Фосфофруктокиназа 1 - Phosphofructokinase 1
| 6-фосфофруктокиназа | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 | |||||||||
| Идентификаторы | |||||||||
| Номер ЕС | 2.7.1.11 | ||||||||
| Количество CAS | 9001-80-3 | ||||||||
| Базы данных | |||||||||
| IntEnz | Просмотр IntEnz | ||||||||
| БРЕНДА | BRENDA запись | ||||||||
| ExPASy | Просмотр NiceZyme | ||||||||
| КЕГГ | Запись в KEGG | ||||||||
| MetaCyc | метаболический путь | ||||||||
| ПРИАМ | профиль | ||||||||
| PDB структуры | RCSB PDB PDBe PDBsum | ||||||||
| Генная онтология | AmiGO / QuickGO | ||||||||
| |||||||||
| Фосфофруктокиназа | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||||
| Символ | ПФК | ||||||||||
| Pfam | PF00365 | ||||||||||
| Pfam клан | CL0240 | ||||||||||
| ИнтерПро | IPR000023 | ||||||||||
| PROSITE | PDOC00336 | ||||||||||
| SCOP2 | 5пфк / Объем / СУПФАМ | ||||||||||
| |||||||||||
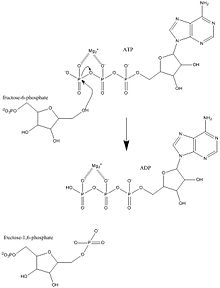
Фосфофруктокиназа-1 (ПФК-1) является одним из важнейших нормативных ферменты (EC 2.7.1.11 ) из гликолиз. Это аллостерический фермент состоит из 4-х подразделений и контролируется многими активаторы и ингибиторы. ПФК-1 катализирует важную «обязательную» стадию гликолиза - превращение фруктозо-6-фосфат и АТФ к 1,6-бисфосфату фруктозы и ADP. Гликолиз является основой дыхания, как анаэробного, так и аэробного. Поскольку фосфофруктокиназа (PFK) катализирует АТФ-зависимое фосфорилирование с превращением фруктозо-6-фосфата во фруктозо-1,6-бисфосфат и АДФ, это один из ключевых регулирующих этапов гликолиза. PFK способен регулировать гликолиз посредством аллостерического ингибирования, и, таким образом, клетка может увеличивать или уменьшать скорость гликолиза в ответ на энергетические потребности клетки. Например, высокое отношение АТФ к АДФ будет ингибировать PFK и гликолиз. Ключевое различие между регуляцией PFK у эукариот и прокариот состоит в том, что у эукариот PFK активируется 2,6-бисфосфатом фруктозы. Назначение фруктозо-2,6-бисфосфата - преодолеть ингибирование АТФ, что позволяет эукариотам иметь большую чувствительность к регуляции гормонами, такими как глюкагон и инсулин.[1]
| β-D-фруктозо-6-фосфат | Фосфофруктокиназа 1 | β-D-фруктозо-1,6-бисфосфат | |
 |  | ||
| АТФ | ADP | ||
| пя | ЧАС2О | ||
| Фруктозобисфосфатаза | |||
Структура
Млекопитающее PFK1 - 340 кД[2] тетрамер состоит из различных комбинаций трех типов субъединиц: мышца (М), печень (Земельные участки тромбоцит (П). Состав ПФК1 тетрамер различается в зависимости от типа ткани, в которой она присутствует. Например, зрелая мышца экспрессирует только M изофермент, следовательно, мышца PFK1 состоит исключительно из гомотетрамеров M4. Печень и почки экспрессируют преимущественно L-изоформу. В эритроциты субъединицы M и L случайным образом тетрамеризуются с образованием M4, L4 и трех гибридных форм фермента (ML3, M2L2, M3L). В результате кинетические и регуляторные свойства различных пулов изоферментов зависят от субъединичного состава. Тканевые изменения активности и изоферментов ПФК вносят значительный вклад в разнообразие гликолитический и глюконеогенный скорости, которые наблюдались для разных тканей.[3]
PFK1 представляет собой аллостерический фермент и имеет структуру, аналогичную структуре гемоглобин поскольку это димер димера.[4] Одна половина каждого димера содержит сайт связывания АТФ, тогда как другая половина сайта связывания субстрата (фруктозо-6-фосфат или (F6P)), а также отдельный аллостерический сайт связывания.[5]
Каждая субъединица тетрамера состоит из 319 аминокислот и состоит из двух доменов: один, который связывает субстрат АТФ, и другой, который связывает фруктозо-6-фосфат. Каждый домен представляет собой b-ствол и имеет цилиндрический b-лист, окруженный альфа-спиралями.
На противоположной стороне каждой субъединицы от каждого активного сайта находится аллостерический сайт на границе раздела между субъединицами в димере. За этот сайт конкурируют ATP и AMP. N-концевой домен играет каталитическую роль в связывании АТФ, а С-конец играет регуляторную роль. [6]
Механизм
PFK1 представляет собой аллостерический фермент, активность которого можно описать с помощью модель симметрии аллостеризма[7] посредством чего происходит согласованный переход от ферментативно неактивного Т-состояния к активному R-состоянию. F6P связывается с высоким сродством с ферментом в состоянии R, но не с ферментом в состоянии T. Для каждой молекулы F6P, которая связывается с PFK1, фермент постепенно переходит из состояния T в состояние R. Таким образом, график, отображающий активность PFK1 в зависимости от увеличения концентрации F6P, будет принимать сигмовидный форма кривой традиционно ассоциируется с аллостерическими ферментами.
PFK1 принадлежит к семейству фосфотрансферазы и он катализирует перенос γ-фосфата от АТФ на фруктозо-6-фосфат. PFK1 активный сайт содержит как сайты связывания ATP-Mg2 +, так и F6P. Некоторые предполагаемые остатки, участвующие в связывании субстрата в Кишечная палочка PFK1 включают Asp127 и Arg171.[8] В B. stearothermophilus PFK1, положительно заряженная боковая цепь остатка Arg162, образует водородный солевой мостик с отрицательно заряженной фосфатной группой F6P, взаимодействие, которое стабилизирует R-состояние относительно T-состояния и частично отвечает за гомотропный эффект связывания F6P. В состоянии T конформация фермента немного сдвигается, так что пространство, ранее занятое Arg162, заменяется на Glu161. Этот обмен положениями между соседними аминокислотными остатками подавляет способность F6P связывать фермент.
Аллостерические активаторы, такие как AMP и ADP связываются с аллостерическим сайтом, чтобы способствовать образованию состояния R, вызывая структурные изменения в ферменте. Аналогичным образом ингибиторы, такие как АТФ и PEP связываются с одним и тем же аллостерическим сайтом и способствуют образованию Т-состояния, тем самым ингибируя активность фермента.
Гидроксильный кислород углерода 1 оказывает нуклеофильную атаку на бета-фосфат АТФ. Эти электроны подталкиваются к ангидриду кислорода между бета- и гамма-фосфатами АТФ.[9][10]
Регулирование
PFK1 является наиболее важным контролирующим участком гликолитического пути млекопитающих. Этот шаг подлежит широкому регулированию, поскольку он не только экзергонический под физиологические условия, но также потому, что это обязательный шаг - первая необратимая реакция, уникальная для гликолитического пути. Это приводит к точному контролю уровня глюкозы и других моносахариды галактоза и фруктоза идет по гликолитическому пути. До реакции этого фермента глюкозо-6-фосфат потенциально может путешествовать по пентозофосфатный путь, или быть преобразованным в глюкозо-1-фосфат для гликогенез.
PFK1 - это аллостерически подавляется высоким уровнем АТФ но АМФ отменяет ингибирующее действие АТФ. Следовательно, активность фермента увеличивается при понижении клеточного отношения АТФ / АМФ. Таким образом, при падении энергетического заряда стимулируется гликолиз. PFK1 имеет два сайта с разным сродством к АТФ, который является субстрат и ингибитор.[2]
PFK1 также ингибируется низким уровнем pH, который усиливает ингибирующий эффект АТФ. PH падает, когда мышцы функционируют анаэробно и производство чрезмерного количества молочная кислота (хотя молочная кислота сама по себе не является причиной снижения pH[11]). Этот тормозящий эффект служит для защиты мышц от повреждений, которые могут возникнуть в результате накопления слишком большого количества кислоты.[2]
Наконец, PFK1 аллостерически ингибируется PEP, цитрат и АТФ. Фосфоенолпировиноградная кислота является продуктом ниже по потоку гликолитический путь. Хотя цитрат действительно накапливается, когда ферменты цикла Кребса достигают своей максимальной скорости, сомнительно, накапливается ли цитрат до концентрации, достаточной для ингибирования PFK-1 в нормальных физиологических условиях.[нужна цитата ]. Повышение концентрации АТФ указывает на избыток энергии и действительно имеет сайт аллостерической модуляции на PFK1, где он снижает сродство PFK1 к его субстрату.
PFK1 - это аллостерически активируется высокой концентрацией AMP, но самым сильным активатором является 2,6-бисфосфат фруктозы, который также производится из фруктозо-6-фосфата путем PFK2. Следовательно, обилие F6P приводит к более высокой концентрации 2,6-бисфосфат фруктозы (Ф-2,6-БП). Связывание F-2,6-BP увеличивает сродство PFK1 к F6P и снижает ингибирующий эффект АТФ. Это пример прямой стимуляции, поскольку гликолиз ускоряется при большом количестве глюкозы.[2]
Активность ПФК снижается за счет подавления синтеза за счет глюкагон. Глюкагон активирует протеинкиназа А что, в свою очередь, отключает киназную активность PFK2. Это отменяет любой синтез F-2,6-BP из F6P и, таким образом, деактивирует PFK1.
Точное регулирование PFK1 предотвращает гликолиз и глюконеогенез от происходящих одновременно. Однако есть цикл субстрата между F6P и F-1,6-BP. Фруктозо-1,6-бисфосфатаза (FBPase) катализирует гидролиз F-1,6-BP обратно до F6P, обратная реакция, катализируемая PFK1. Существует небольшая активность FBPase во время гликолиза и некоторая активность PFK1 во время глюконеогенеза. Этот цикл позволяет усиливать метаболические сигналы, а также выделять тепло за счет гидролиза АТФ.
Серотонин (5-HT) увеличивает PFK путем связывания с рецептором 5-HT (2A), вызывая фосфорилирование тирозинового остатка PFK с помощью фосфолипазы C. Это, в свою очередь, перераспределяет PFK в клетках скелетных мышц. Поскольку PFK регулирует гликолитический поток, серотонин играет регулирующую роль в гликолизе. [12]
Гены
У человека есть три гена фосфофруктокиназы:
Клиническое значение
Генетическая мутация в ПФКМ ген приводит к Болезнь Таруи, которое представляет собой болезнь накопления гликогена, при которой способность определенных типов клеток утилизировать углеводы как источник энергии нарушается.[13]
Болезнь Таруи - это болезнь накопления гликогена с симптомами, включая мышечную слабость (миопатию), спазмы и спазмы, вызванные физической нагрузкой, миоглобинурию (присутствие миоглобина в моче, указывающее на разрушение мышц) и компенсированный гемолиз. АТФ является природным аллостерическим ингибитором ПФК, предотвращающим ненужное производство АТФ посредством гликолиза. Однако мутация в Asp (543) Ala может приводить к более сильному ингибирующему эффекту АТФ (из-за повышенного связывания с ингибирующим аллостерическим сайтом связывания PFK).[14][15]
Мутация фосфофруктокиназы и рак: чтобы раковые клетки удовлетворяли свои потребности в энергии из-за быстрого роста и деления клеток, они выживают более эффективно, когда у них есть гиперактивный фермент фосфофруктокиназа 1.[16][17] Когда раковые клетки быстро растут и делятся, они изначально не имеют достаточного кровоснабжения и, следовательно, могут иметь гипоксию (кислородное голодание), и это запускает O-GlcNAцилирование по серину 529 ПФК. Эта модификация подавляет активность PFK1 и поддерживает пролиферацию рака, в отличие от мнения, что высокая активность PFK1 необходима для рака. Это может быть связано с перенаправлением потока глюкозы в пентозофосфатный путь для выработки НАДФН для детоксикации активных форм кислорода.[18]
Простой герпес типа 1 и фосфофруктокиназа: некоторые вирусы, включая ВИЧ, HCMV и Mayaro, влияют на клеточные метаболические пути, такие как гликолиз, путем зависимого от MOI увеличения активности PFK. Механизм, которым герпес увеличивает активность PFK, заключается в фосфорилировании фермента по остаткам серина. Гликолиз, индуцированный HSV-1, увеличивает содержание АТФ, что имеет решающее значение для репликации вируса.[19]
Смотрите также
- PFK2 (превращает фруктозо-6-фосфат во фруктозо-2,6-бисфосфат на одном участке или наоборот на другом участке)
- PFP (обратимо взаимно превращает фруктозо-6-фосфат и фруктозо-1,6-бисфосфат с использованием неорганических пирофосфат а не АТФ)
- Фруктозобисфосфатаза (гидролизует фруктозо-1,6-бисфосфат до фруктозо-6-фосфата)
Рекомендации
- ^ Усеник А., Легиша М. (ноябрь 2010 г.). Коби Б. (ред.). «Эволюция сайтов связывания аллостерического цитрата на 6-фосфофрукто-1-киназе». PLOS ONE. 5 (11): 677–683. Дои:10.1371 / journal.pone.0015447. ЧВК 2990764. PMID 21124851.
- ^ а б c d Страйер Л., Берг Дж. М., Тимочко Дж. Л. (2007). Биохимия (Шестое изд.). Сан-Франциско: W.H. Фримен. ISBN 978-0-7167-8724-2.
- ^ Данауэй Г.А., Кастен Т.П., Себо Т., Трапп Р. (май 1988 г.). «Анализ субъединиц и изоферментов фосфофруктокиназы в тканях человека». Biochem. J. 251 (3): 677–83. Дои:10.1042 / bj2510677. ЧВК 1149058. PMID 2970843.
- ^ PDB: 4пфк; Эванс П.Р., Фарранс Г.В., Хадсон П.Дж. (июнь 1981 г.). «Фосфофруктокиназа: структура и контроль». Философские труды Королевского общества B. 293 (1063): 53–62. Дои:10.1098 / рстб.1981.0059. PMID 6115424. Сложить резюме – Молекула месяца PDB.
- ^ Shirakihara Y, Evans PR (декабрь 1988 г.). «Кристаллическая структура комплекса фосфофруктокиназы Escherichia coli с продуктами ее реакции». J. Mol. Биол. 204 (4): 973–94. Дои:10.1016/0022-2836(88)90056-3. PMID 2975709.
- ^ Banaszak K, Mechin I, Obmolova G, Oldham M, Chang SH, Ruiz T., Radermacher M, Kopperschläger G, Rypniewski W (март 2011 г.). «Кристаллические структуры эукариотических фосфофруктокиназ из пекарских дрожжей и скелетных мышц кролика». Дж Мол Биол. 407 (7): 284–97. Дои:10.1016 / j.jmb.2011.01.019. PMID 21241708.
- ^ Песков К., Горянин И., Демин О. (август 2008 г.). «Кинетическая модель фосфофруктокиназы-1 из Escherichia coli». J Bioinform Comput Biol. 6 (4): 843–67. Дои:10.1142 / S0219720008003643. PMID 18763746.
- ^ Hellinga HW, Evans PR (1987). «Мутации в активном центре фосфофруктокиназы Escherichia coli». Природа. 327 (6121): 437–9. Дои:10.1038 / 327437a0. PMID 2953977.
- ^ Фонг В.Й., Лин В., Рао С.П., Дик Т., Алонсо С., Пете К. (август 2013 г.). Приход Т. (ред.). «Характеристика активности фосфофруктокиназы в Mycobacterium tuberculosis показывает, что поток функционального гликолитического углерода необходим для ограничения накопления токсичных промежуточных продуктов метаболизма в условиях гипоксии». PLOS ONE. 8 (2): 1198–206. Дои:10.1371 / journal.pone.0056037. ЧВК 3567006. PMID 23409118.
- ^ Папагианни М., Аврамидис Н. (май 2012 г.). «Разработка центральных путей в Lactococcus lactis: функциональная экспрессия генов фосфофруктокиназы (pfk) и альтернативной оксидазы (aox1) из Aspergillus niger в Lactococcus lactis способствует повышению скорости преобразования углерода в окислительных условиях». Ферментные и микробные технологии. 51 (113): 125–30. Дои:10.1016 / j.enzmictec.2012.04.007. PMID 22759530.
- ^ Линдингер, Майкл I .; Ковальчук, Джон М .; Хайгенхаузер, Джордж Дж. Ф. (1 сентября 2005 г.). «Применение физико-химических принципов к кислотно-щелочному статусу скелетных мышц». Американский журнал физиологии. Регуляторная, интегративная и сравнительная физиология. 289 (3): R891 – R894. Дои:10.1152 / ajpregu.00225.2005. ISSN 0363-6119. PMID 16105823.
- ^ Коэльо WS, Сола-Пенна M (январь 2013 г.). «Серотонин регулирует активность 6-фосфофрукто-1-киназы в сигнальном пути, зависимом от PLC-PKC-CaMK II и киназы Janus». Мол. Клетка. Биохим. 372 (1–2): 211–20. Дои:10.1007 / s11010-012-1462-0. PMID 23010892.
- ^ Накадзима Х., Рабен Н., Хамагути Т., Ямасаки Т. (март 2002 г.). «Дефицит фосфофруктокиназы: прошлое, настоящее и будущее». Curr. Мол. Med. 2 (2): 197–212. Дои:10.2174/1566524024605734. PMID 11949936.
- ^ Брюзер А., Кирхбергер Дж., Шенеберг Т. (октябрь 2012 г.). «Измененная аллостерическая регуляция мышечной 6-фосфофруктокиназы вызывает болезнь Таруи». Biochem Biophys Res Commun. 427 (1): 133–7. Дои:10.1016 / j.bbrc.2012.09.024. PMID 22995305.
- ^ Брюзер А., Кирхбергер Дж, Шёнеберг Т. (октябрь 2012 г.). «Измененная аллостерическая регуляция мышечной 6-фосфофруктокиназы вызывает болезнь Таруи». Biochem. Биофиз. Res. Сообщество. 427 (1): 133–7. Дои:10.1016 / j.bbrc.2012.09.024. PMID 22995305.
- ^ Гомес Л.С., Занкан П., Маркондес М.С., Рамос-Сантос Л., Мейер-Фернандес Дж. Р., Сола-Пенна М., Да Силва Д. (февраль 2013 г.). «Ресвератрол снижает жизнеспособность клеток рака груди и метаболизм глюкозы, ингибируя 6-фосфофрукто-1-киназу». Биохимия. 95 (6): 1336–43. Дои:10.1016 / j.biochi.2013.02.013. PMID 23454376.
- ^ Резюме Ваза, Алвес М.Г., Маркес Р., Морейра П.И., Оливейра П.Ф., Майя С.Дж., Сокорро С. (февраль 2013 г.). «Андроген-чувствительные и невосприимчивые клетки рака простаты представляют собой отчетливый профиль гликолитического метаболизма». Int J Biochem Cell Biol. 44 (11): 2077–84. Дои:10.1016 / j.biocel.2012.08.013. PMID 22964025.
- ^ Yi W, Clark PM, Mason DE, Keenan MC, Hill C, Goddard WA, Peters EC, Driggers EM, Hsieh-Wilson LC (август 2012 г.). «Гликозилирование фосфофруктокиназы 1 регулирует рост и метаболизм клеток». Наука. 337 (6097): 975–80. Дои:10.1126 / science.1222278. ЧВК 3534962. PMID 22923583.
- ^ Abrantes JL, Alves CM, Costa J, Almeida FC, Sola-Penna M, Fontes CF, Souza TM (август 2012 г.). «Простой герпес типа 1 активирует гликолиз за счет взаимодействия фермента 6-фосфофрукто-1-киназы (PFK-1)». Biochim Biophys Acta. 1822 (8): 1198–206. Дои:10.1016 / j.bbadis.2012.04.011. PMID 22542512.
внешняя ссылка
- Фосфофруктокиназа-1 в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)
- Proteopedia.org Фосфофруктокиназа
- Обзор всей структурной информации, доступной в PDB за UniProt: P08237 (АТФ-зависимая 6-фосфофруктокиназа, мышечный тип) на PDBe-KB.
- Обзор всей структурной информации, доступной в PDB за UniProt: Q01813 (АТФ-зависимая 6-фосфофруктокиназа, тип тромбоцитов) на PDBe-KB.
АТФ ADP АТФ ADP + + 2 ×  2 ×  2 × 3-фосфоглицерат 2 ×  2 × 2-фосфоглицерат 2 ×  2 × Фосфоенолпируват 2 ×  ADP АТФ 2 × Пируват 2 × |




