Моллюска - Mollusca
| Моллюска | |
|---|---|
 | |
| Тоникелла линейная, а полиплакофоран или хитон, передний (головной) конец вправо | |
| Научная классификация | |
| Королевство: | Animalia |
| Подцарство: | Eumetazoa |
| Clade: | ParaHoxozoa |
| Clade: | Bilateria |
| Clade: | Нефрозоа |
| (без рейтинга): | Протостомия |
| (без рейтинга): | Спиралия |
| Суперфилум: | Лофотрохозоа |
| Тип: | Моллюска Линней, 1758 |
| Классы | |
| Разнообразие[1] | |
| 85000 признанных живых существ. | |

Моллюска второй по величине филюм из беспозвоночный животные после Членистоногие. Члены известны как моллюски или моллюски[а] (/ˈмɒлəsk/). Около 85 000сохранившийся виды моллюсков.[3] Число ископаемых видов оценивается от 60 000 до 100 000 дополнительных видов.[4] Доля неописанных видов очень высока. Многие таксоны остаются малоизученными.[5]
Моллюски - самые крупные морской тип, составляющий около 23% всех названных морских организмы. Многочисленные моллюски также обитают в пресная вода и земной среда обитания. Они очень разнообразны, не только по размеру и анатомический структура, но также в поведении и среде обитания. Тип обычно делится на 8 или 9таксономический классы, из которых два полностью вымерший. Головоногие моллюски моллюски, такие как Кальмар, каракатица, и осьминоги, являются одними из самых неврологически продвинутый из всех беспозвоночные - и либо Гигантский кальмар или колоссальный кальмар - самый крупный из известных видов беспозвоночных. В брюхоногие моллюски (улитки и слизни ) являются самыми многочисленными моллюсками и составляют 80% от общего числа классифицированных видов.
Тремя наиболее универсальными чертами, определяющими современных моллюсков, являются: мантия со значительной полостью, используемой для дыхания и выделение, наличие радула (кроме двустворчатые моллюски ), а структура нервная система. Помимо этих общих элементов, моллюски обладают большим морфологическим разнообразием, поэтому многие учебники основывают свои описания на «гипотетическом предковом моллюске» (см. Изображение ниже). Это сингл "бездомный -любить" оболочка сверху, который сделан из белки и хитин усиленный карбонат кальция, и секретируется мантией, покрывающей всю верхнюю поверхность. Нижняя часть животного состоит из единой мускульной «ступни». Хотя моллюски целоматы, то целомудрие имеет тенденцию быть маленьким. Основная полость тела представляет собой гемоцель через который кровь циркулирует; как таковые, их системы кровообращения в основном открыто. Система питания «генерализованного» моллюска состоит из грубого «языка», радулы и сложной пищеварительной системы, в которой выделяется слизь и микроскопические "волоски" с мышцами, называемые реснички играть различные важные роли. У генерализованного моллюска два парных нервные шнуры, или три в двустворчатые моллюски. В мозг, у видов, которые имеют один, обводит пищевод. Большинство моллюсков имеют глаза, и все они оснащены датчиками для обнаружения химикатов, вибрации и прикосновений. Самый простой вид моллюсков репродуктивная система полагается на внешнее оплодотворение, но встречаются и более сложные варианты. Почти все производят яйца, из которых может возникнуть трохофор личинки, более сложный велигер личинки или миниатюрные взрослые особи. Целомическая полость уменьшена. У них открытая система кровообращения и органы выведения, похожие на почки.
Существуют веские доказательства появления брюхоногих моллюсков, головоногие моллюски, и двустворчатые моллюски в Кембрийский период, 541–485,4 миллиона лет назад. Однако эволюционная история появления обоих моллюсков из предковых Лофотрохозоа и об их разнообразии в хорошо известные жизни и ископаемое формы до сих пор являются предметом жарких споров среди ученых.

Моллюски были и остаются важным источником пищи для анатомически современные люди. Однако существует риск пищевого отравления из-за токсинов, которые могут накапливаться в определенных моллюсках при определенных условиях, и по этой причине во многих странах действуют правила, снижающие этот риск. Моллюски на протяжении веков также были источником важных предметов роскоши, в частности жемчуг, перламутр, Тирийский фиолетовый краситель и морской шелк. Их снаряды тоже были используемый так как Деньги в некоторых доиндустриальных обществах.
Виды моллюсков также могут представлять опасность или вредителей для деятельности человека. Укус синекольчатый осьминог часто приводит к летальному исходу, а Осьминог аполлион причины воспаление это может длиться больше месяца. Укусы некоторых видов крупных тропических конические оболочки также могут убивать, но их сложные, хотя и легко производимые, яды стали важными инструментами в неврологический исследование. Шистосомоз (также известный как бильгарциоз, бильгарциоз или лихорадка улиток) передается людям от водных улиток-хозяев и поражает около 200 миллионов человек. Улитки и слизни также могут быть серьезными сельскохозяйственными вредителями, а случайная или преднамеренная интродукция некоторых видов улиток в новые среды серьезно повредила некоторым из них. экосистемы.
Этимология
Слова моллюск и моллюск произошли от французского моллюск, который возник из латинский моллюск, от моллис, мягкий. Моллюск сам был адаптацией Аристотель с τὰ μαλάκια та малакия (мягкие; < μαλακός Malakós «мягкий»), который он применил среди прочего к каракатица.[6][7] Соответственно, научное изучение моллюсков называется малакология.[8]
Название Molluscoida ранее использовалось для обозначения подразделения животного мира, содержащего брахиоподы, мшанки, и оболочки Предполагалось, что члены трех групп чем-то напоминают моллюсков. Как теперь известно, эти группы не имеют отношения к моллюскам и очень мало связаны друг с другом, поэтому от названия Molluscoida отказались.[9]
Определение
Наиболее универсальными особенностями строения тела моллюсков являются мантия со значительной полостью, используемой для дыхание и выделение, и организация нервной системы. У многих есть известковый оболочка.[10]
Моллюски развили такое разнообразное строение тела, находя синапоморфии (определяющие характеристики) применить ко всем современным группам сложно.[11] Наиболее общая характеристика моллюсков - они не сегментированы и симметричны с двух сторон.[12] Во всех современных моллюсках присутствуют:[13][15]
- В спинной часть стенки тела является мантия (или паллий), который секреты известковый спикулы, тарелки или ракушки. Он перекрывает корпус, оставляя достаточно свободного места, чтобы образовать полость мантии.
- В анус и гениталии открываются в полость мантии.
- Есть две пары основных нервные шнуры.[14]
Другие характеристики, которые обычно встречаются в учебниках, имеют значительные исключения:
| Найдены ли характеристики в этих классы моллюсков | |||||||
| Предполагаемая универсальная характеристика моллюсков[13] | Аплакофора[14](стр. 291–292) | Полиплакофора[14](p292–298) | Моноплакофора[14](p298–300) | Брюхоногие[14](стр300–343) | Головоногие моллюски[14](p343–367) | Двустворчатые моллюски[14](p367–403) | Scaphopoda[14](p403–407) |
|---|---|---|---|---|---|---|---|
| Радула, грубый "язык" с хитиновый зубы | Отсутствует в 20% Неомениоморфа | да | да | да | да | Нет | Внутренний, не может выходить за пределы тела |
| Широкая мускулистая стопа | Снижено или отсутствует | да | да | да | Модифицирован в оружие | да | Маленький, только "перед" |
| Дорсальная концентрация внутренних органов (висцеральная масса) | Не очевидно | да | да | да | да | да | да |
| Большой пищеварительный тракт ceca | Нет слепых стволов у некоторых аплакофор | да | да | да | да | да | Нет |
| Большой комплекс метанефридия («почки») | Никто | да | да | да | да | да | Маленький, простой |
| Один или несколько клапанов / корпусов | Первобытные формы, да; современные формы, нет | да | да | Улитки, да; слизни, в основном да (внутренние рудименты) | Осьминоги, нет; каракатица, наутилус, кальмар, да | да | да |
| Одонтофор | да | да | да | да | да | Нет | да |
Разнообразие


Оценки общепринятых описанных живых видов моллюсков варьируются от 50 000 до максимум 120 000 видов.[1] В 1969 году Дэвид Никол оценил вероятное общее количество живых видов моллюсков в 107 000, из которых около 12 000.пресноводные брюхоногие моллюски и 35 000земной. Bivalvia будет составлять около 14% от общего числа, а другие пять классов - менее 2% от всех живых моллюсков.[17] В 2009 году Чепмен оценил количество описанных живых видов моллюсков в 85 000.[1] Haszprunar в 2001 году оценил около 93 000 названных видов,[18] которые включают 23% всех названных морских организмов.[19] Моллюски уступают только членистоногие по количеству живых видов животных[16] - намного отстает от 1113000 членистоногих, но намного опережает хордовые ' 52,000.[14](pПередний форзац) Всего насчитывается около 200000 живых видов,[1][20] и 70000 ископаемых видов,[13] хотя общее количество видов моллюсков, которые когда-либо существовали, независимо от того, сохранились они или нет, должно быть во много раз больше, чем их количество сегодня.[21]
Моллюски имеют более разнообразную форму, чем любое другое животное. филюм. Они включают улитки, слизни и другие брюхоногие моллюски; моллюски и другие двустворчатые моллюски; кальмары и другие головоногие моллюски; и другие менее известные, но столь же отличительные подгруппы. Большинство видов до сих пор обитают в океанах, от побережья до абиссальная зона, но некоторые образуют значительная часть пресноводной фауны и земной экосистемы. Моллюски чрезвычайно разнообразны по тропический и умеренный регионов, но можно найти вообще широты.[11] Около 80% всех известных видов моллюсков - брюхоногие.[16] Головоногие моллюски такие как Кальмар, каракатица, и осьминоги находятся среди неврологически самый продвинутый из всех беспозвоночных.[22] В Гигантский кальмар, которые до недавнего времени не наблюдались живыми во взрослой форме,[23] один из крупнейших беспозвоночные, но недавно пойманный экземпляр колоссальный кальмар, Длиной 10 м (33 фута) и весом 500 кг (1100 фунтов), возможно, обогнал его.[24]
Пресная вода и наземные моллюски кажутся исключительно уязвимыми для исчезновения. Оценки численности неморских моллюсков сильно различаются, отчасти потому, что многие регионы не были тщательно обследованы. Также не хватает специалистов, которые могут идентифицировать всех животных в любом районе по видам. Однако в 2004 г. Красный список МСОП из находящихся под угрозой исчезновения видов включали около 2000 находящихся под угрозой исчезновения неморских моллюсков. Для сравнения: подавляющее большинство видов моллюсков - морские, но только 41 из них внесен в Красный список 2004 года. Около 42% зарегистрированных исчезновений с 1500 года приходится на моллюсков, почти полностью состоящих из неморских видов.[25]
Гипотетический предковый моллюск
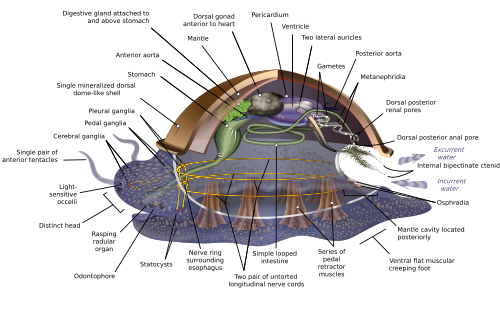
Из-за большого диапазона анатомического разнообразия моллюсков во многих учебниках анатомия моллюсков начинается с описания того, что называется анатомией. архимоллюск, гипотетический генерализованный моллюск, или гипотетический предковый моллюск (ВЕТЧИНА), чтобы проиллюстрировать наиболее общие черты, обнаруженные в пределах филума. Изображение визуально очень похоже на современное. моноплакофораны.[11][15][26]
Генерализованный моллюск билатерально симметричен и имеет один "бездомный -любить" оболочка наверху. Раковина секретируется мантией, покрывающей верхнюю поверхность. Нижняя часть состоит из единой мышечной «ступни».[15] Висцеральная масса или висцерополлий - это мягкая немышечная метаболическая область моллюска. Он содержит органы тела.[12]
Мантия и мантийная полость
Полость мантии, складка в мантии, охватывает значительное пространство. Он выстлан эпидермисом и обнажен, согласно место обитания, в море, пресную воду или воздух. У самых ранних моллюсков полость находилась сзади, но теперь ее положение варьируется от группы к группе. В анус, пара осфрадия (химические датчики) во входящей «полосе», задняя пара жабры и выходных отверстий нефридия («почки») и гонады (репродуктивные органы) находятся в полости мантии.[15] Все мягкое тело двустворчатых моллюсков находится в увеличенной мантийной полости.[12]
Оболочка
Край мантии выделяет раковину (вторично отсутствующую в ряде таксономических групп, таких как голожаберники[12]) который состоит в основном из хитин и конхиолин (а белок закаленный с карбонат кальция ),[15][27] кроме самого внешнего слоя, который почти во всех случаях состоит из конхиолина (см. периостракум ).[15] Моллюски никогда не используют фосфат для создания своих твердых частей,[28] за сомнительным исключением Cobcrephora.[29]В то время как большинство раковин моллюсков состоит в основном из арагонит, те брюхоногие моллюски, которые откладывают яйца с твердой скорлупой, используют кальцит (иногда со следами арагонита) для изготовления яичной скорлупы.[30]
Оболочка состоит из трех слоев: внешнего слоя ( периостракум ) из органического вещества, средний слой из столбчатых кальцит, и внутренний слой, состоящий из слоистого кальцита, часто перламутровый.[12]
В некоторых формах оболочка содержит отверстия. В морские ушки в скорлупе есть отверстия, используемые для дыхания и выхода яйцеклеток и спермы, в наутилус нитка ткани, называемая сифункул проходит через все камеры, и восемь пластин, составляющих оболочку хитоны пронизаны живыми тканями с нервами и сенсорными структурами.[31]
Ступня

Нижняя часть состоит из мускулистой стопы, которая адаптирована для разных целей в разных классах.[32] Нога несет пару статоцисты, которые действуют как датчики баланса. У брюхоногих он выделяет слизь в качестве смазки для движения. В формах, имеющих только верхнюю оболочку, например блюдца нога действует как присоска, прикрепляя животное к твердой поверхности, а вертикальные мышцы зажимают панцирь над ней; у других моллюсков вертикальные мышцы втягивают ступню и другие открытые мягкие части в раковину.[15] У двустворчатых моллюсков ступня приспособлена для закапывания в осадок;[32] у головоногих он используется для реактивного движения,[32] а щупальца и руки происходят от ступни.[33]
Сердечно-сосудистая система
Большинство моллюсков системы кровообращения в основном открыто. Хотя моллюски целоматы, их coeloms сводятся к довольно небольшим пространствам, окружающим сердце и гонады. Основная полость корпуса - это гемоцель через который кровь и целомическая жидкость циркулируют и охватывают большинство других внутренних органов. Эти гемоцелевые пространства действуют как эффективные гидростатический каркас.[12] В крови этих моллюсков содержится респираторный пигмент гемоцианин как кислород -перевозчик. Сердце состоит из одной или нескольких пар предсердий (ушные раковины ), которые получают насыщенную кислородом кровь от жабр и перекачивают ее в желудочек, который закачивает его в аорта (основной артерия ), который довольно короткий и открывается в гемоцель.[15] Предсердия сердца также функционируют как часть выделительная система путем фильтрации продуктов жизнедеятельности из крови и сброса их в целом в виде моча. Пара нефридия («маленькие почки») сзади и соединенные с целомом, извлекают из мочи любые повторно используемые материалы и сбрасывают в нее дополнительные продукты жизнедеятельности, а затем выбрасывают ее через трубки, которые выходят в полость мантии.[15]
Исключением из перечисленного являются моллюски. Planorbidae или улитки из бараньего рога, которые дышат воздухом и используют железо гемоглобин вместо гемоцианина на основе меди, чтобы переносить кислород через кровь.
Дыхание
У большинства моллюсков есть только одна пара жабр или даже одна жабра. Как правило, жабры по форме напоминают перья, хотя у некоторых видов жабры с нитями только с одной стороны. Они разделяют полость мантии, так что вода входит около дна и выходит около вершины. В их нитях есть три типа ресничек, один из которых направляет поток воды через полость мантии, а два других помогают поддерживать чистоту жабр. Если осфрадии обнаруживают вредные химические вещества или, возможно, осадок Попадая в полость мантии, реснички жабр могут перестать биться, пока не прекратятся нежелательные вторжения. Каждая жабра имеет входящий кровеносный сосуд, соединенный с гемоэлем, а выходящий - с сердцем.[15]
Прием пищи, пищеварение и выделение

Члены семейства моллюсков используют внутриклеточное пищеварение функционировать. У большинства моллюсков мускулистый рот с радулы, «языки», несущие множество рядов хитиновых зубов, которые заменяются сзади по мере износа. Радула в первую очередь предназначена для очистки бактерии и водоросли от камней, и связан с одонтофор, хрящевой опорный орган.[12] Радула уникальна для моллюсков и не имеет аналогов у других животных.
Во рту моллюсков также есть железы которые выделяют слизь слизь, к которому прилипает еда. Избиение реснички (крошечные «волоски») направляют слизь к желудку, поэтому слизь образует длинную нить, называемую «пищевой нитью».[15]
На сужающемся заднем конце живота и слегка выступающем в заднюю кишку находится простиль, конус, направленный назад. кал и слизь, которая вращается дополнительными ресничками, так что она действует как катушка, наматывая на себя нитку слизи. Прежде чем полоска слизи достигает простилы, кислотность желудка делает слизь менее липкой и освобождает от нее частицы.[15]
Частицы сортируются по еще одной группе ресничек, которые отправляют более мелкие частицы, в основном минералы, в простилу, так что в конечном итоге они выводятся из организма, в то время как более крупные, в основном пищевые продукты, отправляются в желудок. слепая кишка (мешок, у которого нет другого выхода) для переваривания. Процесс сортировки ни в коем случае не идеален.[15]
Периодически круговые мышцы у входа в заднюю кишку отщипывают и выделяют часть простиля, не давая простилу стать слишком большим. Анус, в части мантийной полости, охвачен выходящей «полосой» тока, создаваемой жабрами. У хищных моллюсков обычно более простая пищеварительная система.[15]
Поскольку у двустворчатых моллюсков голова в значительной степени исчезла, рот был оборудован губными щупиками (по два с каждой стороны рта) для сбора детрит из его слизи.[12]
Нервная система

У головных моллюсков две пары основных нервные шнуры организованы вокруг ряда парных ганглиев, висцеральный шнуры, обслуживающие внутренние органы, и педальные, обслуживающие стопу. Большинство пар соответствующих ганглиев с обеих сторон тела связаны между собой комиссур (относительно большие пучки нервов). Ганглии над кишечником - это церебральный, плевральный и висцеральный нервы, которые расположены над кишечником. пищевод (глотка). Педальные ганглии, контролирующие стопу, находятся ниже пищевода, а их комиссура и соединительные элементы с церебральными и плевральными ганглиями окружают пищевод в виде спайки. кольцо околопищеводного нерва или нервный воротник.[15]
У ацефальных моллюсков (то есть двустворчатых моллюсков) также есть это кольцо, но оно менее очевидно и менее важно. У двустворчатых моллюсков есть только три пары ганглиев - церебральные, педальные и висцеральные, причем висцеральный - самый большой и самый важный из трех, функционирует как главный центр «мышления». Некоторые, такие как гребешки У них есть глаза по краям раковин, которые соединяются с парой петлевых нервов и позволяют различать свет и тень.
Размножение

Самая простая репродуктивная система моллюсков основана на внешнее оплодотворение, но с более сложными вариациями. Все производят яйца, из которых могут появиться трохофор личинки, более сложные велигер личинки или миниатюрные взрослые особи. Два гонады сядьте рядом с целомудрие, небольшая полость, окружающая сердце, в которую они проливают яйцеклетка или сперма. Нефридии извлекают гаметы из целома и выбрасывают их в полость мантии. Моллюски, использующие такую систему, всю жизнь остаются одного пола и полагаются на внешнее оплодотворение. Некоторые моллюски используют внутреннее оплодотворение и / или гермафродиты, функционирующие как представители обоих полов; оба эти метода требуют более сложных репродуктивных систем.[15]
Самый простой моллюск личинка это трохофор, который планктонный и питается плавающими частицами пищи, используя две полосы ресничек вокруг своего «экватора», чтобы переместить пищу в рот, при этом используется больше ресничек, чтобы загнать их в желудок, который использует дополнительные реснички для удаления непереваренных остатков через анус. Новая ткань разрастается в полосах мезодерма во внутренней части, поэтому апикальный пучок и анус раздвигаются дальше по мере роста животного. Стадия трохофоры часто сменяется велигер этап, на котором прототрох, «экваториальная» полоса ресничек, ближайшая к апикальному пучку, развивается в велум («вуаль»), пару лопастей, несущих реснички, с которыми личинка плавает. В конце концов личинка опускается на морское дно и метаморфозы во взрослую форму. В то время как метаморфоз - обычное состояние для моллюсков, головоногие моллюски различаются тем, что демонстрируют прямое развитие: вылупившийся детеныш - это «миниатюрная» форма взрослой особи.[35]
Экология
Кормление
Большинство моллюсков травоядны, питаются водорослями или фильтраторами. Для тех, кто пасется, преобладают две стратегии кормления. Некоторые питаются микроскопическими нитчатыми водорослями, часто используя свою радулу в качестве «грабли» для вычесывания нитей со дна моря. Другие питаются макроскопическими «растениями», такими как водоросли, расчесывая поверхность растения своей радулой. Чтобы использовать эту стратегию, растение должно быть достаточно большим, чтобы моллюск мог «сидеть», поэтому более мелкие макроскопические растения не так часто едят, как их более крупные аналоги.[36]Питатели фильтра это моллюски, которые питаются, отфильтровывая взвешенные вещества и частицы пищи из воды, обычно пропуская воду через свои жабры. Большинство двустворчатых моллюсков являются фильтраторами.
Головоногие моллюски в первую очередь хищны, а радула играет второстепенную роль по сравнению с челюстями и щупальцами в добывании пищи. Моноплакофоран Неопилина использует радулу обычным образом, но в ее рацион входят простейшие, такие как ксенофиофор Stannophyllum.[37] Сакоглоссан морские слизни высасывают сок из водорослей, используя свою однорядную радулу, чтобы пробить клеточные стенки,[38] в то время как Дорид голожаберники и немного Vetigastropoda питаться губками[39][40] и другие питаются гидроидами.[41] (Обширный список моллюсков с необычными привычками питания доступен в приложении к ГРАХЭМ, А. (1955). «Моллюсковые диеты». Журнал исследований моллюсков. 31 (3–4): 144..)
Классификация
Мнения о количестве классы моллюсков; например, в таблице ниже показаны семь классов проживания,[18] и два вымерших. Хотя они вряд ли образуют кладу, некоторые более старые работы сочетают Каудофовеата и Соленогастеры в один класс, Аплакофора.[26][14](стр. 291–292) Два общепризнанных «класса» известны только по окаменелостям.[16]
| Класс | Основные организмы | Описанные живые виды[18] | Распределение |
|---|---|---|---|
| Брюхоногие [14](p300) | все улитки и слизни в том числе морское ушко, блюдца, раковина, голожаберники, морские зайцы, морские бабочки | 70,000 | морской, пресноводный, наземный |
| Двустворчатые моллюски [14](p367) | моллюски, устрицы, гребешки, геоутки, моллюски, рудисты † | 20,000 | морской, пресноводный |
| Полиплакофора [14](стр. 292–298) | хитоны | 1,000 | каменистая приливная зона и морское дно |
| Головоногие моллюски [14](p343) | Кальмар, осьминоги, каракатица, наутилусы, Спирула, белемниты †, аммониты † | 900 | морской |
| Scaphopoda [14](pp403–407) | клыки | 500 | морской 6–7 000 метров (20–22 966 футов) |
| Аплакофора [14](стр. 291–292) | червеобразные моллюски | 320 | морское дно 200–3000 метров (660–9,840 футов) |
| Моноплакофора [14](стр. 298–300) | древняя родословная моллюсков с шляпообразными раковинами | 31 | морское дно 1800–7000 метров (5 900–23 000 футов); один вид 200 метров (660 футов) |
| Ростроконхия †[42] | окаменелости; вероятные предки двустворчатых моллюсков | вымерший | морской |
| Helcionelloida †[43] | окаменелости; моллюски, похожие на улиток, такие как Latouchella | вымерший | морской |
Отнесение этих групп к более высоким таксонам было и остается проблематичным. Филогенетическое исследование предполагает, что полиплакофора образует кладу с монофилетической аплакофорой.[44] Кроме того, это предполагает наличие родственных таксонов между Bivalvia и Gastropoda. Тентакулита также может быть в Mollusca (см. Тентакулиты ).
Эволюция

Окаменелости
Существуют веские доказательства появления брюхоногие моллюски (например., Алданелла ), головоногие моллюски (например., Plectronoceras, ?Нектокарис ) и двустворчатые моллюски (Pojetaia, Fordilla ) ближе к середине Кембрийский период, c. 500 миллион лет назад, хотя, возможно, каждый из них может принадлежать только к основной линии своего соответствующего класса.[45] Однако эволюционная история появления моллюсков из предковой группы Лофотрохозоа, и об их разнообразии в хорошо известных живых и ископаемое форм, все еще активно обсуждается.
Споры ведутся о том, Эдиакарский и рано Кембрийский ископаемые действительно моллюски. Кимберелла, примерно с 555 миллион лет назад, был описан некоторыми палеонтологами как «моллюскоподобный»,[46][47] но другие не хотят идти дальше "вероятного двухсторонний ",[48][49] если это.[50]
Еще более остро ведутся споры о том, Wiwaxia, примерно с 505 миллион лет назад, был моллюском, и многое из этого связано с тем, был ли его питающий аппарат типом радула или более похож на некоторые полихета черви.[48][51] Николас Баттерфилд, который выступает против идеи, что Wiwaxia был моллюском, писал, что раньше микрофоссилий от 515–510 миллион лет назад являются фрагментами действительно похожей на моллюска радулы.[52] Это, по-видимому, противоречит концепции, согласно которой предковая радула моллюсков была минерализована.[53]
 |  |
Однако Гельционеллиды, которые сначала появляются над 540 миллион лет назад в породах раннего кембрия из Сибирь и Китай,[54][55] считаются ранними моллюсками с раковинами, напоминающими улитки. Таким образом, моллюски в панцире предшествуют самым ранним трилобиты.[43] Хотя большинство окаменелостей гелионеллид имеют длину всего несколько миллиметров, были также обнаружены образцы длиной в несколько сантиметров, большинство из которых имеют более бездомный -подобные формы. Предполагается, что крошечные экземпляры - это молодые особи, а более крупные - взрослые особи.[56]
Некоторые анализы гельционеллид пришли к выводу, что это были самые ранние брюхоногие моллюски.[57] Однако другие ученые не уверены, что эти раннекембрийские окаменелости показывают явные признаки кручение , который идентифицирует современных брюхоногих моллюсков, скручивает внутренние органы так, что анус находится над головой.[14](стр300–343)[58][59]

Volborthella, некоторые окаменелости предшествуют 530 миллион лет назаддолгое время считалось головоногим, но открытие более подробных окаменелостей показало, что его панцирь не был секретным, а построен из зерен минерала. диоксид кремния (кремнезем), и он не был разделен на серию отсеков септа как у ископаемых головоногих моллюсков и живых Наутилус находятся. Volborthella'классификация сомнительна.[60] Поздняя кембрийская окаменелость Plectronoceras в настоящее время считается самым ранним окаменелым останком головоногих моллюсков, поскольку его раковина имела перегородки и сифункул, прядь ткани, которая Наутилус используется для удаления воды из отсеков, которые он освободил по мере роста, и что также видно в окаменелостях аммонит снаряды. Однако, Plectronoceras и другие ранние головоногие моллюски ползали по морскому дну вместо того, чтобы плавать, так как их раковины содержали «балласт» каменных отложений на том, что считается нижней стороной, и имели полосы и пятна на том, что считается верхней поверхностью.[61] Все головоногие моллюски с внешней оболочкой, кроме наутилоиды вымерли к концу Меловой период 65 миллион лет назад.[62] Однако безоболочечный Coleoidea (Кальмар, осьминог, каракатица ) сегодня в изобилии.[63]
Окаменелости раннего кембрия Fordilla и Pojetaia рассматриваются как двустворчатые моллюски.[64][65][66][67] «Современные» двустворчатые моллюски появились в Ордовик период 488 по 443 миллион лет назад.[68] Одна группа двустворчатых моллюсков, рудисты, стал основным риф - строители в меловом периоде, но вымерли в Меловое – палеогеновое вымирание.[69] Несмотря на это, двустворчатые моллюски остаются многочисленными и разнообразными.
В Хиолита представляют собой класс вымерших животных с панцирем и крышка это могут быть моллюски. Авторы, которые считают, что заслуживают своего филюм не комментируйте положение этого типа на древе жизни.[70]
Филогения
| Лофотрохозоа |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
В филогения (эволюционное «генеалогическое древо») моллюсков - дискуссионная тема. Помимо дебатов о том, можно ли Кимберелла и любой из "халваксииды "были моллюсками или были близкими родственниками моллюскам,[47][48][51][52] Возникают дискуссии об отношениях между классами живых моллюсков.[49] Фактически, некоторые группы, традиционно классифицируемые как моллюски, возможно, придется переопределить как отдельные, но связанные между собой.[73]
Моллюски обычно считаются членами Лофотрохозоа,[71] группа, определяемая наличием трохофор личинки, а в случае живых Лофофората, питающая структура, называемая лофофор. Другие представители Lophotrochozoa - это кольчатые червя черви и семь морских тип.[74] На диаграмме справа представлена филогения, представленная в 2007 г. без кольчатых червей.
Поскольку отношения между членами генеалогического древа сомнительны, трудно идентифицировать признаки, унаследованные от последнего общего предка всех моллюсков.[75] Например, неясно, был ли предковый моллюск метамерный (состоящий из повторяющихся единиц) - если это так, то можно предположить происхождение от кольчатые червя -подобный червь.[76] Ученые расходятся во мнениях по этому поводу: Гирибет и его коллеги пришли к выводу, что в 2006 году повторение жабр и ретракционных мышц стопы было более поздним развитием.[11] в то время как в 2007 году Зигварт пришел к выводу, что предковый моллюск был метамерным, у него была ступня, используемая для ползания, и «раковина», которая была минерализована.[49] В одной конкретной ветви генеалогического древа оболочка ракообразные считается, что произошел от спикулы (маленькие колючки) из аплакофораны; но это трудно примирить с эмбриологический происхождение спикул.[75]
Раковина моллюска, по-видимому, образовалась из слизи, которая в конечном итоге превратилась в кутикула. Это было бы непроницаемым и, следовательно, привело бы к развитию более сложного дыхательного аппарата в виде жабр.[43] Со временем кутикула минерализовалась,[43] используя тот же генетический механизм (закрепленный ) как и большинство других билатерианских скелеты.[76] Первая раковина моллюска почти наверняка была усилена минералом. арагонит.[27]
Эволюционные отношения внутри моллюсков также обсуждаются, и на приведенных ниже диаграммах показаны две широко поддерживаемые реконструкции:
|
|
Морфологический анализ, как правило, позволяет выявить кладу конхифер, которая в меньшей степени подтверждается молекулярными анализами.[77] хотя эти результаты также приводят к неожиданным парафилиям, например, к рассеянию двустворчатых моллюсков по всем другим группам моллюсков.[78]
Однако анализ, проведенный в 2009 г. с использованием обоих морфологический и молекулярная филогенетика сравнения пришли к выводу, что моллюски не монофилетический; особенно, Scaphopoda и Двустворчатые моллюски обе являются отдельными монофилетическими линиями, не связанными с остальными классами моллюсков; традиционный тип Mollusca - это полифилетический, и это может быть сделано только монофилетическим, если исключить scaphopods и двустворчатые моллюски.[73] Анализ 2010 года выявил традиционные группы конкиферанов и акулиферанов и показал, что моллюски были монофилетическими, что свидетельствует о загрязнении имеющихся данных по соленогастрам.[79] Современные молекулярные данные недостаточны, чтобы ограничить филогенез моллюсков, а поскольку методы, используемые для определения достоверности кладов, склонны к переоценке, рискованно уделять слишком много внимания даже тем областям, по которым разные исследования согласны.[80] Вместо того, чтобы исключить маловероятные отношения, последние исследования добавляют новые перестановки внутренних взаимоотношений моллюсков, даже ставя под сомнение гипотезу раковинных форм.[81]
Человеческое взаимодействие
На протяжении тысячелетий моллюски были источником пищи для людей, а также важными предметами роскоши, в частности жемчуг, перламутр, Тирийский фиолетовый краситель морской шелк, и химические соединения. Их раковины также использовались как форма валюта в некоторых доиндустриальных обществах. Некоторые виды моллюсков могут кусать или ужалить людей, а некоторые из них стали сельскохозяйственными вредителями.
Использование людьми
Моллюски, особенно двустворчатые, такие как моллюски и моллюски, были важным источником пищи, по крайней мере, с момента появления людей современного анатомического типа, и это часто приводило к перелову рыбы.[82] Другие часто употребляемые в пищу моллюски включают: осьминоги и кальмары, щенячки, устрицы, и гребешки.[83] В 2005 г. на долю Китая приходилось 80% мирового улова моллюсков, что составило почти 11 000 000 тонн (11 000 000 длинных тонн; 12 000 000 коротких тонн). В Европе Франция оставалась лидером отрасли.[84] Некоторые страны регулируют импорт и транспортировку моллюсков и других морепродукты, в основном, чтобы минимизировать риск отравления токсины которые иногда могут накапливаться у животных.[85]

Большинство моллюсков с раковинами могут производить жемчуг, но только жемчуг двустворчатые моллюски и немного брюхоногие моллюски, чьи оболочки облицованы перламутр, ценны.[14](стр300–343, 367–403) Лучший натуральный жемчуг производит морской жемчужные устрицы, Pinctada маргаритифера и Pinctada mertensi, которые живут в тропический и субтропический воды Тихий океан. Натуральный жемчуг образуется, когда небольшой посторонний предмет застревает между мантия и оболочка.
Два метода культивирования жемчуг вставьте в устрицы либо «семечки», либо бусинки. В «семенном» методе используются зерна донной скорлупы из пресноводных источников. моллюски, и чрезмерный сбор для этой цели находящихся под угрозой исчезновения несколько видов пресноводных мидий на юго-востоке США.[14](pp367–403) Жемчужная промышленность настолько важна в некоторых областях, что значительные суммы денег тратятся на наблюдение за здоровьем выращиваемых моллюсков.[86]

Другая роскошь и высокаястатус изделия изготавливались из моллюсков. Тирийский фиолетовый, сделанный из чернильных желез мурекс ракушки, "на вес серебра" в четвертом веке до н.э, согласно с Теопомп.[87] Открытие большого количества Murex снаряды на Крит предлагает Минойцы возможно, был пионером добычи «императорского пурпура» в среднеминойский период в 20–18 веках до нашей эры, за столетия до Тирийцы.[88][89] Морской шелк прекрасный, редкий и ценный ткань производится из длинных шелковистых нитей (Бисс ) секретируется несколькими двустворчатыми моллюсками, в частности Pinna nobilis, чтобы прикрепиться к морскому дну.[90] Прокопий, пишущие о персидских войнах около 550 CE, "заявил, что пять потомственных сатрапов (правителей) Армения получившие знаки отличия от римского императора получили хламиды (или плащи) из Лана Пинна. Судя по всему, носить эти хламиды разрешалось только правящим классам ".[91]
Раковины моллюсков, в том числе каури, использовались как своего рода Деньги (ракушечные деньги ) в нескольких доиндустриальных обществах. Однако эти «валюты» в целом существенно отличались от стандартных денег, поддерживаемых и контролируемых государством, знакомых индустриальным обществам. Некоторые "валюты"-оболочки не использовались для коммерческих сделок, а в основном использовались как социальный статус показы в важных случаях, например на свадьбах.[92] При использовании для коммерческих сделок они функционировали как товар Деньги как товар, который можно торговать, стоимость которого варьировалась от места к месту, часто из-за трудностей с транспортировкой, и который был уязвим для неизлечимых инфляция если появится более эффективный транспорт или поведение «золотой лихорадки».[93]
Биоиндикаторы
Двустворчатые моллюски используются как биоиндикаторы для мониторинга состояния водной среды как в пресной, так и в морской среде. Их популяционный статус или структура, физиология, поведение или уровень загрязнения элементами или соединениями могут указывать на состояние загрязнения экосистемы. Они особенно полезны, так как они сидячие и репрезентативны для среды, в которой они отбираются или помещаются.[94] Potamopyrgus antipodarum используется некоторыми водоочистными сооружениями для проверки на эстроген-имитирующие загрязнители от промышленного сельского хозяйства.
Вреден для человека
Укусы и укусы

Некоторые моллюски жалят или кусаются, но смерть от ядов моллюсков составляет менее 10% смертей от медуза укусы.[96]
Все осьминоги ядовиты,[97] но лишь несколько видов представляют значительную угрозу для человека. Осьминоги с синими кольцами в роду Hapalochlaena, которые живут вокруг Австралии и Новой Гвинеи, кусают людей, только если их сильно спровоцировать,[95] но их яд убивает 25% человеческих жертв. Еще один тропический вид, Осьминог аполлион, вызывает серьезные воспаление которые могут длиться более месяца даже при правильном лечении,[98] и укус Осьминог rubescens может вызвать некроз это длится дольше одного месяца при отсутствии лечения, а головные боли и слабость сохраняются до недели, даже если лечить.[99]

Все виды конусные улитки ядовиты и могут болезненно ужалить при обращении с ними, хотя многие виды слишком малы, чтобы представлять большую опасность для людей, и достоверно зарегистрировано лишь несколько смертельных случаев. Их яд представляет собой сложную смесь токсины, некоторые быстро действуют, а другие медленнее, но опаснее.[100][96][101] Воздействие отдельных токсинов конической раковины на нервную систему жертв настолько точно, что может быть полезным инструментом для исследований в неврология, а небольшой размер их молекулы позволяет легко их синтезировать.[100][102]
Переносчики болезней

Шистосомоз (также известная как бильгарциоз, бильгарциоз или улитка), заболевание, вызываемое червем двуустки. Шистосома, является «вторым после малярии самым разрушительным паразитарным заболеванием в тропических странах. По оценкам, этой болезнью инфицированы 200 миллионов человек в 74 странах - 100 миллионов только в Африке».[103] У паразита 13 известных видов, два из которых инфицируют людей. Сам по себе паразит не является моллюском, но у всех видов есть пресноводные улитки. промежуточные хосты.[104]
Вредители
Некоторые виды моллюсков, особенно некоторые улитки и слизни, могут быть серьезными вредителями сельскохозяйственных культур,[105] и при внедрении в новую среду может нарушить баланс локальных экосистемы. Один из таких вредителей - гигантская африканская улитка. Ахатина фулика, был завезен во многие части Азии, а также на многие острова в Индийский океан и Тихий океан. В 1990-х годах этот вид достиг Вест-Индии. Попытки управлять им, введя хищную улитку Euglandina rosea оказался катастрофическим, поскольку хищник проигнорировал Ахатина фулика и вместо этого продолжил истребление нескольких местных видов улиток.[106]
Смотрите также
Заметки
использованная литература
- ^ а б c d Чепмен, А.Д. (2009). Количество живых видов в Австралии и в мире (2-е (печатное) изд.). Канберра: Австралийское исследование биологических ресурсов. ISBN 978-0-642-56860-1. Получено 12 января 2010.; ISBN 978-0-642-56861-8 (интернет-издание).
- ^ Розенберг, Гэри (1996). "Mollusckque - Mollusk vs. Mollusc". Архивировано из оригинал 3 марта 2012 г.
- ^ Розенберг, Гэри (2014). «Новая критическая оценка видового разнообразия современных моллюсков». Американский малакологический бюллетень. 32 (2): 308–322. Дои:10.4003/006.032.0204. S2CID 86761029.
- ^ Taylor, P.D .; Льюис, Д.Н. (2005). Ископаемые беспозвоночные. Издательство Гарвардского университета.
- ^ Федосов, Александр Е .; Пуйландре, Николя (2012). «Филогения и таксономия комплекса рода Kermia – Pseudodaphnella (Mollusca: Gastropoda: Raphitomidae): замечательное излучение за счет диверсификации развития личинок» (PDF). Систематика и биоразнообразие. 10 (4): 447–477. Дои:10.1080/14772000.2012.753137. S2CID 55028766.
- ^ μαλάκια, μαλακός. Лидделл, Генри Джордж; Скотт, Роберт; Греко-английский лексикон на Проект Персей.
- ^ Аристотель. «Книга I часть 1, Книга IV часть 1 и т. Д.». История животных.
- ^ Little, L .; Fowler, H.W .; Coulson, J .; Onions, C.T., eds. (1964). «Малакология». Краткий оксфордский словарь английского языка. Издательство Оксфордского университета.
- ^ Чисхолм, Хью, изд. (1911). . Британская энциклопедия. 18 (11-е изд.). Издательство Кембриджского университета. п. 675.
- ^ Хоган, С. Майкл. (2010). «Кальций». В Jorgensen, A .; Кливленд, К. (ред.). Энциклопедия Земли. Национальный совет по науке и окружающей среде.
- ^ а б c d Giribet, G .; Окусу, А .; Lindgren, A.R .; Хафф, S.W .; Schrödl, M .; Нисигучи, М. (Май 2006 г.). «Доказательства клады, состоящей из моллюсков с последовательно повторяющимися структурами: моноплакофораны родственны хитонам». Труды Национальной академии наук Соединенных Штатов Америки. 103 (20): 7723–7728. Bibcode:2006PNAS..103.7723G. Дои:10.1073 / pnas.0602578103. ЧВК 1472512. PMID 16675549.
- ^ а б c d е ж г час Хейворд, П.Дж. (1996). Справочник морской фауны Северо-Западной Европы. Издательство Оксфордского университета. С. 484–628. ISBN 978-0-19-854055-7.
- ^ а б c Бруска, Р. И Бруска, Г.Дж. (2003). Беспозвоночные (2-е изд.). Sinauer Associates. п. 702. ISBN 978-0-87893-097-5.
- ^ а б c d е ж г час я j k л м п о п q р s т ты v Ruppert, E.E .; Fox, R.S .; Барнс, Р.Д. (2004). Зоология беспозвоночных (7-е изд.). Брукс / Коул. ISBN 978-0-03-025982-1.
- ^ а б c d е ж г час я j k л м п о п [14](p284–291)
- ^ а б c d Ponder, W.F .; Линдберг, Д. Р., ред. (2008). Филогения и эволюция Mollusca. Беркли, Калифорния: Калифорнийский университет Press. п. 481. ISBN 978-0-520-25092-5.
- ^ Николь, Дэвид (июнь 1969). «Число живых видов моллюсков». Систематическая зоология. 18 (2): 251–254. Дои:10.2307/2412618. JSTOR 2412618.
- ^ а б c Хашпрунар, Г. (2001). «Моллюски (Моллюски)». Энциклопедия наук о жизни. John Wiley & Sons, Ltd. Дои:10.1038 / npg.els.0001598. ISBN 978-0470016176.
- ^ Хэнкок, Ребекка (2008). «Признание исследования моллюсков». Австралийский музей. Архивировано из оригинал 30 мая 2009 г.. Получено 9 марта 2009.
- ^ Вдумайтесь, Уинстон Ф. и Линдберг, Дэвид Р. (2004). «Филогения моллюсков» (Пресс-релиз). Всемирный конгресс малакологии. Получено 9 марта 2009.
- ^ Рауп, Дэвид М. и Стэнли, Стивен М. (1978). Принципы палеонтологии (2-е изд.). W.H. Фримен и Ко, стр. 4–5. ISBN 978-0716700227.
- ^ Barnes, R.S.K .; Calow, P .; Olive, P.J.W .; Golding, D.W .; Спайсер, Дж. (2001). Беспозвоночные: синтез (3-е изд.). Великобритания: Blackwell Science.
- ^ Кубодера, Т .; Мори, К. (22 декабря 2005 г.). «Первые наблюдения за живым гигантским кальмаром в дикой природе» (PDF). Труды Королевского общества B. 272 (1581): 2583–2586. Дои:10.1098 / rspb.2005.3158. ЧВК 1559985. PMID 16321779. Архивировано из оригинал (PDF) 3 июня 2016 г.. Получено 22 октября 2008.
- ^ Блэк, Ричард (26 апреля 2008 г.). «Колоссальный кальмар из морозилки». Новости BBC. Получено 1 октября 2008.
- ^ Lydeard, C .; Cowie, R .; Ponder, W.F .; и другие. (Апрель 2004 г.). «Глобальное сокращение неморских моллюсков» (PDF). Бионаука. 54 (4): 321–330. Дои:10.1641 / 0006-3568 (2004) 054 [0321: TGDONM] 2.0.CO; 2. Архивировано 31 марта 2007 года.CS1 maint: неподходящий URL (ссылка на сайт)
- ^ а б Хили, Дж. М. (2001). «Моллюска». В Андерсоне, Д.Т. (ред.). Зоология беспозвоночных (2-е изд.). Издательство Оксфордского университета. С. 120–171. ISBN 978-0-19-551368-4.
- ^ а б Портер, С. (1 июня 2007 г.). «Химия морской воды и ранняя биоминерализация карбонатов». Наука. 316 (5829): 1302. Bibcode:2007Научный ... 316.1302P. Дои:10.1126 / science.1137284. PMID 17540895. S2CID 27418253.
- ^ Йохельсон, Э. Л. (1975). «Обсуждение раннекембрийских» моллюсков"" (PDF). Журнал геологического общества. 131 (6): 661–662. Bibcode:1975JGSoc.131..661.. Дои:10.1144 / gsjgs.131.6.0661. S2CID 219540340.
- ^ Чернс, Л. (декабрь 2004 г.). «Раннепалеозойское разнообразие хитонов (Polyplacophora, Mollusca) на основе новых данных из силурия Готланда, Швеция». Lethaia. 37 (4): 445–456. Дои:10.1080/00241160410002180.
- ^ Томпа, А. С. (декабрь 1976 г.). «Сравнительное исследование ультраструктуры и минералогии кальцинированных яиц наземных улиток (Pulmonata: Stylommatophora)» (PDF). Журнал морфологии. 150 (4): 861–887. Дои:10.1002 / jmor.1051500406. HDL:2027.42/50263. PMID 30257539. S2CID 52844967.
- ^ Мур, Джанет (21 сентября 2006 г.). Знакомство с беспозвоночными. ISBN 9781139458474. Архивировано из оригинал на 2020-01-14. Получено 2019-06-04.
- ^ а б c Wilbur, Karl M .; Trueman, E.R .; Кларк, М.Р., ред. (1985), Моллюска, 11. Форма и функция, Нью-Йорк: Academic Press, ISBN 0-12-728702-7 стр. 4
- ^ Shigeno, S .; Сасаки, Т .; Moritaki, T .; Касугай, Т .; Vecchione, M .; Агата, К. (январь 2008 г.). «Эволюция головного комплекса головоногих моллюсков путем сборки нескольких частей тела: свидетельства эмбрионального развития наутилуса». Журнал морфологии. 269 (1): 1–17. Дои:10.1002 / jmor.10564. PMID 17654542. S2CID 13109195.
- ^ Ruppert, E.E .; Фокс, Р. И Барнс, Р.Д. (2004). «Моллюска». Зоология беспозвоночных (7-е изд.). Брукс / Коул. С. 290–291. ISBN 0030259827.
- ^ Марин, Ф .; Люке, Г. (октябрь 2004 г.). «Белки раковины моллюсков». Comptes Rendus Palevol. 3 (6–7): 469. Дои:10.1016 / j.crpv.2004.07.009.
- ^ Steneck, R.S .; Уотлинг, Л. (июль 1982 г.). «Возможности кормления и ограничения растительноядных моллюсков: функционально-групповой подход». Морская биология. 68 (3): 299–319. Дои:10.1007 / BF00409596. S2CID 84207061.
- ^ Тендал О.С. (1985). «Xenophyophores (Protozoa, Sarcodina) в рационе Neopilina galatheae (Mollusca, Monoplacophora) " (PDF). Отчет Галатеи. 16: 95–98. Архивировано из оригинал (PDF) в 2012-11-30. Получено 2013-09-14.
- ^ Дженсен, К. Р. (февраль 1993 г.). «Морфологические адаптации и пластичность радулярных зубов Sacoglossa (= Ascoglossa) (Mollusca: Opisthobranchia) по отношению к их пищевым растениям». Биологический журнал Линнеевского общества. 48 (2): 135–155. Дои:10.1111 / j.1095-8312.1993.tb00883.x.
- ^ Вегеле, Х. (март 1989 г.). «Рацион некоторых антарктических голожаберников (Gastropoda, Opisthobranchia, Nudibranchia)». Морская биология. 100 (4): 439–441. Дои:10.1007 / BF00394819. S2CID 83444088.
- ^ Издатели, Bentham Science (июль 1999 г.). Современная органическая химия. Издательство Bentham Science.
- ^ Ламберт, W.J.(1 октября 1991 г.). "Сосуществование голожаберников, питающихся гидроидами: имеет ли значение биология питания и среда обитания?".[постоянная мертвая ссылка ]
- ^ Кларксон, E.N.K. (1998). Палеонтология и эволюция беспозвоночных. Блэквелл. п. 221. ISBN 978-0-632-05238-7.
- ^ а б c d е Runnegar, B .; Pojeta Jr, J. (октябрь 1974 г.). «Филогения моллюсков: палеонтологическая точка зрения». Наука. 186 (4161): 311–317. Bibcode:1974Наука ... 186..311R. Дои:10.1126 / science.186.4161.311. JSTOR 1739764. PMID 17839855. S2CID 46429653.
- ^ Kocot, K. M .; Cannon, J. T .; Todt, C .; Citarella, M. R .; Кон, А. Б .; Мейер, А .; Santos, S. R .; Schander, C .; Мороз, Л. Л .; и другие. (22 сентября 2011 г.). «Филогеномика обнаруживает глубокие взаимоотношения моллюсков». Природа. 477 (7365): 452–456. Bibcode:2011Натура.477..452K. Дои:10.1038 / природа10382. ЧВК 4024475. PMID 21892190.
- ^ Бадд, Г. Э. и Дженсен, С. Критическая переоценка летописи окаменелостей билатерианского типа. Биол. Ред. 75, 253–295 (2000).
- ^ Федонкин, М.А .; Ваггонер, Б. (28 августа 1997 г.). «Позднедокембрийская окаменелость Kimberella - это двустворчатый организм, подобный моллюску». Природа. 388 (6645): 868. Bibcode:1997Натура.388..868F. Дои:10.1038/42242. S2CID 4395089.
- ^ а б Федонкин, М.А .; Simonetta, A .; Иванцов, А. (2007). «Новые данные о кимберелле, вендском моллюскоподобном организме (Белое море, Россия): палеоэкологические и эволюционные последствия» (PDF). Геологическое общество, Лондон, Специальные публикации. 286 (1): 157–179. Bibcode:2007GSLSP.286..157F. Дои:10.1144 / SP286.12. S2CID 331187. Архивировано из оригинал (PDF) на 2012-11-22. Получено 2008-07-10.
- ^ а б c Баттерфилд, Нью-Джерси (2006). «Зацепление некоторых« червей »стеблевой группы: ископаемых лофотрохозоидов в сланцах Берджесс». BioEssays. 28 (12): 1161–6. Дои:10.1002 / bies.20507. PMID 17120226. S2CID 29130876.
- ^ а б c Sigwart, J.D .; Саттон, М. Д. (октябрь 2007 г.). «Глубокая филогения моллюсков: синтез палеонтологических и неонтологических данных». Труды Королевского общества B: биологические науки. 274 (1624): 2413–2419. Дои:10.1098 / rspb.2007.0701. ЧВК 2274978. PMID 17652065. Для краткого обзора см. "Моллюска". Музей палеонтологии Калифорнийского университета. Получено 2008-10-02.
- ^ Бадд, Г. Э. и С. Йенсен. 2016: Происхождение животных и гипотеза «саванны» о ранней билатерианской эволюции. Биологические обзоры 7: Интернет впереди печати.
- ^ а б Caron, J.B .; Scheltema, A .; Schander, C .; Рудкин Д. (13 июля 2006 г.). «Мягкотелый моллюск с радулой из среднекембрийских сланцев Берджесс». Природа. 442 (7099): 159–163. Bibcode:2006Натура 442..159С. Дои:10.1038 / природа04894. HDL:1912/1404. PMID 16838013. S2CID 4431853.
- ^ а б Баттерфилд, Нью-Джерси (май 2008 г.). «Раннекембрийская радула». Журнал палеонтологии. 82 (3): 543–554. Дои:10.1666/07-066.1. S2CID 86083492.
- ^ Cruz, R .; Lins, U .; Фарина, М. (1998). «Минералы радулярного аппарата Falcidens sp. (Caudofoveata) и эволюционные последствия для Phylum Mollusca ". Биологический бюллетень. 194 (2): 224–230. Дои:10.2307/1543051. JSTOR 1543051. PMID 28570844.
- ^ Пархаев, П.Ю. (2007). Кембрийский «фундамент» эволюции брюхоногих моллюсков. Геологическое общество, Лондон, Специальные публикации. 286. С. 415–421. Bibcode:2007GSLSP.286..415P. Дои:10.1144 / SP286.31. ISBN 978-1-86239-233-5. S2CID 130979274. Получено 2009-11-01.
- ^ Steiner, M .; Li, G .; Qian, Y .; Zhu, M .; Erdtmann, B.D. (2007). «Неопротерозойские и ранние кембрийские комплексы окаменелостей ракушечников и пересмотренная биостратиграфическая корреляция платформы Янцзы (Китай)». Палеогеография, палеоклиматология, палеоэкология. 254 (1–2): 67. Bibcode:2007ППП ... 254 ... 67С. Дои:10.1016 / j.palaeo.2007.03.046.
- ^ Mus, M.M .; Palacios, T .; Дженсен, С. (2008). «Размер самых ранних моллюсков: выросли ли маленькие гелионеллиды, чтобы стать большими взрослыми?». Геология. 36 (2): 175. Bibcode:2008Гео .... 36..175M. Дои:10.1130 / G24218A.1.
- ^ Посадка, Е .; Гейер, G .; Бартовски, К. (2002). «Последние раннекембрийские окаменелости малых раковин, трилобиты и дизаэробный интервал Хэтч-Хилл на континентальном склоне Квебека». Журнал палеонтологии. 76 (2): 287–305. Дои:10.1666 / 0022-3360 (2002) 076 <0287: LECSSF> 2.0.CO; 2. JSTOR 1307143.
- ^ Frýda, J .; Nützel, A .; Вагнер, П.Дж. (2008). «Палеозойские брюхоногие моллюски». In Ponder, W.F .; Линдберг, Д. (ред.). Филогения и эволюция Mollusca. California Press. С. 239–264. ISBN 978-0-520-25092-5.
- ^ Кучинский, А. (2000). «Микроструктуры раковин раннекембрийских моллюсков» (PDF). Acta Palaeontologica Polonica. 45 (2): 119–150. Получено 4 ноября 2009.
- ^ Хагадорн, Дж. И Ваггонер, Б. (2002). «Раннекембрийская проблемная окаменелость Volborthella: новые взгляды на бассейн и хребет». В Корсетти, Ф.А. (ред.). Протерозой-кембрий Большого бассейна и за его пределами, Тихоокеанский раздел SEPM Book 93 (PDF). SEPM (Общество осадочной геологии). С. 135–150. Архивировано 11 сентября 2006 года.CS1 maint: BOT: статус исходного URL-адреса неизвестен (ссылка на сайт)
- ^ Vickers-Rich, P .; Fenton, C.L .; Fenton, M.A .; Рич, Т. (1997). Книга окаменелостей: летопись доисторической жизни. Courier Dover Publications. стр.269–272. ISBN 978-0-486-29371-4.
- ^ Marshall C.R .; Уорд П.Д. (1996). «Внезапное и постепенное вымирание моллюсков в позднем меловом периоде западноевропейского Тетиса». Наука. 274 (5291): 1360–1363. Bibcode:1996Научный ... 274.1360M. Дои:10.1126 / science.274.5291.1360. PMID 8910273. S2CID 1837900.
- ^ Монахов, Н. "Широкая история головоногих моллюсков". Получено 2009-03-21.
- ^ Pojeta, J. (2000). «Кембрийские Pelecypoda (Mollusca)». Американский малакологический бюллетень. 15: 157–166.
- ^ Шнайдер, Дж. (2001). «Систематика двустворчатых моллюсков в ХХ веке». Журнал палеонтологии. 75 (6): 1119–1127. Дои:10.1666 / 0022-3360 (2001) 075 <1119: BSDTC> 2.0.CO; 2.
- ^ Губанов, А.П .; Кучинский, А.В .; Пил, Дж. (2007). «Первая эволюционно-адаптивная линия ископаемых моллюсков». Lethaia. 32 (2): 155. Дои:10.1111 / j.1502-3931.1999.tb00534.x.
- ^ Губанов, А.П .; Пил, Дж. (2003). "Раннекембрийский гелионеллоидный моллюск Анабарелла Востокова ». Палеонтология. 46 (5): 1073–1087. Дои:10.1111/1475-4983.00334.
- ^ Цзун-Цзе, Ф. (2006). «Введение в ордовикских двустворчатых моллюсков южного Китая с обсуждением ранней эволюции двустворчатых моллюсков». Геологический журнал. 41 (3–4): 303–328. Дои:10.1002 / gj.1048.
- ^ Raup, D.M .; Яблонски, Д. (1993). «География вымирания морских двустворчатых моллюсков в конце мелового периода». Наука. 260 (5110): 971–973. Bibcode:1993Научный ... 260..971R. Дои:10.1126 / science.11537491. PMID 11537491.
- ^ Малинки, Дж. М. (2009). «Пермская хиолитида из Австралии: последний из хиолитов?». Журнал палеонтологии. 83: 147–152. Дои:10.1666 / 08-094R.1. S2CID 85924056.
- ^ а б c d е Sigwart, J.D .; Саттон, доктор медицины (октябрь 2007 г.). «Глубокая филогения моллюсков: синтез палеонтологических и неонтологических данных». Труды Королевского общества B. 274 (1624): 2413–2419. Дои:10.1098 / rspb.2007.0701. ЧВК 2274978. PMID 17652065. Для краткого обзора см. "Моллюска". Музей палеонтологии Калифорнийского университета. Получено 2008-10-02.
- ^ "Моллюска". Музей палеонтологии Калифорнийского университета. Получено 2008-10-02.
- ^ а б Голобов, Пабло А .; Каталано, Сантьяго А .; Миранде, Дж. Маркос; Szumik, Claudia A .; Ариас, Дж. Сальвадор; Келлэрсьё, Мари; Фаррис, Джеймс С. (2009). «Филогенетический анализ 73 060 таксонов подтверждает основные группы эукариот». Кладистика. 25 (3): 211–230. Дои:10.1111 / j.1096-0031.2009.00255.x.
- ^ «Знакомство с Lophotrochozoa». Музей палеонтологии Калифорнийского университета. Получено 2008-10-02.
- ^ а б Генри, Дж .; Окусу, А .; Мартиндейл, М. (2004). «Клеточная линия полиплакофорана, Chaetopleura apiculata: вариации в спиральной программе и значение для эволюции моллюсков». Биология развития. 272 (1): 145–160. Дои:10.1016 / j.ydbio.2004.04.027. PMID 15242797.
- ^ а б Джейкобс, Д.К .; Wray, C.G .; Wedeen, C.J .; Кострикен, Р .; Desalle, R .; Staton, J. L .; Гейтс, Р.Д .; Линдберг, Д. (2000). «Закрепленное выражение моллюсков, серийная организация и эволюция раковины». Эволюция и развитие. 2 (6): 340–347. Дои:10.1046 / j.1525-142x.2000.00077.x. PMID 11256378. S2CID 25274057.
- ^ Виннепеннинкс, В; Бакеляу, Т; Де Вахтер, Р. (1996). «Исследование филогении моллюсков на основе последовательностей 18S рРНК». Молекулярная биология и эволюция. 13 (10): 1306–1317. Дои:10.1093 / oxfordjournals.molbev.a025577. PMID 8952075.
- ^ Passamaneck, Y .; Schander, C .; Галаныч, К. (2004). «Исследование филогении моллюсков с использованием больших и малых субъединиц ядерных последовательностей рРНК». Молекулярная филогенетика и эволюция. 32 (1): 25–38. Дои:10.1016 / j.ympev.2003.12.016. PMID 15186794.
- ^ Wilson, N .; Rouse, G .; Гирибет, Г. (2010). «Оценка гипотезы моллюсков Serialia (Monoplacophora + Polyplacophora) с использованием новых молекулярных данных». Молекулярная филогенетика и эволюция. 54 (1): 187–193. Дои:10.1016 / j.ympev.2009.07.028. PMID 19647088.
- ^ Wägele, J .; Letsch, H .; Klussmann-Kolb, A .; Mayer, C .; Misof, B .; Вегеле, Х. (2009). «Значения филогенетической поддержки не обязательно информативны: случай гипотезы Serialia (филогения моллюска)». Границы зоологии. 6 (1): 12. Дои:10.1186/1742-9994-6-12. ЧВК 2710323. PMID 19555513.
- ^ Vinther, J .; Sperling, E. A .; Бриггс, Д.Э.; Петерсон, К. Дж. (2011). «Молекулярная палеобиологическая гипотеза происхождения моллюсков-аплакофоранов и их происхождение от хитоноподобных предков». Труды Королевского общества B: биологические науки. 279 (1732): 1259–68. Дои:10.1098 / rspb.2011.1773. ЧВК 3282371. PMID 21976685.
- ^ Mannino, M.A .; Томас, К. (2002). «Истощение ресурсов? Влияние доисторических поисков пищи человеком на сообщества приливных моллюсков и его значение для расселения, мобильности и расселения людей». Мировая археология. 33 (3): 452–474. Дои:10.1080/00438240120107477. JSTOR 827879. S2CID 161085658.
- ^ Garrow, J.S .; Ральф, А .; Джеймс, W.P.T. (2000). Питание человека и диетология. Elsevier Health Sciences. п. 370. ISBN 978-0-443-05627-7.
- ^ «В 2005 году Китай выловил почти 11 млн тонн моллюсков». ФАО. Получено 2008-10-03.
- ^ «Импорт рыбной продукции или двустворчатых моллюсков». Великобритания: Агентство пищевых стандартов. Получено 2008-10-02.
- ^ Jones, J.B .; Крипер, Дж. (Апрель 2006 г.). «Болезни жемчужных устриц и других моллюсков: перспектива Западной Австралии». Журнал исследований моллюсков. 25 (1): 233–238. Дои:10.2983 / 0730-8000 (2006) 25 [233: DOPOAO] 2.0.CO; 2.
- ^ Четвертый век до н.э историк Теопомп цитируется Афиней (12: 526) около 200 г. до н. э .; согласно с Гулик, C.B. (1941). Афиней, Деипнософисты. Кембридж, Массачусетс: Издательство Гарвардского университета. ISBN 978-0-674-99380-8.
- ^ Риз, Д.С. (1987). «Производство ракушек Палаикастро и пурпурных красок бронзового века в Средиземноморском бассейне». Ежегодник Британской школы археологии в Афинах. 82: 201–6. Дои:10,1017 / с0068245400020438.
- ^ Штиглиц, Р.Р. (март 1994). «Минойское происхождение тирского пурпура». Библейский археолог. 57 (1): 46–54. Дои:10.2307/3210395. JSTOR 3210395. S2CID 163601220.
- ^ Третий новый международный словарь Вебстера (без сокращений) 1976. G. & C. Merriam Co., p. 307.
- ^ Тернер, Р.Д .; Роузуотер, Дж. (Июнь 1958 г.). «Семья Pinnidae в Западной Атлантике». Johnsonia. 3 (38): 294.
- ^ Маурер, Б. (октябрь 2006 г.). «Антропология денег» (PDF). Ежегодный обзор антропологии. 35: 15–36. Дои:10.1146 / annurev.anthro.35.081705.123127. Архивировано из оригинал (PDF) на 2007-08-16.
- ^ Хогендорн, Дж. И Джонсон, М. (2003). Ракушечные деньги работорговли. Издательство Кембриджского университета. ISBN 978-0521541107. В частности, главы «Бум и спад в торговле каури» (страницы 64–79) и «Каури как деньги: транспортные расходы, ценности и инфляция» (страницы 125–147)
- ^ Университет Бордо; и другие. "МоллюСКАН" глаз проект ". Получено 2017-01-28.
- ^ а б Алафачи, А. (5 июня 2018 г.). «Синекольчатый осьминог». Австралийский отдел исследования яда. Получено 2008-10-03.
- ^ а б Williamson, J.A .; Fenner, P.J .; Burnett, J.W .; Рифкин, Дж. (1996). Ядовитые и ядовитые морские животные: медико-биологический справочник. UNSW Press. С. 65–68. ISBN 978-0-86840-279-6.
- ^ Андерсон, Р. (1995). «Аквариумное разведение гигантского тихоокеанского осьминога». Барабан и горбун. 26: 14–23.
- ^ Brazzelli, V .; Baldini, F .; Nolli, G .; Borghini, F .; Боррони, Г. (март 1999 г.). "Осьминог аполлион укус ». Контактный дерматит. 40 (3): 169–70. Дои:10.1111 / j.1600-0536.1999.tb06025.x. PMID 10073455. S2CID 35988014.
- ^ Андерсон, Р. (1999). «Укус осьминога и его лечение». Фестивус. 31: 45–46.
- ^ а б c Конкар, Д. (19 октября 1996 г.). «Докторская улитка - смертельный для рыб, а иногда и людей, яд конусной улитки содержит фармакопею точных лекарств». Новый ученый. Получено 2008-10-03.
- ^ Ливетт, Б. "Отравление конусным моллюском, с отчетом о смертельном случае". Кафедра биохимии и молекулярной биологии Университета Мельбурн. Архивировано из оригинал на 2012-12-01. Получено 2008-10-03.
- ^ Haddad Junior, V .; Паула Нето, J.O.B.D .; Кобо, В.Л.Дж. (Сентябрь – октябрь 2006 г.). «Ядовитые моллюски: риски несчастных случаев, вызванных улитками-конусами (брюхоногие моллюски: Conidae) в Бразилии». Revista da Sociedade Brasileira de Medicina Tropical. 39 (5): 498–500. Дои:10.1590 / S0037-86822006000500015. PMID 17160331.
- ^ "Программа борьбы с шистосомозом Центра Картера". В Картер Центр. Получено 2008-10-03.
- ^ Браун, Д.С. (1994). Пресноводные улитки Африки и их медицинское значение. CRC Press. п. 305. ISBN 978-0-7484-0026-3.
- ^ Баркер, Г. (2002). Моллюски как вредители сельскохозяйственных культур. CABI Publications. ISBN 978-0-85199-320-1.
- ^ Civeyrel, L .; Симберлофф, Д. (октябрь 1996 г.). «Сказка о двух улитках: неужели лекарство хуже болезни?». Биоразнообразие и сохранение. 5 (10): 1231–1252. Дои:10.1007 / BF00051574. S2CID 43071631.
дальнейшее чтение
- Sturm, C .; Пирс, Т. И Вальдес, А. Моллюски: руководство по их изучению, сбору и сохранению. Универсальные издатели. 2006. 454 с. ISBN 1581129300
- Trigo, J.E .; Díaz Agras, G.J .; Гарсиа-Альварес, О.Л .; Guerra, A .; Moreira, J .; Pérez, J .; Rolán, E .; Тронкосо, Дж. И Ургорри, В. (2018). Troncoso, J.S., Trigo, J.E. & Rolán, E., ed. Guía de los Moluscos Marinos de Galicia. Виго: Служба публикаций университета Виго. 836 страниц. ISBN 978-84-8158-787-6
внешние ссылки
 Данные, относящиеся к Моллюска в Wikispecies
Данные, относящиеся к Моллюска в Wikispecies- «Моллюска» на Энциклопедия жизни

- Исследователи составили эволюционное древо моллюсков; 26 октября 2011 г.
- Интернет-руководство Харди по морским брюхоногим моллюскам
- Роттердамский музей естественной истории Галерея изображений Shell
- Программа наблюдения за мидиями
- Онлайн-биомониторинг активности двустворчатых моллюсков, 24/7: MolluSCAN глаз