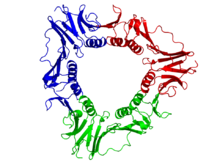
Вид сверху и сбоку гомотример человека PCNA скользящий зажим (цвет радуги, N-конец = синий, C-конец = красный) с двухнитевым ДНК моделируется через центральную пору (маджента).[1]
Крио-ЭМ структура ДНК-связанного процессивного комплекса PolD – PCNA
Структурная основа связывания ДНК комплексом PolD – PCNA
А Зажим ДНК, также известный как скользящий зажим или β-зажим, является белковый комплекс что служит процессивность фактор продвижения Репликация ДНК. В качестве важнейшего компонента Холофермент ДНК-полимераза III, фиксирующий белок связывает ДНК-полимераза и предотвращает это фермент от отделения от шаблона ДНК прядь. Зажим-полимераза белок-белковые взаимодействия являются более сильными и специфичными, чем прямые взаимодействия между полимеразой и цепью матричной ДНК; потому что один из шаги, ограничивающие скорость в реакции синтеза ДНК происходит ассоциация полимеразы с матрицей ДНК, наличие скользящего зажима резко увеличивает количество нуклеотиды что полимераза может добавить к растущей нити за одно событие ассоциации. Наличие зажима ДНК может увеличить скорость синтеза ДНК до 1000 раз по сравнению с непроцессивной полимеразой.[2]
Зажимная складка ДНК представляет собой α + β белок, который собирается в мультимерную структуру, которая полностью окружает двойную спираль ДНК, когда полимераза добавляет нуклеотиды к растущей прядке.[3] Зажим ДНК собирается на ДНК в вилка репликации и «скользит» по ДНК вместе с продвигающейся полимеразой, чему способствует слой воды молекулы в центральной поре зажима между ДНК и поверхностью белка. Из-за тороидальный форма собранного мультимера, зажим не может отделиться от цепи шаблона, не диссоциируя на мономеры.
Зажимная складка ДНК находится в бактерии, археи, эукариоты и немного вирусы. У бактерий скользящий зажим представляет собой гомодимер состоит из двух идентичных бета-субъединиц ДНК-полимераза III и поэтому называется бета-зажимом. В архее[4] и эукариот, это тример, состоящий из трех молекул PCNA. В Бактериофаг Т4 также используется скользящий зажим, называемый gp45, который представляет собой тример, сходный по структуре с PCNA, но не имеющий гомологии последовательности ни с PCNA, ни с бактериальным бета-зажимом.[3]
В бета-зажим является специфическим зажимом ДНК и субъединицей ДНК-полимераза III холоэнзим содержится в бактериях. Две бета-субъединицы собираются вокруг ДНК посредством гамма-субъединицы и гидролиза АТФ; эта сборка называется предпусковой комплекс. После сборки вокруг ДНК сродство бета-субъединиц к субъединице гамма заменяется сродством к альфа- и эпсилон-субъединицам, которые вместе создают полный холофермент.[6][7][8] ДНК-полимераза III - это первичный ферментный комплекс, участвующий в прокариотическийРепликация ДНК.
Гамма-комплекс ДНК-полимеразы III, состоящий из субъединиц γδδ'χψ, катализирует АТФ шаперон двух бета-субъединиц для связывания с ДНК. После связывания с ДНК бета-субъединицы могут свободно скользить по двухцепочечной ДНК. Бета-субъединицы, в свою очередь, связывают комплекс полимеразы αε. Субъединица α обладает ДНК-полимераза активности, а субъединица ε представляет собой 3’-5 ’ экзонуклеаза.[8]
Бета-цепь бактериальной ДНК-полимеразы III состоит из трех топологически эквивалентных домены (N-концевой, центральный и C-терминал ). Две молекулы бета-цепи тесно связаны, образуя замкнутое кольцо, окружающее дуплексную ДНК.
Определенный НПВП (карпрофен, бромфенак и ведапрофен) демонстрируют некоторое подавление репликации бактериальной ДНК за счет ингибирования зажима бактериальной ДНК.[9]
Скользящий зажим у эукариот собирается из определенной субъединицы ДНК-полимераза дельта называется ядерным антигеном пролиферирующих клеток (PCNA ). В N-концевой и C-терминал домены PCNA топологически идентичны. Три молекулы PCNA тесно связаны, образуя замкнутое кольцо, охватывающее дуплекс ДНК.
Последовательность PCNA хорошо сохраняется у растений, животных и грибов, что указывает на сильное избирательное давление для сохранения структуры и предполагает, что этот тип механизма репликации ДНК сохраняется у эукариот.[11][12] Гомологи PCNA также были идентифицированы в археи (Euryarchaeota и Crenarchaeota ) и в Paramecium bursaria Chlorella virus 1 (PBCV-1) и в вирусы ядерного полиэдроза.
Белок субъединицы скользящего зажима вируса gp45 содержит два домена. Каждый домен состоит из двух альфа-спиралей и двух бета-листов - складка дублирована и имеет внутреннюю псевдодвойную симметрию.[14] Три молекулы gp45 тесно связаны с образованием замкнутого кольца дуплексной ДНК.
Скользящие зажимы загружаются на связанные с ними цепи матрицы ДНК с помощью специализированных белков, известных как "погрузчики с подвижным зажимом ", которые также разбирают зажимы после завершения репликации. Сайты связывания этих белков-инициаторов перекрываются с сайтами связывания ДНК-полимеразы, поэтому зажим не может одновременно связываться с загрузчиком зажима и с полимеразой. Таким образом, зажим не будет активно разбирается, в то время как полимераза остается связанной. Зажимы ДНК также связаны с другими факторами, участвующими в гомеостазе ДНК и генома, такими как нуклеосома коэффициенты сборки, Фрагмент Окадзаки лигазы и Ремонт ДНК белки. Все эти белки также имеют общий сайт связывания на зажиме ДНК, который перекрывается с сайтом загрузчика зажима, гарантируя, что зажим не будет удален, пока какой-либо фермент все еще работает с ДНК. Работа погрузчика зажима требует Гидролиз АТФ чтобы «закрыть» зажим вокруг ДНК.
^Мизрахи В., Генри Р.Н., Марлиер Дж. Ф., Джонсон К. А., Бенкович С. Дж. (Июль 1985 г.). «Ограничивающие скорость стадии в пути реакции ДНК-полимеразы I». Биохимия. 24 (15): 4010–8. Дои:10.1021 / bi00336a031. PMID3902078.
^PDB: 1CZD; Моарефи I, Джерузалми Д., Тернер Дж., О'Доннелл М., Куриян Дж. (Март 2000 г.). «Кристаллическая структура фактора процессивности ДНК-полимеразы бактериофага Т4». J. Mol. Биол. 296 (5): 1215–23. Дои:10.1006 / jmbi.1999.3511. PMID10698628.
^Steitz TA, Shamoo Y (1999). «Построение реплисомы из взаимодействующих частей: скользящий зажим в комплексе с пептидом из ДНК-полимеразы и комплексом редактирования полимеразы». Клетка. 99 (2): 155–166. Дои:10.1016 / S0092-8674 (00) 81647-5. PMID10535734. S2CID18103622.
Уотсон Дж. Д., Бейкер Т. А., Белл С. П., Ганн А., Левин М., Лосик Р. (2004). Молекулярная биология гена. Сан-Франциско: Пирсон / Бенджамин Каммингс. ISBN978-0-8053-4635-0.