Мархантиофита - Marchantiophyta
| Печеночники | |
|---|---|
 | |
| "Hepaticae" из Эрнст Геккель с Kunstformen der Natur, 1904 | |
| Научная классификация | |
| Королевство: | Plantae |
| Clade: | Эмбриофиты |
| Подразделение: | Мархантиофита Stotler & Stotl.-Crand., 1977.[2] исправить. 2000 г.[3] |
| Классы и заказы | |
В Мархантиофита /мɑːrˌkæптяˈɒжɪтə/ (![]() Слушать) являются подразделением несосудистый наземные растения обычно называют гепатит или печеночники. подобно мхи и роголистник, у них есть гаметофит -доминантный жизненный цикл, при котором клетки растения несут только один набор генетической информации.
Слушать) являются подразделением несосудистый наземные растения обычно называют гепатит или печеночники. подобно мхи и роголистник, у них есть гаметофит -доминантный жизненный цикл, при котором клетки растения несут только один набор генетической информации.
По оценкам, существует около 9000 видов печеночников.[4] Некоторые из более знакомых видов растут в виде плоских безлистных слоевище, но большинство видов листовые и имеют форму, очень похожую на сплющенную. мох. Лиственные виды можно отличить от внешне похожих мхов по ряду признаков, включая их одноклеточные. ризоиды. Печеночники листовые также отличаются от большинства (но не всех) мхов тем, что на их листьях никогда не бывает Коста (присутствует во многих мхах) и может нести маргинальные реснички (очень редко во мхах). Другие различия не универсальны для всех мхов и печеночников, но наличие трехуровневых листьев, наличие глубоких лопастей или сегментированных листьев или отсутствие четко дифференцированных стеблей и листьев - все указывает на то, что растение является печеночником.
Печеночники обычно маленькие, обычно от 2 до 20 мм в ширину с отдельными растениями менее 10 см в длину, поэтому их часто упускают из виду. Однако некоторые виды могут покрывать большие участки земли, камней, деревьев или любого другого достаточно твердого субстрата, на котором они встречаются. Они распространены по всему миру почти во всех доступных средах обитания, чаще всего во влажных местах, хотя встречаются также пустынные и арктические виды. Некоторые виды могут быть помехой в тенистых теплицах или сорняком в садах.[5]
Физические характеристики
Описание
Большинство печеночников небольшие, размером от 2 до 20 миллиметров (0,08-0,8 дюйма) в ширину, с отдельными растениями менее 10 сантиметров (4 дюйма) в длину,[6] поэтому их часто упускают из виду. Наиболее известные печеночники состоят из распростертой, уплощенной, ленточной или ветвистой структуры, называемой слоевище (тело растения); эти печеночники называются печеночники таллозы. Однако у большинства печеночников стебли плоские с перекрывающимися чешуями или листья в двух или более рядах, средний ряд часто заметно отличается от внешних рядов; они называются печеночники листовые или печеночники чешуи.[7][8] (Увидеть галерея ниже для примеров.)

Печеночники наиболее надежно отличить от внешне похожих мхи их одноклеточными ризоиды.[9] Другие различия не универсальны для всех мхов и печеночников;[8] но отсутствие четко дифференцированных стеблей и листьев у видов таллозы или у листовых видов наличие глубоко лопастных или сегментированных листьев и наличие листьев, расположенных в три ряда, все указывает на то, что растение является печеночником.[10][11] В отличие от любых других эмбриофитов, большинство печеночников содержат уникальные мембраносвязанные масляные тела содержащие изопреноиды по крайней мере в некоторых из своих клеток, липидные капли в цитоплазме всех других растений остаются незамкнутыми.[12] Общее физическое сходство некоторых мхов и печеночников листовых означает, что подтверждение идентификации некоторых групп может быть выполнено с уверенностью только с помощью микроскопия или опытный бриолог .
Печеночники, как и другие мохообразные, обладают гаметофит -доминантный жизненный цикл, с спорофит зависит от гаметофита.[12] Каждая клетка типичного печеночника содержит только один набор генетической информации, поэтому клетки растения гаплоидный большую часть своего жизненного цикла. Это резко контрастирует с картиной, наблюдаемой почти у всех животных и сосудистых растений. В более знакомом семенные растения гаплоидное поколение представлено только крошечными пыльца и яйцеклетка, в то время диплоид поколение - это знакомое дерево или другое растение.[13] Другой необычной особенностью жизненного цикла печеночника является то, что спорофиты (т.е. диплоид body) очень недолговечны, отмирают вскоре после выпуска спор.[14] У мхов спорофит более стойкий, а у роголистника спорофит рассеивает споры в течение длительного времени.[нужна цитата ]
Жизненный цикл

Жизнь печеночника начинается с прорастания гаплоида. спора произвести протонема, который представляет собой либо массу нитевидных волокон, либо уплощенный слоевище.[15][16] Протонема - это переходный этап в жизни печеночника, из которого вырастет зрелый гаметофор ("гамета -bearer ") растение, производящее половые органы. Мужские органы известны как антеридия (единственное число: antheridium) и продуцируют сперматозоиды. Кластеры антеридий окружены защитным слоем клеток, называемым перигоний (множественное число: перигония). Как и у других наземных растений, женские органы известны как архегония (единственное число: archegonium) и защищены тонким окружением перихетум (множественное число: perichaeta).[8] Каждый архегоний имеет тонкую полую трубку, «шейку», по которой сперматозоиды плывут, чтобы достичь яйцеклетки.
Виды печеночника могут быть двояковыпуклый или однообразный. У двудольных печеночников женские и мужские половые органы переносятся на разных и отдельных гаметофитных растениях. У одноплодных печеночников два вида репродуктивных структур расположены на разных ветвях одного и того же растения.[17] В любом случае сперматозоиды должны перемещаться из антеридиев, где они производятся, в архегоний, где содержатся яйца. В сперма печеночников - это двустворчатый, т.е. имеют две хвостовидные жгутики позволяющие плавать на короткие дистанции,[18] при условии наличия хотя бы тонкой пленки воды. Их путешествию могут помочь брызги дождевых капель. В 2008 году японские исследователи обнаружили, что некоторые печеночники способны выпускать содержащую сперму воду на высоту до 15 см, что позволяет им оплодотворять женские растения, растущие на расстоянии более метра от ближайшего мужского пола.[19]
Когда сперма достигает архегонии, оплодотворение происходит, что приводит к образованию диплоидного спорофита. После оплодотворения неполовозрелые спорофит внутри архегония развиваются три отдельные области: (1) ступня, который закрепляет спорофит на месте и получает питательные вещества от своего «материнского» растения (2) сферической или эллипсоидной формы. капсула, внутри которого споры будут продуцироваться для распространения в новые места, и (3) щетинка (стебель), который лежит между двумя другими регионами и соединяет их.[18] Когда спорофит разовьется во всех трех областях, щетинка удлиняется, выталкиваясь из архегониума и разрывая его. В то время как ножка остается закрепленной внутри родительского растения, капсула вытесняется щетинкой и вытягивается от растения в воздух. Внутри капсулы клетки делятся, производя как Elater клетки и споровые клетки. Элатеры имеют форму пружины и толкают стенку капсулы, чтобы рассыпаться, когда капсула лопается. Клетки, продуцирующие споры, подвергаются мейоз образовывать гаплоид споры рассредоточиться, после чего жизненный цикл может начаться снова.
Бесполое размножение
Некоторые печеночники способны бесполое размножение; для мохообразных в целом «почти верно было бы сказать, что вегетативное размножение является правилом, а не исключением».[20] Например, в Riccia, когда более старые части раздвоенных слоевищ отмирают, молодые кончики становятся отдельными особями.[20]
Некоторые печеночники таллозы, такие как Marchantia polymorpha и Lunularia cruciata производить небольшие дискообразные геммы в неглубоких чашках.[21] Маршанция gemmae могут быть разнесены на глубину до 120 см при попадании дождя в чашки.[22] В Мецгерия, геммы растут по краям слоевища.[23] Marchantia polymorpha является обычным сорняком в теплицах, часто покрывающим всю поверхность контейнеров;[24]:230 Распространение геммы является «основным механизмом распространения печеночника по питомнику или теплице».[24]:231
Экология
Сегодня печеночники можно найти во многих экосистемах по всей планете, за исключением моря и чрезмерно засушливой окружающей среды или тех, которые подвергаются высоким уровням прямого солнечного излучения.[25] Как и большинство групп живых растений, они наиболее распространены (как по количеству, так и по видам) во влажных тропических районах.[26] Печеночники чаще встречаются в тени от умеренной до глубокой, хотя пустынные виды могут терпеть прямой солнечный свет и периоды полного высыхания.
Классификация
Отношение к другим растениям
Традиционно печеночники сгруппировали вместе с другими мохообразные (мхи и роголистник ) в отделе Bryophyta, в котором печеночники составляли класс Hepaticae (также называемый Marchantiopsida).[8][27] Однако, поскольку эта группировка делает Bryophyta парафилетический, печеночникам теперь обычно дают отдельное подразделение.[28] Использование названия подразделения Bryophyta Sensu lato все еще встречается в литературе, но чаще это имя используется в ограниченном смысле, чтобы включать только мхи.
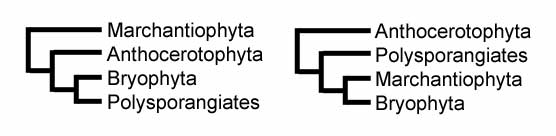
Еще одна причина того, что печеночники теперь классифицируются отдельно, заключается в том, что они, по-видимому, отошли от всех остальных. эмбриофит растения в начале своего эволюция. Самым убедительным подтверждающим доказательством является то, что печеночники - единственная живая группа наземных растений, у которых нет устьица на спорофит поколение.[29] Среди самых ранних окаменелости Считается, что печеночники окаменелости сжатия из Паллавицинииты с Верхнего Девонский из Нью-Йорк.[30] Эти окаменелости напоминают современные виды в Мецгериалес.[31] Другая девонская окаменелость называется Протосальвиния также похож на печеночник, но его отношение к другим растениям все еще неясно, поэтому он может не принадлежать к Marchantiophyta. В 2007 году были объявлены самые старые окаменелости, относящиеся к печеночникам. Metzgeriothallus sharonae от Живетян (Средний Девонский ) из Нью-Йорк, Соединенные Штаты.[32] Однако в 2010 году в Аргентине было обнаружено пять различных типов окаменелых спор печеночника, относящихся к гораздо более раннему среднему периоду. Ордовик около 470 миллионов лет назад.[1][33]
Внутренняя классификация
Бриологи классифицировать печеночники по разделу Мархантиофита. Это разделительное название основано на названии наиболее общепризнанного рода печеночников. Маршанция.[34] В добавок к этому таксон -основное название, печеночники часто называют Гепатикофита. Это название происходит от их общего латинского названия, поскольку латинский язык был языком, на котором ботаники публиковали свои описания видов. Это название привело к некоторой путанице,[нужна цитата ] отчасти потому, что это название таксона, производное от род Hepatica что на самом деле цветущее растение из семейства лютиковых Лютиковые. Кроме того, название Hepaticophyta часто ошибочно пишется в учебниках как Гепатофита, что только добавляет путаницы.
Хотя среди бриологов нет единого мнения относительно классификации печеночников выше семейного ранга,[35] Marchantiophyta можно разделить на три класса:[36][37][38][39]
- В Юнгерманиопсида включает два заказы Мецгериалес (простые таллоиды) и Jungermanniales (печеночники листовые).
- В Марчантиопсида включает три ордена Маршантиалес (сложные слоевища печеночников) и Sphaerocarpales (бутылочные гепатики), а также Blasiales (ранее относился к Metzgeriales).[36][40] Он также включает проблемный род Моноклея, который иногда помещается в отдельный заказ Monocleales.[41]
- Третий класс, Haplomitriopsida недавно признан сестринской группой других печеночников;[39] он включает в себя роды Гапломитрий, Treubia, и Apotreubia.
На диаграмме справа показана часть 2006 г. кладистический анализ печеночников на основе трех генов хлоропластов, одного ядерного гена и одного митохондриального гена.[36] |
|
Обновленная классификация Söderström et al. 2016 г.[42]
- Мархантиофита Stotler & Crandall-Stotler 2000
- Haplomitriopsida Stotler & Crandall-Stotler 1977 г.
- Haplomitriales Хэмлин 1972
- Treubiales Шляков 1972
- Марчантиопсида Кронквист, Тахтаджан и Циммерманн, 1966 г.
- Blasiidae He-Nygrén et al. 2006 г.
- Blasiales Stotler & Crandall-Stotler 2000
- Marchantiidae Engler 1893 sensu He-Nygrén et al. 2006 г.
- Neohodgsoniales Длинный 2006
- Sphaerocarpales Спелеологи 1910 (печеночник бутылочный)
- Lunulariales Длинный 2006
- Маршантиалес Limpricht 1877 г. (сложные таллоиды)
- Blasiidae He-Nygrén et al. 2006 г.
- Юнгерманиопсида Stotler & Crandall-Stotler 1977 г.
- Pelliidae He-Nygrén et al. 2006 г.
- Пеллиалес He-Nygrén et al. 2006 г.
- Паллавициниалес Фрей и Стеч 2005
- Fossombroniales Шляков 1972
- Metzgeriidae Варфоломей-Беган 1990
- Pleuroziales Шляков 1972
- Мецгериалес Шало 1930
- Jungermanniidae Энглер 1893 (печеночники листовые)
- Porellales Шляков 1972
- Ptilidiales Шляков 1972
- Jungermanniales фон Клингграфф 1858
- Pelliidae He-Nygrén et al. 2006 г.
- Haplomitriopsida Stotler & Crandall-Stotler 1977 г.
По оценкам, существует около 9000 видов печеночников, по крайней мере 85% из которых относятся к группе листовых.[3][43] Несмотря на это, на сегодняшний день геномы печеночника не секвенированы, а идентифицировано и охарактеризовано лишь несколько генов.[44]
Экономическое значение
В древности считалось, что печеночники излечивают болезни печень, отсюда и название.[45] В Древнеанглийский, слово печеночник буквально означает печеночное растение.[46] Вероятно, это произошло из-за внешнего вида некоторых таллоидных печеночников, которые по очертаниям напоминают печень, и привело к общепринятому названию группы как гепатит, от латинский слово гепатикус за «принадлежность к печени». Несвязанное цветущее растение, Hepatica, иногда также называют печеночником, потому что когда-то он также использовался при лечении заболеваний печени. Это архаичное отношение формы растения к функции было основано на «Доктрина подписей».[47]
Печеночники сегодня не имеют прямого экономического значения. Наибольшее влияние они оказывают косвенным образом, через уменьшение эрозии вдоль берегов рек, их сбор и удержание воды в тропических лесах, а также формирование почвенные корки в пустынях и полярных регионах. Однако некоторые виды используются людьми напрямую. Несколько видов, например Riccia fluitans, водные печеночники таллозы, продаваемые для использования в аквариумы. Их тонкие, тонкие ветви плавают по поверхности воды и являются средой обитания как для мелких беспозвоночных, так и для рыб, которые ими питаются.
Галерея
Небольшая подборка изображений, демонстрирующих структуру и разнообразие печеночника:

Маршанция полиморфный, с антеридиальными и архегониальными стеблями. 
В архегоний из Порелла. 
Спорофит Порелла выходит из своего архегония. 
Порелла платифилла комок, растущий на дереве. 
Пеллия эпифилла, растет на влажной почве. 
Плагиохила асплениоид, а печеночник листовой. 
Riccia fluitans, водный таллоза печеночника. 
Conocephalum коникум, крупный слоевище печеночника.








Смотрите также
использованная литература
- ^ а б Уокер, Мэтт. «Окаменелости самых ранних наземных растений, обнаруженных в Аргентине» [1]. (BBC, Earth News, 2010).
- ^ Stotler, Raymond E .; Барбара Дж. Кэндалл-Стотлер (1977). «Контрольный список печеночников и роголистников Северной Америки». Бриолог. Американское бриологическое и лихенологическое общество. 80 (3): 405–428. Дои:10.2307/3242017. JSTOR 3242017.
- ^ а б Крэндалл-Стотлер, Барбара; Стотлер, Раймонд Э. (2000). «Морфология и классификация Marchantiophyta». В А. Джонатане Шоу; Бернар Гоффине (ред.). Биология мохообразных. Кембридж: Издательство Кембриджского университета. п. 21. ISBN 0-521-66097-1.
- ^ Печеночники Домашняя страница | UNB
- ^ Шустер, Рудольф М. Hepaticae и Anthocerotae Северной Америки, том VI, стр. 19. (Чикаго: Филдский музей естественной истории, 1992). ISBN 0-914868-21-7.
- ^ Шустер, Рудольф М. Hepaticae и Anthocerotae Северной Америки, т. I. С. 243–244. (Нью-Йорк: издательство Колумбийского университета, 1966 г.)
- ^ Кашьяп, Шив Рам. Печеночники Западных Гималаев и равнины Пенджаб, т. I, стр. 1. (Нью-Дели: Ботаническая хроника, 1929 г.)
- ^ а б c d Шофилд, В. Б. Введение в бриологиюС. 135–140. (Нью-Йорк: Макмиллан, 1985). ISBN 0-02-949660-8.
- ^ Нехира, Кунито. "Прорастание спор, развитие протонем и развитие спорообразования", с. 347 в Рудольф М. Шустер (ред.), Новое руководство по бриологии, том I. (Нитинан, Миядзаки, Япония: Ботаническая лаборатория Хаттори, 1983). ISBN 49381633045.
- ^ Эллисон, К. У. и Джон Чайлд. Печеночники Новой ЗеландииС. 13–14. (Данидин: University of Otago Press, 1975).
- ^ Конард, Генри С. и Пол Л. Редферн-младший. Как узнать мхи и печеночники, перераб. ред., с. 12–23. (Дубьюк, Айова: William C. Brown Co., 1979) ISBN 0-697-04768-7
- ^ а б Гарольд К. Болд, К. Дж. Алексопулос и Т. Делеворяс. Морфология растений и грибов, 5 изд., С. 189. (Нью-Йорк: Харпер-Коллинз, 1987). ISBN 0-06-040839-1.
- ^ Фоскет, Дональд Э. Рост и развитие растений: молекулярный подход, п. 27. (Сан-Диего: Academic Press, 1994). ISBN 0-12-262430-0.
- ^ Хикс, Мари Л. Путеводитель по печеночникам Северной Каролины, п. 10. (Дарем: издательство Duke University Press, 1992). ISBN 0-8223-1175-5.
- ^ Нехира, Кунито. "Прорастание спор, развитие протонемат и развитие спорообразования", стр. 358–374 в Рудольф М. Шустер (ред.), Новое руководство по бриологии, том I. (Нитинан, Миядзаки, Япония: Ботаническая лаборатория Хаттори, 1983). ISBN 49381633045.
- ^ Чопра, Р. Н. и П. К. Кумра. Биология мохообразных, стр. 1–38. (Нью-Йорк: John Wiley & Sons, 1988). ISBN 0-470-21359-0.
- ^ Малькольм, Билл и Нэнси Малкольм. Мхи и другие мохообразные: иллюстрированный глоссарий, стр. 6 и 128. (Новая Зеландия: Micro-Optics Press, 2000). ISBN 0-473-06730-7.
- ^ а б Кэмпбелл, Дуглас Х. Строение и развитие мхов и папоротниковС. 73–74. (Лондон: Macmillan Co., 1918).
- ^ Боль, С. (2010). «Ботаническая баллистика». Новый ученый. 208 (2792/3): 45–47. Дои:10.1016 / s0262-4079 (10) 63177-6.
- ^ а б Лепп, Хейно (15 апреля 2008 г.). «Вегетативное размножение». Австралийские мохообразные. Австралийский национальный ботанический сад. Получено 22 декабря, 2011.
- ^ Смит, AJE (1989) Печеночники Британии и Ирландии, Cambridge University Press, Кембридж.
- ^ Equihua, C. (1987). "Разброс гемм в печеночнике Marchantia-Polymorpha". Cryptogamie, Bryologie, Lichénologie. 8 (3): 199–217. Архивировано из оригинал на 2012-04-26.
- ^ Лепп, Хейно (28 февраля 2008 г.). "Воспроизведение и распространение". Австралийские мохообразные. Австралийский национальный ботанический сад. Получено 22 декабря, 2011.
- ^ а б Ньюби, Адам и Джеймс Э. Альтланд, Чарльз Х. Гиллиам и Гленн Вехтье (декабрь 2006 г.). "J. Environ. Hort. 24 (4)" (PDF). Послевсходовая борьба с печеночником в питомниках, выращиваемых в контейнерах1. Научно-исследовательский институт садоводства. С. 230–236. Архивировано из оригинал (PDF) 24 июля 2012 г.. Получено 24 декабря, 2011.CS1 maint: несколько имен: список авторов (ссылка на сайт)
- ^ Шустер, Рудольф М. Hepaticae и Anthocerotae Северной Америки, т. I. С. 243–249. (Нью-Йорк: издательство Колумбийского университета, 1966).
- ^ Почс, Тамаш. «Тропические лесные мохообразные», с. 59 в А. Дж. Э. Смит (ред.) Экология мохообразных. (Лондон: Чепмен и Холл, 1982). ISBN 0-412-22340-6.
- ^ Крэндалл-Стотлер, Барбара. & Стотлер, Раймонд Э. "Морфология и классификация Marchantiophyta". стр. 36–38 в А. Джонатан Шоу и Бернард Гоффине (редакторы), Биология мохообразных. (Кембридж: Издательство Кембриджского университета: 2000). ISBN 0-521-66097-1
- ^ Гоффине, Бернар. «Происхождение и филогенетические взаимоотношения мохообразных». стр. 124–149 в А. Джонатан Шоу и Бернард Гоффине (редакторы), Биология мохообразных. (Кембридж: Издательство Кембриджского университета:! 2000). ISBN 0-521-66097-1
- ^ Кенрик, Пол и Питер Р. Крейн. Происхождение и ранняя диверсификация наземных растений: кладистическое исследование, п. 59. (Вашингтон, округ Колумбия: Smithsonian Institution Press, 1997). ISBN 1-56098-730-8.
- ^ Тейлор, Томас Н. и Эдит Л. Тейлор. Биология и эволюция ископаемых растений, п. 139. (Englewood Cliffs, NJ: Prentice Hall, 1993). ISBN 0-13-651589-4.
- ^ Остендорп, Кора. Мохообразные палеозоя и мезозояС. 70–71. (Bryophytum Bibliotheca, Band 34, 1987). ISBN 3-443-62006-X.
- ^ VanAller Hernick, L .; Посадка, Е .; Бартовски, К. (2008). «Древнейшие печеночники Земли - Metzgeriothallus sharonae sp. ноя из среднего девона (живетского) восточного Нью-Йорка, США ". Обзор палеоботаники и палинологии. 148 (2–4): 154–162. Дои:10.1016 / j.revpalbo.2007.09.002.
- ^ Rubinstein, C.V .; Gerrienne, P .; De La Puente, G.S .; Astini, R.A .; Стиманс, П. (2010). «Свидетельства раннего среднего ордовика для наземных растений в Аргентине (восточная Гондвана)». Новый Фитолог. 188 (2): 365–369. Дои:10.1111 / j.1469-8137.2010.03433.x. PMID 20731783.
- ^ Крэндалл-Стотлер, Барбара. & Стотлер, Раймонд Э. "Морфология и классификация Marchantiophyta". п. 63 в А. Джонатан Шоу и Бернард Гоффине (редакторы), Биология мохообразных. (Кембридж: Издательство Кембриджского университета: 2000). ISBN 0-521-66097-1
- ^ Джонс, Э. У. (2004). Флора печеночника и роголистника Западной Африки. Scripta Botnica Belgica. 30. Мейсе: Национальный ботанический сад (Бельгия). п. 30. ISBN 90-72619-61-7.
- ^ а б c Форрест, Лаура Л .; Кристин Э. Дэвис; Дэвид Г. Лонг; Барбара Дж. Крэндалл-Стотлер; Александра Кларк; Мишель Л. Холлингсворт (2006). «Раскрытие истории эволюции печеночников (Marchantiophyta): многочисленные таксоны, геномы и анализы». Бриолог. 109 (3): 303–334. Дои:10.1639 / 0007-2745 (2006) 109 [303: UTEHOT] 2.0.CO; 2.
- ^ Генрихс, Йохен; С. Робберт Градштейн; Розмари Уилсон; Харальд Шнайдер (2005). «К естественной классификации печеночников (Marchantiophyta) на основе гена хлоропластов rbcL». Cryptogamie Bryologie. 26 (2): 131–150.
- ^ Хэ-Нюгрен, Сяолань; Айно Юслен; Инкери Ахонен; Дэвид Гленни; Синикка Пийппо (2006). «Освещение эволюционной истории печеночников (Marchantiophyta) - к естественной классификации». Кладистика. 22 (1): 1–31. Дои:10.1111 / j.1096-0031.2006.00089.x. S2CID 86082381.
- ^ а б Renzaglia, Karen S .; Скотт Шютт; Р. Джоэл Дафф; Роберто Лигроне; А. Джонатан Шоу; Брент Д. Мишлер; Джеффри Дж. Дакетт (2007). «Филогения мохообразных: расширение молекулярных и морфологических границ». Бриолог. 110 (2): 179–213. Дои:10.1639 / 0007-2745 (2007) 110 [179: BPATMA] 2.0.CO; 2.
- ^ Форрест, Лаура Л .; Барбара Дж. Крэндалл-Стотлер (2004). «Филогения простых таллоидных печеночников (Jungermanniopsida, Metzgeriidae) на основе пяти генов хлоропластов». Монографии по систематической ботанике. Молекулярная систематика мохообразных. Пресса Ботанического сада Миссури. 98: 119–140.
- ^ Шустер, Рудольф М. Hepaticae и Anthocerotae Северной Америки, т. VI, стр. 26. (Чикаго: Филдский музей естественной истории, 1992). ISBN 0-914868-21-7.
- ^ Седерстрём; и другие. (2016). «Всемирный контрольный список роголистников и печеночников». Фитоключи. 59 (59): 1–826. Дои:10.3897 / фитоключи.59.6261. ЧВК 4758082. PMID 26929706.
- ^ Садава, Давид; Дэвид М. Хиллис; Х. Крейг Хеллер; Мэй Беренбаум (2009). Жизнь: Наука биологии (9-е изд.). Нью-Йорк: У. Х. Фриман. п. 599. ISBN 978-1429246446.
- ^ Sierocka, I; Козловский, Л. П .; Bujnicki, J.M .; Ярмоловски, А; Szweykowska-Kulinska, Z (2014). «Специфическая для самок экспрессия гена в печеночнике двудомной Pellia endiviifolia регулируется в процессе развития и связана с производством архегоний». BMC Биология растений. 14: 168. Дои:10.1186/1471-2229-14-168. ЧВК 4074843. PMID 24939387.
- ^ Диттмер, Говард Дж. Филогения и форма в царстве растений, п. 286. (Торонто: Д. Ван Ностранд Ко., 1964)
- ^ Рэйвен П. Х., Р. Ф. Эверт и С. Э. Эйххорн. Биология растений, 7 изд., С. 351. (Нью-Йорк: В. Х. Фриман, 2005). ISBN 0-7167-1007-2.
- ^ Стерн, Кингсли Р. Вводная биология растений, 5 изд., С. 338. (Дубьюк, Айова: издательство Wm. C. Brown Publishers, 1991). ISBN 0-697-09947-4.
внешние ссылки
- Строение печеночника в картинках
- LiToL: Сборка Древа Жизни Печеночника (примечание: 500 000 миллионов лет назад читать "480 миллионов лет назад".)
- Взаимоотношения мхов, печеночников и роголистников
- Дополнительная информация о печеночниках
- Печеночники