А-ДНК - A-DNA
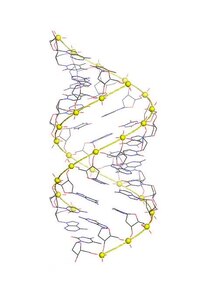
А-ДНК одна из возможных двойных спиральных структур, которые ДНК можно принять. Считается, что А-ДНК является одной из трех биологически активных двойных спиральных структур наряду с B-ДНК и Z-ДНК. Это правая двойная спираль, очень похожая на более распространенную форму B-ДНК, но с более короткой и компактной спиральной структурой, которая пар оснований не перпендикулярны оси спирали, как в B-ДНК. Это было обнаружено Розалинд Франклин, которые также назвали формы A и B. Она показала, что ДНК превращается в форму А в условиях дегидратации. Такие условия обычно используются для образования кристаллов, и многие кристаллические структуры ДНК имеют форму A.[1] Такая же спиральная конформация встречается в двухцепочечных РНК и в гибридных двойных спиралях ДНК-РНК.
Структура
A-ДНК довольно похожа на B-ДНК, учитывая, что это правая двойная спираль с большими и малыми бороздками. Однако, как показано в сравнительной таблице ниже, наблюдается небольшое увеличение количества пар оснований (bp) на оборот (что приводит к меньшему углу закручивания) и меньшее увеличение на пару оснований (делая A-ДНК 20-25% короче, чем B-ДНК). Большая бороздка А-ДНК глубокая и узкая, а малая бороздка широкая и неглубокая. A-ДНК более широкая и, по-видимому, более сжатая по своей оси, чем B-ДНК.[2]
Сравнительная геометрия наиболее распространенных форм ДНК
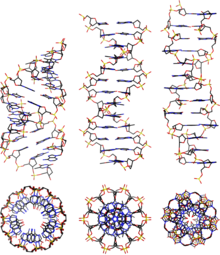

| Атрибут геометрии: | Форма | B-форма | Z-форма |
|---|---|---|---|
| Чувство спирали | правша | правша | левша |
| Повторяющийся блок | 1 п.н. | 1 п.н. | 2 п.н. |
| Вращение / уд. | 32.7° | 34.3° | 60°/2 |
| Среднее б.п. / оборот | 11 | 10 | 12 |
| Наклон н.п. к оси | +19° | −1.2° | −9° |
| Подъем / уд. По оси | 2,6 Å (0,26 нм) | 3,4 Å (0,34 нм) | 3,7 Å (0,37 нм) |
| Подъем / поворот спирали | 28,6 Å (2,86 нм) | 35,7 Å (3,57 нм) | 45,6 Å (4,56 нм) |
| Средняя крутка пропеллера | +18° | +16° | 0° |
| Гликозильный угол | анти | анти | пиримидин: анти, пурин: син |
| Расстояние между нуклеотидным фосфатом и фосфатом | 5,9 Å | 7,0 Å | С: 5,7 Å, G: 6,1 Å |
| Сахарная морщинка | C3'-эндо | C2'-эндо | C: C2'-эндо, G: C3'-эндо |
| Диаметр | 23 Å (2,3 нм) | 20 Å (2,0 нм) | 18 Å (1,8 нм) |
Биологическая функция
Обезвоживание ДНК переводит ее в форму А, и это, по-видимому, защищает ДНК в таких условиях, как экстремальное обезвоживание бактерий.[3] Связывание с белками также может удалять растворитель с ДНК и преобразовывать его в форму А, что подтверждается структурой нескольких гипертермофильных вирусов архей, включая палочковидные вирусы. рудивирусы SIRV2 [4] и SSRV1,[5] обволакивающий нитчатый липотриксвирусы AFV1,[6] SFV1 [7] и SIFV,[5] тристромавирус PFV2 [8] а также икосаэдр портоглобовирус SPV1.[9] Считается, что ДНК А-формы является одной из адаптаций гипертермофильных вирусов архей к суровым условиям окружающей среды, в которых эти вирусы процветают.
Было высказано предположение, что двигатели, которые упаковывают двухцепочечную ДНК в бактериофагах, используют тот факт, что A-ДНК короче, чем B-ДНК, и что конформационные изменения в самой ДНК являются источником больших сил, генерируемых этими двигателями.[10] Экспериментальные доказательства того, что A-ДНК является промежуточным звеном в вирусной биомоторной упаковке, получены из двойного красителя. Фёрстеровский резонансный перенос энергии измерения, показывающие, что B-ДНК укорачивается на 24% в остановившемся («сломанном») промежуточном продукте А-формы.[11][12] В этой модели гидролиз АТФ используется для управления конформационными изменениями белков, которые альтернативно дегидратируют и регидратируют ДНК, а цикл укорочения / удлинения ДНК связан с циклом захвата / высвобождения белок-ДНК для создания поступательного движения, которое перемещает ДНК в капсид. .
Смотрите также
Рекомендации
- ^ Розалинд, Франклин (1953). «Структура тимонуклеатных волокон натрия. I. Влияние содержания воды» (PDF). Acta Crystallographica. 6 (8): 673–677. Дои:10.1107 / s0365110x53001939.
- ^ Дикерсон, Ричард Э. (1992). Структура ДНК от А до Я. Методы в энзимологии. 211. стр.67–111. Дои:10.1016/0076-6879(92)11007-6. ISBN 9780121821128. PMID 1406328 - через Elsevier Science Direct.
- ^ Уилан Д. Р. и др. (2014). «Обнаружение массового и обратимого конформационного перехода B- в A-ДНК у прокариот в ответ на высыхание». Интерфейс J R Soc. 11 (97): 20140454. Дои:10.1098 / rsif.2014.0454. ЧВК 4208382. PMID 24898023.
- ^ Ди Майо Ф., Эгельман Э. Х. и др. (2015). «Вирус, поражающий гипертермофил, инкапсидирует ДНК А-формы». Наука. 348 (6237): 914–917. Bibcode:2015Научный ... 348..914D. Дои:10.1126 / science.aaa4181. ЧВК 5512286. PMID 25999507.
- ^ а б Ванга, Ф; Бакеро, Д.П .; Beltran, LC; Вс, З; Осинский, Т; Чжэн, Вт; Прангишвили, Д; Крупович, М; Эгельман, EH (5 августа 2020 г.). «Структуры нитчатых вирусов, заражающих гипертермофильные археи, объясняют стабилизацию ДНК в экстремальных условиях». Труды Национальной академии наук Соединенных Штатов Америки. 117 (33): 19643–19652. Дои:10.1073 / pnas.2011125117. PMID 32759221.
- ^ Kasson, P; DiMaio, F; Ю, Х; Lucas-Staat, S; Крупович, М; Schouten, S; Прангишвили, Д; Эгельман, EH (2017). «Модель новой мембранной оболочки нитчатого гипертермофильного вируса». eLife. 6: e26268. Дои:10.7554 / eLife.26268. ЧВК 5517147. PMID 28639939.
- ^ Лю, Y; Осинский, Т; Ванга, Ф; Крупович, М; Schouten, S; Kasson, P; Прангишвили, Д; Эгельман, EH (2018). «Структурная консервация в оболочечном нитчатом вирусе, заражающем гипертермофильный ацидофил». Nature Communications. 9 (1): 3360. Bibcode:2018NatCo ... 9.3360L. Дои:10.1038 / s41467-018-05684-6. ЧВК 6105669. PMID 30135568.
- ^ Ванга, Ф; Бакеро, Д.П .; Вс, З; Осинский, Т; Прангишвили, Д; Egelman, EH; Крупович, М (2020). «Структура нитевидного вируса раскрывает семейные связи в виросфере архей». Эволюция вирусов. 6 (1): veaa023. Дои:10.1093 / ve / veaa023. ЧВК 7189273. PMID 32368353.
- ^ Ванга, Ф; Лю, Y; Вс, З; Осинский, Т; де Оливейра, GAP; Конвей, Дж. Ф.; Schouten, S; Крупович, М; Прангишвили, Д; Эгельман, EH (2019). «Упаковка для ДНК А-формы в икосаэдрическом вирусе». Труды Национальной академии наук Соединенных Штатов Америки. 116 (45): 22591–22597. Дои:10.1073 / пнас.1908242116. ЧВК 6842630. PMID 31636205.
- ^ Харви, SC (2015). «Гипотеза червяка: переходы между A-ДНК и B-ДНК обеспечивают движущую силу для упаковки генома в двухцепочечных ДНК бактериофагах». Журнал структурной биологии. 189 (1): 1–8. Дои:10.1016 / j.jsb.2014.11.012. ЧВК 4357361. PMID 25486612.
- ^ Орам, М. (2008). «Модуляция реакции упаковки t4-терминазы бактериофага структурой ДНК». Дж Мол Биол. 381 (1): 61–72. Дои:10.1016 / j.jmb.2008.05.074. ЧВК 2528301. PMID 18586272.
- ^ Рэй, К. (2010). «Хруст ДНК вирусным мотором упаковки: сжатие прокапсидного портального остановленного субстрата Y-ДНК». Вирусология. 398 (2): 224–232. Дои:10.1016 / j.virol.2009.11.047. ЧВК 2824061. PMID 20060554.
внешняя ссылка
Виды нуклеиновые кислоты | |||||||
|---|---|---|---|---|---|---|---|
| Избиратели | |||||||
| Рибонуклеиновые кислоты (кодирование, некодирование ) |
| ||||||
| Дезоксирибонуклеиновые кислоты | |||||||
| Аналоги | |||||||
| Клонирование векторов | |||||||
| |||||||