Россманн фолд - Rossmann fold
| Россманноподобная альфа / бета / альфа-сэндвич-складка | |
|---|---|
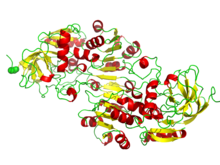 НАД / НАДФ связываются с доменами россмановской складки. На рисунке изображен бета-альфа-фолдинг в алкогольдегидрогеназе. | |
| Идентификаторы | |
| Символ | Россманн-like_a / b / a_fold |
| Pfam клан | CL0039 |
| ИнтерПро | IPR014729 |
В Россманн фолд третичная складка, найденная в белки это связывает нуклеотиды, например, фермент кофакторы FAD, НАД+, и НАДФ+. Эта складка состоит из чередующихся бета-нити и альфа спиральный сегменты, в которых бета-нити связаны водородными связями друг с другом, образуя протяженную бета-лист а альфа-спирали окружают обе стороны листа, образуя трехслойный сэндвич. Классическая складка Россманна содержит шесть бета-тяжей, тогда как складки, подобные Россманну, иногда называют Россманоидные складки, содержат всего пять нитей. Начальная бета-альфа-бета (bab) складка является наиболее консервативным сегментом складки Россмана.[1] Мотив назван в честь Майкл Россманн кто первым заметил этот структурный мотив в ферменте лактатдегидрогеназа в 1970 г. и который позже заметил, что это часто встречающийся мотив в нуклеотидсвязывающих белках.[2]
История
Складку Россманна впервые описал д-р. Майкл Россманн и коллеги в 1974 году.[3] Он был первым, кто установил структуру лактатдегидрогеназы и охарактеризовал структурный мотив внутри этого фермента, который позже будет назван складкой Россмана. Впоследствии было обнаружено, что большинство дегидрогеназ, которые используют НАД или НАДФ, содержат этот же структурно консервативный мотив складки Россмана.[3][4]
В 1989 г. Исраэль Ханукоглу от Институт науки Вейцмана обнаружил, что консенсусная последовательность НАДФ+ сайт связывания в некоторых ферментах, которые используют НАДФ+ отличается от НАД+ обязательный мотив.[5] Это открытие было использовано для изменения коферментной специфичности ферментов.[6]
Структура


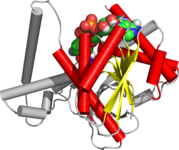
Складка Россманна состоит из шести параллельных бета-нити которые образуют расширенный бета-лист. Первые три пряди соединены между собой α- спирали в результате получается структура бета-альфа-бета-альфа-бета. Этот образец дублируется один раз, чтобы получить перевернутый тандемный повтор, содержащий шесть нитей. В целом жилы расположены в порядке 321456 (1 = N-концевой, 6 = C-концевой).[7] Пять многожильных складок типа Россманна расположены в порядке 32145.[8] Общая третичная структура складки напоминает трехслойный сэндвич, в котором начинка состоит из удлиненного бета-листа, а два ломтика хлеба образованы соединяющимися параллельными альфа-спиралями.[1]
Одна из особенностей складки Россманна - это ее кофактор специфичность связывания. Путем анализа четырех НАДН-связывающих ферментов было обнаружено, что во всех четырех ферментах нуклеотидный кофактор имеет одинаковую конформацию и ориентацию по отношению к полипептидной цепи.[1]
Сгиб может содержать дополнительные нити, соединенные короткими спиралями или витками.[1] Наиболее консервативный сегмент складок Россмана - это первый бета-альфа-бета сегмент. Поскольку этот сегмент контактирует с ADP часть динуклеотидов, таких как FAD, НАД и НАДФ его также называют «бета-бета-складкой, связывающей АДФ».
Функция
Функция укладки Россмана в ферментах заключается в связывании кофакторов нуклеотидов. Он также часто способствует связыванию субстрата.
Метаболические ферменты обычно выполняют одну конкретную функцию, а в случае UDP-глюкозо 6-дегидрогеназа, основная функция - катализировать двухстадийное NAD (+) - зависимое окисление UDP-глюкоза в UDP-глюкуроновая кислота.[9] N- и C-концевые домены UgdG имеют общие структурные особенности с древними митохондриальными рибонуклеазами, названными MAR. MAR присутствуют в низших эукариотических микроорганизмах, имеют Россманноидную складку и принадлежат к надсемейству изохоризматаз. Это наблюдение подтверждает, что структурные мотивы Россмана, обнаруженные в NAD (+) - зависимых дегидрогеназах, могут выполнять двойную функцию, работая как домен связывания нуклеотидного кофактора и как рибонуклеаза.
Эволюция
Эволюционная связь между складками Россмана и Россманн-подобными складками неясна. Эти складки получили название россманноидов. Была выдвинута гипотеза, что все эти складки, включая складку Россмана, произошли от единой общей предковой складки, которая помимо неспецифической каталитической активности обладала способностью связывания нуклеотидов.[3]
Филогенетический анализ фермента, связывающего НАДФ адренодоксинредуктаза выявили, что от прокариот до метазоа и вплоть до приматов отличие мотивов последовательности от большинства сайтов связывания FAD и NAD является строго консервативным.[10]
Во многих статьях и учебниках фолд Россмана определяется как строго повторяющаяся серия структуры βαβ. Тем не менее, всестороннее изучение складок Россмана во многих сайтах связывания NAD (P) и FAD показало, что строго консервативна только первая βα структура. В некоторых ферментах может быть много петель и несколько спиралей (то есть не одна спираль) между бета-цепями, которые образуют бета-лист.[1][10] Несмотря на это расхождение, консервативная последовательность и структурные особенности указывают на общее эволюционное происхождение складок Россмана.[10]
Рекомендации
- ^ а б c d е Ханукоглу I (2015). «Proteopedia: Rossmann fold: бета-альфа-бета складка на сайтах связывания динуклеотидов». Биохимия и молекулярная биология образование. 43 (3): 206–9. Дои:10.1002 / bmb.20849. PMID 25704928.
- ^ Кокс ММ, Нельсон Д.Л. (2013). Принципы биохимии Ленингера (6-е изд.). Нью-Йорк: W.H. Фримен. ISBN 978-1-4292-3414-6.
- ^ а б c Кессель А (2010). Введение в белки: структура, функции и движение. Флорида: CRC Press. п. 143. ISBN 978-1-4398-1071-2.
- ^ Рао С.Т., Россманн М.Г. (май 1973 г.). «Сравнение супервторичных структур в белках». Журнал молекулярной биологии. 76 (2): 241–56. Дои:10.1016/0022-2836(73)90388-4. PMID 4737475.
- ^ Ханукоглу И., Гутфингер Т. (март 1989 г.). «Последовательность кДНК адренодоксинредуктазы. Идентификация сайтов связывания НАДФ в оксидоредуктазах» (PDF). Европейский журнал биохимии. 180 (2): 479–84. Дои:10.1111 / j.1432-1033.1989.tb14671.x. PMID 2924777.
- ^ Скраттон Н.С., Берри А., Перхэм Р.Н. (январь 1990 г.). «Перепроектирование коферментной специфичности дегидрогеназы с помощью белковой инженерии». Природа. 343 (6253): 38–43. Дои:10.1038 / 343038a0. PMID 2296288.
- ^ «NAD (P) -связывающие Россманновские складчатые домены». SCOP: структурная классификация белков.
- ^ «Нуклеотид-связывающий домен». SCOP: структурная классификация белков.
- ^ Бхаттачарья М., Упадхьяй Р., Вишвешвара С. (2012). «Сигнатуры взаимодействия, стабилизирующие NAD (P) -связывающую складку Россмана: подход структурной сети». PLOS ONE. 7 (12): e51676. Дои:10.1371 / journal.pone.0051676. ЧВК 3524241. PMID 23284738.
- ^ а б c Ханукоглу I (2017). «Сохранение интерфейсов фермент-кофермент в FAD и NADP-связывающем адренодоксинредуктазе-А повсеместном ферменте». Журнал молекулярной эволюции. 85 (5): 205–218. Дои:10.1007 / s00239-017-9821-9. PMID 29177972.