Антисмысловая РНК - Antisense RNA

Антисмысловая РНК (asRNA), также называемый антисмысловым транскриптом,[1] естественный антисмысловой транскрипт (NAT)[2][3][4] или антисмысловой олигонуклеотид,[5] одноцепочечный РНК что комплементарно кодирующему белку информационная РНК (мРНК), с которой он гибридизуется, и тем самым блокирует его перевод в белок. asRNAs (которые встречаются в природе) были обнаружены в обоих прокариоты и эукариоты,[1] антисмысловые транскрипты можно разделить на короткие (<200 нуклеотидов) и длинные (> 200 нуклеотидов). некодирующие РНК (нкРНК).[4] Основная функция asRNA - регулирование экспрессия гена. asRNA также могут быть получены синтетическим путем и нашли широкое распространение в качестве инструментов исследования для нокдаун генов. Они также могут иметь терапевтическое применение.[6][1][4]
Открытие и история разработки лекарств
Некоторые из самых ранних asRNA были обнаружены при исследовании функциональных белков. Пример был micF asRNA. Характеризуя внешнюю мембрану порин ompC в Кишечная палочка некоторые из наблюдаемых клонов промотора ompC были способны подавлять экспрессию другого мембранного порина, такого как ompF. Было обнаружено, что область, ответственная за эту функцию репрессии, находится в локусе из 300 пар оснований выше промотора ompC. Эта область из 300 пар оснований на 70% гомологична по последовательности с 5 'конец мРНК ompF и, таким образом, транскрипт этого локуса из 300 пар оснований комплементарен мРНК ompF. Позже этот транскрипт, обозначенный как micF, оказался asRNA ompF и способен подавлять экспрессию ompF при стрессе за счет образования дуплекса с мРНК ompF. Это вызывает деградацию мРНК ompF.[2]
В отличие от РНК micF, обнаруженной случайно, большинство asRNA были обнаружены в результате полногеномного поиска малых регуляторных РНК и с помощью транскриптом анализ. Обычно первый шаг включает в себя вычислительные предсказания, основанные на некоторых известных характеристиках асРНК. Во время вычислительного поиска исключаются области кодирования. Области, которые, как предполагается, имеют консервативные структуры РНК и действуют как орфанные промоторы и Ро независимые терминаторы предпочтение отдается во время анализа. Поскольку вычислительный поиск сосредоточен на межгенная область, asRNAs, которые транскрибируются с противоположной цепи кодирующего гена, вероятно, будут пропущены с помощью этого метода. Чтобы обнаружить асРНК, транскрибируемую из кодирующей области, олигонуклеотидные микрочипы может быть использован. В этом методе одну или обе цепи кодирующих генов можно использовать в качестве зондов. Помимо компьютерного поиска и микроматриц, некоторые asRNA были обнаружены путем секвенирования клонов кДНК, а также картирования промоторных элементов.[7] Хотя многие результаты упомянутых выше подходов привели к появлению множества возможных асРНК, лишь немногие из них оказались действительными асРНК с помощью дальнейших функциональных тестов. Чтобы свести к минимуму количество ложноположительных результатов, новые подходы последних лет были сосредоточены на специфичной для цепи транскрипции, хроматин связывание некодирующих РНК и исследования отдельных клеток.[1]
Идея асРНК как мишеней для лекарств возникла в 1978 году, когда Замечник и Стефенсон обнаружили антисмысловой олигонуклеотид к вирусной РНК вируса скаркомы Рауса, который был способен ингибировать репликацию вируса и синтез белка. С тех пор много усилий было направлено на разработку асРНК в качестве кандидатов в лекарства. В 1998 году был выпущен первый препарат асРНК, Fomivirsen, был одобрен FDA. Фомивирсен, олигонуклеотид из 21 пары оснований, был разработан для лечения цитомегаловирусный ретинит у больных СПИДом. Он работает путем нацеливания на транскрибируемую мРНК вируса и, следовательно, ингибирования репликации цитомегаловируса. Несмотря на то, что производство фомивирсена было прекращено в 2004 году из-за потери рынка, он послужил успешным и вдохновляющим примером использования асРНК в качестве мишеней или кандидатов в лекарства.[5]
Другой пример использования асРНК в качестве терапевтического агента: Мипомерсен, который был одобрен FDA в 2013 году. Мипомерсен был разработан для управления уровнем холестерин липопротеинов низкой плотности (ЛПНП) у пациентов с гомозиготным семейная гиперхолестеринемия (HoFH), которое является редким аутосомно-доминантным генетическим заболеванием. Из-за высокого уровня общего холестерина (650–1000 мг / дл) и рецепторов ЛПНП (выше 600 мг / дл) в HoFH пациенты с HoFH имеют высокий риск ишемической болезни сердца. Потому что белок апо-В-100 было установлено, что требуется для производства липопротеины очень низкой плотности (ЛПОНП) и ЛПНП, мипомерсен дополняет мРНК апо-B-100 и нацеливает ее на РНКаза H зависимая деградация. В конечном итоге мипомерсен способен снижать уровень ЛПНП.[8]
Примеры для разных видов
Первоначальные обнаруженные асРНК были у прокариот, включая плазмиды, бактериофаг и бактерии. Например, в плазмиде ColE1 асРНК, называемая РНК I, играет важную роль в определении числа копий плазмиды путем контроля репликации. Репликация ColE1 зависит от транскрипции праймерной РНК, называемой РНК II. После того, как РНК II транскрибируется, она гибридизуется со своей ДНК-матрицей, а затем расщепляется РНКазой Н. В присутствии асРНК РНК I, РНК I и РНК II образуют дуплекс, который вносит конформационное изменение РНК II. Следовательно, РНК II не может гибридизоваться со своей ДНК-матрицей, что приводит к низкому количеству копий ColE1. В бактериофаге P22 сар асРНК помогает регулировать литический и лизогенный цикл, контролируя экспрессию Ant.[9] Помимо экспрессии в прокариотах, асРНК также были обнаружены в растениях. Наиболее хорошо описанный пример регуляции асРНК у растений приведен на Цветущий Локус C (FLC) ген. Ген FLC в Arabidopsis thaliana кодирует фактор транскрипции, который предотвращает экспрессию ряда генов, вызывающих переход цветков. В холодной среде asRNA гена FLC, обозначаемая COOLAIR, экспрессируется и ингибирует экспрессию FLC посредством модификации хроматина, что, следовательно, способствует цветению.[10] В клетках млекопитающих типичным примером регуляции асРНК является инактивация Х-хромосомы. Xist, асРНК, может рекрутировать поликомб репрессивный комплекс 2 (PRC2), что приводит к гетерохроматинизации Х-хромосомы.[3]
Классификация
Антисмысловые РНК можно классифицировать по-разному. Что касается регуляторных механизмов, некоторые авторы группируют асРНК во взаимодействиях РНК-ДНК, взаимодействия РНК-РНК либо в ядро или же цитоплазма и РНК-белковые взаимодействия (эпигенетический ).[3] Антисмысловые РНК можно классифицировать по типу промоторов, которые инициируют экспрессию асРНК: независимые промоторы, общие двунаправленные промоторы или скрытые промоторы. Что касается длины, хотя асРНК в целом классифицируется как днРНК, существуют короткие асРНК с длиной менее 200 пар оснований. Поскольку установлено, что регуляторный механизм асРНК является видоспецифичным, асРНК также можно классифицировать по видам.[1] Один из распространенных способов классификации асРНК - это то, где асРНК транскрибируются относительно своих генов-мишеней: цис-действующий и трансакционный.
Цис-действующий
Цис-действующий asRNA транскрибируются с противоположной цепи целевого гена в локусе целевого гена. Они часто демонстрируют высокую степень или полную комплементарность с целевым геном. Если цис-действующая асРНК регулирует экспрессию гена, воздействуя на мРНК, она может нацеливаться только на индивидуальную мРНК. При взаимодействии с нацеливающими мРНК цис-действующие асРНК могут либо блокировать связывание рибосом, либо рекрутировать РНКазу для разрушения нацеливающих мРНК. Следовательно, функция этих цис-действующих асРНК заключается в репрессии трансляции направленных мРНК.[2] Помимо цис-действующих асРНК, нацеленных на мРНК, существуют цис-действующие эпигенетические глушители и активаторы. С точки зрения эпигенетической модификации цис-действие относится к природе этих асРНК, которые регулируют эпигенетические изменения вокруг места где они расшифрованы. Вместо нацеливания на отдельные мРНК эти цис-действующие эпигенетические регуляторы могут привлекать модифицирующие хроматин ферменты, которые могут оказывать влияние как на локусы транскрипции, так и на соседние гены.[3]
Транс-действующий
Транс-действующий asRNA транскрибируются из локусов, удаленных от нацеливающих генов. В отличие от цис-действующих асРНК, они демонстрируют низкую степень комплементарности с целевым геном, но могут быть длиннее, чем цис-действующие асРНК. Они также могут нацеливаться на несколько локусов. Из-за этих свойств транс-действующих асРНК они образуют менее стабильные комплексы со своими транскриптами нацеливания и иногда требуют помощи от РНК. шаперонный белок такие как Hfq для выполнения своих функций. Из-за сложности транс-действующих asRNAs в настоящее время они рассматриваются как менее поддающиеся воздействию лекарств мишени.[2]
Функция
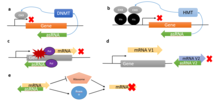
Эпигенетическая регуляция
Многие примеры asRNA демонстрируют ингибирующий эффект на инициацию транскрипции посредством эпигенетических модификаций.
Метилирование ДНК
Метилирование ДНК может привести к долгосрочному подавлению специфических генов. Репрессия функциональных белков посредством индуцированного асРНК метилирования ДНК была обнаружена при нескольких заболеваниях человека. В классе альфа-талассемия, тип заболевания крови, при котором снижается уровень гемоглобина, что приводит к недостатку кислорода в тканях,[11] гемоглобин альфа1 ген (HBA1) подавляется аномальным транскриптом предполагаемого РНК-связывающего белка Luc7-подобного (LUC71), который служит асРНК для HBA1 и индуцирует метилирование промотора HBA1.[1] Другой пример - подавление гена-супрессора опухолей p15INK4b, также называемого CDKN2B, при остром лимфобластном лейкозе и остром миелоидном лейкозе. АсРНК, которая отвечает за этот эффект сайленсинга, представляет собой антисмысловую некодирующую РНК в локусе INK (АНРИЛ ), который экспрессируется в том же локусе, который кодирует p15INK4b.[3]
Модификация гистона
В эукариотических клетках ДНК плотно упакована гистоны. Модификация гистонов может изменить взаимодействия с ДНК, что может в дальнейшем вызвать изменения в экспрессия гена. Биологические последствия метилирование гистонов зависят от контекста. В общем, метилирование гистонов приводит к репрессии гена, но также может быть достигнута активация гена.[12] Доказательства показали, что метилирование гистонов может быть индуцировано асРНК. Например, ANRIL, помимо способности индуцировать метилирование ДНК, также может репрессировать соседний ген CDKN2B, CDKN2A, путем набора поликомб репрессивный комплекс 2 (PRC2), что приводит к метилированию гистонов (H3K27me). Другой классический пример - инактивация Х-хромосомы XIST.[1]
Эпигенетическая модификация, вызванная ANRIL, является примером цис-действующей эпигенетической регуляции.[3] Кроме того, модификация хроматина, индуцированная антисмысловой РНК, может быть как транс-действующей. Например, у млекопитающих asRNA ГОРЯЧИЙ ВОЗДУХ записано с гомеобокс C (HOXC) локус, но он рекрутирует PRC2 в HOXD, который накладывает H3K27 и заглушает HOXD. HOTAIR высоко экспрессируется в первичных опухолях груди.[1]
Ко-транскрипционная регуляция
Эпигенетические регуляции, такие как метилирование ДНК и метилирование гистонов, могут подавлять экспрессию генов, подавляя инициацию транскрипции. Однако иногда репрессия генов может быть достигнута путем преждевременного прекращения или замедления процесса транскрипции. AsRNA могут участвовать в регуляции этого уровня генов. Например, в бактериальных или эукариотических клетках, где присутствуют сложные РНК-полимеразы, двунаправленная транскрипция в одном и том же локусе может привести к конфликту полимераз и привести к прекращению транскрипции. Даже когда столкновение полимеразы маловероятно во время слабой транскрипции, также может происходить пауза полимеразы, которая блокирует элонгацию и приводит к репрессии гена. Один из примеров - подавление IME4 ген его asRNA RME2. Другой способ ко-транскрипционного воздействия на транскрипцию - блокирование сплайсинга. Один классический пример из жизни человека: цинковый палец E-box связывание с геном гомеобокса 2 (ZEB2 ), который кодирует E-кадгерин, репрессор транскрипции. Для эффективной трансляции мРНК ZEB2 требуется наличие внутренний сайт входа рибосомы (IRES) в интроне мРНК на 5 'конец. При экспрессии asRNA ZEB2 она может маскировать сайт сплайсинга и поддерживать IRES в мРНК, что приводит к эффективному синтезу E-кадгерина. Наконец, в зависимости от уровня экспрессии асРНК могут быть получены различные изоформы смыслового транскрипта. Следовательно, asRNA-зависимая регуляция не ограничивается механизмом включения / выключения; скорее, он представляет собой прекрасную систему управления тоном.[1]
Посттранскрипционная регуляция
Прямая посттранскрипционная модуляция с помощью asRNA относится к мРНК, на которую непосредственно нацелены asRNA; таким образом, это влияет на перевод. Некоторые характеристики этого типа асРНК описаны в цис- и трансформирующих асРНК. Этот механизм относительно быстр, потому что и мРНК нацеливания, и ее асРНК должны одновременно присутствовать в одной и той же клетке. Как описано в цис-действующих асРНК, спаривание мРНК-асРНК может привести к блокированию входа в рибосомы и зависимой от РНКазы Н деградации. В целом, асРНК, нацеленные на мРНК, могут либо активировать, либо ингибировать трансляцию смысловых мРНК, при этом ингибирующий эффект является наиболее выраженным.[1]
Лечебный потенциал
Как регуляторный элемент asRNAs имеют много преимуществ, чтобы их можно было рассматривать в качестве мишени для лекарств. Прежде всего, асРНК регулируют экспрессию генов на нескольких уровнях, включая транскрипцию, посттранскрипцию и эпигенетические модификации. Во-вторых, цис-действующие асРНК специфичны для последовательности и демонстрируют высокую степень комплементарности с нацеливающими генами.[1] В-третьих, уровень экспрессии асРНК очень мал по сравнению с уровнем экспрессии мРНК нацеливания; следовательно, для достижения эффекта требуется лишь небольшое количество асРНК. С точки зрения мишеней для лекарств это является огромным преимуществом, поскольку для их эффективности требуется только низкая дозировка.[4]
В последние годы идея нацеливания на asRNAs для увеличения экспрессии генов специфическим для локуса образом привлекает большое внимание. Из-за характера разработки лекарств всегда легче получить лекарственные средства, действующие как ингибиторы или подавители. Однако существует потребность в разработке лекарств, которые могут активировать или повышать экспрессию генов, таких как гены-супрессоры опухолей, нейропротективные факторы роста и гены, которые, как обнаружено, подавляются при определенных менделевских расстройствах. В настоящее время подход к восстановлению недостаточной экспрессии генов или функции белка включает заместительную ферментативную терапию, микроРНК терапии и доставки функциональной кДНК. Однако у каждого есть свои недостатки. Например, синтезированный белок, используемый в заместительной ферментной терапии, часто не может имитировать всю функцию эндогенного белка. Кроме того, заместительная ферментная терапия - это пожизненное обязательство и большое финансовое бремя для пациента. Из-за локус-специфической природы асРНК и свидетельств изменений в экспрессии асРНК при многих заболеваниях были попытки создать одноцепочечные олигонуклеотиды, называемые антагоНАТ, для ингибирования асРНК и, в конечном итоге, для повышения экспрессии специфических генов.[4]
Несмотря на обещания асРНК в качестве мишеней для лекарств или кандидатов в лекарственные средства, остается еще несколько проблем, которые необходимо решить.[13] Прежде всего, асРНК и антагоНАТ могут легко разрушаться РНКазой или другими разрушающими ферментами. Чтобы предотвратить деградацию терапевтических олигонуклеотидов, обычно требуется химическая модификация. Наиболее распространенная химическая модификация олигонуклеотидов - это добавление фосфоротиоат связь с позвоночником.[5] Однако модификация фосфротиоата может быть провоспалительной. Побочные эффекты, включая жар, озноб или тошноту, наблюдались после местной инъекции олигонуклеотидов, модифицированных фосфротиоатом. Во-вторых, нецелевое значение токсичности также представляет большую проблему. Несмотря на локус-специфичную природу эндогенных асРНК, только 10-50% синтезированных олигонуклеотидов показали ожидаемый целевой эффект. Одна из возможных причин этой проблемы - высокие требования к структуре асРНК, которая должна распознаваться последовательностью-мишенью и РНКазой H. Единичное несоответствие может привести к искажению вторичной структуры и привести к нецелевым эффектам.[4] Наконец, было показано, что искусственные asRNA имеют ограниченное внутриклеточное поглощение.[5] Хотя было показано, что нейроны и глия обладают способностью свободно поглощать голые антисмысловые олигонуклеотиды, прослеживаемые носители, такие как вирус и липидные везикулы, по-прежнему будут идеальными для контроля и мониторинга внутриклеточной концентрации и метаболизма.[4]
Смотрите также
Рекомендации
- ^ а б c d е ж грамм час я j k Пелехано В., Штейнмец Л.М. (декабрь 2013 г.). «Регулирование генов с помощью антисмысловой транскрипции». Обзоры природы. Генетика. 14 (12): 880–893. Дои:10.1038 / nrg3594. PMID 24217315.
- ^ а б c d Сабери Ф., Камали М., Наджафи А., Язданпараст А., Могхаддам М. М. (2016-07-28). «Природные антисмысловые РНК как регуляторные элементы мРНК в бактериях: обзор функций и применения». Письма о клеточной и молекулярной биологии. 21: 6. Дои:10.1186 / s11658-016-0007-z. ЧВК 5415839. PMID 28536609.
- ^ а б c d е ж Magistri M, Faghihi MA, St Laurent G, Wahlestedt C (август 2012 г.). «Регулирование структуры хроматина с помощью длинных некодирующих РНК: основное внимание уделяется естественным антисмысловым транскриптам». Тенденции в генетике. 28 (8): 389–396. Дои:10.1016 / j.tig.2012.03.013. ЧВК 3768148. PMID 22541732.
- ^ а б c d е ж грамм Wahlestedt C (июнь 2013 г.). «Нацеливание на длинную некодирующую РНК для терапевтического повышения экспрессии гена». Обзоры природы. Открытие наркотиков. 12 (6): 433–446. Дои:10.1038 / nrd4018. PMID 23722346.
- ^ а б c d Коле Р., Крайнер А.Р., Альтман С. (январь 2012 г.). «РНК-терапия: помимо РНК-интерференции и антисмысловых олигонуклеотидов». Обзоры природы. Открытие наркотиков. 11 (2): 125–140. Дои:10.1038 / nrd3625. ЧВК 4743652. PMID 22262036.
- ^ Вайс Б., Давидкова Г., Чжоу Л.В. (март 1999 г.). «Генная терапия антисмысловой РНК для изучения и модуляции биологических процессов». Клеточные и молекулярные науки о жизни. 55 (3): 334–358. Дои:10.1007 / с000180050296. PMID 10228554.
- ^ Томасон МК, Сторц Г. (2010). «Бактериальные антисмысловые РНК: сколько их и что они делают?». Ежегодный обзор генетики. 44 (1): 167–188. Дои:10.1146 / annurev-genet-102209-163523. ЧВК 3030471. PMID 20707673.
- ^ Вонг Э., Голдберг Т. (февраль 2014 г.). «Мипомерсен (кинамро): новый антисмысловой олигонуклеотидный ингибитор для лечения гомозиготной семейной гиперхолестеринемии». P&T. 39 (2): 119–122. ЧВК 3956393. PMID 24669178.
- ^ Саймонс Р.В. (декабрь 1988 г.). «Естественный контроль антисмысловой РНК - краткий обзор». Ген. 72 (1–2): 35–44. Дои:10.1016/0378-1119(88)90125-4. PMID 2468573.
- ^ Ietswaart R, Wu Z, Dean C (сентябрь 2012 г.). «Контроль времени цветения: еще одно окно для связи антисмысловой РНК и хроматина». Тенденции в генетике. 28 (9): 445–453. Дои:10.1016 / j.tig.2012.06.002. PMID 22785023.
- ^ «альфа-талассемия». Домашний справочник по генетике. Национальная медицинская библиотека США NIH. 14 ноября 2017.
- ^ Ветстайн JR (2010). «Метилирование гистонов». Справочник по сотовой сигнализации (Второе изд.). С. 2389–2397. Дои:10.1016 / b978-0-12-374145-5.00287-4. ISBN 978-0-12-374148-6.
- ^ Вайс Б. (ред.): Антисмысловые олигодезоксинуклеотиды и антисмысловая РНК: новые фармакологические и терапевтические агенты, CRC Press, Boca Raton, FL, 1997.
Виды нуклеиновые кислоты | |||||||
|---|---|---|---|---|---|---|---|
| Избиратели | |||||||
| Рибонуклеиновые кислоты (кодирование, некодирование ) |
| ||||||
| Дезоксирибонуклеиновые кислоты | |||||||
| Аналоги | |||||||
| Клонирование векторов | |||||||
| |||||||