Ферментный катализ - Enzyme catalysis
Ферментный катализ увеличение ставка из процесс по биологическая молекула, "фермент ". Большинство ферментов являются белками, и большинство таких процессов представляют собой химические реакции. Внутри фермента катализ обычно происходит в локализованном месте, называемом активный сайт.
Наиболее ферменты состоят преимущественно из белков, либо одной белковой цепи, либо многих таких цепей в многосубъединичный комплекс. Ферменты часто также включают небелковые компоненты, такие как ионы металлов или специализированные органические молекулы, известные как кофактор (например. аденозинтрифосфат ). Многие кофакторы являются витаминами, и их роль как витаминов напрямую связана с их использованием в катализе биологических процессов метаболизма. Катализ из биохимические реакции в клетка жизненно важен, поскольку многие, но не все метаболически важные реакции в некаталитическом состоянии имеют очень низкую скорость. Одним из драйверов эволюции белка является оптимизация такой каталитической активности, хотя только самые важные ферменты работают вблизи пределов каталитической эффективности, а многие ферменты далеки от оптимального. Важными факторами в ферментативном катализе являются общий кислотный и основной катализ, орбитальное управление, энтропийное ограничение, эффекты ориентации (например, катализ с замком и ключом), а также двигательные эффекты, включающие динамика белка[1]
Механизмы ферментного катализа различаются, но все они в принципе аналогичны другим типам ферментов. химический катализ в том, что решающим фактором является снижение энергетического барьера (ов), отделяющего реагенты от продуктов.[2]. Уменьшение энергия активации (Eа) увеличивает долю молекул реагента, которые могут преодолеть этот барьер и образовать продукт. Важным принципом является то, что, поскольку они только уменьшают энергетические барьеры между продуктами и реагентами, ферменты всегда катализируют реакции в обоих направлениях и не могут продвигать реакцию вперед или влиять на положение равновесия - только на скорость, с которой она достигается. Как и в случае с другими катализаторами, фермент не расходуется и не изменяется в результате реакции (как субстрат), а рециркулируется, так что один фермент выполняет много циклов катализа.
Индуцированная посадка
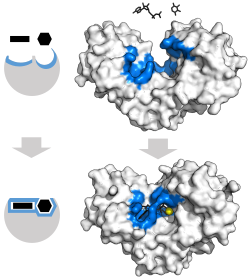
Классическая модель фермента -субстрат взаимодействие - это индуцированная модель соответствия.[3] Эта модель предполагает, что начальное взаимодействие между ферментом и субстратом относительно слабое, но эти слабые взаимодействия быстро вызывают конформационные изменения в ферменте, который усиливает связывание.
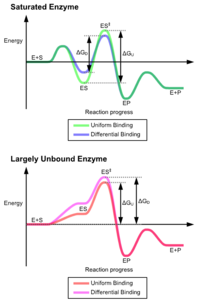
Преимущества механизма индуцированной подгонки возникают из-за стабилизирующего эффекта сильного связывания ферментов. Существует два различных механизма связывания субстрата: однородное связывание, которое имеет сильное связывание с субстратом, и дифференциальное связывание, которое имеет сильное связывание в переходном состоянии. Стабилизирующий эффект равномерного связывания увеличивает аффинность связывания как субстрата, так и переходного состояния, тогда как дифференциальное связывание увеличивает только аффинность связывания в переходном состоянии. Оба они используются ферментами и были выбраны эволюционным путем, чтобы минимизировать энергию активации реакции. Ферменты, которые являются насыщенными, то есть имеют высокое сродство связывания с субстратом, требуют дифференциального связывания для снижения энергии активации, тогда как небольшие несвязанные с субстратом ферменты могут использовать либо дифференциальное, либо равномерное связывание.[4]
Эти эффекты привели к тому, что большинство белков используют механизм дифференциального связывания для снижения энергии активации, поэтому большинство субстратов имеют высокое сродство к ферменту в переходном состоянии. Дифференциальное связывание осуществляется с помощью механизма индуцированной подгонки - субстрат сначала связывается слабо, затем фермент меняет конформацию, увеличивая сродство к переходному состоянию и стабилизируя его, таким образом уменьшая энергию активации для его достижения.
Однако важно уточнить, что концепция индуцированной подгонки не может использоваться для рационализации катализа. То есть химический катализ определяется как уменьшение Eа‡ (когда система уже находится в ES‡) относительно Eа‡ в некаталитической реакции в воде (без фермента). Индуцированная подгонка только предполагает, что барьер ниже в закрытой форме фермента, но не говорит нам, какова причина снижения барьера.
Индуцированная подгонка может быть полезна для точности молекулярного распознавания в присутствии конкуренции и шума через конформационная корректура механизм.[5]
Механизмы альтернативного пути реакции
Эти конформационные изменения также приносят каталитические остатки в активный сайт близко к химическим связям в субстрате, которые будут изменены в реакции. После связывания один или несколько механизмов катализа понижают энергию реакции переходное состояние, обеспечивая альтернативный химический путь для реакции. Существует шесть возможных механизмов катализа «через барьер», а также механизм «через барьер»:
Близость и ориентация
Взаимодействия фермент-субстрат выравнивают реактивные химические группы и удерживают их близко друг к другу в оптимальной геометрии, что увеличивает скорость реакции. Это снижает энтропия реагентов и, таким образом, делает реакции присоединения или переноса менее неблагоприятными из-за снижения общей энтропии, когда два реагента становятся одним продуктом. Однако это общий эффект, который наблюдается в реакциях без добавления или переноса, когда он возникает из-за увеличения «эффективной концентрации» реагентов. Это понимается при рассмотрении того, как увеличение концентрации приводит к увеличению скорости реакции: по существу, когда реагенты более концентрированы, они чаще сталкиваются и поэтому реагируют чаще. При ферментативном катализе связывание реагентов с ферментом ограничивает конформационное пространство реагентов, удерживая их в `` правильной ориентации '' и близко друг к другу, так что сталкиваются чаще и с правильной геометрией, чтобы облегчить желаемая реакция. «Эффективная концентрация» - это концентрация, которой реагент должен быть в свободном состоянии в растворе, чтобы испытать ту же частоту столкновений. Часто такие теоретические эффективные концентрации нефизичны и невозможно реализовать в реальности, что является свидетельством огромной каталитической силы многих ферментов с огромным увеличением скорости по сравнению с некаталитическим состоянием.
| Например: |
| Подобные реакции будут происходить намного быстрее, если реакция будет внутримолекулярной. |
 |
| Эффективная концентрация ацетат во внутримолекулярной реакции можно оценить как k2/ k1 = 2 х 105 Молярный. |
Однако ситуация может быть более сложной, поскольку современные компьютерные исследования установили, что традиционные примеры эффектов близости не могут быть напрямую связаны с энтропийными эффектами ферментов.[6][7][8] Кроме того, первоначальное энтропийное предложение[9] Было обнаружено, что в значительной степени переоценивается вклад ориентационной энтропии в катализ.[10]
Доноры или акцепторы протонов
Доноры и акцепторы протонов, т.е. кислоты и основание может отдавать и принимать протоны, чтобы стабилизировать развивающиеся заряды в переходном состоянии. Это связано с общим принципом катализа, принципом уменьшения энергетических барьеров, поскольку в целом переходные состояния являются состояниями с высокой энергией, и за счет их стабилизации эта высокая энергия снижается, понижая барьер. Ключевой особенностью ферментного катализа по сравнению с многими другими видами катализа небиологического происхождения является то, что и кислотный, и основной катализ могут быть объединены в одной реакции. Во многих абиотических системах кислоты (большие [H +]) или основания (стоки H + с большой концентрацией или разновидности с электронными парами) могут увеличивать скорость реакции; но, конечно, среда может иметь только один общий pH (показатель кислотности или щелочности (щелочности)). Однако, поскольку ферменты представляют собой большие молекулы, они могут позиционировать как кислотные группы, так и основные группы в своем активном центре для взаимодействия со своими субстратами и использовать оба режима независимо от общего pH.
Часто для активации используется общий кислотный или щелочной катализ. нуклеофил и / или электрофил группы или стабилизирующие уходящие группы. Многие аминокислоты с кислотными или основными группами используются в активном центре, такие как глутаминовая и аспарагиновая кислоты, гистидин, цистин, тирозин, лизин и аргинин, а также серин и треонин. Кроме того, часто используется пептидный каркас с карбонильными и амидными N-группами. Цистин и Гистидин очень часто задействованы, поскольку у них обоих есть pKa близко к нейтральному pH и поэтому может как принимать, так и отдавать протоны.
Многие механизмы реакции, включающие кислотно-основной катализ, предполагают существенно измененное значение pKa. Это изменение pKa возможно через локальное окружение остатка.[нужна цитата ].
| Условия | Кислоты | Базы |
|---|---|---|
| Гидрофобная среда | Увеличить pKa | Уменьшить pKa |
| Смежные остатки одинакового заряда | Увеличить pKa | Уменьшить pKa |
| Солевой мостик (и водород связь) образование | Уменьшить pKa | Увеличить pKa |
На pKa также может значительно влиять окружающая среда, до такой степени, что остатки, которые являются основными в растворе, могут действовать как доноры протонов, и наоборот.
| Например: |
| Каталитическая триада из сериновая протеаза |
 |
| Начальный этап каталитического механизма сериновой протеазы включает гистидин активного центра, принимающий протон от серинового остатка. Это подготавливает серин в качестве нуклеофила для атаки амидной связи субстрата. Этот механизм включает передачу протона от серина (основание, pKa 14) гистидину (кислота, pKa 6), что стало возможным благодаря локальному окружению оснований. |
Важно уточнить, что модификация pKa является чистой частью электростатического механизма.[11] Кроме того, каталитический эффект приведенного выше примера в основном связан с уменьшением pKa оксианиона и увеличением pKa гистидина, в то время как перенос протона от серина к гистидину не катализируется в значительной степени, поскольку он не барьер, определяющий скорость.[12]
Электростатический катализ
Стабилизация заряженных переходных состояний также может происходить за счет остатков в активном центре, образующих ионные связи (или взаимодействия частичного ионного заряда) с промежуточным продуктом. Эти связи могут происходить либо от кислый или же базовый боковые цепи найдены на аминокислоты Такие как лизин, аргинин, аспарагиновая кислота или же глютаминовая кислота или из металла кофакторы Такие как цинк. Ионы металлов особенно эффективны и могут снизить pKa воды настолько, чтобы сделать ее эффективным нуклеофилом.
Систематические исследования с компьютерным моделированием показали, что электростатические эффекты, безусловно, вносят наибольший вклад в катализ.[11] Это может увеличить скорость реакции до 10 раз.7.[13] В частности, было обнаружено, что фермент обеспечивает более полярную среду, чем вода, и что ионные переходные состояния стабилизируются фиксированными диполями. Это сильно отличается от стабилизации переходного состояния в воде, где молекулы воды должны платить «энергией реорганизации».[14] Для стабилизации ионного и заряженного состояний. Таким образом, катализ связан с тем, что полярные группы ферментов заранее организованы. [15]
Было показано, что величина электростатического поля, создаваемого активным центром фермента, сильно коррелирует с увеличением каталитической скорости фермента.[16][17]
Связывание субстрата обычно исключает воду из активного центра, тем самым понижая локальную диэлектрическую проницаемость до диэлектрической проницаемости органического растворителя. Это усиливает электростатические взаимодействия между заряженными / полярными подложками и активными центрами. Кроме того, исследования показали, что распределения заряда вокруг активных центров устроены таким образом, чтобы стабилизировать переходные состояния катализированных реакций. В некоторых ферментах эти распределения заряда, по-видимому, служат для направления полярных субстратов к их сайтам связывания, так что скорости этих ферментативных реакций превышают их очевидные контролируемые диффузией пределы.[нужна цитата ].
| Например: |
| Карбоксипептидаза каталитический механизм |
 |
| Тетраэдрический интермедиат стабилизируется частичной ионной связью между Zn2+ ион и отрицательный заряд на кислороде. |
Ковалентный катализ
Ковалентный катализ вовлекает субстрат, образующий временный Ковалентная связь с остатками в активном центре фермента или с кофактором. Это добавляет к реакции дополнительный ковалентный промежуточный продукт и помогает снизить энергию более поздних переходных состояний реакции. Ковалентная связь должна быть разорвана на более поздней стадии реакции для регенерации фермента. Этот механизм используется каталитическая триада ферментов, таких как протеазы подобно химотрипсин и трипсин, где образуется промежуточный ацил-фермент. Альтернативный механизм база шиффа формирование с использованием бесплатных амин из лизин остаток, как видно из фермента альдолаза в течение гликолиз.
Некоторые ферменты используют не аминокислоты кофакторы Такие как пиридоксальфосфат (PLP) или пирофосфат тиамина (TPP) с образованием ковалентных промежуточных продуктов с молекулами реагентов.[18][19] Такие ковалентные промежуточные соединения действуют для снижения энергии более поздних переходных состояний, подобно тому, как ковалентные промежуточные соединения, образованные с помощью аминокислотных остатков активного центра, обеспечивают стабилизацию, но возможности кофакторов позволяют ферментам проводить реакции, которые не могут выполнять только боковые аминокислотные остатки. Ферменты, использующие такие кофакторы, включают PLP-зависимый фермент аспартат трансаминаза и TPP-зависимый фермент пируватдегидрогеназа.[20][21]
Вместо снижения энергии активации пути реакции ковалентный катализ предоставляет альтернативу. путь для реакции (через ковалентное промежуточное соединение) и поэтому отличается от истинного катализа.[11] Например, энергетику ковалентной связи с молекулой серина в химотрипсине следует сравнивать с хорошо изученной ковалентной связью с нуклеофилом в реакции некаталитического раствора. Истинное предложение ковалентного катализа (где барьер ниже, чем соответствующий барьер в растворе) потребует, например, частичной ковалентной связи с переходным состоянием группой фермента (например, очень прочной водородной связи) и т. эффекты не вносят значительного вклада в катализ.
Катализ ионами металлов
Ион металла в активном центре участвует в катализе, координируя стабилизацию заряда и экранирование. Из-за положительного заряда металла только отрицательные заряды могут быть стабилизированы ионами металлов.[22] Однако ионы металлов выгодны в биологическом катализе, потому что на них не влияют изменения pH.[23] Ионы металлов также могут ионизировать воду, действуя как Кислота Льюиса.[24] Ионы металлов также могут быть агентами окисления и восстановления.[25]
Напряжение связи
Это основной эффект индуцированного связывания, когда сродство фермента к переходному состоянию больше, чем к самому субстрату. Это вызывает структурные перестройки, которые деформируют субстрат, связывая его в положение, более близкое к конформации переходного состояния, таким образом уменьшая разность энергий между субстратом и переходным состоянием и помогая катализировать реакцию.
Однако эффект деформации на самом деле является эффектом дестабилизации основного состояния, а не эффектом стабилизации переходного состояния.[11][26][страница нужна ] Кроме того, ферменты очень гибкие и не могут оказывать большого деформационного эффекта.[27]
Помимо напряжения связи в субстрате, внутри самого фермента также может индуцироваться напряжение связи для активации остатков в активном центре.
| Например: |
| Конформации субстрата, связанного субстрата и переходного состояния лизоцим. |
| Субстрат при связывании искажается из конформации полукресла гексозного кольца (из-за стерических препятствий с аминокислотами белка, вынуждающих экваториальный с6 находиться в осевом положении) в конформацию кресла[28][страница нужна ] |
Квантовое туннелирование
Этим традиционным механизмам «через барьер» в некоторых случаях бросали вызов модели и наблюдения за механизмами «сквозь барьер» (квантовое туннелирование ). Некоторые ферменты работают с кинетикой, которая быстрее, чем можно было бы предсказать с помощью классической ΔG‡. В моделях «сквозь барьер» протон или электрон могут туннелировать через активационные барьеры.[29][30] Квантовое туннелирование протонов наблюдалось в триптамин окисление ароматическая аминдегидрогеназа.[31]
Квантовое туннелирование, по-видимому, не дает большого каталитического преимущества, поскольку вклады туннелирования одинаковы в катализированных и некаталитических реакциях в растворе.[30][32][33][34] Однако туннельный вклад (обычно увеличивающий константы скорости в ~ 1000 раз)[31] по сравнению со скоростью реакции для классического пути «через барьер»), вероятно, имеет решающее значение для жизнеспособности биологических организмов. Это подчеркивает общую важность туннельных реакций в биологии.
В 1971-1972 годах была сформулирована первая квантово-механическая модель ферментативного катализа.[35][36][требуется сторонний источник ]
Активный фермент
Энергия связи комплекса фермент-субстрат не может рассматриваться как внешняя энергия, необходимая для активации субстрата. Фермент с высоким содержанием энергии может сначала передавать некоторую определенную энергетическую группу X1 от каталитического центра фермента до конечного места первого связанного реагента, затем другой группы X2 из второго связанного реагента (или из второй группы единственного реагента) должен быть перенесен в активный центр для завершения превращения субстрата в продукт и регенерации фермента.[37]
Мы можем представить всю ферментативную реакцию как две реакции сочетания:
(1)

(2)

Это видно из реакции (1), что группа X1 активного фермента появляется в продукте из-за возможности реакции обмена внутри фермента, чтобы избежать как электростатического ингибирования, так и отталкивания атомов. Таким образом, мы представляем активный фермент как мощный реагент ферментативной реакции. Реакция (2) показывает неполное превращение субстрата, поскольку его группа X2 остается внутри фермента. Этот подход как идея ранее предлагался, полагаясь на гипотетически чрезвычайно высокие ферментативные превращения (каталитически совершенный фермент).[38]
Решающим моментом для проверки настоящего подхода является то, что катализатор должен быть комплексом фермента с передающей группой реакции. Этот химический аспект подтверждается хорошо изученными механизмами нескольких ферментативных реакций. Рассмотрим реакцию гидролиза пептидной связи, катализируемую чистым белком α-химотрипсином (фермент, действующий без кофактора), который является хорошо изученным членом семейства сериновых протеаз, см.[39]
Мы представляем экспериментальные результаты для этой реакции в виде двух химических этапов:
(3)

(4)

где S1 полипептид, P1 и P2 продукты. Первая химическая ступень (3) включает образование ковалентного промежуточного ацилфермента. Второй шаг (4) - стадия деацилирования. Важно отметить, что группа H +, изначально обнаруженная на ферменте, но не в воде, появляется в продукте до стадии гидролиза, поэтому ее можно рассматривать как дополнительную группу ферментативной реакции.
Таким образом, реакция (3) показывает, что фермент действует как мощный реагент реакции. Согласно предложенной концепции, транспорт H от фермента способствует первому превращению реагента, разрыву первой начальной химической связи (между группами P1 и P2). Стадия гидролиза приводит к разрыву второй химической связи и регенерации фермента.
Предлагаемый химический механизм не зависит от концентрации субстратов или продуктов в среде. Однако изменение их концентрации в основном вызывает изменения свободной энергии на первой и конечной стадиях реакций (1) и (2) из-за изменений содержания свободной энергии каждой молекулы, будь то S или P, в водном растворе. Этот подход соответствует следующему механизму сокращения мышц. Заключительным этапом гидролиза АТФ в скелетных мышцах является высвобождение продукта, вызванное ассоциацией миозиновых головок с актином.[40] Закрытие актин-связывающей щели во время реакции ассоциации структурно связано с открытием нуклеотид-связывающего кармана на активном сайте миозина.[41]
Примечательно, что заключительные стадии гидролиза АТФ включают быстрое высвобождение фосфата и медленное высвобождение АДФ.[42][43]Высвобождение фосфатного аниона из связанного аниона АДФ в водный раствор можно рассматривать как экзергоническую реакцию, поскольку фосфатный анион имеет низкую молекулярную массу.
Таким образом, мы приходим к выводу, что первичное выделение неорганического фосфата H2PO4− приводит к преобразованию значительной части свободной энергии гидролиза АТФ в кинетическую энергию сольватированного фосфата, вызывая активное течение. Это предположение о локальной механохимической трансдукции согласуется с механизмом сокращения мышц Тироша, в котором сила мышц возникает в результате интегрированного действия активного потока, создаваемого гидролизом АТФ.[44][45]
Примеры каталитических механизмов
На самом деле, большинство ферментных механизмов включает комбинацию нескольких различных типов катализа.
Триозофосфатизомераза
Триозофосфатизомераза (EC 5.3.1.1 ) катализирует обратимое взаимопревращение двух триоза фосфаты изомеры дигидроксиацетонфосфат и D-глицеральдегид-3-фосфат.
Трипсин
Трипсин (EC 3.4.21.4 ) это сериновая протеаза что раскалывает белок субстраты после лизин или же аргинин остатки с использованием каталитическая триада для выполнения ковалентного катализа и оксианионная дыра для стабилизации накопления заряда на переходные состояния.
Альдолаза
Альдолаза (EC 4.1.2.13 ) катализирует распад фруктозо-1,6-бисфосфат (Ф-1,6-БП) в глицеральдегид-3-фосфат и дигидроксиацетонфосфат (DHAP ).
Коэффициент диффузии ферментов
Появление исследования одиночных молекул привели в 2010-х годах к наблюдению, что движение несвязанных ферментов увеличивается с увеличением концентрации субстрата и увеличением энтальпия реакции.[46] Последующие наблюдения показывают, что это увеличение диффузионность обусловлено временным перемещением фермента центр массы, что приводит к «эффекту отдачи, который приводит в движение фермент».[47]
Сходство реакции
Сходство ферментативных реакций (EC ) можно рассчитать, используя изменения связей, реакционные центры или метрики субструктуры (EC-BLAST ).[48]
Смотрите также
- Каталитическая триада
- Ферментный анализ
- Кинетика ферментов
- Ферментная распущенность
- Белковая динамика
- Псевдоферменты, чья повсеместность, несмотря на их каталитическую неактивность, предполагает omic подразумеваемое
- Квантовое туннелирование
- Карта протеолиза
- Кристаллография с временным разрешением
Рекомендации
- ^ Камерлин, С. С .; Варшел, А (2010). «На заре 21 века: динамика - недостающее звено для понимания ферментативного катализа?». Белки: структура, функции и биоинформатика. 78 (6): 1339–75. Дои:10.1002 / prot.22654. ЧВК 2841229. PMID 20099310.
- ^ Шринивасан, Бхарат (27 сентября 2020 г.). «Совет: обучение кинетике ферментов». Журнал FEBS. Дои:10.1111 / фев.15537. ISSN 1742-464X.
- ^ Кошланд DE (Февраль 1958 г.). «Применение теории ферментной специфичности к синтезу белков». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 44 (2): 98–104. Bibcode:1958ПНАС ... 44 ... 98К. Дои:10.1073 / пнас.44.2.98. ЧВК 335371. PMID 16590179.

- ^ Anslyn, E.V .; Догерти, Д.А. (2006). Современная физико-органическая химия. Книги университетских наук. ISBN 978-1-891389-31-3.
- ^ Савир Y; Тласти Т (2007). Скалас, Энрико (ред.). «Конформационная корректура: влияние конформационных изменений на специфику молекулярного распознавания» (PDF). PLOS One. 2 (5): e468. Bibcode:2007PLoSO ... 2..468S. Дои:10.1371 / journal.pone.0000468. ЧВК 1868595. PMID 17520027. Архивировано из оригинал (PDF) 14 мая 2011 г.. Получено 22 августа 2010.
- ^ Stanton, R.V .; Perakyla, M .; Bakowies, D .; Коллман, П.А. (1998). «Комбинированные расчеты ab initio и свободной энергии для изучения реакций в ферментах и растворах: гидролиз амидов в трипсине и водном растворе». Варенье. Chem. Soc. 120 (14): 3448–3457. Дои:10.1021 / ja972723x.
- ^ Kuhn, B .; Коллман, П.А. (2000). "QM-FE и расчеты молекулярной динамики катехол-O-метилтрансферазы: свободная энергия активации в ферменте и в водном растворе и региоселективность реакции, катализируемой ферментом". Варенье. Chem. Soc. 122 (11): 2586–2596. Дои:10.1021 / ja992218v.
- ^ Bruice, T.C .; Лайтстоун, футбольный клуб (1999). «Основное состояние и переходное состояние вкладов в скорость внутримолекулярных и ферментативных реакций». Соотв. Chem. Res. 32 (2): 127–136. Дои:10.1021 / ar960131y.
- ^ Page, M.I .; Дженкс, В. (1971). «Энтропийный вклад в скорость ускорения ферментативных и внутримолекулярных реакций и хелатный эффект». Proc. Natl. Акад. Sci. Соединенные Штаты Америки. 68 (8): 1678–1683. Bibcode:1971ПНАС ... 68.1678П. Дои:10.1073 / pnas.68.8.1678. ЧВК 389269. PMID 5288752.
- ^ Warshel, A .; Парсон, W.W. (2001). «Динамика биохимических и биофизических реакций: взгляд на компьютерное моделирование». Кварта. Rev. Biophys. 34 (4): 563–679. Дои:10,1017 / с0033583501003730. PMID 11852595.
- ^ а б c d Warshel, A .; Шарма, П.К .; Като, М .; Xiang, Y .; Liu, H .; Olsson, M.H.M. (2006). «Электростатические основы ферментного катализа». Chem. Rev. 106 (8): 3210–3235. Дои:10.1021 / cr0503106. PMID 16895325.
- ^ Warshel, A .; Naray-Szabo, G .; Sussman, F .; Хван, Дж .-К. (1989). «Как на самом деле работают сериновые протеазы?». Биохимия. 28 (9): 3629–37. Дои:10.1021 / bi00435a001. PMID 2665806.
- ^ Воет, Дональд. (коп. 2011). Биохимия. Джон Вили и сыновья. OCLC 808679090. Проверить значения даты в:
| дата =(помощь) - ^ Маркус, Р. А. (1965). "К теории реакций с переносом электрона. VI. Единый подход к гомогенным и электродным реакциям" (PDF). J. Chem. Phys. 43 (2): 679–701. Bibcode:1965ЖЧФ..43..679М. Дои:10.1063/1.1696792.
- ^ Варшел, А (1978). «Энергетика ферментного катализа». Proc. Natl. Акад. Sci. Соединенные Штаты Америки. 75 (11): 5250–5254. Bibcode:1978PNAS ... 75,5 250 Вт. Дои:10.1073 / пнас.75.11.5250. ЧВК 392938. PMID 281676.
- ^ «Как работают ферменты»
- ^ «ЭКСТРЕМАЛЬНЫЙ КАТАЛИЗ МОЩНОСТИ ЭЛЕКТРИЧЕСКИХ ПОЛЕЙ В АКТИВНОМ САЙТЕ КЕТОСТЕРОИДНОЙ ИЗОМЕРАЗЫ», [1]
- ^ Тони, М. Д. "Специфичность реакции в пиридоксальных ферментах". Архив биохимии и биофизики (2005) 433: 279-287
- ^ Информационный центр по микронутриентам, Университет штата Орегон
- ^ Воет, Дональд; Джудит Воет (2004). Биохимия. John Wiley & Sons Inc., стр.986–989. ISBN 978-0-471-25090-6.
- ^ Воет, Дональд; Джудит Воет (2004). Биохимия. John Wiley & Sons Inc., стр.604–606. ISBN 978-0-471-25090-6.
- ^ Piccirilli, Joseph A .; Vyle, Joseph S .; Карутерс, Марвин Х .; Чех, Томас Р. (7 января 1993 г.). «Катализ ионов металлов в рибозимной реакции Tetrahymena». Природа. 361 (6407): 85–88. Bibcode:1993Натура 361 ... 85П. Дои:10.1038 / 361085a0. PMID 8421499. S2CID 4326584.
- ^ БЕНДЕР, МИРОН Л. (1 января 1962 г.). «Металлический ионный катализ нуклеофильных органических реакций в растворе». Реакции координированных лигандов. Успехи химии. 37. АМЕРИКАНСКОЕ ХИМИЧЕСКОЕ ОБЩЕСТВО. С. 19–36. Дои:10.1021 / ba-1963-0037.ch002. ISBN 978-0841200388.
- ^ Файф, Томас Х .; Пржистас, Теодор Дж. (1 февраля 1985 г.). «Катализ ионов двухвалентного металла при гидролизе сложных эфиров пиколиновой кислоты. Ион металла способствует реакциям, катализируемым ионами гидроксида и водой». Журнал Американского химического общества. 107 (4): 1041–1047. Дои:10.1021 / ja00290a048. ISSN 0002-7863.
- ^ Штадтман, Э. Р. (1 января 1990 г.). «Катализируемое ионами металлов окисление белков: биохимический механизм и биологические последствия». Свободная радикальная биология и медицина. 9 (4): 315–325. Дои:10.1016/0891-5849(90)90006-5. ISSN 0891-5849. PMID 2283087.
- ^ Дженкс, Уильям П. (1987) [1969]. Катализ в химии и энзимологии. Серия Макгроу-Хилла по продвинутой химии (переиздание). Нью-Йорк: Dover Publications. ISBN 9780486654607.
- ^ Warshel, A .; Левитт, М. (1976). «Теоретические исследования ферментативных реакций: диэлектрическая электростатическая и стерическая стабилизация иона карбония в реакции лизоцима». Журнал молекулярной биологии. 103 (2): 227–49. Дои:10.1016/0022-2836(76)90311-9. PMID 985660.
- ^ <основы биохимии Voet, Voet and Pratt 4th edition>, которая по форме похожа на переходное состояние.
- ^ Гарсия-Вилока, М; Гао, Дж; Карплюс, М; Трулар, Д.Г. (2004). «Как работают ферменты: анализ с помощью современной теории скорости и компьютерного моделирования». Наука. 303 (5655): 186–95. Bibcode:2004Наука ... 303..186Г. Дои:10.1126 / science.1088172. PMID 14716003. S2CID 17498715.
- ^ а б Olsson, MH; Зигбан, ЧП; Варшел, А (2004). «Моделирование большого кинетического изотопного эффекта и температурной зависимости переноса атома водорода в липоксигеназе». Журнал Американского химического общества. 126 (9): 2820–8. Дои:10.1021 / ja037233l. PMID 14995199.
- ^ а б Масграу, L; Ружейникова, А; Johannissen, LO; Hothi, P; Басран, Дж; Ранаган, KE; Малхолланд, AJ; Сатклифф, MJ; и другие. (2006). «Атомное описание ферментативной реакции, в которой преобладает туннелирование протонов». Наука. 312 (5771): 237–41. Bibcode:2006Научный ... 312..237М. Дои:10.1126 / science.1126002. PMID 16614214. S2CID 27201250.
- ^ Hwang, J.-K .; Варшел, А. (1996). «Насколько важны квантово-механические движения ядер в ферментативном катализе». Варенье. Chem. Soc. 118 (47): 11745–11751. Дои:10.1021 / ja962007f.
- ^ Болл, П. (2004). «Ферменты: случайно или намеренно?». Природа. 431 (7007): 396–397. Bibcode:2004Натура 431..396Б. Дои:10.1038 / 431396a. PMID 15385982. S2CID 228263.
- ^ Olsson, M.H.M .; Parson, W.W .; Варшел, А. (2006). «Динамический вклад в ферментный катализ: критические проверки популярной гипотезы». Chem. Rev. 106 (5): 1737–1756. Дои:10.1021 / cr040427e. PMID 16683752.
- ^ Волькенштейн М.В., Догонадзе Р.Р., Мадумаров А.К., Урушадзе З.Д., Харкац Ю.И. Теория ферментного катализа.- Молекулярная биология, Москва, 6, 1972, 431-439.
- ^ Волькенштейн М.В., Догонадзе Р.Р., Мадумаров А.К., Урушадзе З.Д., Харкац Ю.И. Электронные и конформационные взаимодействия в ферментативном катализе. В: E.L. Андроникашвили (Ред.), Конфигурационные изменения Биополимеров в Растворах, Издательство "Наука", Москва, 1973, 153-157
- ^ Фойгель, Александр Г. (2011). «Является ли фермент мощным реагентом биохимической реакции?». Молекулярная и клеточная биохимия. 352 (1–2): 87–9. Дои:10.1007 / s11010-011-0742-4. PMID 21318350. S2CID 11133081.
- ^ Фогель, А.Г. (1982). «Кооперативность ферментативных реакций и молекулярные аспекты преобразования энергии». Мол. Клетка. Биохим. 47 (1): 59–64. Дои:10.1007 / bf00241567. PMID 7132966. S2CID 21790380.
- ^ Hengge, AC; Стейн, Р.Л. (2004). «Роль конформационной подвижности белков в ферментативном катализе: ацилирование альфа-химотрипсина специфическими пептидными субстратами». Биохимия. 43 (3): 742–747. Дои:10.1021 / bi030222k. PMID 14730979.
- ^ Lymn, RW; Тейлор, EW. (1971). «Механизм гидролиза аденозинтрифосфата актомиозином». Биохимия. 10 (25): 4617–4624. Дои:10.1021 / bi00801a004. PMID 4258719.
- ^ Холмс, KC; Ангерт, I; Кулл, Ф.Г .; Ян, Вт; Шредер, Р. (2003). «Электронная криомикроскопия показывает, насколько сильное связывание миозина с актином высвобождает нуклеотид». Природа. 425 (6956): 423–427. Bibcode:2003Натура.425..423H. Дои:10.1038 / nature02005. PMID 14508495. S2CID 2686184.
- ^ Симанковский, РФ; Wiseman, Миссури; Белый, HD. (1985). «Диссоциация АДФ из субфрагмента 1 актомиозина является достаточно медленной, чтобы ограничить ненагруженную скорость укорочения в мышцах позвоночных». Proc. Natl. Акад. Sci. Соединенные Штаты Америки. 82 (3): 658–662. Bibcode:1985ПНАС ... 82..658С. Дои:10.1073 / pnas.82.3.658. ЧВК 397104. PMID 3871943.
- ^ Белый, HD; Белкнап, Б; Уэбб, MR. (1997). «Кинетика стадий расщепления нуклеозидтрифосфата и высвобождения фосфата ассоциированным актомиозином скелета кролика, измеренная с использованием нового флуоресцентного зонда для определения фосфата». Биохимия. 36 (39): 11828–11836. Дои:10.1021 / bi970540h. PMID 9305974.
- ^ Тирош, Р; Низкий, WZ; Оплатка, А. (1990). «Поступательное движение актиновых филаментов в присутствии тяжелого меромиозина и MgATP, измеренное с помощью доплеровского уширения рассеяния лазерного света». Биохим. Биофиз. Acta. 1037 (3): 274–280. Дои:10.1016 / 0167-4838 (90) 90025-б. PMID 2178685.
- ^ Тирош, Р. (2006). «Баллистические протоны и микроволновые водные растворы (солитоны) в биоэнергетических превращениях». Int. J. Mol. Наука. 7 (9): 320–345. Дои:10.3390 / i7090320.
- ^ Муддана, Хари С .; Сенгупта, Самудра; Маллук, Томас Э .; и другие. (28 января 2010 г.). «Катализ субстрата усиливает диффузию одного фермента». Журнал Американского химического общества. 132 (7): 2110–1. Дои:10.1021 / ja908773a. ЧВК 2832858. PMID 20108965.

- ^ Ридель, Клемент; Габизон, Ронен; Wilson, Christian A.M .; и другие. (8 января 2015 г.). «Тепло, выделяющееся во время каталитического обмена, усиливает диффузию фермента». Природа. 517 (7533): 227–30. Bibcode:2015Натура.517..227р. Дои:10.1038 / природа14043. ЧВК 4363105. PMID 25487146. Сложить резюме – Природа: Новости и просмотры (8 января 2015 г.).
- ^ Рахман, С.А.; Куэста, СМ; Furnham, N; Холлидей, GL; Торнтон, Дж. М. (2014). «EC-BLAST: инструмент для автоматического поиска и сравнения ферментативных реакций». Методы природы. 11 (2): 171–174. Дои:10.1038 / nmeth.2803. ЧВК 4122987. PMID 24412978.
дальнейшее чтение
- Алан Фершт, Структура и механизм в науке о белке: руководство по ферментативному катализу и сворачиванию белков. У. Х. Фриман, 1998. ISBN 0-7167-3268-8
- Специальный выпуск Философские труды B по квантовому катализу в ферментах в свободном доступе.[постоянная мертвая ссылка ]
внешняя ссылка
 СМИ, связанные с Ферментный катализ в Wikimedia Commons
СМИ, связанные с Ферментный катализ в Wikimedia Commons