Сперматозоид - Википедия - Spermatozoon
| Сперматозоид | |
|---|---|
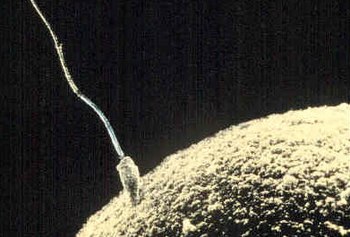 Сперматозоид пытается проникнуть в яйцеклетка пальто к удобрять Это. | |
 Схема человеческого сперматозоида | |
| Подробности | |
| Идентификаторы | |
| латинский | сперматозоид |
| MeSH | D013094 |
| Анатомическая терминология | |
А сперматозоид (произносится /ˌsпɜːrмæтəˈzoʊəп/, альтернативное написание сперматозоид; множественное число сперматозоиды; из Древнегреческий: σπέρμα («семя») и Древнегреческий: ζῷον ("живое существо")) является подвижный сперма клетка, или подвижная форма гаплоидный клетка это мужчина гамета. Сперматозоид присоединяется ан яйцеклетка сформировать зигота. (Зигота - это отдельная клетка с полным набором хромосомы, который обычно превращается в эмбрион.)
Сперматозоиды составляют примерно половину ядерной генетическая информация к диплоид потомство (исключая, в большинстве случаев, митохондриальная ДНК ). У млекопитающих секс потомства определяется сперматозоидом: сперматозоид, несущий Х хромосома приведет к женский (XX) потомство, один из которых Y-хромосома приведет к мужской (XY) потомство. Сперматозоиды впервые были обнаружены у Антони ван Левенгук лаборатория в 1677 году.[1]
Строение, функции и размер сперматозоидов млекопитающих
Люди
Сперматозоид человека является репродуктивной клеткой у мужчин и может выжить только в теплой среде; как только он покидает мужское тело, вероятность выживания сперматозоидов снижается, и они могут погибнуть, тем самым уменьшая общее качество спермы. Сперматозоиды бывают двух типов: «женские» и «мужские». Сперматозоиды, дающие начало потомству женского пола (XX) после оплодотворения, отличаются тем, что несут X-хромосому, а сперматозоиды, дающие потомство мужского пола (XY), несут Y-хромосому.
Сперматозоид человека состоит из плоской головки в форме диска 5.1.мкм на 3,1 мкм и хвостик длиной 50 мкм.[2] Хвост жгутиконосцы, который продвигает сперматозоиды (у человека со скоростью примерно 1–3 мм / мин), взбивая их по эллиптическому конусу.[3] У спермы есть обоняние механизм наведения, и, достигнув фаллопиевых труб, должен пройти период емкости до проникновения в яйцеклетку.[4]
Голова: Он имеет компактное ядро, содержащее только хроматическое вещество, и окружено только тонким ободком цитоплазмы. Над ядром находится шапкообразная структура, называемая акросома, образованный в результате модификации тела Гольджи, секретирующего фермент спермлизин (гиалуронидаза, проникающий в корону фермент, zona eyesin или аэрозин), которые необходимы для оплодотворения. Акросомальная область экспериментирует с акросомной реакцией, она заключается в слиянии плазматической мембраны сперматозоида с внешней акросомной мембраной.[5] На поверхности головы лежит обезвреживающее вещество, которое удаляется перед оплодотворением.
Шея: Это самая маленькая часть (0,03 × 10−6 м) и имеет проксимальную центриоль параллельно основанию ядра и дистально центриоль перпендикулярно предыдущему. Проксимальная центриоль присутствует также в зрелом сперматозоиде; дистальная центриоль исчезает после сборки аксонемы. Проксимальная центриоль входит в яйцеклетку во время оплодотворения и начинает первое деление яйцеклетки, не имеющее центриоли. Дистальная центриоль дает начало осевой нити, которая образует хвост и имеет расположение (9 + 2). Переходная мембрана, называемая Manchette лежит в средней части.
Средняя часть: Он имеет 10–14 спиралей митохондрий, окружающих аксиальный филамент в цитоплазме. Он обеспечивает подвижность и поэтому называется электростанцией спермы. Он также имеет кольцевую центриоль (кольцо), которая образует диффузионный барьер между средней частью и основной частью и служит стабилизирующей структурой для жесткости хвостовой части.[6]
Хвост: Это самая длинная часть (50 × 10−6 м), имеющий аксиальный филамент, окруженный цитоплазмой и плазматической мембраной, но на заднем конце аксиальный филамент голый. Это толкающий механизм.
Сперма имеет щелочную природу, и сперматозоиды не достигают полной подвижности (гипермобильности), пока не достигнут влагалище, где щелочной pH нейтрализуется кислыми влагалищными жидкостями. Этот постепенный процесс занимает 20–30 минут. В течение этого периода, фибриноген от семенные пузырьки образует сгусток, закрепляющий и защищающий сперму. Так же, как они становятся гипермобильными, фибринолизин от простата растворяет сгусток, обеспечивая оптимальное развитие сперматозоидов.
Сперматозоид характеризуется минимумом цитоплазма и самая плотно упакованная ДНК, известная в эукариоты. В сравнении с митотический хромосомы в соматические клетки, ДНК сперматозоидов как минимум в шесть раз более конденсирована.[7]
Образец способствует ДНК /хроматин, а центриоль, и, возможно, также ооцит -активирующий фактор (OAF).[8] Это также может способствовать отцовскому информационная РНК (мРНК), также способствующая эмбриональному развитию.[8]

Электрон микрофотография сперматозоидов человека увеличены в 3140 раз.

Сперматозоиды в образце мочи 45-летнего пациента мужского пола, которого наблюдают с диагнозом: доброкачественная гиперплазия простаты.

Размеры головки человеческого сперматозоида измерены у 39-летнего здорового человека.



Сперматозоид человека содержит не менее 7500 различных белки.[9]
Генетика спермы человека связана с эволюция человека, согласно исследованию 2020 года.[10][11][12]
Повреждение и восстановление ДНК
Наличие повреждений ДНК в сперматозоидах в период после мейоз но прежде оплодотворение может быть восстановлен в оплодотворенной яйцеклетке, но, если его не исправить, может иметь серьезные пагубные последствия для фертильности и развивающегося эмбриона. Сперматозоиды человека особенно уязвимы для атаки свободных радикалов и образования окислительного повреждения ДНК.[13][14] (см., например, 8-оксо-2'-дезоксигуанозин )
Воздействие на мужчин определенного образа жизни, окружающей среды или профессиональных опасностей может увеличить риск анеуплоидный сперматозоиды.[15] В частности, риск анеуплоидии увеличивается при курении табака,[16][17] и профессиональное воздействие бензола,[18] инсектициды,[19][20] и перфторированные соединения.[21] Повышенная анеуплоидия сперматозоидов часто возникает в связи с повышенным повреждением ДНК. Фрагментация ДНК и повышенная восприимчивость ДНК к денатурации in situ. апоптоз соматических клеток, характеризуют аномальные сперматозоиды в случаях мужское бесплодие.[22][23]
Избежание реакции иммунной системы
Гликопротеин Молекулы на поверхности эякулированных сперматозоидов распознаются всеми женскими иммунными системами человека и интерпретируются как сигнал о том, что клетка не должна отторгаться. В противном случае женская иммунная система могла бы атаковать сперму в половые пути. Специфические гликопротеины, покрывающие сперматозоиды, также используются некоторыми раковыми и бактериальными клетками, некоторыми паразитическими червями и ВИЧ-инфицированными лейкоцитами, что позволяет избежать иммунного ответа со стороны организм хозяина.[24]
В гемато-яичковый барьер, поддерживаемые плотными стыками между Клетки Сертоли семенных канальцев, предотвращает связь между образующимися сперматозоидами в семенниках и кровеносными сосудами (и иммунными клетками, циркулирующими в них) внутри межстраничное пространство. Это не дает им вызвать иммунный ответ. Барьер кровь-яички также важен для предотвращения нарушения сперматогенеза токсичными веществами.
Сперматозоиды в других организмах

Животные
Оплодотворение зависит от сперматозоидов большинства животных с половым размножением.
Некоторые виды плодовая муха производят самый крупный из известных в природе сперматозоидов.[25][26] Drosophila melanogaster производит сперму размером до 1,8 мм,[27] в то время как его родственник Дрозофила бифурка производит самый крупный из известных сперматозоидов, его длина превышает 58 мм.[25] У Drosophila melanogaster весь сперматозоид, включая хвост, включается в ооцит цитоплазма однако у Drosophila bifurca в ооцит входит лишь небольшая часть хвоста.[28]
Деревянная мышь Apodemus sylvaticus имеет сперматозоиды серповидной морфологии. Еще одна особенность, которая делает эти гаметоциты уникальными, - это наличие апикального крючка на головке сперматозоида. Этот крючок используется для прикрепления к крючкам или жгутикам других сперматозоидов. Агрегация вызывается этими навесными приспособлениями и результатом подвижных поездов. Эти поезда улучшают моторику женских репродуктивных путей и являются средством, способствующим оплодотворению.[29]
Постмейотическая фаза сперматогенеза мышей очень чувствительна к окружающей среде. генотоксичный агентов, потому что по мере того, как мужские половые клетки образуют зрелые сперматозоиды, они постепенно теряют способность восстанавливать повреждения ДНК.[30] Облучение мышей-самцов во время позднего сперматогенеза может вызвать повреждение, которое сохраняется не менее 7 дней в оплодотворяющих сперматозоидах, а нарушение путей восстановления двухцепочечных разрывов материнской ДНК увеличивает хромосомные аберрации, происходящие из сперматозоидов.[31] Лечение мышей-самцов с мелфалан, бифункциональный алкилирующий агент часто применяемый в химиотерапии, вызывает повреждения ДНК во время мейоза, которые могут сохраняться в неизлеченном состоянии по мере того, как половые клетки проходят через компетентные к репарации ДНК фазы сперматогенного развития.[32] Такие неизлечимые повреждения ДНК в сперматозоидах после оплодотворения могут привести к появлению у потомства различных аномалий.
Морские ежи Такие как Arbacia punctulata являются идеальными организмами для использования в исследовании сперматозоидов, они порождают большое количество сперматозоидов в море, что делает их хорошо подходящими в качестве модельные организмы для экспериментов.[нужна цитата ]
Сперматозоиды сумчатые обычно длиннее, чем у плацентарные млекопитающие.[33]
Растения, водоросли и грибы
В гаметофиты из мохообразные, папоротники и немного голосеменные производить подвижный сперма клетки, в отличие от пыльца зерна, используемые у большинства голосеменных и всех покрытосеменные. Это обеспечивает половое размножение при отсутствии воды невозможно, поскольку вода является необходимой средой для встречи сперматозоидов и яйцеклетки. Сперматозоиды водорослей и низших растений часто имеют множество жгутиков (см. Изображение) и поэтому морфологически отличаются от сперматозоидов животных.
Некоторые водоросли и грибы производят неподвижные сперматозоиды, называемые спермиями. У высших растений, некоторых водорослей и грибов оплодотворение включает миграцию ядра сперматозоидов через трубку для оплодотворения (например, пыльцевая трубка у высших растений), чтобы достичь яйцеклетки.
Производство сперматозоидов у млекопитающих
Сперматозоиды производятся в семенные канальцы из яички в процессе, называемом сперматогенезом. Круглые клетки называются сперматогония делятся и дифференцируются в конечном итоге, чтобы стать сперматозоидами. В течение совокупление то клоака или же влагалище получает оплодотворенный, а затем сперматозоиды проходят через хемотаксис к яйцеклетке внутри Фаллопиевых труб или матка.
Активация сперматозоидов
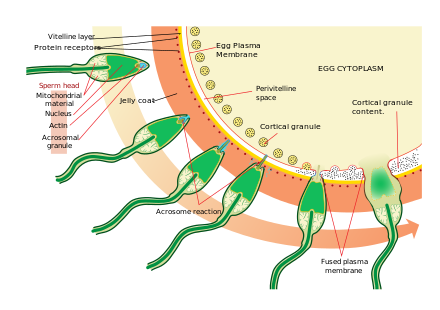
Приближение к яйцеклетке - довольно сложный, многоэтапный процесс. хемотаксис руководствуясь различными химическими веществами / стимулами на отдельных уровнях филогении. Одним из наиболее значимых и распространенных сигнальных знаков события является то, что прототип профессиональных рецепторов хемотаксиса, рецептор формилового пептида (60000 рецепторов на клетку), а также активаторная способность его лиганда формил Met-Leu-Phe была продемонстрирована в поверхностной мембране даже в случае спермы человека.[34] Сперматозоиды млекопитающих становятся еще более активными, когда они приближаются к яйцеклетке в процессе, называемом активация спермы. Было показано, что активация спермы вызывается: кальций ионофоры in vitro, прогестерон выпущен поблизости кучевые клетки и привязка к ZP3 из zona pellucida. В кучевые клетки заключены в гелеобразное вещество, состоящее в основном из гиалуроновой кислоты, развиваются в яичнике вместе с яйцеклеткой и поддерживают его при росте.
Первоначальное изменение называется «гиперактивацией», которое вызывает изменение подвижности сперматозоидов. Они плавают быстрее, а их движения хвоста становятся более резкими и беспорядочными.
Недавнее открытие связывает гиперактивацию с внезапным притоком ионов кальция в хвосты. Хлыстоподобный хвост (жгутик) сперматозоидов усыпан ионные каналы образованный белками, называемыми CatSper. Эти каналы избирательны, пропускают только ионы кальция. Открытие каналов CatSper отвечает за приток кальция. Внезапное повышение уровня кальция приводит к тому, что жгутик формирует более глубокие изгибы, продвигая сперматозоиды с большей силой через вязкую среду. Гиперактивность сперматозоидов необходима для преодоления двух физических барьеров, защищающих яйцеклетку от оплодотворения.
Второй процесс активации сперматозоидов - это акросомная реакция. Это включает высвобождение содержимого акросомы, которое рассеивается, и воздействие ферментов, прикрепленных к внутренней акросомной мембране сперматозоидов. Это происходит после того, как сперма впервые встречается с яйцеклеткой. Этот механизм типа «замок и ключ» зависит от вида и предотвращает слияние сперматозоидов и яйцеклеток разных видов. Есть некоторые свидетельства того, что именно эта привязка запускает акросома для высвобождения ферментов, которые позволяют сперматозоиду сливаться с яйцеклеткой.
ZP3, один из белков, составляющих блестящую оболочку, затем связывается с молекулой-партнером в сперме. Ферменты на внутренней акросомной мембране переваривают блестящую оболочку. После того, как сперматозоид проникает через блестящую зону, часть мембраны сперматозоидов, затем предохранители с мембраной яйцеклетки, и содержимое головки диффундирует в яйцо.
Считается, что после проникновения ооцит стал активирован. Он подвергается вторичному мейотическому делению, и два гаплоидных ядра (отцовское и материнское) сливаются с образованием зигота. Чтобы предотвратить полиспермия и свести к минимуму возможность получения триплоид zygote, несколько изменений в пеллюцидной зоне яйца делают их непроницаемыми вскоре после того, как в яйцеклетку попадает первый сперматозоид.
Искусственное хранилище
Сперматозоиды можно хранить в разбавителях, таких как Переменная температура Иллини (IVT) разбавитель, который, как сообщается, способен сохранять высокую фертильность сперматозоидов в течение более семи дней.[35] Разбавитель для IVT состоит из нескольких солей, сахаров и антибактериальных агентов и насыщен газами. CO2.[35]
Криоконсервация спермы можно использовать для более длительного хранения. Для сперматозоидов человека самый длительный успешный срок хранения с использованием этого метода - 21 год.[36]
История
- В 1677 г. микробиолог Антони ван Левенгук обнаружил сперматозоиды.
- В 1841 году швейцарский анатом Альберт фон Кёлликер писал о сперматозоидах в своей работе Untersuchungen über die Bedeutung der Samenfäden (Исследования о важности сперматозоидов).
Смотрите также
Рекомендации
- ^ «Хронология: вспомогательная репродукция и контроль рождаемости». CBC Новости. Получено 2006-04-06.
- ^ Смит, Д.Дж. (2009). «Скопление человеческой спермы у поверхности: исследование с помощью моделирования» (PDF). Журнал гидромеханики. 621: 295. Bibcode:2009JFM ... 621..289S. Дои:10.1017 / S0022112008004953. Получено 20 мая 2012.
- ^ Исидзима, Сумио; Ошио, Сигеру; Мохри, Хидео (1986). "Жгутиковые движения сперматозоидов человека". Gamete Research. 13 (3): 185–197. Дои:10.1002 / мрд.1120130302.
- ^ Лаура Гиохалас.Направление спермы у млекопитающих Nature Reviews: молекулярная клеточная биология. Май 2006 г.
- ^ дель Рио, Мария Хосе; Годой, Ана; Торо, Алехандра; Орельяна, Ренан; Кортес, Мануэль Э .; Морено, Рикардо Д.; Vigil, Пилар (октябрь 2007 г.). "La reacción acrosómica del espermatozoide: avances recientes". Revista Internacional de Andrología. 5 (4): 368–373. Дои:10.1016 / S1698-031X (07) 74086-4.
- ^ "кольцо спермы | SGD". www.yeastgenome.org. Получено 2019-02-22.
- ^ Уорд WS, Коффи Д.С. (1991). «Упаковка и организация ДНК в сперматозоидах млекопитающих: сравнение с соматическими клетками». Биология размножения. 44 (4): 569–74. Дои:10.1095 / биолрепрод44.4.569. PMID 2043729.
- ^ а б Херардо Баррозу, доктор медицинских наук, магистр наук, Карлос Вальдеспин, доктор медицинских наук, Ева Вега, магистр наук, Рубен Кершенович, доктор медицинских наук, Розаура Авила, бакалавр наук, Конрадо Авенданьо, доктор медицинских наук, Серджио Энингер, доктор медицинских наук, доктор философии .Db Вклад сперматозоидов в развитие: оплодотворение и не только Фертильность и бесплодие, Том 92, выпуск 3, страницы 835-848 (сентябрь 2009 г.)
- ^ Amaral, A .; Castillo, J .; Ramalho-Santos, J .; Олива, Р. (2013). «Комбинированный протеом спермы человека: клеточные пути и значение для фундаментальной и клинической науки». Обновление репродукции человека. 20 (1): 40–62. Дои:10.1093 / humupd / dmt046. PMID 24082039.
- ^ https://www.cell.com/cell/pdf/S0092-8674(19)31377-7.pdf
- ^ https://phys.org/news/2020-01-scanning-sperm-human-evolution.html
- ^ https://markets.businessinsider.com/news/stocks/genetic-scanning-system-in-sperm-may-control-rate-of-human-evolution-1028840950
- ^ Гаврилюк, Дан; Эйткен, Роберт Джон (2015). «Повреждение ДНК сперматозоидов, опосредованное реактивными видами кислорода: его влияние на репродуктивную способность человека и траекторию здоровья потомства». Роль мужчин в потере беременности и неудачной имплантации эмбриона. Успехи экспериментальной медицины и биологии. 868. С. 23–47. Дои:10.1007/978-3-319-18881-2_2. ISBN 978-3-319-18880-5. PMID 26178844.
- ^ Lozano, G.M .; Bejarano, I .; Espino, J .; González, D .; Ортис, А .; García, J.F .; Родригес, А.Б .; Париенте, Дж. (2009). «Капситация с градиентом плотности - наиболее подходящий метод для улучшения оплодотворения и снижения фрагментации ДНК сперматозоидов бесплодных мужчин». Анатолийский журнал акушерства и гинекологии. 3 (1): 1–7.
- ^ Templado C, Uroz L, Estop A (2013). «Новые взгляды на происхождение и актуальность анеуплоидии в сперматозоидах человека». Молекулярная репродукция человека. 19 (10): 634–43. Дои:10,1093 / мольхр / gat039. PMID 23720770.
- ^ Ши Кью, Ко Э, Барклай Л., Хоанг Т., Радемейкер А., Мартин Р. (2001). «Курение сигарет и анеуплоидия в сперме человека». Молекулярное воспроизводство и развитие. 59 (4): 417–21. Дои:10.1002 / мрд.1048. PMID 11468778.
- ^ Рубес Дж, Лоу X, Мур Д., Перро С., Слот V, Эвенсон Д., Селеван С.Г., Вайробек А.Дж. (1998). «Курение сигарет связано с повышенной дисомией сперматозоидов у мужчин-подростков». Фертильность и бесплодие. 70 (4): 715–23. Дои:10.1016 / S0015-0282 (98) 00261-1. PMID 9797104.
- ^ Xing C, Marchetti F, Li G, Weldon RH, Kurtovich E, Young S, Schmid TE, Zhang L, Rappaport S, Waidyanatha S, Wyrobek AJ, Eskenazi B (2010). «Воздействие бензола вблизи допустимого предела в США связано с анеуплоидией сперматозоидов». Перспективы гигиены окружающей среды. 118 (6): 833–9. Дои:10.1289 / ehp.0901531. ЧВК 2898861. PMID 20418200.
- ^ Ся Й, Бянь Q, Сюй Л., Ченг С., Сонг Л., Лю Дж., Ву В., Ван С., Ван Х (2004). «Генотоксическое воздействие на сперматозоиды человека у рабочих завода по производству пестицидов, подвергшихся воздействию фенвалерата». Токсикология. 203 (1–3): 49–60. Дои:10.1016 / j.tox.2004.05.018. PMID 15363581.
- ^ Ся И, Ченг С., Биан Кью, Сюй Л., Коллинз, доктор медицины, Чанг Х.С., Сон Л., Лю Дж., Ван С., Ван Х (2005). «Генотоксическое воздействие на сперматозоиды рабочих, подвергшихся воздействию карбарила». Токсикологические науки. 85 (1): 615–23. Дои:10.1093 / toxsci / kfi066. PMID 15615886.
- ^ Governini L, Guerranti C, De Leo V, Boschi L, Luddi A, Gori M, Orvieto R, Piomboni P (2014). «Хромосомные анеуплоидии и фрагментация ДНК сперматозоидов человека от пациентов, подвергшихся воздействию перфторированных соединений». Андрология. 47 (9): 1012–9. Дои:10.1111 / и.12371. PMID 25382683.
- ^ Горчица, З; Traganos, F; Jesionowska, H; Darzynkiewicz, Z (1993). «Наличие разрывов цепи ДНК и повышенная чувствительность ДНК in situ к денатурации в аномальных сперматозоидах человека. Аналогия апоптозу соматических клеток». Exp Cell Res. 207 (1): 202–205. Дои:10.1006 / excr.1993.1182. PMID 8391465.
- ^ Эвенсон, Д.П .; Darzynkiewicz, Z; Меламед, MR (1980). «Связь неоднородности хроматина сперматозоидов с фертильностью». Наука. 210 (4474): 1131–1133. Bibcode:1980Sci ... 210.1131E. Дои:10.1126 / science.7444440. PMID 7444440.
- ^ "Сперма ключ к иммунитету от болезней"'". Новости BBC. 2007-12-17.
- ^ а б Питник, S; Спайсер, GS; Марков Т.А. (11 мая 1995 г.). «Какова длина гигантской спермы?». Природа. 375 (6527): 109. Bibcode:1995 Натур.375Q.109P. Дои:10.1038 / 375109a0. PMID 7753164. S2CID 4368953.
- ^ Питник, S; Марков Т.А. (27 сентября 1994 г.). «Преимущества крупных самцов, связанные с затратами на производство спермы у Drosophila hydei, вида с гигантскими сперматозоидами». Труды Национальной академии наук Соединенных Штатов Америки. 91 (20): 9277–81. Bibcode:1994PNAS ... 91.9277P. Дои:10.1073 / пнас.91.20.9277. ЧВК 44795. PMID 7937755.
- ^ Купер, К. (1950). Демерец, М. (ред.). Биология дрозофилы. Нью-Йорк: Вили. С. 1–61.
- ^ Pitnick, S .; Спайсер, Г. С .; Марков, Т.А. (1995). «Какова длина гигантской спермы». Природа. 375 (6527): 109. Bibcode:1995 Натур.375Q.109P. Дои:10.1038 / 375109a0. PMID 7753164. S2CID 4368953.
- ^ Мур, H; Дворакова, К; Jenkins, N; Порода, W (2002). «Исключительное сотрудничество спермы в Wood Mouse» (PDF). Природа. 418 (6894): 174–177. Bibcode:2002Натура.418..174М. Дои:10.1038 / природа00832. PMID 12110888. S2CID 4413444.
- ^ Маркетти Ф., Выробек А.Дж. (2008). «Снижение репарации ДНК во время спермиогенеза мышей приводит к накоплению наследственных повреждений ДНК». Ремонт ДНК. 7 (4): 572–81. Дои:10.1016 / j.dnarep.2007.12.011. PMID 18282746.
- ^ Маркетти Ф., Эссерс Дж., Канаар Р., Выробек А. Дж. (2007). «Нарушение репарации материнской ДНК увеличивает хромосомные аберрации сперматозоидов». Труды Национальной академии наук Соединенных Штатов Америки. 104 (45): 17725–9. Bibcode:2007ПНАС..10417725М. Дои:10.1073 / pnas.0705257104. ЧВК 2077046. PMID 17978187.
- ^ Маркетти Ф., Епископ Дж., Джинджерих Дж., Выробек А.Дж. (2015). «Повреждение мейотической межцепочечной ДНК ускользает от отцовской репарации и вызывает хромосомные аберрации в зиготе из-за неправильного материнского ремонта». Научные отчеты. 5: 7689. Bibcode:2015НатСР ... 5Э7689М. Дои:10.1038 / srep07689. ЧВК 4286742. PMID 25567288.
- ^ Ларри Фогельнест; Тимоти Портас (1 мая 2019 г.). Современная терапия австралийских млекопитающих. Csiro Publishing. ISBN 978-1-4863-0752-4.
- ^ Гнесси Л., Фаббри А., Сильвестрони Л., Моретти С., Фрайоли Ф., Перт CB, Исидори А. (1986). «Доказательства наличия специфических рецепторов для N-формилхемотаксических пептидов на сперматозоидах человека». Журнал клинической эндокринологии и метаболизма. 63 (4): 841–6. Дои:10.1210 / jcem-63-4-841. PMID 3018025.
- ^ а б Уотсон, П. Ф. (1993). «Потенциальное влияние технологии инкапсуляции сперматозоидов на важность выбора времени искусственного оплодотворения: перспектива в свете опубликованных работ». Размножение, фертильность и развитие. 5 (6): 691–9. Дои:10.1071 / RD9930691. PMID 9627729.
- ^ НОВОСТИ и пресс-релизы Planer> Ребенок, родившийся после 21 года хранения спермы в морозильной камере с контролируемой скоростью Planer 14/10/2004
внешняя ссылка
| Биологические термины | |
|---|---|
| Сексуальный воспроизведение | |
| Сексуальность | |
| |
| Микроскопические открытия1 |
|  | ||||
|---|---|---|---|---|---|---|
| Общие темы | ||||||
| похожие темы |
| |||||
| Связанные люди |
| |||||
| Признания | ||||||
| ||||||