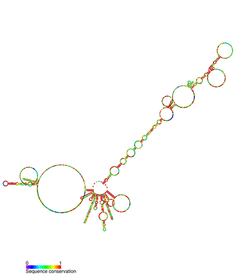
Компонент теломеразной РНК, также известный как TR, TER или TERC, является нкРНК нашел в эукариоты это компонент теломераза, фермент, используемый для увеличения теломеры.[2][3] TERC служит шаблоном для теломер репликация (обратная транскрипция ) теломеразой. Теломеразные РНК сильно различаются по последовательности и структуре у позвоночных, инфузорий и дрожжей, но они имеют общий 5'псевдоузел структура близка к шаблонной последовательности. РНК теломеразы позвоночных имеют 3 ' H / ACAsnoRNA -подобный домен.[4][5][6]
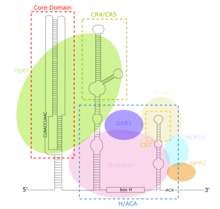
TERC - это Длинная некодирующая РНК (lncRNA) длиной от ~ 150 нт у инфузорий до 400-600 нт у позвоночных и 1300 нт у дрожжей (Алнафах). Зрелый человеческий TERC (hTR) имеет длину 451 н.[7] TERC имеет обширные вторичные структурные особенности более 4 основных консервативных доменов.[8] Основной домен, самый большой домен на 5 ’конце TERC, содержит CUAAC Теломер последовательность шаблона. Его вторичная структура состоит из большой петли, содержащей матричную последовательность, спирали, замыкающей петлю P1, и P2 / P3 псевдоузел.[9] Основной домен и консервативный домен CR4 / CR5 ассоциируются с TERT, и являются единственными доменами TERC, необходимыми для каталитической активности теломеразы in vitro.[10] 3 ’конец TERC состоит из консервативного домена H / ACA,[9] структура из 2 шпилек, соединенных однонитевым шарниром и окаймленных на 3 ’конце однонитевой последовательностью АСА.[7] Домен H / ACA связывает Дискерин, GAR1, NOP10, NHP2, чтобы сформировать H / ACA RNP сложный.[9] Консервативный домен CR7 также локализован на 3 ’конце TERC и содержит 3nt CAB (Тело Кахала Локализация), который связывает TCAB1.[9]
Иллюстрация: hTR и ассоциированные белки теломеразного комплекса
Функция
Теломераза - это рибонуклеопротеиновая полимераза, которая поддерживает концы теломер за счет добавления теломерного повтора TTAGGG. Это повторение различается в зависимости от эукариоты (см. таблицу на теломер статью для полного списка). Фермент состоит из белкового компонента (TERT ) с обратная транскриптаза активность и компонент РНК, кодируемый этим геном, который служит шаблоном для теломерного повтора. CCCUAA, обнаруженный около позиции 50 последовательности TERC позвоночных, действует как матрица. Экспрессия теломеразы играет роль в клеточном старение, поскольку это обычно подавляется в послеродовой период соматические клетки в результате прогрессирующего укорачивания теломер. Нарушение регуляции экспрессии теломеразы в соматических клетках может быть вовлечено в онкогенез. Исследования на мышах показывают, что теломераза также участвует в хромосомный ремонт, поскольку de novo синтез теломерных повторов могут встречаться двухцепочечные разрывы.[11]Гомологи TERC также можно найти в Вирусы галлидного герпеса.[12]
Основной домен TERC содержит РНК шаблон, из которого TERT синтезирует теломерные повторы TTAGGG.[9] В отличие от других RNP, в теломераза, белок TERT является каталитическим, в то время как днРНК TERC является структурным, а не действует как рибозим.[13] Основной регион TERC и TERT достаточны для восстановления каталитической активности теломеразы in vitro.[9][10] Домен H / ACA TERC принимает на работу Дискерин сложный (DKC1, GAR1, NOP10, NHP2 ), который стабилизирует TERC, увеличивая образование комплекса теломеразы и общую каталитическую активность.[9] Домен CR7 связывает TCAB1, который локализует теломеразу в кахальные тела, дополнительно увеличивая каталитическую активность теломеразы.[9] TERC экспрессируется повсеместно, даже в клетках, лишенных теломеразной активности и экспрессии TERT.[14] В результате были предложены различные независимые от TERT функциональные роли TERC. 14 гены содержащие мотив связывания TERC, непосредственно транскрипционно регулируемый с помощью TERC посредством увеличения экспрессии, опосредованного образованием триплекса РНК-ДНК. TERC-опосредованная активация Lin37, Trpg1l, Tyrobp, УСП16 стимулирует NF-κB пути, что приводит к увеличению экспрессии и секреции воспалительных цитокины.[15]
Биосинтез
В отличие от большинства днРНК которые собраны из интроны посредством сплайсосома, hTR напрямую записано из специального промоутерский сайт[7] расположен в геномном локусе 3q26.2[16] к РНК-полимераза II.[7] Зрелый hTR имеет длину 451nt, но примерно 1/3 клеточных транскриптов hTR в устойчивом состоянии имеют ~ 10nt геномно кодируемых 3 ’хвостов. Большинство этих удлиненных видов hTR имеют дополнительное удлинение олиго-A 3 ’.[7] Обработка незрелого 3’-хвостового hTR до зрелого 451nt hTR может быть выполнена прямым 3’-5 ’ эксорибонуклеолитический деградация или косвенный путь олигоаденилирования с помощью PAPD5, удаление 3’-олиго-A-хвоста экзонуклеазой 3’-5 ’РНК ПАРН и последующее 3’-5 ’экзорибонуклеолитическое разложение.[7] Расширенные транскрипты hTR также расщепляются РНК. экзосома.[7]
5’-концы транскриптов hTR также дополнительно процессируются. ТГС-1гиперметилирование то 5'-метилгуанозиновый кэп к кэпу N2,2,7 триметилгуанозина (TMG), который ингибирует созревание hTR.[17] Связывание Дискерин комплекс с транскрибируемыми доменами H / ACA hTR во время транскрипции способствует прекращению транскрипции.[7] Контроль относительной скорости этих различных конкурирующих путей, которые активируют или ингибируют созревание hTR, является важным элементом регуляции общей активности теломеразы.
TERC использовался как защитный остеопороз, с его повышенной экспрессией, задерживающей скорость остеогенез.[23] Из-за его сверхэкспрессии в ряде фенотипов рака, TERC был исследован как потенциальный биомаркер рака. Было обнаружено, что он является эффективным биомаркером плоскоклеточный рак легкого (LUSC).[24]
^Лингнер Дж., Хендрик Л.Л., Чех Т.Р. (август 1994 г.). «Теломеразные РНК разных инфузорий имеют общую вторичную структуру и пермутированную матрицу». Гены и развитие. 8 (16): 1984–98. Дои:10.1101 / гад.8.16.1984. PMID7958872.
^Theimer CA, Feigon J (июнь 2006 г.). «Структура и функции теломеразной РНК». Текущее мнение в структурной биологии. 16 (3): 307–18. Дои:10.1016 / j.sbi.2006.05.005. PMID16713250.
^Fragnet L, Kut E, Rasschaert D (июнь 2005 г.). «Сравнительное функциональное исследование теломеразной РНК вируса на основе естественных мутаций». Журнал биологической химии. 280 (25): 23502–15. Дои:10.1074 / jbc.M501163200. PMID15811851. S2CID24301693.
^Гао ГК, Ян Д.В., Лю В. (январь 2020 г.). «LncRNA TERC облегчает прогрессирование остеопороза, поглощая miRNA-217, чтобы активировать RUNX2». Европейский обзор медицинских и фармакологических наук. 24 (2): 526–534. Дои:10.26355 / eurrev_202001_20029. PMID32016954.
^Storti CB, de Oliveira RA, de Carvalho M, Hasimoto EN, Cataneo DC, Cataneo AJ, et al. (Февраль 2020 г.). «Теломерные гены и теломерные днРНК являются кандидатами в биомаркеры плоскоклеточного рака легких (LUSC)». Экспериментальная и молекулярная патология. 112: 104354. Дои:10.1016 / j.yexmp.2019.104354. PMID31837325.
Маррон А., Докал I (декабрь 2004 г.). «Врожденный дискератоз: молекулярное понимание функции теломеразы, старения и рака». Обзоры экспертов в области молекулярной медицины. 6 (26): 1–23. Дои:10.1017 / S1462399404008671. PMID15613268.
Ямагути Х (июнь 2007 г.). «Мутации генов комплекса теломеразы, связанные с отказами костного мозга». Журнал медицинской школы Ниппон. 74 (3): 202–9. Дои:10.1272 / jnms.74.202. PMID17625368.
Вонг К.К., Чанг С., Вейлер С.Р., Ганесан С., Чаудхури Дж., Чжу С. и др. (Сентябрь 2000 г.). «Дисфункция теломер нарушает восстановление ДНК и увеличивает чувствительность к ионизирующему излучению». Природа Генетика. 26 (1): 85–8. Дои:10.1038/79232. PMID10973255. S2CID1873111.
Митчелл Дж. Р., Коллинз К. (август 2000 г.). «Активация теломеразы человека требует двух независимых взаимодействий между теломеразной РНК и обратной транскриптазой теломеразы». Молекулярная клетка. 6 (2): 361–71. Дои:10.1016 / S1097-2765 (00) 00036-8. PMID10983983.
Имото И., Пимхаокхам А., Фукуда Й., Янг З. К., Шимада Ю., Номура Н. и др. (Август 2001 г.). «SNO является вероятной мишенью для амплификации гена на 3q26 в плоскоклеточном раке пищевода». Сообщения о биохимических и биофизических исследованиях. 286 (3): 559–65. Дои:10.1006 / bbrc.2001.5428. PMID11511096.
Вуллиами Т., Маррон А., Голдман Ф, Дирлав А., Бесслер М., Мейсон П. Дж., Докал I (сентябрь 2001 г.). «РНК-компонент теломеразы мутирован при аутосомно-доминантном врожденном дискератозе». Природа. 413 (6854): 432–5. Bibcode:2001Натура.413..432В. Дои:10.1038/35096585. PMID11574891. S2CID4348062.
Чжан Р.Г., Чжан Р.П., Ван XW, Се Х. (март 2002 г.). «Влияние цисплатина на активность теломеразы и длину теломер в клетках гепатомы человека BEL-7404». Клеточные исследования. 12 (1): 55–62. Дои:10.1038 / sj.cr.7290110. PMID11942411. S2CID36839452.
Ян И, Чен И, Чжан Ц., Хуанг Х, Вайсман С.М. (июль 2002 г.). «Ядрышковая локализация белка hTERT связана с функцией теломеразы». Экспериментальные исследования клеток. 277 (2): 201–9. Дои:10.1006 / excr.2002.5541. PMID12083802.
Чанг Дж. Т., Чен Ю. Л., Ян Х. Т., Чен К. Ю., Ченг А. Дж. (Июль 2002 г.). «Дифференциальная регуляция активности теломеразы шестью субъединицами теломеразы». Европейский журнал биохимии. 269 (14): 3442–50. Дои:10.1046 / j.1432-1033.2002.03025.x. PMID12135483.
Суд А.К., Гроб Дж., Джаббари С., Буллер Р. Э., Хендрикс М. Дж., Клингельхутц А. (2003). «Нулевые мутации p53 связаны с теломеразонегативным фенотипом при карциноме яичников». Биология и терапия рака. 1 (5): 511–7. Дои:10.4161 / cbt.1.5.167. PMID12496479. S2CID45381814.