Инфузория - Ciliate
| Инфузория | |
|---|---|
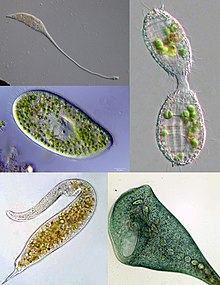 | |
| Некоторые примеры разнообразия инфузорий. По часовой стрелке сверху слева: Лакримария, Coleps, Стентор, Дилепт, Парамеций | |
| Научная классификация | |
| Clade: | SAR |
| Infrakingdom: | Альвеолаты |
| Тип: | Цилиофора Дофлейн, 1901 исправить. |
| Подтипы и классы[1] | |
См. Текст для подклассов. | |
| Синонимы | |
| |
В инфузории группа простейшие характеризуется наличием волосовидных органелл, называемых реснички, которые по структуре идентичны эукариотические жгутики, но в целом они короче и представлены в гораздо большем количестве, с разными волнистый рисунок, чем жгутики. Реснички встречаются у всех членов группы (хотя своеобразный Суктория есть они только для части их жизненный цикл ) и по-разному используются при плавании, ползании, привязанности, кормлении и ощущении.
Инфузории - важная группа протисты, обычен почти везде, где есть вода - в озерах, прудах, океанах, реках и почвах. Описано около 4500 уникальных свободноживущих видов, а потенциальное количество современных видов оценивается в 27 000–40 000.[2] В это число входят многие эктосимбиотический и эндосимбиотический виды, а также некоторые обязать и оппортунистический паразиты. Размеры инфузорий варьируются от 10 до 10 штук. мкм в некоторых кольподеи до 4 мм в длину в некоторых гелейиды, и включать некоторые из самых морфологически сложные простейшие.[3][4]
В большинстве систем таксономия, "Цилиофора"оценивается как филюм[5] под любым из нескольких королевства, в том числе Chromista,[6] Протиста[7] или Простейшие.[8] В некоторых старых системах классификации, таких как влиятельные таксономические работы Альфред Каль мерцательные простейшие помещаются в класс "Реснички"[9][10] (термин, который также может относиться к род рыб ). В таксономической схеме, одобренной Международное общество протистологов, что исключает формальные ранг такие обозначения, как «тип» и «класс», «цилиофора» не имеет рейтинга таксон в пределах Альвеолаты.[11][12]
Структура клетки

Ядра
В отличие от большинства других эукариоты, инфузории имеют два разных типа ядра: крошечный, диплоид микронуклеус («генеративное ядро», несущее зародышевый ячейки), и большой, полиплоид макронуклеус («вегетативное ядро», которое заботится об общей регуляции клеток, выражая фенотип организма). Последний образуется из микроядра путем усиления геном и тяжелое редактирование. Микронуклеус передает свой генетический материал потомству, но не экспрессирует свои гены. Макронуклеус обеспечивает малая ядерная РНК для вегетативного роста.
Деление макронуклеуса происходит за счет амитоз, и сегрегация хромосомы происходит в результате процесса, механизм которого неизвестен. Этот процесс не идеален, и примерно через 200 поколений клетка проявляет признаки старения. Периодически макронуклеусы необходимо регенерировать из микроядер. В большинстве случаев это происходит во время спряжение. Здесь выстраиваются две клетки, микроядра претерпевают мейоз, некоторые из гаплоидный дочери обмениваются, а затем сливаются, образуя новые микроядра и макронуклеусы.
Цитоплазма
Еда вакуоли сформированы через фагоцитоз и обычно следуют определенному пути через ячейку, поскольку их содержимое переваривается и разбивается лизосомы Таким образом, вещества, содержащиеся в вакуоли, достаточно малы, чтобы размытый через мембрану пищевой вакуоли в клетку. Все, что осталось в пищевой вакуоли к моменту достижения цитопрокта (анальная пора ) разряжается экзоцитоз. У большинства инфузорий также есть одна или несколько выдающихся сократительные вакуоли, которые собирают воду и выводят ее из клетки для поддержания осмотическое давление, или в какой-либо функции для поддержания ионного баланса. У некоторых родов, например Парамеций, они имеют характерную форму звезды, причем каждая точка представляет собой собирающую трубку.
Специализированные структуры у инфузорий
Реснички расположены рядами, называемыми кинети. В некоторых формах есть и поликинетиды тела, например, среди спиротрихи где они обычно образуют щетину, называемую Cirri. Чаще реснички тела располагаются в мононуклеоз- и дикинетиды, которые включают соответственно один и два кинетосомы (базальные тельца), каждое из которых может поддерживать ресничку. Они расположены в ряды, называемые кинети, которые идут от передней части клетки к задней. Тело и оральные кинетиды составляют недоразвитый, организация, уникальная для инфузорий и важная для их классификации, и включает различные фибриллы и микротрубочки участвует в координации ресничек.
Инфрацилиатура - один из основных компонентов клеточная кора. Остальные альвеолы, маленькие пузырьки под клеточной мембраной, которые упаковываются напротив нее, образуя пленка поддержание формы клетки, которая варьируется от гибкой и сократительной до жесткой. Многочисленные митохондрии и экструзомы также обычно присутствуют. Наличие альвеол, строение ресничек, форма митоза и различные другие детали указывают на тесную связь между инфузориями, Apicomplexa, и динофлагелляты. Эти внешне непохожие группы составляют альвеолиты.
Кормление
Большинство инфузорий гетеротрофы, питаясь более мелкими организмами, такими как бактерии и водоросли, а детрит попадал в ротовую борозду (рот) модифицированными ресничками ротовой полости. Обычно это включает в себя серию мембранеллы слева от рта и пароральная перепонка справа, оба из которых возникают из поликинетиды, группы многих ресничек вместе с ассоциированными структурами. Пища перемещается ресничками через поры рта в пищевод, в результате чего образуются пищевые вакуоли.
Однако методы кормления значительно различаются. Некоторые инфузории не имеют рта и питаются абсорбцией (осмотрофия ), в то время как другие являются хищниками и питаются другими простейшими и, в частности, другими инфузориями. Некоторые инфузории паразитируют животные, хотя только один вид, Balantidium coli, как известно, вызывает заболевание у людей.[13]
Размножение и сексуальные явления
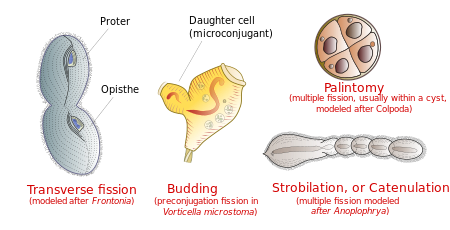
Размножение
Инфузории размножаются бесполым путем, различными видами деление.[14] Во время деления микроядро подвергается митоз а макронуклеус удлиняется и подвергается амитоз (кроме Кариореликтовский инфузории, макронуклеары которых не делятся). Затем клетка делится на две, и каждая новая клетка получает копию микроядра и макронуклеуса.

Обычно ячейка делится поперечно, причем передний половина инфузории ( протер) образуя один новый организм, а задний половина ( Опист) образуя другой. Однако в некоторых группах инфузорий происходят и другие типы деления. Они включают подающий надежды (появление из тела зрелого родителя маленького реснитчатого потомства, или «роевиков»); стробиляция (множественные деления вдоль тела клетки, производящие цепочку новых организмов); и палинтомия (множественные деления, обычно в пределах киста ).[15]
Деление может происходить спонтанно, как часть вегетативного клеточный цикл. Как вариант, это может происходить в результате самооплодотворения (автогамия ),[16] или это может последовать спряжение, сексуальный феномен, при котором инфузории совместимых типы вязки обмениваться генетическим материалом. Хотя конъюгацию иногда называют формой воспроизводства, она не связана напрямую с репродуктивными процессами и не приводит напрямую к увеличению числа отдельных инфузорий или их потомства.[17]
Конъюгация
- Обзор
Спряжение инфузорий - это сексуальное явление, которое приводит к генетическая рекомбинация и ядерная реорганизация внутри клетки.[17][15] Во время конъюгации две инфузории совместимого типа спаривания образуют мост между своими цитоплазмы. Микроядра претерпевают мейоз, макронуклеусы исчезают, и гаплоидный микроядра обмениваются через мост. У некоторых инфузорий (перитрих, хонотрихи и немного сукторианцы ), конъюгированные клетки сливаются навсегда, и один конъюгант поглощается другим.[13][18] Однако в большинстве групп инфузорий клетки отделяются после конъюгации, и оба образуют новые макронуклеары из своих микроядер.[19] Спряжение и автогамия всегда сопровождаются делением.[15]
У многих инфузорий, таких как Парамецийпартнеры по спряжению (гамонты) похожи или неотличимы по размеру и форме. Это называется «изогамонтической» конъюгацией. В некоторых группах партнеры разные по размеру и форме. Это называется «анизогамонтическим» конъюгацией. В сидячие перитрихи например, один половой партнер (микроконъюгант) маленький и подвижный, а другой (макроконъюгант) большой и сидячий.[17]
- Этапы спряжения
 Этапы конъюгации у Paramecium caudatum
Этапы конъюгации у Paramecium caudatum

В Paramecium caudatum, этапы сопряжения следующие (см. диаграмму справа):
- Совместимые деформации спаривания встречаются и частично сливаются
- Микроядра подвергаются мейозу, образуя четыре гаплоидных микроядра на клетку.
- Три из этих микроядер распадаются. Четвертый подвергается митозу.
- Две клетки обмениваются микроядром.
- Затем клетки разделяются.
- Микроядра в каждой клетке сливаются, образуя диплоидное микроядро.
- Митоз возникает трижды, в результате чего образуется восемь микроядер.
- Четыре новых микроядра превращаются в макронуклеусы, а старые макронуклеусы распадаются.
- Двойное деление происходит дважды, образуя четыре идентичные дочерние клетки.
Перестройки ДНК (скремблирование генов)
Инфузории содержат два типа ядер: соматический "макронуклеус "и зародышевый "микронуклеус ". Только ДНК в микроядре передается во время полового размножения (конъюгации). С другой стороны, только ДНК в макронуклеусе активно экспрессируется и определяет фенотип организма. Макроядерная ДНК получена из микроядерной ДНК удивительным образом обширная перестройка и амплификация ДНК.
Макронуклеус начинается как копия микронуклеуса. Микроядерные хромосомы фрагментируются на множество более мелких частей и усиливаются, чтобы получить множество копий. Образующиеся в результате макроядерные хромосомы часто содержат только одну ген. В Тетрахимена у микроядра 10 хромосом (по пять на гаплоидный геном), а у макронуклеуса более 20 000 хромосом.[20]
Кроме того, микроядерные гены прерываются многочисленными «внутренними элиминированными последовательностями» (IES). Во время развития макронуклеуса IES удаляются, а оставшиеся генные сегменты, макронуклеарные последовательности (MDS), сращиваются вместе, чтобы получить рабочий ген. Тетрахимена имеет около 6000 IES, и около 15% микроядерной ДНК удаляется во время этого процесса. В процессе руководствуются малые РНК и эпигенетический хроматин Метки.[20]
В Spirotrich инфузории (такие как Oxytricha ), этот процесс еще более сложен из-за «скремблирования генов»: MDS в микроядре часто имеют другой порядок и ориентацию, чем в макронуклеарном гене, и поэтому помимо делеции ДНК инверсия и перемещение требуются для «расшифровки». Этим процессом управляют длинные РНК, происходящие из родительского макронуклеуса. Более 95% микроядерной ДНК удаляется во время развития макронуклеаров спираотрихов.[20]
Окаменелости
До недавнего времени самые старые известные окаменелости инфузорий были тинтинниды от Ордовикский период. В 2007 году Ли и другие. опубликовали описание ископаемых инфузорий из Формация Доушантуо, около 580 миллионов лет назад, в Эдиакарский период. К ним относятся два типа тинтиннидов и возможный предковый сукториан.[21] Ископаемое Вортичелла был обнаружен внутри кокона пиявки из Триасовый период, около 200 миллионов лет назад.[22]
Классификация

Для инфузорий было предложено несколько различных схем классификации. Следующая схема основана на молекулярном филогенетический анализ до четырех генов 152 видов, представляющих 110 семейств:[1]
Подтип Postciliodesmatophora
- Класс Гетеротричея (например. Стентор )
- Класс Кариореликтея
Subphylum Intramacronucleata

- Класс Armophorea
- Класс Кариакотрихия (только один вид, Кариакотрикс хвостатый )
- Класс Муранотричея
- Класс Parablepharismea
- Класс Кольподея (например. Colpoda )
- Класс Litostomatea
- Подкласс Haptoria (например. Дидиниум )
- Подкласс Ринхостоматия
- Подкласс Трихостоматия (например. Балантидиум )
- Класс Нософорея
- Класс Филлофарингея
- Подкласс Хонотрихия
- Подкласс Киртофория
- Подкласс Ринходия
- Подкласс Суктория (например. Подофира )
- Подкласс Синимения
- Класс Олигогименофорея
- Подкласс Апостоматия
- Подкласс Astomatia
- Подкласс Гименостоматия (например. Тетрахимена )
- Подкласс Пеникулия (например. Парамеций )
- Подкласс Перитрихия (например. Вортичелла )
- Подкласс Scuticociliatia
- Класс Плагиопилея
- Класс Простома (например. Coleps )
- Класс Протокрузия
- Класс Спиротричея
- Подкласс Хореотрихия
- Подкласс Евплотия
- Подкласс Гипотрихия
- Подкласс Ликнофория
- Подкласс Олиготрихия
- Подкласс Phacodiniidea
- Подкласс Протогипотрихия
Другой
Включены некоторые старые классификации Опалиновые у инфузорий. Принципиальное отличие мультикалиатной жгутиконосцы (например., гемимастигиды, Стефанопогон, Multicilia, опалины ) и инфузорий - наличие макронуклеусов только в инфузориях.[23]
Патогенность
Единственный известный представитель филума инфузорий патогенный для людей это Balantidium coli,[24][25] что вызывает болезнь балантидиаз. Не патогенен для домашней свиньи.[26]
использованная литература
- ^ а б Гао, Фэн; Уоррен, Алан; Чжан, Цяньцянь; Гонг, июнь; Мяо, Мяо; Солнце, Пинг; Сюй, Дапэн; Хуанг, Цзе; Йи, Чжэньчжэнь (2016-04-29). «Основанная на всех данных эволюционная гипотеза реснитчатых протистов с пересмотренной классификацией типа Ciliophora (Eukaryota, Alveolata)». Научные отчеты. 6: 24874. Bibcode:2016НатСР ... 624874Г. Дои:10.1038 / srep24874. ISSN 2045-2322. ЧВК 4850378. PMID 27126745.
- ^ Фойсснер, В .; Хоксворт, Дэвид, ред. (2009). Разнообразие протистов и географическое распространение. Темы биоразнообразия и сохранения. Springer Нидерланды. п. 111. ISBN 9789048128006.
- ^ Нильсен, Торкель Гиссель; Kiørboe, Томас (1994). «Регулирование биомассы и продукции зоопланктона в прибрежных экосистемах умеренного пояса. 2. Инфузории». Лимнология и океанография. 39 (3): 508–519. Bibcode:1994LimOc..39..508N. Дои:10.4319 / lo.1994.39.3.0508.
- ^ Линн, Денис (2008). Ресничные простейшие 3-е издание. Springer. стр.129. ISBN 978-1-4020-8238-2.
- ^ «Отчет ИТИС». Интегрированная система таксономической информации. Получено 11 декабря 2014.
- ^ Кавальер-Смит, Томас (01.01.2018). «Kingdom Chromista и его восемь типов: новый синтез, подчеркивающий нацеливание на перипластидные белки, эволюцию цитоскелета и перипластид, а также древние расхождения». Протоплазма. 255 (1): 297–357. Дои:10.1007 / s00709-017-1147-3. ISSN 1615-6102. ЧВК 5756292. PMID 28875267.
- ^ Йи З, Сон В., Зажим Дж. К., Чен З, Гао С., Чжан Кью (декабрь 2008 г.). «Пересмотр систематических взаимоотношений внутри отряда Euplotida (Protista, Ciliophora) с использованием новых последовательностей гена, кодирующего малую субъединицу рРНК, и тестирование использования комбинированных наборов данных для построения филогении комплекса Diophrys». Мол. Филогенет. Evol. 50 (3): 599–607. Дои:10.1016 / j.ympev.2008.12.006. PMID 19121402.
- ^ Miao M, Song W, Chen Z и др. (2007). «Уникальный эуплотидный инфузорий, Gastrocirrhus (Protozoa, Ciliophora): оценка его филогенетического положения на основании последовательности гена малой субъединицы рРНК». J. Eukaryot. Микробиол. 54 (4): 371–8. Дои:10.1111 / j.1550-7408.2007.00271.x. PMID 17669163.
- ^ Альфред Каль (1930). Urtiere oder Protozoa I: Wimpertiere oder Ciliata - Volume I General Section And Prostomata.
- ^ «Медицинское определение CILIATA». www.merriam-webster.com. Получено 2017-12-22.
- ^ Adl, Sina M .; Басс, Дэвид; Lane, Christopher E .; Лукеш, Юлий; Schoch, Conrad L .; Смирнов, Алексей; Агата, Сабина; Берни, Седрик; Браун, Мэтью У .; Бурки, Фабьен; Карденас, Пако (2019). «Поправки к классификации, номенклатуре и разнообразию эукариот». Журнал эукариотической микробиологии. 66 (1): 4–119. Дои:10.1111 / jeu.12691. ISSN 1550-7408. ЧВК 6492006. PMID 30257078.
- ^ Adl, Sina M .; и другие. (2005). «Новая классификация эукариот более высокого уровня с упором на таксономию простейших». Журнал эукариотической микробиологии. 52 (5): 399–451. Дои:10.1111 / j.1550-7408.2005.00053.x. PMID 16248873.
- ^ а б Линн, Денис (2008). Ресничные простейшие: характеристика, классификация и руководство по литературе (3-е изд.). Springer. стр.58. ISBN 978-1-4020-8238-2.
1007/978-1-4020-8239-9
- ^ Х., Линн, Денис (2008). Ресничные простейшие: характеристика, классификация и руководство по литературе. Нью-Йорк: Спрингер. п. 30. ISBN 9781402082382. OCLC 272311632.
- ^ а б c Х., Линн, Денис (2008). Ресничные простейшие: характеристика, классификация и руководство по литературе. Нью-Йорк: Спрингер. п. 23. ISBN 9781402082382. OCLC 272311632.
- ^ Бергер Дж. Д. (октябрь 1986 г.). «Аутогамия в Paramecium. Приверженность стадии клеточного цикла к мейозу». Exp. Cell Res. 166 (2): 475–85. Дои:10.1016/0014-4827(86)90492-1. PMID 3743667.
- ^ а б c Райков И.Б. (1972). «Ядерные явления при конъюгации и автогамии у инфузорий». Исследования в протозоологии. 4: 149.
- ^ Финли, Гарольд Э. "Сопряжение Микростома вортичеллы. "Труды Американского общества микроскопии (1943): 97-121.
- ^ «Знакомство с ресничками». Получено 2009-01-16.
- ^ а б c Мотидзуки, Кадзуфуми (2010). «Перестройки ДНК, направленные некодирующими РНК в инфузориях». Междисциплинарные обзоры Wiley: РНК. 1 (3): 376–387. Дои:10.1002 / wrna.34. ЧВК 3746294. PMID 21956937.
- ^ Li, C.-W .; и другие. (2007). «Ресничные простейшие из докембрийской формации Доушантуо, Венган, Южный Китай». Геологическое общество, Лондон, Специальные публикации. 286 (1): 151–156. Bibcode:2007GSLSP.286..151L. Дои:10.1144 / SP286.11.
- ^ Бомфлер, Бенджамин; Керп, Ганс; Тейлор, Томас Н .; Моэструп, Эйвинд; Тейлор, Эдит Л. (18 декабря 2012 г.). «Кокон триасовой пиявки из Антарктиды содержит ископаемое животное-колокольчик». Труды Национальной академии наук Соединенных Штатов Америки. 109 (51): 20971–20974. Bibcode:2012PNAS..10920971B. Дои:10.1073 / pnas.1218879109. ISSN 1091-6490. ЧВК 3529092. PMID 23213234.
- ^ Кавальер-Смит, Т. (2000). Мегаэволюция жгутиков: основа диверсификации эукариот. В: Leadbeater, B.S.C., Green, J.C. (ред.). Жгутиковые. Единство, разнообразие и эволюция. Лондон: Тейлор и Фрэнсис, стр. 361-390, стр. 362, г. [1].
- ^ «Балантидиаз». DPDx - Лабораторная идентификация паразитарных заболеваний, вызывающих озабоченность общественного здравоохранения. Центры по контролю и профилактике заболеваний. 2013.
- ^ Рамачандран, Амбили (23 мая 2003 г.). "Введение". Паразит: Balantidium coli Болезнь: балантидиаз. ParaSite. Стэндфордский Университет.CS1 maint: ref = harv (ссылка на сайт)
- ^ Шистер, Фредерик Л. и Линн Рамирес-Авила (октябрь 2008 г.). "Текущее мировое состояние Balantidium coli". Обзоры клинической микробиологии. 21 (4): 626–638. Дои:10.1128 / CMR.00021-08. ЧВК 2570149. PMID 18854484.
дальнейшее чтение
- Линн, Денис Х. (2008). Ресничные простейшие: характеристика, классификация и руководство по литературе. Нью-Йорк: Спрингер. ISBN 9781402082382. OCLC 272311632.
- Инфузории: клетки как организмы. Хаусманн, Клаус., Брэдбери, Филлис К. (Phyllis Clarke). Штутгарт: Густав Фишер Верлаг. 1996 г. ISBN 978-3437250361. OCLC 34782787.CS1 maint: другие (ссылка на сайт)
- Иллюстрированный справочник по простейшим: организмы, традиционно называемые простейшими, или недавно обнаруженные группы.. Ли, Джон Дж., Лидейл, Гордон Ф., Брэдбери, Филлис К. (Филлис Кларк) (2-е изд.). Лоуренс, Канзас, США: Общество протозоологов. 2000 г. ISBN 9781891276224. OCLC 49191284.CS1 maint: другие (ссылка на сайт)
| Микроскопические открытия1 |
|  | ||||
|---|---|---|---|---|---|---|
| Общие темы | ||||||
| похожие темы |
| |||||
| Связанные люди |
| |||||
| Признания | ||||||
| ||||||