительно, что активность РНКП длительная, сложная и строго регулируется. В кишечная палочка бактерии, более 100 факторы транскрипции были идентифицированы, которые модифицируют активность RNAP.[10]
RNAP может инициировать транскрипцию в определенных последовательностях ДНК, известных как промоутеры. Затем он производит цепь РНК, которая дополнительный к цепи ДНК-матрицы. Процесс добавления нуклеотиды к цепи РНК называется удлинением; у эукариот RNAP может строить цепочки длиной до 2,4 миллиона нуклеотиды (полная длина дистрофин ген). RNAP будет преимущественно высвобождать свой транскрипт РНК в определенных последовательностях ДНК, кодируемых на концах генов, которые известны как терминаторы.
Продукция RNAP включает:
- Посланник РНК (мРНК) - шаблон для синтез белков к рибосомы.
- Некодирующая РНК или «гены РНК» - широкий класс генов, кодирующих РНК, не транслируемую в белок. Наиболее яркими примерами генов РНК являются: переносить РНК (тРНК) и рибосомная РНК (рРНК), обе из которых участвуют в процессе перевод. Однако с конца 1990-х годов было обнаружено много новых генов РНК, и поэтому гены РНК могут играть гораздо более важную роль, чем считалось ранее.
- Передача РНК (тРНК) - специфичные для трансферов аминокислоты к росту полипептид цепи на рибосомальном участке синтеза белка во время перевод
- Рибосомная РНК (рРНК) - компонент рибосом
- Микро РНК - регулирует активность генов
- Каталитическая РНК (Рибозим ) - ферментативно активные молекулы РНК.
RNAP выполняет de novo синтез. Это возможно благодаря специфическим взаимодействиям с инициирующим нуклеотидом, которые жестко удерживают RNAP на месте, облегчая химическую атаку на поступающий нуклеотид. Такие специфические взаимодействия объясняют, почему RNAP предпочитает запускать транскрипты с АТФ (за которым следуют GTP, UTP и затем CTP). В отличие от ДНК-полимераза, RNAP включает геликаза активности, поэтому для раскрутки ДНК не требуется отдельный фермент.
Действие
Инициация
Связывание РНК-полимеразы в бактериях включает фактор сигма распознавание области корового промотора, содержащей элементы -35 и -10 (расположенные перед начало транскрибируемой последовательности), а также, на некоторых промоторах, С-концевой домен α-субъединицы, распознающий расположенные выше элементы промотора.[11] Существует несколько взаимозаменяемых сигма-факторов, каждый из которых распознает определенный набор промоторов. Например, в Кишечная палочка, σ70 экспрессируется в нормальных условиях и распознает промоторы генов, необходимых в нормальных условиях ("гены домашнего хозяйства "), а σ32 распознает промоторы генов, необходимые при высоких температурах ("гены теплового шока "). У архей и эукариот функции бактериального общего фактора транскрипции сигма выполняются множеством общие факторы транскрипции которые работают вместе. Замкнутый комплекс РНК-полимераза-промотор обычно называют "комплекс преинициации транскрипции."[12][13]
После связывания с ДНК РНК-полимераза переключается с закрытого комплекса на открытый. Это изменение включает разделение цепей ДНК с образованием развернутого участка ДНК размером примерно 13 п.н., называемого "пузырь транскрипции ". Суперспирализация играет важную роль в активности полимеразы из-за раскручивания и перемотки ДНК. Поскольку участки ДНК перед RNAP разматываются, возникают компенсаторные положительные суперспирали. Области за RNAP перемотаны, и присутствуют отрицательные суперспирали.[13]
Побег промоутера
Затем РНК-полимераза начинает синтезировать исходный гетеродуплекс ДНК-РНК с рибонуклеотидными основаниями, спаренными с цепью матричной ДНК в соответствии с взаимодействиями пар оснований Уотсона-Крика. Как отмечалось выше, РНК-полимераза контактирует с промоторной областью. Однако эти стабилизирующие контакты подавляют способность фермента получать доступ к ДНК дальше по течению и, таким образом, синтез полноразмерного продукта. Чтобы продолжить синтез РНК, РНК-полимераза должна ускользнуть от промотора. Он должен поддерживать контакты промотора, раскручивая более низкую ДНК для синтеза, "хруст" ДНК в инициирующий комплекс.[14] Во время перехода от промотора РНК-полимераза считается «промежуточным продуктом стресса». Термодинамически стресс накапливается в результате действий по раскручиванию ДНК и уплотнению ДНК. Как только гетеродуплекс ДНК-РНК становится достаточно длинным (~ 10 п.н.), РНК-полимераза освобождает свои вышестоящие контакты и эффективно достигает перехода от промотора к фазе элонгации. Гетеродуплекс в активном центре стабилизирует комплекс элонгации.
Однако побег промоутера - не единственный результат. РНК-полимераза также может снимать стресс, освобождая свои нижележащие контакты, останавливая транскрипцию. Приостановленный транскрибирующий комплекс имеет два варианта: (1) высвободить зарождающийся транскрипт и начать заново с промотора или (2) восстановить новый 3'OH на зарождающемся транскрипте в активном центре за счет каталитической активности РНК-полимеразы и возобновить скручивание ДНК для достижения побег промоутера. Неудачное начало, непродуктивный цикл РНК-полимеразы до перехода от промотора, приводит к образованию коротких фрагментов РНК длиной около 9 п.н. в процессе, известном как прерванная транскрипция. Степень прерванной инициации зависит от присутствия факторов транскрипции и силы промоторных контактов.[15]
Удлинение
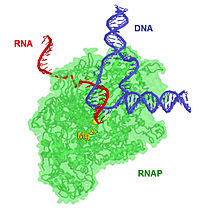
Транскрипционный комплекс длиной 17 пар оснований содержит гибрид ДНК-РНК длиной 8 пар оснований, то есть 8 пар оснований включают транскрипт РНК, связанный с цепью матрицы ДНК.[нужна цитата ] По мере развития транскрипции рибонуклеотиды добавляются к 3'-концу транскрипта РНК, и комплекс РНКП перемещается по ДНК. Характерные скорости удлинения у прокариот и эукариот составляют около 10–100 нт / сек.[16]
Аспартил (жерех ) остатки в RNAP будут удерживать Mg2+ ионы, которые, в свою очередь, будут координировать фосфаты рибонуклеотидов. Первый Mg2+ будет удерживать альфа-фосфат добавляемого NTP. Это позволяет осуществлять нуклеофильную атаку 3'OH из транскрипта РНК, добавляя еще один NTP к цепи. Второй Mg2+ будет удерживать пирофосфат NTP.[17] Общее уравнение реакции:
- (NMP)п + NTP → (NMP)п + 1 + ППя
верность
В отличие от механизмов корректуры ДНК-полимераза те из RNAP только недавно были исследованы. Вычитка начинается с отделения неправильно включенного нуклеотида от матрицы ДНК. Это приостанавливает транскрипцию. Затем полимераза возвращается на одну позицию и расщепляет динуклеотид, содержащий несовпадающий нуклеотид. В РНК-полимеразе это происходит в том же самом активном центре, который используется для полимеризации, и поэтому заметно отличается от ДНК-полимеразы, где проверка проводится в отдельном активном центре нуклеазы.[18]
Общий коэффициент ошибок составляет около 10−4 до 10−6.[19]
Прекращение
У бактерий прекращение транскрипции РНК может быть rho-зависимым или rho-независимым. Первый полагается на ро-фактор, который разрушает гетеродуплекс ДНК-РНК и вызывает высвобождение РНК.[20] Последний, также известный как внутреннее прекращение, полагается на палиндромную область ДНК. Транскрипция области вызывает образование структуры «шпильки» из петли транскрипции РНК и связывания с собой. Эта структура шпильки часто богата парами оснований G-C, что делает ее более стабильной, чем сам гибрид ДНК-РНК. В результате гибрид ДНК-РНК из 8 п.н. в транскрипционном комплексе переходит в гибрид из 4 п.н. Эти последние 4 пары оснований представляют собой слабые пары оснований A-U, и весь транскрипт РНК выпадет из ДНК.
Терминация транскрипции у эукариот менее изучена, чем у бактерий, но включает расщепление нового транскрипта с последующим независимым от матрицы добавлением аденинов на его новом 3'-конце в процессе, называемом полиаденилирование.[21]
Другие организмы
Учитывая, что ДНК и РНК-полимеразы осуществляют зависимую от матрицы нуклеотидную полимеризацию, можно было ожидать, что эти два типа ферментов будут структурно родственны. Однако, рентгеновский кристаллографический исследования обоих типов ферментов показывают, что кроме критического содержания Mg2+ ион на каталитическом сайте, они практически не связаны друг с другом; действительно, матрично-зависимые ферменты нуклеотидной полимеризации, по-видимому, независимо возникли дважды в течение ранней эволюции клеток. Одна линия привела к появлению современных ДНК-полимераз и обратных транскриптаз, а также к нескольким односубъединичным РНК-полимеразам (ssRNAP) из фагов и органелл.[2] Другая мультисубъединичная линия РНКП сформировала все современные клеточные РНК-полимеразы.[22][1]
Бактерии
В бактерии, тот же фермент катализирует синтез мРНК и некодирующая РНК (нкРНК).
РНКП - большая молекула. Основной фермент состоит из пяти субъединиц (~ 400 кДа ):[23]
- β ': субъединица β' является самой большой субъединицей и кодируется геном rpoC.[24] Субъединица β 'содержит часть активного центра, ответственного за синтез РНК, и некоторые из детерминант неспецифических взаимодействий с ДНК и возникающей РНК. В цианобактериях и хлоропластах он разделен на две субъединицы.[25]
- β: субъединица β является второй по величине субъединицей и кодируется rpoB ген. Субъединица β содержит оставшуюся часть активного центра, ответственного за синтез РНК, и остальные детерминанты для неспецифичных для последовательности взаимодействий с ДНК и возникающей РНК.
- α: субъединица α является третьей по величине субъединицей и присутствует в двух копиях на молекулу RNAP, αя и αII (один и два). Каждая субъединица α содержит два домена: αNTD (N-концевой домен) и αCTD (C-концевой домен). αNTD содержит детерминанты сборки RNAP. αCTD (С-концевой домен) содержит детерминанты для взаимодействия с промоторной ДНК, обеспечивая неспецифические для последовательности взаимодействия на большинстве промоторов и специфичные для последовательности взаимодействия на промоторах, содержащих вышестоящие элементы, и содержит детерминанты для взаимодействий с регуляторными факторами.
- ω: субъединица ω самая маленькая субъединица. Субъединица ω облегчает сборку RNAP и стабилизирует собранную RNAP.[26]
Для связывания промоторов ядро RNAP связывается с фактором инициации транскрипции. сигма (σ) с образованием холофермента РНК-полимеразы. Sigma снижает сродство RNAP к неспецифической ДНК, увеличивая при этом специфичность для промоторов, что позволяет транскрипции инициировать в правильных сайтах. Таким образом, полный холофермент состоит из 6 субъединиц: β'βαя и αIIωσ (~ 450 кДа).
Эукариоты

Эукариоты имеют несколько типов ядерных РНКП, каждый из которых отвечает за синтез определенного подмножества РНК. Все они структурно и механически связаны друг с другом и с бактериальным РНКП:
- РНК-полимераза I синтезирует пре-рРНК 45S (35S у дрожжей), который превращается в 28S, 18S и 5.8S рРНК, которые будут формировать основные участки РНК рибосома.[27]
- РНК-полимераза II синтезирует прекурсоры мРНК и большинство мяРНК и микроРНК.[28] Это наиболее изученный тип, и из-за высокого уровня контроля, необходимого для транскрипции, диапазон факторы транскрипции необходимы для его связывания с промоторами.
- РНК-полимераза III синтезирует тРНК, рРНК 5S и другие малые РНК найдено в ядро и цитозоль.[29]
- РНК-полимераза IV синтезирует миРНК в растениях.[30]
- РНК-полимераза V синтезирует РНК, участвующие в миРНК -направленный гетерохроматин образование у растений.[31]
Эукариотический хлоропласты содержат РНКП, очень похожую на бактериальную РНКП («пластид-кодируемая полимераза, PEP»). Они используют сигма-факторы, закодированные в ядерном геноме.[32]
Хлоропласт также содержит вторую, структурно и механически не связанную, одинарную субъединицу RNAP («кодируемая ядром полимераза, NEP»). Эукариотический митохондрии использовать POLRMT (человек), односубъединичная РНКП, кодируемая ядром.[2] Такие фагоподобные полимеразы в растениях называют RpoT.[32]
Археи
Археи имеют один тип РНКП, отвечающий за синтез всех РНК. Архейская RNAP структурно и механически подобна бактериальной RNAP и эукариотической ядерной RNAP I-V, и особенно тесно структурно и механически связана с эукариотической ядерной RNAP II.[7][33]История открытия РНК-полимеразы архей началась совсем недавно. Первый анализ RNAP архея был проведен в 1971 г., когда RNAP с крайнего галофил Halobacterium cutirubrum был выделен и очищен.[34] Кристаллические структуры РНКП из Sulfolobus solfataricus и Sulfolobus shibatae установить общее количество идентифицированных архейных субъединиц на уровне тринадцати.[7][35]
У архей есть субъединица, соответствующая эукариотическому Rpb1, разделенная на две части. Нет гомолога эукариотическому Rpb9 (POLR2I ) в S. shibatae комплекс, хотя TFS (гомолог TFIIS) был предложен как один, основанный на сходстве. Существует дополнительная субъединица, названная Rpo13; вместе с Rpo5 он занимает пространство, заполненное вставкой, обнаруженной в бактериальных β 'субъединицах (1,377–1,420 в Taq).[7] Более раннее исследование с более низким разрешением S. solfataricus структура не нашла Rpo13 и присвоила пространство только Rpo5 / Rpb5. Rpo3 примечателен тем, что это железо-серный белок. Субъединица AC40 RNAP I / III, обнаруженная у некоторых эукариот, имеет сходные последовательности,[35] но железо не связывает.[36] В любом случае этот домен выполняет структурную функцию.[37]
Субъединица RNAP архей ранее использовала номенклатуру «RpoX», где каждой субъединице присваивается буква, не связанная с какими-либо другими системами.[1] В 2009 г. была предложена новая номенклатура, основанная на нумерации субъединицы «Rpb» Pol II эукариот.[7]
Вирусы

Ортопоксвирусы и некоторые другие нуклеоцитоплазматические большие ДНК-вирусы синтезировать РНК, используя кодируемую вирусами мульти-субъединичную РНКП. Они наиболее похожи на эукариотические РНКП, но некоторые субъединицы минифицированы или удалены.[38] На какой именно RNAP они больше всего похожи, является предметом споров.[39] Большинство других вирусов, синтезирующих РНК, используют несвязанные механизмы.
Многие вирусы используют односубъединичную ДНК-зависимую РНКП (ssRNAP), которая структурно и механически связана с односубъединичной РНКП эукариотических хлоропластов (RpoT) и митохондрий (POLRMT ) и, более отдаленно, к ДНК-полимеразы и обратные транскриптазы. Пожалуй, наиболее широко изученная такая односубъединичная РНКП - это бактериофаг РНК-полимераза Т7. ssRNAP не могут быть проверены.[2]
Другие вирусы используют РНК-зависимая РНКП (RNAP, который использует РНК в качестве матрицы вместо ДНК). Это происходит в вирусы с отрицательной цепью РНК и дцРНК вирусы, оба из которых существуют в течение части своего жизненного цикла как двухцепочечные РНК. Однако некоторые вирусы с положительной цепью РНК, такие как полиовирус, также содержат РНК-зависимую РНКП.[40]
История
RNAP был открыт независимо Чарльзом Лоу, Одри Стивенс, и Джерард Гурвиц в 1960 г.[41] К этому времени половина 1959 г. Нобелевская премия в медицине был награжден Северо-Очоа за открытие того, что считалось RNAP,[42] но вместо этого оказалось полинуклеотидфосфорилаза.
Очищение
РНК-полимеразу можно выделить следующими способами:
- Автор фосфоцеллюлозная колонка.[43]
- К центрифугирование в градиенте глицерина.[44]
- Автор Колонка ДНК.
- Автор ионная хроматография столбец.[45]
А также комбинации вышеперечисленных приемов.
Смотрите также
Рекомендации
- ^ а б c d Вернер Ф., Громанн Д. (февраль 2011 г.). «Эволюция мультисубъединичных РНК-полимераз в трех областях жизни». Обзоры природы. Микробиология. 9 (2): 85–98. Дои:10.1038 / nrmicro2507. PMID 21233849. См. Также Cramer 2002: Крамер П. (2002). «Многосубъединичные РНК-полимеразы». Curr Opin Struct Biol. 12 (1): 89–97. Дои:10.1016 / s0959-440x (02) 00294-4. PMID 11839495.
- ^ а б c d Cermakian N, Ikeda TM, Miramontes P, Lang BF, Gray MW, Cedergren R (декабрь 1997 г.). «Об эволюции односубъединичных РНК-полимераз». Журнал молекулярной эволюции. 45 (6): 671–81. Bibcode:1997JMolE..45..671C. CiteSeerX 10.1.1.520.3555. Дои:10.1007 / PL00006271. PMID 9419244.
- ^ Нобелевская премия по химии 2006 г.
- ^ Гриффитс AJF, Миллер JH, Suzuki DT и др. Введение в генетический анализ. 7-е издание. Нью-Йорк: У. Х. Фриман; 2000. Глава 10.
- ^ Финн Р.Д., Орлова Е.В., Гоуэн Б., Бак М., Ван Хеель М. (декабрь 2000 г.). «Ядро РНК-полимеразы Escherichia coli и структуры холофермента». Журнал EMBO. 19 (24): 6833–44. Дои:10.1093 / emboj / 19.24.6833. ЧВК 305883. PMID 11118218.
- ^ Чжан Г., Кэмпбелл Е.А., Минахин Л., Рихтер С., Северинов К., Дарст С.А. (сентябрь 1999 г.). «Кристаллическая структура ядерной РНК-полимеразы Thermus aquaticus при разрешении 3,3 A». Клетка. 98 (6): 811–24. Дои:10.1016 / S0092-8674 (00) 81515-9. PMID 10499798.
- ^ а б c d е Корхин Ю., Unligil UM, Литтлфилд О., Нельсон П.Дж., Стюарт Д.И., Сиглер П.Б., Белл С.Д., Абрешия Н.Г. (май 2009 г.). «Эволюция сложных РНК-полимераз: полная структура РНК-полимеразы архей». PLOS Биология. 7 (5): e1000102. Дои:10.1371 / journal.pbio.1000102. ЧВК 2675907. PMID 19419240.
- ^ Альбертс Б. (18 ноября 2014 г.). Молекулярная биология клетки (Шестое изд.). Нью-Йорк, штат Нью-Йорк. ISBN 9780815344322. OCLC 887605755.
- ^ Марков Д., Нарышкина Т., Мустаев А., Северинов К. (сентябрь 1999 г.). «Сайт связывания цинка в самой большой субъединице ДНК-зависимой РНК-полимеразы участвует в сборке фермента». Гены и развитие. 13 (18): 2439–48. Дои:10.1101 / gad.13.18.2439. ЧВК 317019. PMID 10500100.
- ^ Исихама А (2000). «Функциональная модуляция РНК-полимеразы Escherichia coli». Ежегодный обзор микробиологии. 54: 499–518. Дои:10.1146 / annurev.micro.54.1.499. PMID 11018136.
- ^ ИнтерПро: IPR011260
- ^ Рёдер, Роберт Г. (1991). «Сложности инициации эукариотической транскрипции: регуляция сборки преинициационного комплекса». Тенденции в биохимических науках. 16 (11): 402–408. Дои:10.1016 / 0968-0004 (91) 90164-Q. ISSN 0968-0004. PMID 1776168.
- ^ а б Уотсон Дж. Д., Бейкер Т. А., Белл С. П., Ганн А. А., Левин М., Лосик Р. М. (2013). Молекулярная биология гена (7-е изд.). Пирсон.
- ^ Ревякин А., Лю С., Эбрайт Р.Х., Стрик Т.Р. (ноябрь 2006 г.). «Прерывистая инициация и продуктивная инициация с помощью РНК-полимеразы включают сжатие ДНК». Наука. 314 (5802): 1139–43. Bibcode:2006Научный ... 314.1139R. Дои:10.1126 / science.1131398. ЧВК 2754787. PMID 17110577.
- ^ Гольдман С.Р., Ebright RH, Nickels BE (май 2009 г.). «Прямое обнаружение абортивных транскриптов РНК in vivo». Наука. 324 (5929): 927–8. Bibcode:2009Sci ... 324..927G. Дои:10.1126 / science.1169237. ЧВК 2718712. PMID 19443781.
- ^ Майло Р., Филипс Р. «Клеточная биология в цифрах: что быстрее, транскрипция или перевод?». book.bionumbers.org. В архиве с оригинала 20 апреля 2017 г.. Получено 8 марта 2017.
- ^ Светлов В, Нудлер Э (январь 2013). «Основной механизм транскрипции РНК-полимеразой II». Biochimica et Biophysica Acta (BBA) - механизмы регуляции генов. 1829 (1): 20–8. Дои:10.1016 / j.bbagrm.2012.08.009. ЧВК 3545073. PMID 22982365.
- ^ Sydow JF, Cramer P (декабрь 2009 г.). «Верность РНК-полимеразы и проверка транскрипции» (PDF). Текущее мнение в структурной биологии. 19 (6): 732–9. Дои:10.1016 / j.sbi.2009.10.009. PMID 19914059.
- ^ Филипс Р., Майло Р. "Какова частота ошибок при транскрипции и переводе?". Получено 26 марта 2019.
- ^ Ричардсон JP (сентябрь 2002 г.). «Rho-зависимая терминация и АТФазы в терминации транскрипта». Biochimica et Biophysica Acta (BBA) - Структура и экспрессия гена. 1577 (2): 251–260. Дои:10.1016 / S0167-4781 (02) 00456-6. PMID 12213656.
- ^ Lykke-Andersen S, Jensen TH (октябрь 2007 г.). «Перекрывающиеся пути диктуют прекращение транскрипции РНК-полимеразы II». Биохимия. 89 (10): 1177–82. Дои:10.1016 / j.biochi.2007.05.007. PMID 17629387.
- ^ Стиллер Дж. У., Даффилд ЕС, Холл Б. Д. (сентябрь 1998 г.). «Амитохондриальные амебы и эволюция ДНК-зависимой РНК-полимеразы II». Труды Национальной академии наук Соединенных Штатов Америки. 95 (20): 11769–74. Bibcode:1998PNAS ... 9511769S. Дои:10.1073 / pnas.95.20.11769. ЧВК 21715. PMID 9751740.
- ^ Ebright RH (Декабрь 2000 г.). «РНК-полимераза: структурное сходство между бактериальной РНК-полимеразой и эукариотической РНК-полимеразой II». Журнал молекулярной биологии. 304 (5): 687–98. Дои:10.1006 / jmbi.2000.4309. PMID 11124018.
- ^ Монастырская Г.С., Губанов В.В., Гурьев С.О., Саломатина И.С., Шуваева Т.М., Липкин В.М., Свердлов Е.Д. (июль 1982 г.). «Первичная структура РНК-полимеразы E. coli, нуклеотидная последовательность гена rpoC и аминокислотная последовательность бета'-субъединицы». Исследования нуклеиновых кислот. 10 (13): 4035–44. Дои:10.1093 / nar / 10.13.4035. ЧВК 320776. PMID 6287430.
- ^ Бергсланд К.Дж., Хазелкорн Р. (июнь 1991 г.). «Эволюционные взаимоотношения между эубактериями, цианобактериями и хлоропластами: данные по гену rpoC1 штамма PCC 7120 Anabaena sp.». Журнал бактериологии. 173 (11): 3446–55. Дои:10.1128 / jb.173.11.3446-3455.1991. ЧВК 207958. PMID 1904436.
- ^ Мэтью Р., Чаттерджи Д. (октябрь 2006 г.). «История развития омега-субъединицы бактериальной РНК-полимеразы». Тенденции в микробиологии. 14 (10): 450–5. Дои:10.1016 / j.tim.2006.08.002. PMID 16908155.
- ^ Груммт I (1999). Регуляция транскрипции рибосомных генов млекопитающих с помощью РНК-полимеразы I. Прогресс в исследованиях нуклеиновых кислот и молекулярной биологии. 62. С. 109–54. Дои:10.1016 / S0079-6603 (08) 60506-1. ISBN 9780125400626. PMID 9932453.
- ^ Ли Й; Kim M; Хан Дж; Yeom KH; Lee S; Baek SH; Ким В.Н. (Октябрь 2004 г.). «Гены микроРНК транскрибируются РНК-полимеразой II». EMBO J. 23 (20): 4051–60. Дои:10.1038 / sj.emboj.7600385. ЧВК 524334. PMID 15372072.
- ^ Уиллис И.М. (Февраль 1993 г.). «РНК-полимераза III. Гены, факторы и специфичность транскрипции». Евро. J. Biochem. 212 (1): 1–11. Дои:10.1111 / j.1432-1033.1993.tb17626.x. PMID 8444147.
- ^ Herr AJ, Jensen MB, Dalmay T, Baulcombe, округ Колумбия (апрель 2005 г.). «РНК-полимераза IV направляет подавление эндогенной ДНК». Наука. 308 (5718): 118–20. Bibcode:2005Наука ... 308..118H. Дои:10.1126 / science.1106910. PMID 15692015.
- ^ Вежбицкий А.Т., Реам Т.С., Хааг-младший, Пикард С.С. (май 2009 г.). «Транскрипция РНК-полимеразы V направляет ARGONAUTE4 к хроматину». Природа Генетика. 41 (5): 630–4. Дои:10,1038 / нг.365. ЧВК 2674513. PMID 19377477.
- ^ а б Schweer J, Türkeri H, Kolpack A, Link G (декабрь 2010 г.). «Роль и регуляция пластидных сигма-факторов и их функциональных взаимодействующих факторов во время транскрипции хлоропластов - последние уроки Arabidopsis thaliana». Европейский журнал клеточной биологии. 89 (12): 940–6. Дои:10.1016 / j.ejcb.2010.06.016. PMID 20701995.
- ^ Вернер Ф (сентябрь 2007 г.). «Строение и функции РНК-полимераз архей». Молекулярная микробиология. 65 (6): 1395–404. Дои:10.1111 / j.1365-2958.2007.05876.x. PMID 17697097.
- ^ Луи Б.Г., Фитт П.С. (февраль 1971 г.). "Энзимология нуклеиновых кислот чрезвычайно галофильных бактерий. Halobacterium cutirubrum дезоксирибонуклеиновая кислота-зависимая полимераза рибонуклеиновой кислоты". Биохимический журнал. 121 (4): 621–7. Дои:10.1042 / bj1210621. ЧВК 1176638. PMID 4940048.
- ^ а б Хирата А., Кляйн Б.Дж., Мураками К.С. (февраль 2008 г.). «Рентгеновская кристаллическая структура РНК-полимеразы архей». Природа. 451 (7180): 851–4. Bibcode:2008Натура.451..851H. Дои:10.1038 / природа06530. ЧВК 2805805. PMID 18235446.
- ^ Фернандес-Торнеро К., Морено-Морсилло М., Рашид У. Дж., Тейлор Н. М., Руис Ф. М., Грюн Т., Легран П., Стойервальд Ю., Мюллер К. В. (октябрь 2013 г.). «Кристаллическая структура 14-субъединичной РНК-полимеразы I». Природа. 502 (7473): 644–9. Bibcode:2013Натура.502..644F. Дои:10.1038 / природа12636. PMID 24153184.
- ^ Дженнингс М.Э., Лесснер Ф.Х., Карр Е.А., Лесснер Д.Д. (февраль 2017 г.). «Кластеры [4Fe-4S] Rpo3 являются ключевыми детерминантами в образовании пост-Rpo3 / Rpo11 гетеродимера РНК-полимеразы в Methanosarcina acetivorans». МикробиологияOpen. 6 (1): e00399. Дои:10.1002 / mbo3.399. ЧВК 5300874. PMID 27557794.
- ^ Мирзаханян Ю., Гершон П.Д. (сентябрь 2017 г.). "Многосубъединичные ДНК-зависимые РНК-полимеразы вируса осповакцины и других нуклеоцитоплазматических вирусов большой ДНК: впечатления от эпохи структуры". Обзоры микробиологии и молекулярной биологии. 81 (3). Дои:10.1128 / MMBR.00010-17. ЧВК 5584312. PMID 28701329.
- ^ Гульельмини, Жюльен; Ву, Энтони С .; Крупович, Март; Фортер, Патрик; Гайя, Морган (10.09.2019). «Диверсификация гигантских и крупных эукариотических вирусов дцДНК предшествовала возникновению современных эукариот». Труды Национальной академии наук. 116 (39): 19585–19592. Дои:10.1073 / пнас.1912006116. ISSN 0027-8424. ЧВК 6765235. PMID 31506349.
- ^ Алквист П. (май 2002 г.). «РНК-зависимые РНК-полимеразы, вирусы и сайленсинг РНК». Наука. 296 (5571): 1270–3. Bibcode:2002Наука ... 296.1270A. Дои:10.1126 / science.1069132. PMID 12016304.
- ^ Гурвиц Дж (декабрь 2005 г.). «Открытие РНК-полимеразы». Журнал биологической химии. 280 (52): 42477–85. Дои:10.1074 / jbc.X500006200. PMID 16230341.
- ^ Нобелевская премия 1959 г.
- ^ Келли Дж. Л., Леман И. Р. (август 1986 г.). «Митохондриальная РНК-полимераза дрожжей. Очистка и свойства каталитической субъединицы». Журнал биологической химии. 261 (22): 10340–7. PMID 3525543.
- ^ Хонда А., Мукаигава Дж., Йокояма А., Като А., Уэда С., Нагата К., Кристал М., Наяк Д. П., Исихама А. (апрель 1990 г.). «Очистка и молекулярная структура РНК-полимеразы вируса гриппа A / PR8». Журнал биохимии. 107 (4): 624–8. Дои:10.1093 / oxfordjournals.jbchem.a123097. PMID 2358436.
- ^ Хагер Д.А., Джин DJ, Берджесс Р.Р. (август 1990 г.). «Использование ионообменной хроматографии высокого разрешения Mono Q для получения высокочистой и активной РНК-полимеразы Escherichia coli». Биохимия. 29 (34): 7890–4. Дои:10.1021 / bi00486a016. PMID 2261443.
внешняя ссылка
- ДНКи - DNA Interactive, включая информацию и флеш-ролики о РНК-полимеразе.
- РНК + полимераза в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)
- ЕС 2.7.7.6
- РНК-полимераза - синтез РНК из ДНК-матрицы
(Wayback Machine копия)
| Авторитетный контроль |
|---|