Мшанки - Bryozoa
| Мшанки | |
|---|---|
 | |
| «Мшанки», с Эрнст Геккель с Kunstformen der Natur, 1904 | |
| Научная классификация | |
| Королевство: | Animalia |
| Подцарство: | Eumetazoa |
| Clade: | ParaHoxozoa |
| Clade: | Bilateria |
| Clade: | Нефрозоа |
| (без рейтинга): | Протостомия |
| (без рейтинга): | Спиралия |
| Суперфилум: | Лофотрохозоа |
| Clade: | Лофофората |
| Тип: | Мшанки Эренберг, 1831 г.[3] |
| Классы | |
| Синонимы[4] | |
Ectoprocta (Nitsche, 1869) (ранее подтип Bryozoa) | |
Мшанки (также известный как Полизоа, Эктопрокта или обычно как мох животные)[5] площадь филюм из водный беспозвоночный животные. Обычно около 0,5 миллиметра (1⁄64 дюйм) в длину, они питатели-фильтры которые отсеивают частицы пищи из воды с помощью выдвижного лофофор, "корона" щупальца выложено реснички. Наиболее морской виды обитают в тропических водах, но некоторые встречаются в океанические желоба, а другие находятся в полярный воды. Один учебный класс живет только во множестве пресная вода окружающей среде, и некоторые представители в основном морского класса предпочитают солоноватая вода. 5869[6] живые виды известны. Один род одинокий, а остальные колониальный.
Первоначально этот тип назывался «Polyzoa», но в 1831 году этот термин был заменен на «Bryozoa». Другая группа животных, обнаруженная впоследствии, чей механизм фильтрации выглядел похожим, также была включена в «Bryozoa» до 1869 года, когда были отмечены две группы. сильно отличаться внутренне. Недавно обнаруженная группа получила название Энтопрокта, в то время как оригинальные «мшанки» назывались «Ectoprocta». Однако «мшанки» оставались более широко используемым термином для последней группы.
Особей в колониях мшанок (эктопроктов) называют зооиды, поскольку они не являются полностью независимыми животными. Все колонии содержат автозооиды, отвечающие за питание и выделение. Колонии некоторых классы имеют различные типы специализированных зооидов без кормления, некоторые из которых являются инкубаториями для оплодотворенных яиц, а некоторые классы также имеют специальные зооиды для защиты колонии. Класс Хейлостомы имеет наибольшее количество видов, возможно, потому, что у них самый широкий спектр специализированных зооидов. Некоторые виды могут ползать очень медленно, используя колючие защитные зооиды в качестве ног. Автозооиды поставляют питательные вещества непищающим зооидам по каналам, которые различаются между классами. Все зооиды, в том числе и одиночные, состоят из цистид который обеспечивает стенку тела и производит экзоскелет и полипид который содержит внутренние органы и лофофор или другие специальные расширения. У зооидов нет специальных выделительных органов, и полипиды автозооидов списываются, когда полипиды становятся перегруженными продуктами жизнедеятельности; обычно стенка тела затем вырастает замещающим полипидом. У автозооидов кишечник имеет U-образную форму, с ртом внутри «короны» щупалец и анусом снаружи. Колонии бывают самых разных форм, включая вееры, кусты и листы. Хейлостомы производят минерализованный экзоскелеты и образуют однослойные листы, покрывающие поверхности.
Зооиды всех пресноводных видов одновременны гермафродиты. Хотя представители многих морских видов функционируют сначала как самцы, а затем как самки, их колонии всегда содержат комбинацию зооидов, находящихся на мужской и женской стадиях. Все виды излучают сперма в воду. Некоторые также выпускают яйцеклетка в воду, в то время как другие захватывают сперму через щупальца, чтобы оплодотворить яйцеклетку изнутри. У некоторых видов личинки иметь большой желтки, идите кормить и быстро оседают на поверхности. У других личинок мало желтка, но они плавают и питаются несколько дней, прежде чем осесть. После заселения все личинки проходят радикальную метаморфоза который разрушает и восстанавливает почти все внутренние ткани. Пресноводные виды также производят статобласты которые лежат в спящем состоянии до тех пор, пока условия не станут благоприятными, что позволяет родословной колонии выжить, даже если суровые условия убивают материнскую колонию.
К хищникам морских мшанок относятся: голожаберники (морские слизни), рыба, морские ежи, пикногониды, ракообразные, клещи и морская звезда. Пресноводными мшанками охотятся улитки, насекомые и рыбы. В Таиланд, многие популяции одного пресноводного вида были уничтожены интродуцированные виды улитки. Быстрорастущие инвазивные мшанки у северо-восточного и северо-западного побережья США уменьшили ламинария лесов настолько сильно, что это повлияло на местные популяции рыб и беспозвоночных. Мшанки распространили болезни на рыбные хозяйства и рыбаки. Химические вещества, извлеченные из видов морских мшанок, были исследованы для лечения рака и Болезнь Альцгеймера, но анализы не обнадеживают.[7]
Минерализованные скелеты мшанок впервые появляются в породах с раннего Ордовик период,[1] что делает его последним крупным филумом, появившимся в летописи окаменелостей. Это заставило исследователей подозревать, что мшанки возникли раньше, но изначально были неминерализованными и могли значительно отличаться от окаменелых и современных форм. Ранние окаменелости имели в основном прямостоячие формы, но постепенно стали преобладать корковые формы. Неизвестно, является ли тип монофилетический. Эволюционные отношения мшанок с другими типами также неясны, отчасти потому, что на взгляд ученых на генеалогическое древо животных в основном влияют более известные типы. Обе морфологический и молекулярная филогения анализы расходятся во мнениях относительно взаимоотношений мшанок с энтопроками, относительно того, следует ли группировать мшанок с брахиоподы и форониды в Лофофората, и следует ли учитывать мшанок протостомы или же дейтеростомы.
Описание
Отличительные черты
Мшанки, форониды и брахиоподы процедить еду из воды с помощью лофофор, «корона» из полых щупалец. Мшанки образуют колонии, состоящие из клоны называемые зооидами, размер которых обычно составляет около 0,5 мм (1⁄64 в) длинный.[8] Форониды напоминают мшанок зооидов, но имеют длину от 2 до 20 см (от 1 до 8 дюймов) и, хотя часто растут группами, не образуют колоний, состоящих из клонов.[9] Брахиоподы, обычно считающиеся близкими родственниками мшанок и форонид, отличаются наличием раковин, похожих на раковины двустворчатые моллюски.[10] Все три из них тип есть целомудрие, внутренняя полость выстлана мезотелий.[8][9][10]Некоторые корковые колонии мшанок с минерализованный экзоскелеты очень похожи на маленькие кораллы. Однако колонии мшанок основаны анцеструлой, которая имеет округлую форму, а не форму нормального зооида этого вида. С другой стороны, основание полипа коралла имеет форму его дочерних полипов, а коралловые зооиды не имеют формы. целомудрие или же лофофор.[11]
Энтопрокты, другой тип фильтраторов, больше похожи на мшанок, но их лофофор -подобная структура питания имеет твердые щупальца, их анус лежит внутри, а не снаружи основания "короны" и у них нет целомудрие.[12]
| Мшанки[8] (Эктопрокта) | Другой лофофораты[13] | Другой Лофотрохозоа | Похожие на вид типы | |||
|---|---|---|---|---|---|---|
| Phoronida[9] | Брахиоподы[10] | Аннелида, Моллюска | Энтопрокта[12] | Кораллы (класс в типе Книдария )[11] | ||
| Целом | Трехчастный, если включена полость эпистома | Трехчастный | По одному на сегмент в базовой форме; слился с некоторыми таксоны | никто | ||
| Формирование целомудрие | Неуверенный, потому что метаморфоза личинок во взрослых особей невозможно отследить | Энтерокоэли | Schizocoely | непригодный | ||
| Лофофор | С полыми щупальцами | никто | Похожая на вид структура питания, но с твердыми щупальцами. | никто | ||
| Питающий ток | От кончиков до оснований щупалец | непригодный | От оснований до кончиков щупалец | непригодный | ||
| Многогранный клетки в эпителий | да[14] | нет[14] | да[14] | непригодный | ||
| Положение анус | За пределами базы лофофор | Варьируется, у некоторых видов отсутствует | Задняя часть, но нет Siboglinidae | Внутри базы лофофор -подобный орган | никто | |
| Колониальный | В большинстве своем колонии клонов; один одинокий род | Sessile виды часто образуют группы, но без активного сотрудничества | Колонии клонов у некоторых видов; некоторые одиночные виды | Колонии клонов | ||
| Форма основателя зооида | Круглый, в отличие от обычных зооидов[11] | непригодный | Как и другие зооиды | |||
| Минерализованный экзоскелеты | Немного таксоны | нет | Двустворчатый -подобные снаряды | У некоторых сидячих кольчатых червей образуются минерализованные трубки;[15] у большинства моллюсков есть раковины, но большинство современных головоногие моллюски есть внутренние оболочки или нет.[16] | нет | Некоторые таксоны |
Виды зооидов
Все мшанки колониальны, кроме одного род, Монобриозоид.[17][18] Размер отдельных членов колонии мшанок составляет около 0,5 мм (1⁄64 в) длинные и известны как зооиды,[8] поскольку они не являются полностью независимыми животными.[19] Все колонии содержат питающихся зооидов, известных как автозооиды, а в колониях некоторых групп также содержатся неспециализированные гетерозооиды;[18] члены колонии генетически идентичны и взаимодействуют друг с другом, как органы более крупных животных.[8] Какой тип зооидов растет в какой-либо колонии, определяется химическими сигналами от колонии в целом или иногда в ответ на запах хищников или конкурирующих колоний.[18]
Корпуса всех типов состоят из двух основных частей. В цистид состоит из стенки корпуса и любого типа экзоскелет является секретный посредством эпидермис. Экзоскелет может быть органическим (хитин, полисахарид или же белок ) или из минерала карбонат кальция. Стенка тела состоит из эпидермиса, базальная пластинка (мат из неячеистого материала), соединительная ткань, мышцы и мезотелий который выравнивает целомудрие (основная полость тела)[8] - разве что в одном учебный класс мезотелий разделен на два отдельных слоя: внутренний образует перепончатый мешок, который свободно плавает и содержит целом, а внешний, прикрепленный к стенке тела и окружающий перепончатый мешок в псевдоцелом.[20] Другая основная часть тела мшанки, известная как полипид и расположенный почти полностью внутри цистиды, содержит нервную систему, пищеварительную систему, некоторые специализированные мышцы и питающий аппарат или другие специализированные органы, которые заменяют питающий аппарат.[8]
Кормление зооидов
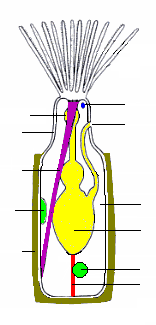
Наиболее распространенным типом зооидов является питающийся автозооид, в котором полипид несет «корону» из полых щупалец, называемую лофофор, который захватывает частицы пищи из воды.[18] Во всех колониях большой процент зооидов - автозооиды, а некоторые полностью состоят из автозооидов, некоторые из которых также участвуют в воспроизводстве.[21]
Основная форма «короны» - полный круг. в учебный класс Phylactolaemata корона кажется U-образной, но это впечатление создается глубокой вмятиной на краю короны, которая не имеет разрыва на краю щупалец.[8] По бокам щупалец покрыты тонкими волосками, которые называются реснички, чье биение заставляет водный поток от кончиков щупалец к их основаниям, где он и выходит. Частицы пищи, которые сталкиваются с щупальцами, улавливаются слизь, и дополнительные реснички на внутренней поверхности щупалец переносят частицы к рту, который находится в центре основания «короны».[22] Метод, используемый эктопроками, известен как «сбор вверх по потоку», поскольку частицы пищи улавливаются до того, как они пройдут через поле ресничек, которое создает питающий ток. Этот метод также используется форониды, брахиоподы и крылышки.[23]
Лофофор и рот устанавливаются на гибкой трубке, называемой «перевернутой», потому что ее можно вывернуть наизнанку и втянуть в полипид.[8] скорее как палец резиновой перчатки; в этом положении лофофор лежит внутри перевернутого и сложен, как спицы зонта. Обращение снимается, иногда в пределах 60миллисекунды парой втягивающих мышц, закрепленных на дальнем конце цистиды. Датчики на концах щупалец могут проверить признаки опасности до того, как перевернутый и лофофор полностью расправятся. Разгибание обусловлено увеличением внутреннего давления жидкости, которое виды с гибкими экзоскелетами производят за счет сокращения круговых мышц, лежащих внутри стенки тела,[8] в то время как виды с перепончатым мешком используют круговые мышцы, чтобы сжимать его.[20] Некоторые виды с жесткими экзоскелетами имеют гибкую мембрану, которая заменяет часть экзоскелета, а поперечные мышцы, закрепленные на дальней стороне экзоскелета, увеличивают давление жидкости, втягивая мембрану внутрь.[8] В других случаях в защитном скелете нет разрыва, и поперечные мышцы тянут гибкий мешок, который соединен с водой снаружи через небольшую пору; расширение мешка увеличивает давление внутри тела и выталкивает перевернутый и лофофор наружу.[8] У некоторых видов втянутый перевернутый и лофофор защищен крышечкой (крышкой), которая закрывается мышцами и открывается давлением жидкости. В одной учебный класс полая мочка, называемая «эпистомом», нависает над ртом.[8]
Кишечник имеет U-образную форму, идёт ото рта в центре лофофора вниз во внутреннюю часть животного, а затем обратно к нему. анус, который расположен на перевернутом, снаружи и обычно ниже лофофора.[8] Сеть нитей мезотелий называется "funiculi" ("веревочки"[24]) соединяет мезотелий, покрывающий кишечник, с выстилающей стенкой тела. Стенка каждой нити состоит из мезотелия и окружает пространство, заполненное жидкостью, которая считается кровью.[8] Зооиды колонии связаны, что позволяет автозооидам делиться пищей друг с другом и с любыми гетерозооидами, не питающимися.[8] Способы связи варьируются между различными классами мшанок: от довольно больших промежутков в стенках тела до небольших пор, через которые питательные вещества проходят через канатики.[8][20]
Есть нервное кольцо вокруг глотки (горла) и ганглий это служит мозгом для одной стороны этого. Нервы идут от кольца и ганглия к щупальцам и остальной части тела.[8] У мшанок нет специализированных органов чувств, но реснички на щупальцах действуют как сенсоры. Члены род Бугула расти к солнцу, и поэтому должен уметь обнаруживать свет.[8] В колониях некоторых видов сигналы передаются между зооидами через нервы, которые проходят через поры в стенках тела и координируют такие действия, как питание и втягивание лофофоров.[8]
Одинокие особи Монобриозоид автозооиды с грушевидным телом. Более широкие концы имеют до 15 коротких мускулистых выступов, с помощью которых животные прикрепляются к песку или гравию.[25] и протащатся сквозь отложения.[26]
Авикулярия и вибракула
Некоторые авторитеты используют термин авикулярия (пл. из авикулярий ) для обозначения любого типа зооида, в котором лофофор заменен расширением, выполняющим некоторую защитную функцию,[21] в то время как другие ограничивают этот термин теми, кто защищает колонию, хватая захватчиков и мелких хищников, убивая некоторых и кусая придатки других.[8] У некоторых видов щелкающие зооиды установлены на стебле (стебле), их птицеподобный вид является причиной появления термина - Чарльз Дарвин описал их как «голову и клюв стервятника в миниатюре, сидящего на шее и способного двигаться».[8][21] Стебельчатые авикулярии кладут на стебель в перевернутом виде.[18] «Нижние челюсти» представляют собой модифицированные версии жаберных крышек, которые защищают втянутые лофофоры у автозооидов некоторых видов и закрываются «как мышеловка» аналогичными мышцами.[8] а верхняя челюсть в форме клюва - перевернутая стенка тела.[18] У других видов авикулярии представляют собой неподвижные коробчатые зооиды, расположенные нормальным образом вверх, так что модифицированная жаберная крышка прижимается к стенке тела.[18] У обоих типов измененная жаберная крышка открывается другими прикрепленными к ней мышцами,[21] или внутренними мышцами, которые повышают давление жидкости, натягивая гибкую мембрану.[8] Действия этих щелкающих зооидов контролируются небольшими, сильно модифицированными полипами, которые расположены внутри «рта» и несут пучки коротких сенсорных реснички.[8][18] Эти зооиды появляются в различных положениях: некоторые занимают место автозооидов, некоторые помещаются в небольшие промежутки между автозооидами, а маленькие авикулярии могут встречаться на поверхностях других зооидов.[21]
У вибракулы, которую некоторые считают разновидностью авикулярий, жаберная крышка видоизменена, образуя длинную щетину с широким диапазоном движений. Они могут действовать как защита от хищников и захватчиков или как чистильщики. У некоторых видов, образующих мобильные колонии, вибракулы по краям используются в качестве ног для рытья и ходьбы.[8][21]
Другие виды колониальных зооидов
Кенозооиды (от греческого κενος, что означает «пустой»[27]) состоят только из стенки кузова и нитей фуникулера, пересекающих интерьер,[8] и никакого полипида.[18] У некоторых видов они образуют стебли ветвящихся структур, у других они действуют как спейсеры, позволяющие колониям быстро расти в новом направлении.[18][21]
Спинозооиды образуют защитные шипы и иногда появляются поверх автозооидов. Гонозооиды действуют как выводковые камеры для оплодотворенных яиц.[18] У некоторых видов есть миниатюрные нанозооиды с небольшими полипидами с одним щупальцем, и они могут расти на других зооидах или в стенках тела автозооидов, которые выродились.[21]
Формы и состав колоний
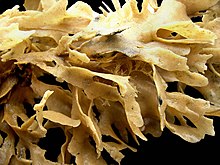

Хотя зооиды микроскопические, размер колоний варьируется от 1 см (1⁄2 дюймов) до более 1 м (3 футов 3 дюйма).[8] Однако большинство из них имеют диаметр менее 10 см (4 дюйма).[11] Формы колоний сильно различаются, в зависимости от модели бутонизации, по которой они растут, разнообразия присутствующих зооидов, а также типа и количества скелетного материала, который они выделять.[8]
Некоторые морские виды напоминают кусты или вееры, опираясь на «стволы» и «ветви», образованные кенозооидами, из которых растут питающиеся автозооиды. Колонии этих типов обычно неминерализованный но может иметь экзоскелеты сделано из хитин.[8] Другие выглядят маленькими кораллы, производя тяжелые каркасы извести.[28] Многие виды образуют колонии, состоящие из листов автозооидов. Эти листы могут образовывать листья, пучки или, в род Thalmoporella, структуры, напоминающие открытую кочан салата.[8]
Однако наиболее распространенной морской формой является корка, при которой однослойный лист зооидов покрывает твердую поверхность или водоросли. Некоторые колонии, покрытые коркой, могут вырастать до более 50 см (1 фут 8 дюймов) и содержать около 2 000 000 зооидов.[8] Эти виды обычно имеют экзоскелеты, усиленные карбонат кальция, а отверстия, через которые выступают лофофоры, находятся на верхней или внешней поверхности.[8] Подобный мху внешний вид покрывающих корку колоний отвечает за название филума (Древнегреческий слова βρύον Brúon что означает "мох" и ζῷον Zôion означает «животное»).[29] Большие колонии корковых видов часто имеют "дымоходы ", щели в покрове лофофоров, через которые они быстро вытесняют просеянную воду и, таким образом, избегают повторной фильтрации воды, которая уже исчерпана.[30] Они образованы пятнами непищевых гетерозооидов.[31] Новые дымоходы появляются у краев расширяющихся колоний, в точках, где скорость оттока уже высока, и не меняют своего положения при изменении потока воды.[32]
Некоторые пресноводные виды выделяют массу студенистого материала диаметром до 1 м (3 фута 3 дюйма), к которому прилипают зооиды. Другие пресноводные виды имеют форму растений с «стволами» и «ветвями», которые могут стоять прямо или раскидываться по поверхности. Некоторые виды могут ползать примерно на 2 см (3⁄4 в) в сутки.[8]
Каждая колония растет на бесполый подающий надежды от единственного зооида, известного как анцеструла,[8] который имеет скорее округлую форму, чем форму обычного зооида.[11] Это происходит на концах «стволов» или «ветвей» форм, имеющих такую структуру. По их краям вырастают корковые колонии. У видов с известковый экзоскелеты, они не минерализуются, пока зооиды не вырастут полностью. Продолжительность жизни колонии колеблется от одного до примерно 12 лет, и короткоживущие виды проходят через несколько поколений за один сезон.[8]
Виды, которые производят защитные зооиды, делают это только тогда, когда угрозы уже появились, и могут сделать это в течение 48 часов.[18] Теория «индуцированной защиты» предполагает, что производство средств защиты обходится дорого и что колонии, которые защищаются слишком рано или слишком сильно, будут иметь сниженные темпы роста и продолжительность жизни. Такой подход к защите «в последнюю минуту» возможен, потому что потеря зооидов в результате одной атаки вряд ли будет значительной.[18] Колонии некоторых покрывающих корку видов также производят специальные гетерозооиды, чтобы ограничить распространение других корковых организмов, особенно других мшанок. В некоторых случаях этот ответ более агрессивен, если противник меньше, что говорит о том, что зооиды на краю колонии могут каким-то образом ощущать размер противника. Некоторые виды постоянно преобладают над другими, но большинство дерновые войны нерешительны, и комбатанты вскоре начинают расти на неоспоримых территориях.[18] Мшанки, соревнующиеся за территорию, не используют изощренные методы, используемые губки или же кораллы возможно потому, что из-за короткой продолжительности жизни мшанок тяжелые вложения в войны за территорию становятся невыгодными.[18]
Мшанки вносят свой вклад в карбонатное осаждение морской жизни с ордовикского периода. Мшанки берут на себя ответственность за многие формы колоний, которые эволюционировали в разные таксономические группы и различаются по способности образовывать отложения. Девять основных форм колоний мшанок включают: корковые, куполообразные, пальчатые, листовые, фенестратные, сильное ветвление, нежное ветвление, сочлененные и свободно живущие. Большинство этих отложений происходит из двух отдельных групп колоний: домовых, с тонким ветвлением, с сильным ветвлением и пальчатыми; и фенестрат. Колонии фенестрата генерируют грубые частицы как в виде осадка, так и в качестве компонентов коралловых рифов строматопориодов. Однако хрупкие колонии образуют как грубые отложения, так и ядра глубоководных субфотных биогенных насыпей. Почти все постмшанчатые отложения состоят из форм роста, с добавлением свободноживущих колоний, которые включают значительное количество различных колоний. «В отличие от палеозоя, постпалеозойские мшанки образовывали отложения, которые более широко варьируются в зависимости от размера их зерен; они растут, когда переходят от грязи к песку и гравию ».[33]
Таксономия


Первоначально этот тип назывался «Polyzoa», но вскоре это название было заменено термином Эренберга «Bryozoa».[34][35] Название «Bryozoa» первоначально применялось только к животным, также известным как «Ectoprocta», у которых анус лежит за пределами «короны» щупалец (по Древнегреческий приставка ἐκτο, означающая «снаружи», и слово πρωκτος, означающее «анус»).[36] После открытия Энтопрокта, в котором анус находится внутри «короны» из щупалец (на основе древнегреческого префикса ἐντο, означающего «внутри», и слова πρωκτος, означающего «анус»[37]) название «Bryozoa» использовалось в филюм уровень, чтобы включить два классы Эктопрокта и Энтопрокта.[38] Однако в 1869 году Хинрих Ницше считал эти две группы совершенно разными по разным причинам и придумал название «Ectoprocta» для «Bryozoa» Эренберга.[4][39] Несмотря на внешне похожие методы кормления, они заметно различались анатомически; в дополнение к разному расположению ануса, эктопрокты имеют полые щупальца и целомудрие, а у энтопроков твердые щупальца и нет целома. Следовательно, эти две группы теперь широко рассматриваются как отдельные типы, и название «Bryozoa» теперь является синонимом «Ectoprocta».[38] С тех пор это мнение остается большинством, хотя большинство публикаций предпочитают название «Bryozoa», а не «Ectoprocta».[35] Тем не менее, некоторые известные ученые продолжали рассматривать "Ectoprocta" и "Entoprocta" как близких родственников и объединять их в группу "Bryozoa".[39]
Неопределенность в отношении объема названия «Bryozoa» привела к предложениям в 1960-х и 1970-х годах, что его следует избегать и использовать однозначный термин «Ectoprocta».[40] Однако это изменение затруднило бы поиск более старых работ, в которых тип был назван «Bryozoa», а стремление избежать двусмысленности, если бы оно применялось последовательно ко всем классификациям, потребовало бы переименования нескольких других типов и многих более низких. группы уровней.[34] На практике зоологические наименования разделенных или объединенных групп животных сложны и не вполне последовательны.[41] В произведениях с 2000 года для устранения двусмысленности использовались различные названия, в том числе: «Bryozoa»,[8][11] «Эктопрокта»,[14][18] "Bryozoa (Ectoprocta)",[20] и «Ectoprocta (Bryozoa)».[42] Некоторые использовали более одного подхода в одной работе.[43]
Общее название «моховые животные» происходит от греческих βρυόν (мох) и ζῷα (животные) и относится к покрытым мхом видам покрывающих корку видов.[44]
Вплоть до недавнего времени (2008 г.) существовали «недостаточно известные и неправильно понятые типовые виды, принадлежащие к семейству Cyclostome Bryozoan Oncousoeciidae». (Taylor, Zaton 2008). Были проведены современные исследования и эксперименты с использованием сканирующей электронной микроскопии в низком вакууме материалов без покрытия, чтобы критически изучить и, возможно, пересмотреть таксономию трех родов, принадлежащих к этому семейству, включая Онкусоэция, Microeciella, и Эвристрот. Этот метод позволяет получать данные, которые трудно распознать в оптический микроскоп. Допустимый типовой вид Онкусоэция оказался Oncousoecia lobulata. Эта интерпретация стабилизирует Онкусоэция путем установления типового вида, соответствующего общему использованию этого рода. Товарищ Онкусоэциид Эвристрот теперь считается, что не является сородичем О. лобулата, как предполагалось ранее, но показывает достаточно сходств, чтобы считаться младшим синонимом Онкусоэция. Microeciella suborbicularus также недавно был выделен из О. лобулата и О. дилатанс, используя этот современный метод сканирования в низком вакууме, которому он был неточно синонимизирован в прошлом. Недавно был открыт новый род, названный Юнероссия в семействе Stomachetosellidae, наряду с 10 относительно новыми видами мшанок, такими как Альдерина флавента, Corbulella extenuata, Puellina septemcryptica, Юнероссия копиоза, Калиптотека капааенсис, Bryopesanser зубчатая, Cribellopora souleorum, Metacleidochasma verrucosa, Disporella compta, и Favosipora adunca.[45]
Классификация и разнообразие
Количество официально описанных видов колеблется от 4000 до 4500.[46] Gymnolaemata и особенно Cheilostomata имеют наибольшее количество видов, возможно, из-за широкого круга специализированных зооидов.[18] Под Линнеевская система классификации, который до сих пор используется как удобный способ пометить группы организмов,[47] живущие члены филюм Мшанки делятся на:[8][18]
| Учебный класс | Phylactolaemata | Stenolaemata | Gymnolaemata | |
|---|---|---|---|---|
| Заказ | Plumatellida[48] | Циклостоматида | Ктеностомы | Хейлостомы |
| Среды | Пресная вода | морской | В основном морской | |
| Губообразный эпистом, нависающий над ртом | да | никто | ||
| Формы колоний | Желатиновые массы или трубчатые разветвленные структуры[49] | Возводить или покрывать коркой[50] | Прямой, инкрустирующий или свободно живущий | |
| Материал экзоскелета | Студенистый или пленчатый; неминерализованный | Минерализованный | Хитин студенистые или перепончатые; неминерализованный | Минерализованный |
| Operculum («крышка») | никто | никто[50] (кроме семьи Eleidae[51]) | Нет у большинства видов | Да (кроме рода Бугула ) |
| Форма лофофора | U-образный вид (кроме род Фредеричелла, у которого лофофор округлый) | Круговой | ||
| Как распространился лофофор | Сжатие всей стенки тела | Сдавливание перепончатого мешка (отдельный внутренний слой эпителий что выстилает целом) | Сжатие всей стенки тела | Втягивание внутрь гибкого участка стенки тела или расширение внутреннего мешка. |
| Виды зооидов | Только автозооиды | Ограниченные гетерозооиды, в основном гонозооиды[52] | Столоны и шипы, а также автозооиды[52] | Полный спектр типов |
Окаменелости

| ||
| ||
| ||
| ||
| ||
| Ископаемый скелет Архимед Мшанник |

Найдены окаменелости около 15 000 видов мшанок. Мшанки - одна из трех доминирующих групп палеозойских окаменелостей.[53] Самый древний вид с минерализованный скелет встречается в нижнем ордовике.[1] Вполне вероятно, что первые мшанки появились намного раньше и были полностью мягкотелыми, а окаменелости ордовика фиксируют появление минерализованных скелетов в этом типе.[4] Аренигским этапом раннего Ордовик период,[11][54] о 480 миллион лет назад, все современные заказы из стенолематы были представлены,[55] и ктеностома порядок гимнасты появился к среднему ордовику, около 465 миллион лет назад. Окаменелости раннего ордовика могут также представлять формы, которые уже стали значительно отличаться от первоначальных представителей этого типа.[55] Ктеностомы с фосфатированной мягкой тканью известны с девона.[56] Другие виды питатели-фильтры появились примерно в то же время, что говорит о том, что некоторые изменения сделали среду более благоприятной для этого образа жизни.[11] Окаменелости хейлостомы, еще один отряд гимназистов, впервые появляется в середине Юрский, о 172 миллион лет назад, и это были самые многочисленные и разнообразные мшанки из Меловой в настоящее время.[11] Данные, собранные за последние 100 миллионов лет, показывают, что хейлостомы постоянно росли над циклостоматами в территориальной борьбе, что может помочь объяснить, как хейлостомы вытеснили циклостоматов в качестве доминирующих морских мшанок.[57] Морские окаменелости из Палеозой эпоха, которая закончилась 251 миллион лет назад, имеют в основном прямостоячие формы, из Мезозойский довольно поровну разделены на прямостоячие и корковые формы, а более поздние - преимущественно корковые.[58] Окаменелости мягкой пресной воды филактолематы очень редки,[11] появляются в и после поздней перми (которая началась около 260 миллион лет назад) и полностью состоят из прочных статобластов.[49] Нет никаких известных окаменелостей пресноводных представителей других классов.[49]
Эволюционное генеалогическое древо

Ученые разделились во мнениях относительно того, являются ли Bryozoa (Ectoprocta) монофилетический группы (включают ли они все и только одного вида-предка и всех его потомков), о том, каковы ближайшие родственники этого типа в генеалогическом древе животных, и даже о том, следует ли считать их членами протостомы или же дейтеростомы, две основные группы, которые составляют всех животных средней сложности.
Молекулярная филогения, которая пытается разработать эволюционное генеалогическое древо организмов, сравнивая их биохимия и особенно их гены, много сделал для прояснения отношений между наиболее известными беспозвоночный тип.[38] Однако нехватка генетических данных о "второстепенных типах", таких как мшанки и энтопрокты оставил свои отношения с другими группами неясными.[39]
Традиционный вид
Традиционная точка зрения состоит в том, что Bryozoa представляют собой монофилетическую группу, в которой учебный класс Phylactolaemata наиболее тесно связан с Stenolaemata и Ктеностомы, классы, которые появляются раньше всего в летописи окаменелостей.[59] Однако в 2005 г. молекулярная филогения исследование, посвященное филактолематам, пришло к выводу, что они более тесно связаны с типом Phoronida, и особенно к единственному виду форонид, который является колониальным, чем к другим классам эктопроков. Это означает, что Entoprocta не являются монофилетическими, поскольку Phoronida являются подгруппой эктопроктов, но стандартное определение Entoprocta исключает Phoronida.[59]

В 2009 году еще один молекулярная филогения исследования, используя комбинацию генов из митохондрии и ядро клетки пришли к выводу, что мшанки - это монофилетический phylum, другими словами, включает всех потомков общего предка, который сам является мшанкой. Анализ также пришел к выводу, что классы Phylactolaemata, Stenolaemata и Gymnolaemata также являются монофилетическими, но не удалось определить, Stenolaemata более тесно связаны с Phylactolaemata или Gymnolaemata. Gymnolaemata традиционно делятся на мягкотелых. Ктеностомы и минерализованный Cheilostomata, но анализ 2009 г. посчитал более вероятным, что ни один из этих заказы является монофилетическим и минерализованным скелеты вероятно, не раз эволюционировал в пределах ранних Gymnolaemata.[4]
Отношения мшанок с другими типами сомнительны и противоречивы. Традиционная филогения, основанная на анатомия и по развитию взрослых форм от эмбрионы, не привело к устойчивому консенсусу по поводу положения эктопродуктов.[14] Попытки восстановить генеалогическое древо животных в значительной степени игнорировали эктопрокты и другие «второстепенные типы», которые мало изучены научными исследованиями, поскольку они, как правило, крошечные, имеют относительно простой строение тела и мало влияют на человеческую экономику - несмотря на то, "второстепенные типы" включают в себя большую часть всего разнообразия в истории эволюции животных.[61]
По мнению Рут Дьюэл, Джудит Уинстон и Фрэнка МакКинни "Наша стандартная интерпретация мшанок морфология и эмбриология представляет собой конструкцию, возникшую в результате более чем 100-летних попыток синтезировать единую структуру для всех беспозвоночных », и мало учитывает некоторые особенности эктопроктов.[55]


У эктопрокта все внутренние органы личинки разрушаются во время метаморфоза до взрослой формы, и органы взрослой особи строятся из личинки. эпидермис и мезодерма, а в других билатерии некоторые органы, включая кишечник, построены из энтодерма. У большинства двусторонних эмбрионов бластопор, вмятина на внешней стенке, углубляется и становится кишечником личинки, но при эктопрокте бластопор исчезает, и новая вмятина становится точкой роста кишечника. Целом эктопрокта не образуется ни одним из процессов, используемых другими билатериянами, энтерокоэли, в которых мешочки, образующиеся на стенке кишечника, становятся отдельными полостями, ни шизокоэлия, при котором ткань между кишечником и стенкой тела расщепляется, образуя парные полости.[55]
Энтопрокты
Когда в XIX веке были открыты энтопрокты, они и мшанки (эктопрокты) рассматривались как классы внутри типа Bryozoa, поскольку обе группы были сидячий животные, которые с фильтром посредством венца из щупалец, несущих реснички.
Начиная с 1869 г., растет понимание различий, в том числе положения энтопрокта. анус внутри структуры кормления и разница в раннем схема разделения ячеек в их эмбрионы, заставило ученых рассматривать две группы как отдельные типы,[39] а «мшанки» стали просто альтернативным названием эктопроктов, у которых анус находится за пределами питающего органа.[38] Серия исследований молекулярной филогении с 1996 по 2006 год также пришла к выводу, что мшанки (эктопрокты) и энтопрокты не являются сестринскими группами.[39]
Однако два известных зоолога Клаус Нильсен и Томас Кавалье-Смит, утверждают на основании анатомических особенностей и особенностей развития, что мшанки и энтопрокты принадлежат к одному и тому же типу - Bryozoa. Исследование молекулярной филогении в 2007 году также поддержало эту старую идею, в то время как его выводы о других типах совпали с выводами нескольких других анализов.[39]
Группировка в лофофората
К 1891 г. мшанки (эктопрокты) были отнесены к форониды в суперфиле под названием «Тентакулата». В 1970-х годах сравнения личинок форонид и Cyphonautes Личинки некоторых гимнолатных мшанок предположили, что мшанки, большинство из которых являются колониальными, произошли от полуколониальных видов форонидов.[62] Брахиоподы также были отнесены к «Tentaculata», которые были переименованы Лофофората поскольку все они используют лофофор для подачи фильтра.[38]
Большинство ученых с этим согласны,[38] но Клаус Нильсен считает эти сходства поверхностными.[14] Лофофората обычно определяется как животные с лофофором, трехчастным целомом и U-образной кишкой.[62] По мнению Нильсена, лофофоры форонид и брахиопод больше похожи на лофофоры крылышки,[14] которые являются членами филума Hemichordata.[63] Щупальца мшанок несут клетки с множеством реснички, в то время как соответствующие клетки лофофоров форонид, брахиопод и птеробранхов имеют по одной ресничке на клетку; а щупальца мшанок не имеют гемального канала («кровеносного сосуда»), который есть у других трех типов.[14]
Если группировка мшанок с форонидами и брахиопод в Lophophorata верна, то следующий вопрос заключается в том, являются ли Lophophorata протостомы, наряду с большинством типов беспозвоночных, или дейтеростомы, вместе с хордовые, полухордовые и иглокожие.
Традиционно считалось, что лофофораты представляют собой смесь протостомов и дейтеростомов. Исследования 1970-х годов показали, что это дейтеростомы из-за некоторых особенностей, которые считались характерными для дейтеростомов: трехчастный целом; радиальное, а не спиральное дробление в развитии эмбриона;[38] и формирование целома энтерокоэли.[14] Однако целом у личинок эктопрока не видно деления на три части.[62] и у взрослых эктопроктов отличается от других целомат phyla, поскольку он построен заново из эпидермиса и мезодермы после того, как метаморфоз разрушил личинку целом.[55]
Молекулярная филогенетика лофофоратов
Молекулярный филогенетический анализ, начиная с 1995 г., с использованием разнообразных биохимических данных и аналитических методов показал, что лофофораты являются протостомами и тесно связаны с кольчатые червя и моллюски в суперфиле под названием Лофотрохозоа.[38][64] Анализ «полных доказательств», в котором использовались как морфологические признаки, так и относительно небольшой набор генов, позволил прийти к различным выводам, в основном в пользу тесной связи между лофофоратами и Lophotrochozoa.[64] Исследование 2008 года с использованием большего набора генов пришло к выводу, что лофофораты были ближе к Lophotrochozoa, чем к дейтеростомам, но также что лофофораты не были монофилетическими. Вместо этого он пришел к выводу, что брахиоподы и форониды образуют монофилетическую группу, но мшанки (эктопрокты) были наиболее близки к энтопрокам, что подтверждает первоначальное определение «мшанок».[64]
Они также являются единственным основным типом исключительно клональных животных и все являются колониальными. Это колонии модульных единиц, известных как зооиды. Поскольку они процветают в колониях, колониальный рост позволяет им развивать неограниченные вариации формы. Несмотря на это, было обнаружено лишь небольшое количество основных форм роста, которые обычно вновь появлялись на протяжении всей истории мшанок.[53]
Эктопрокт молекулярная филогенетика
Филогенетическое положение ectoproct bryozoans остается неопределенным, но остается уверенным, что они принадлежат к Protostomia, а точнее к Lophoctrochozoa. Это означает, что личинка эктопрока является трохофорой, а корона - гомологом прототроха; это подтверждается сходством между венечными личинками и личинками перикалиммы Типа 1 некоторых моллюсков и сипункул, где зона прототроха расширяется, чтобы покрыть гипосферу.[65]
Изучение последовательности митохондриальной ДНК предполагает, что Bryozoa могут быть связаны с Chaetognatha.[66]
Физиология
Кормление и выделение
Большинство видов питатели-фильтры которые просеивают мелкие частицы, в основном фитопланктон (микроскопические плавающие растения) из воды.[8] Пресноводные виды Plumatella emarginata питается диатомеи, зеленые водоросли, цианобактерии, не-фотосинтетический бактерии динофлагелляты, коловратки, простейшие, маленький нематоды и микроскопические ракообразные.[67] Хотя токи, которые генерируют мшанки для притягивания пищи ко рту, хорошо известны, точный метод захвата все еще обсуждается. Все виды также смахивают щупальцами более крупные частицы ко рту, а некоторые захватывают их. зоопланктон (планктонные животные), используя свои щупальца в качестве клеток. Кроме того, щупальца, площадь поверхности которых увеличена на микроворсинки (небольшие волоски и складки), впитывают органические соединения растворяется в воде.[8] Нежелательные частицы можно отогнать щупальцами или закрыть, закрыв рот.[8] Исследование, проведенное в 2008 году, показало, что как покрытые коркой, так и стоячие колонии питались быстрее и росли быстрее при слабом, чем при сильном течении.[68]
У некоторых видов первая часть желудка образует мышечный желудок выложено хитиновый зубы, которые раздавливают бронированную добычу, например диатомеи. Волнообразный перистальтический сокращения перемещают пищу через желудок для переваривания. Последняя часть желудка выстлана реснички (мельчайшие волоски), которые сжимают непереваренные твердые частицы, которые затем проходят через кишечник и через анус.[8]
Нет нефридия («маленькие почки») или другие выделительный органы у мшанок,[18] и считается, что аммиак распространяется через стенку тела и лофофор.[8] Более сложные отходы не выводятся, а накапливаются в полипид, который вырождается через несколько недель. Часть старого полипида перерабатывается, но большая его часть остается в виде большой массы умирающих клеток, содержащих накопившиеся отходы, которые спрессовываются в «коричневое тело». Когда дегенерация завершена, цистида (внешняя часть животного) производит новый полипид, а коричневое тело остается в целомудрие, или в желудке нового полипида, и в следующий раз, когда животное испражняется.[8]
Дыхание и кровообращение
Органов дыхания нет, сердце или же кровеносный сосуд. Вместо этого зооиды поглощают кислород и выводят углекислый газ путем диффузии. Мшанки осуществляют диффузию с помощью тонкой мембраны (в случае анасканы и некоторые polyzoa) или через псудопоры, расположенные на внешней дерме зооида.[69] Различные группы мшанок используют разные методы для обмена питательными веществами и кислородом между зооидами: у некоторых есть довольно большие промежутки в стенках тела, что позволяет целомическая жидкость свободно циркулировать; в других - фуникулеры (внутренние "веревочки")[24]) соседних зооидов соединяются через небольшие поры в стенке тела.[8][20]
Размножение и жизненные циклы

Зооиды всех видов филактолемат одновременны. гермафродиты. Хотя представители многих морских видов являются протандрическими, другими словами, функционируют сначала как самцы, а затем как самки, их колонии содержат комбинацию зооидов, находящихся на мужской и женской стадиях. У всех видов яичники развиваются на внутренней стороне стенки тела, и яички на семяпроводе, соединяющем желудок со стенкой тела.[18] Яйца и сперма выделяются в целом, и сперма выходит в воду через поры на кончиках некоторых щупалец, а затем захватывается питательными потоками зооидов, производящих яйца.[8] Яйца некоторых видов оплодотворяются извне после выхода через пору между двумя щупальцами, которая в некоторых случаях находится на кончике небольшого выступа, называемого «интертентакулярный орган», в основании пары щупалец. Другие оплодотворяются внутренне, в межжелудочковом органе или целом.[8] В ктеностомах мать обеспечивает выводковую камеру для оплодотворенных яиц, а ее полипид распадается, обеспечивая питание для потомства. эмбрион. Стенолематы производят специализированные зооиды, служащие помещениями для расплода, и их яйца делятся внутри них, давая до 100 идентичных эмбрионов.[18]
В расщепление яиц мшанок двулучевая, т. е. ранние стадии двусторонне симметричны. Неизвестно, как образуется колеом, так как метаморфоза из личинка взрослому человеку разрушает все внутренние ткани личинки. У многих животных бластопор, отверстие в поверхности раннего эмбриона, проходит сквозь него, образуя кишечник. Однако у мшанок бластопор закрывается, и появляется новое отверстие, образующее рот.[8]
Личинки мшанок различаются по форме, но все они имеют полосу ресничек вокруг тела, которая позволяет им плавать, пучок ресничек наверху и липкий мешок, который выворачивает и фиксирует их, когда они оседают на поверхности.[8] Некоторые виды гимнолемат производят личинок cyphonautes, у которых мало желтка, но хорошо развиты рот и кишечник, и они живут как планктон в течение значительного времени до заселения. Эти личинки имеют треугольные оболочки из хитина с одним углом вверху и открытым основанием, образуя капюшон вокруг рта, обращенного вниз.[18] В 2006 году сообщалось, что реснички личинок cyphonautes используют тот же набор методов, что и взрослые особи, для захвата пищи.[70] Виды, вынашивающие свои эмбрионы, образуют личинки, которые питаются желтки, не имеют кишечника и не питаются, а такие личинки быстро оседают на поверхности.[8] У всех морских видов личинки производят коконы, в которых они метаморфоза полностью после заселения: личинки эпидермис становится подкладкой целомудрие, а внутренние ткани превращаются в запас пищи, который питает развивающийся зооид, пока он не будет готов к кормлению.[8] Личинки филактолематов продуцируют несколько полипидов, так что каждая новая колония начинается с нескольких зооидов.[8] У всех видов зооиды-основатели затем выращивают новые колонии подающий надежды клоны самих себя. У филактолематов зооиды умирают после создания нескольких клонов, так что живые зооиды встречаются только по краям колонии.[8]
Филактолематы также могут размножаться бесполым путем, используя метод, который позволяет колонии выжить в изменчивых и неопределенных условиях пресноводной среды.[18] В течение лета и осени они производят дискообразные статобласты, массы клеток, которые функционируют как «стручки выживания», скорее похожие на геммулы губок.[8] Статобласты формируются на семенных канатиках, связанных с кишечником родителей, которые питают их.[18] По мере роста статобласты развивают защитные свойства. двустворчатый снаряды из хитин. Когда они созревают, некоторые статобласты прилипают к родительской колонии, некоторые падают на дно («сессобласты»), некоторые содержат воздушные пространства, которые позволяют им плавать («флотобласты»),[8] а некоторые остаются в цистиде родителя, чтобы восстановить колонию в случае ее смерти.[18] Статобласты могут оставаться в спящем состоянии в течение значительных периодов времени, а в спящем состоянии могут выжить в суровых условиях, таких как замораживание и высыхание. Их можно переносить на большие расстояния животными, плавучей растительностью, течениями.[8] и ветры,[18] и даже в кишечнике более крупных животных.[71] Когда условия улучшаются, створки раковины отделяются, и клетки внутри превращаются в зооид, который пытается сформировать новую колонию. Plumatella emarginata производит как «сессобласты», которые позволяют клону контролировать хорошую территорию, даже если в тяжелые времена уничтожаются родительские колонии, так и «флотобласты», которые распространяются на новые участки. Новые колонии Plumatella repens производят в основном «сессобласты», в то время как зрелые переходят в «флотобласты».[67] По оценкам исследования, одна группа колоний на участке размером 1 квадратный метр (11 квадратных футов) производила 800 000 статобластов.[8]
Cupuladriid Bryozoa способны к половому и бесполому размножению. Колонии, размножающиеся половым путем (аклональные), являются результатом роста личиночных купуладриидов до взрослой стадии, тогда как бесполые колонии (клональные) являются результатом роста фрагмента колонии купуладриидов в свою собственную колонию. Различные формы воспроизводства купуладриид достигаются с помощью различных методов в зависимости от морфологии и классификации зооида.[72]
Экология
Среда обитания и распространение
Большинство морских видов обитает в тропических водах на глубине менее 100 метров (330 футов; 55 морских саженей). Однако некоторые были найдены в глубоком море. траншеи,[73] особенно вокруг холодные просачивания, и другие рядом с полюса.[74][75] Подавляющее большинство из них сидячий. Формы корки являются наиболее распространенными из них на мелководье, но прямостоячие формы становятся более распространенными по мере увеличения глубины.[74] Несколько форм, таких как Кристелла может двигаться, и Антарктика разновидность, Alcyonidium pelagosphaera, состоит из плавающих колоний. Пелагический вид имеет диаметр от 5,0 до 23,0 мм, имеет форму полой сферы и состоит из одного слоя автозооидов. До сих пор неизвестно, являются ли эти колонии пелагическими всю свою жизнь или представляют собой временно и ранее не описанную стадию молоди.[74][76]
В 2014 г. сообщалось, что мшанки Fenestrulina rugula стали доминирующим видом в некоторых частях Антарктиды. Глобальное потепление увеличил скорость очистки на айсберги, и этот вид особенно хорошо приспособлен к повторному заселению вымытых территорий.[77]
Филактолематы обитают во всех типах пресноводных сред - озерах и прудах, реках и ручьях, а также в эстуариях.[49] - и являются одними из самых многочисленных сидячих пресноводных животных.[59] Некоторые ктеностомы исключительно пресноводные, другие предпочитают солоноватую воду, но могут выжить в пресной воде.[49] Знания ученых о популяциях пресноводных мшанок во многих частях мира неполны, даже в некоторых частях Европы. Долгое время считалось, что некоторые пресноводные виды встречаются во всем мире, но с 2002 года все они были разделены на более локализованные виды.[49]
Мшанки в основном неподвижны, обычно обитают на твердых субстратах, включая камни, песок или ракушки. [78]
Мшанки растут в клональный колонии. Личинка мшанок поселяется на твердом веществе и производит колонию бесполым путем через почкование. Эти колонии могут вырастить тысячи отдельных зооидов за относительно короткий период времени. Несмотря на то, что колонии зооидов растут за счет бесполого размножения, мшанки являются гермафродитами, и новые колонии могут образовываться в результате полового размножения и образования свободно плавающих личинок. Однако, когда колонии становятся слишком большими, они могут разделиться на две части. Это единственный случай, когда в результате бесполого размножения новая колония отделяется от своей предшественницы. Большинство колоний стационарны. Действительно, эти колонии обычно оседают на неподвижных веществах, таких как отложения и грубые вещества. Есть несколько колоний пресноводных видов, таких как Кристателла муседо которые способны медленно передвигаться на ползучей ноге.[79]
Взаимодействие с нечеловеческими организмами

Морские виды распространены на коралловые рифы, но редко значительная часть общего биомасса. В умеренных водах скелеты погибших колоний составляют значительный компонент раковины. гравий, и живые в этих местах в изобилии.[80] Морская кружевная мшанка Membraniporambranacea производит шипы в ответ на нападение нескольких видов голожаберники («морские слизни»).[81] Другие хищники морских мшанок включают рыбу, морские ежи, пикногониды, ракообразные, клещи[82] и морская звезда.[83] В целом морской иглокожие и моллюски поедают массы зооидов, раздробив части колоний, ломая их минерализованные «домики», в то время как большинство членистоногие хищники на мшанках поедают отдельных зооидов.[84]
В пресной воде мшанки - одни из самых важных питатели-фильтры, вместе с губки и моллюски.[85] Пресноводные мшанки подвергаются нападению многих хищников, в том числе улиток, насекомых и рыб.[67]
В Таиланд то интродуцированные виды Pomacea canaliculata (золотая яблочная улитка), которая, как правило, разрушительна травоядное животное, уничтожил популяции филактолематов везде, где бы они ни появлялись. P. canaliculata также охотится на обыкновенного пресноводного гимнолемата, но с меньшим разрушительным эффектом. Аборигенные улитки мшанками не питаются.[86]
Несколько видов гидроид семья Zancleidae имеют симбиотические отношения с мшанками, некоторые из которых полезны для гидроидов, а другие - паразитический. Изменения проявляются в форме некоторых этих гидроидов, например, более мелкие щупальца или инкрустация корней мшанками.[87] Мшанки Альционидиум узловатый защищает ублюдок Burnupena papyracea против хищничества сильных и ненасытных лангуст Jasus lalandii. Хотя раковины моллюсков, покрытые оболочкой мшанок, более прочны, чем раковины без этого укрепления, химическая защита, производимая мшанками, вероятно, является более значительным сдерживающим фактором.[88]

в Banc d'Arguin офшорный Мавритания виды Комменсальная акантодезия, который обычно растет прикрепленным к гравию и твердому субстрату, сформировал необязательные симбиотические отношения с раки-отшельники вида Pseudopagurus cf. гранулиман в результате образуются структуры размером с яйцо, известные как бриолиты.[89] Зародившись на пустой раковине брюхоногих моллюсков, колонии мшанок образуют многослойные скелетные корки, которые образуют сферические инкрустации и расширяют жизненную камеру рака-отшельника за счет спирально-спирального трубчатого роста.
Некоторые виды филактолематов являются промежуточными хозяевами для группы миксозоа которые также вызывают пролиферативная болезнь почек, что часто приводит к летальному исходу лосось рыбы,[90] и резко сократила популяции диких рыб в Европе и Северной Америке.[49]
Membraniporambranacea, чьи колонии питаются и растут исключительно быстро в широком диапазоне скоростей течения, был впервые замечен в Залив Мэн в 1987 году и быстро стал самым многочисленным организмом, живущим на водоросли.[68] Этот вторжение сократили популяцию ламинарии, сломав их листья,[8] так что его место в качестве доминирующей «растительности» в некоторых областях занял другой захватчик, крупный водоросль Кодиум хрупкий tomentosoides.[68] Эти изменения уменьшили площадь среды обитания, доступную для местных рыб и беспозвоночных. М. мембранацеа также вторгся на северо-западное побережье США.[8] Несколько пресноводных видов также были обнаружены за тысячи километров от их естественных ареалов. Некоторые из них могли транспортироваться естественным путем в виде статобластов. Другие, более вероятно, были распространены людьми, например, через завезенные водные растения или в качестве безбилетных пассажиров на кораблях.[71]
Взаимодействие с людьми
Рыбные хозяйства и инкубаторы потеряли запасы пролиферативная болезнь почек, что вызвано одним или несколькими миксозои которые используют мшанок в качестве альтернативных хозяев.[90]
Некоторые рыбаки в Северное море пришлось искать другую работу из-за формы экзема (кожное заболевание), известное как "Доггер Банк зуд ",[74] вызвано контактом с мшанками, приставшими к сетям и кастрюлям для омаров.[91]
Морские мшанки часто ответственны за биообрастание на корпусах судов, в доках и маринах, а также на морских сооружениях. Они одни из первых колонизаторов новых или недавно очищенных построек.[80] Пресноводные виды - случайные помехи в водопроводных трубах, оборудовании для очистки питьевой воды, очистных сооружениях и охлаждающих трубах электростанций.[49][92]
Группа химикатов под названием бриостатины можно извлечь из морских мшанок Бугула неритина. В 2001 году фармацевтическая компания GPC Biotech лицензированный бриостатин 1 от Государственный университет Аризоны для коммерческого развития в качестве лечения рака. GPC Biotech отменила разработку в 2003 году, заявив, что бриостатин 1 показал небольшую эффективность и некоторые токсические побочные эффекты.[93] В январе 2008 г. клиническое испытание был отправлен в США Национальные институты здоровья для измерения безопасности и эффективности Бриостатина 1 при лечении Болезнь Альцгеймера. Однако к концу декабря 2008 года, когда планировалось завершить исследование, участников набрать не было.[94] Более поздняя работа показывает, что он положительно влияет на познавательные способности у больных Болезнь Альцгеймера с небольшими побочными эффектами.[95] Около 1000 кг (2200 фунтов) мшанок необходимо переработать для получения 1 грамма (1⁄32 унция) бриостатина. В результате были разработаны синтетические эквиваленты, которые проще производить и, по-видимому, не менее эффективны.[96]
Смотрите также
Рекомендации
- ^ а б c Taylor, P.D .; Бернинг, В .; Уилсон, М.А. (ноябрь 2013 г.). «Переосмысление кембрийского« мшанки » Pywackia как октокораль ". Журнал палеонтологии. 87 (6): 984–990. Дои:10.1666/13-029. S2CID 129113026.
- ^ Ма, Джунье; Тейлор, Пол Д .; Ся, Фэншэн; Жан, Ренбин (сентябрь 2015 г.). «Самый старый из известных мшанок: Профилактика (Cryptostomata) из нижнего тремадока (нижний ордовик) Люцзячан, юго-запад провинции Хубэй, центральный Китай ". Палеонтология. 58 (5): 925–934. Дои:10.1111 / pala.12189.
- ^ Эрнст, А. (2007). «Цистопоральный вид мшанок из Цехштейна (поздняя пермь)». Paläontologische Zeitschrift. 81 (2): 113–117. Дои:10.1007 / BF02988385. S2CID 129637643.
- ^ а б c d Fuchs, J .; Обст, М; Сундберг, П. (июль 2009 г.). «Первая всеобъемлющая молекулярная филогения Bryozoa (Ectoprocta), основанная на комбинированном анализе ядерных и митохондриальных генов». Молекулярная филогенетика и эволюция. 52 (1): 225–233. Дои:10.1016 / j.ympev.2009.01.021. PMID 19475710.
- ^ Бруска; Бруска. "21: Тип лофофората". Беспозвоночные.
- ^ Bock, P .; Гордон, Д. (Август 2013). "Тип Bryozoa Ehrenberg, 1831". Zootaxa. 3703 (1): 67–74. Дои:10.11646 / zootaxa.3703.1.14.
- ^ «Знакомство с мшанками». Калифорнийский университет Беркли. Получено 8 декабря 2019.
- ^ а б c d е ж грамм час я j k л м п о п q р s т ты v ш Икс у z аа ab ac объявление ае аф аг ах ай эй ак аль являюсь ан ао ap водный ар в качестве в au средний ау топор ай az ба bb до н.э bd быть парень bg бх Ruppert, E.E .; Фокс, Р. И Барнс, Р.Д. (2004). «Лофопората». Зоология беспозвоночных (7-е изд.). Брукс / Коул. стр.829–845. ISBN 978-0-03-025982-1.
- ^ а б c Ruppert, E.E .; Фокс, Р. И Барнс, Р.Д. (2004). «Лофопората». Зоология беспозвоночных (7-е изд.). Брукс / Коул. стр.817–821. ISBN 978-0-03-025982-1.
- ^ а б c Ruppert, E.E .; Фокс, Р. И Барнс, Р.Д. (2004). «Лофопората». Зоология беспозвоночных (7-е изд.). Брукс / Коул. стр.821–829. ISBN 978-0-03-025982-1.
- ^ а б c d е ж грамм час я j Rich, T.H .; Fenton, M.A .; Фентон, К. (1997). ""Моховые животные ", или мшанки". Книга окаменелостей. Dover Publications. стр.142–152. ISBN 978-0-486-29371-4. Получено 7 августа 2009.
- ^ а б Ruppert, E.E .; Фокс, Р. И Барнс, Р.Д. (2004). «Камптозоа и циклиофора». Зоология беспозвоночных (7-е изд.). Брукс / Коул. стр.808–812. ISBN 978-0-03-025982-1.
- ^ Ruppert, E.E .; Фокс, Р. И Барнс, Р.Д. (2004). «Лофопората». Зоология беспозвоночных (7-е изд.). Брукс / Коул. п.817. ISBN 978-0-03-025982-1.
- ^ а б c d е ж грамм час я Нильсен, К. (июль 2002 г.). «Филогенетическое положение Entoprocta, Ectoprocta, Phoronida и Brachiopoda». Интегративная и сравнительная биология. 42 (3): 685–691. Дои:10.1093 / icb / 42.3.685. PMID 21708765.
- ^ Ruppert, E.E .; Фокс, Р. И Барнс, Р.Д. (2004). «Аннелида». Зоология беспозвоночных (7-е изд.). Брукс / Коул. стр.414–420. ISBN 978-0-03-025982-1.
- ^ Ruppert, E.E .; Фокс, Р. И Барнс, Р.Д. (2004). Зоология беспозвоночных (7-е изд.). Брукс / Коул. стр.284–291. ISBN 978-0-03-025982-1.
- ^ Гьер, О. (2009). «Тентакулата». Мейобентология (2-е изд.). Springer Verlag. п. 227. ISBN 978-3-540-68657-6. Получено 7 июля 2009.
- ^ а б c d е ж грамм час я j k л м п о п q р s т ты v ш Икс у z Доэрти, П.Дж. (2001). «Лофофораты». В Андерсоне, Д.Т. (ред.). Зоология беспозвоночных (2-е изд.). Издательство Оксфордского университета. С. 363–373. ISBN 978-0-19-551368-4.
- ^ Little, W .; Фаулер, H.W; Коулсон, Дж. И Луки, К. (1964). «Зооид». Краткий оксфордский словарь английского языка. Издательство Оксфордского университета. ISBN 978-0-19-860613-0.
- ^ а б c d е Нильсен, К. (2001). «Bryozoa (Ectoprocta: 'Moss' Animals)». Энциклопедия наук о жизни. John Wiley & Sons, Ltd. Дои:10.1038 / npg.els.0001613. ISBN 978-0470016176.
- ^ а б c d е ж грамм час McKinney, F.K .; Джексон, J.B.C. (1991). «Мшанки как модульные машины». Эволюция мшанок. Издательство Чикагского университета. С. 1–13. ISBN 978-0-226-56047-2. Получено 29 июля 2009.
- ^ Ruppert, E.E .; Фокс, Р. И Барнс, Р.Д. (2004). «Лофопората». Зоология беспозвоночных (7-е изд.). Брукс / Коул. п.817. ISBN 978-0-03-025982-1.
- ^ Riisgård, H.U .; Нильсен, К; Ларсен, PS (2000). «Сбор нисходящего потока в питателях цилиарной взвеси: принцип наверстывания» (PDF). Серия "Прогресс морской экологии". 207: 33–51. Bibcode:2000MEPS..207 ... 33R. Дои:10,3354 / MEPS207033. Получено 12 сентября 2009.
- ^ а б "канатик". Словарь случайных домов. Случайный дом. Получено 2 августа 2009.
- ^ Хейворд, П.Дж. (1985). «Систематическая часть». Ctenostome Bryozoans. Конспекты британской фауны. Линнеевское общество Лондона. С. 106–107. ISBN 978-90-04-07583-2. Получено 2 августа 2009.
- ^ Гьер, О. (2009). «Тентакулата». Мейобентология (2-е изд.). Springer-Verlag. п. 227. ISBN 978-3-540-68657-6. Получено 2 августа 2009.
- ^ Liddell, H.G .; Скотт Р. (1940). "кенос". Греко-английский лексикон. Кларендон Пресс. ISBN 978-0-19-864226-8. Получено 1 августа 2009.
- ^ Branch, M.L .; Griffiths, C.I .; Бекли, Л. (2007). «Мшанки: мох или кружевные животные». Два океана - Путеводитель по морской жизни в южной части Африки. Струик. С. 104–110. ISBN 978-1-77007-633-4. Получено 2 августа 2009.
- ^ Little, W .; Fowler, H.W .; Коулсон, Дж. И Луки, К. (1959). «Мшанки». Краткий оксфордский словарь английского языка. Оксфордский университет. ISBN 978-0-19-860613-0.
- ^ Eckman, J.E .; Окамура, Б. (декабрь 1998 г.). «Модель захвата частиц мшанками в турбулентном потоке: значение формы колонии». Американский натуралист. 152 (6): 861–880. Дои:10.1086/286214. PMID 18811433. S2CID 5535013.
- ^ Фогель, С. (1996). «Жизнь в градиентах скорости». Жизнь в движущихся жидкостях (2-е изд.). Издательство Принстонского университета. п. 191. ISBN 978-0-691-02616-9. Получено 5 августа 2009.
- ^ фон Дассов, М. (1 августа 2006 г.). «Функционально-зависимое развитие колониального животного». Биологический бюллетень. 211 (1): 76–82. Дои:10.2307/4134580. ISSN 0006-3185. JSTOR 4134580. PMID 16946244. Архивировано из оригинал 6 июля 2009 г.. Получено 5 августа 2009.
- ^ Тейлор, Пол Д .; Джеймс, Ноэль П. (август 2013 г.). «Вековые изменения колоний и карбонатных отложений мшанок в геологической истории». Седиментология. 60 (5): 1184–1212. Дои:10.1111 / сед.12032.
- ^ а б Beatty, J.A .; Блэквелдер (1974). «Имена типа беспозвоночных». Систематическая зоология. 23 (4): 545–547. Дои:10.2307/2412472. JSTOR 2412472.
- ^ а б Майр, Э. (июнь 1968 г.). «Мшанки против Ectoprocta». Систематическая зоология. 17 (2): 213–216. Дои:10.2307/2412368. JSTOR 2412368.
- ^ Little, W .; Фаулер, H.W; Коулсон, Дж. И Луки, К. (1964). «Экто-». Краткий оксфордский словарь английского языка. Издательство Оксфордского университета. ISBN 978-0-19-860613-0.
- ^ Little, W .; Фаулер, H.W; Коулсон, Дж. И Луки, К. (1964). «Энто-». Краткий оксфордский словарь английского языка. Издательство Оксфордского университета. ISBN 978-0-19-860613-0.
- ^ а б c d е ж грамм час Галаныч, К.М. (2004). «Новый взгляд на филогению животных» (PDF). Ежегодный обзор экологии, эволюции и систематики. 35: 229–256. Дои:10.1146 / annurev.ecolsys.35.112202.130124. Получено 26 августа 2016.
- ^ а б c d е ж Hausdorf, B .; Helmkampf, M; Мейер, А; Витек, А; Herlyn, H; Брухауз, I; Ганкельн, Т; Ударил, TH; Либ, Б. (2007). «Спиральная филогеномика поддерживает воскрешение мшанок, включающих Ectoprocta и Entoprocta». Молекулярная биология и эволюция. 24 (12): 2723–2729. Дои:10,1093 / молбев / мсм214. PMID 17921486.
- ^ Каффи, Р. Дж. (1969). «Мшанки против Ectoprocta - Необходимость точности». Систематическая зоология. 18 (2): 250–251. Дои:10.2307/2412617. JSTOR 2412617.
- ^ Гизелин, М. (1977). «Об изменении названий высших таксонов». Систематическая зоология. 26 (3): 346–349. Дои:10.2307/2412681. JSTOR 2412681.
- ^ Yokobori, S .; Изето, Т; Асакава, S; Сасаки, Т; Shimizu, N; Ямагиши, А; Осима, Т; Хиросе, Э (май 2008 г.). «Полные нуклеотидные последовательности митохондриальных геномов двух одиночных энтопроктов, Локсокорон аллакс и Loxosomella aloxiata: Значение для филогении лофотрохозойных ". Молекулярная филогенетика и эволюция. 47 (2): 612–628. Дои:10.1016 / j.ympev.2008.02.013. PMID 18374604.
- ^ Рейнольдс, К. (2000). «Таксономически важные особенности на поверхности флотобластов в Plumatella (Bryozoa) ". Микроскопия и микроанализ. 6 (3): 202–210. Bibcode:2000MiMic ... 6..202R. Дои:10.1017 / S1431927600000349. PMID 10790488. Текст начинается «Тип Ectoprocta (Bryozoa) ...»
- ^ Трамбл, Вт; Браун, L (2002). «Мшанки». Краткий оксфордский словарь английского языка. Oxford University Press, США. ISBN 978-0-19-860457-0.
- ^ Тейлор, Пол Д (октябрь 2008 г.). «Таксономия родов мшанок Oncousoecia, Microeciella и Eurystrotos». Журнал естественной истории. 42 (39–40): 2557–2574. Дои:10.1080/00222930802277640. S2CID 84315311.
- ^ Чепмен, А.Д. (2006). Количество живых видов в Австралии и в мире (PDF). Департамент окружающей среды и наследия правительства Австралии. п. 34. ISBN 978-0-642-56849-6. Получено 7 августа 2009.
- ^ Ruppert, E.E .; Фокс, Р. И Барнс, Р.Д. (2004). «Знакомство с беспозвоночными». Зоология беспозвоночных (7-е изд.). Брукс / Коул. стр.2–9. ISBN 978-0-03-025982-1.
- ^ "Стандартная страница отчета ITIS: Phylactolaemata". Интегрированная система таксономической информации. Получено 12 августа 2009.
- ^ а б c d е ж грамм час Massard, J.A .; Геймер, Габи (2008). «Глобальное разнообразие мшанок (Bryozoa или Ectoprocta) в пресной воде». Гидробиология. 595: 93–99. Дои:10.1007 / s10750-007-9007-3. S2CID 13057599.
- ^ а б Fish, J.D .; Рыба, С. (1996). «Мшанки». Путеводитель по морскому берегу для студентов. (2-е изд.). Кембридж: Издательство Кембриджского университета. стр.418–419. ISBN 978-0-521-46819-0.
- ^ Яблонски, Д .; Lidgard, S .; Тейлор, П. (1997). «Сравнительная экология излучений мшанок: происхождение новинок в циклостомах и хейлостомах». ПАЛАИ. 12 (6): 505–523. Bibcode:1997Палай..12..505J. Дои:10.2307/3515408. JSTOR 3515408.
- ^ а б Hayward, P.J .; Райланд, Дж. (1985). «Определитель высших таксонов морских Bryozoa». Cyclostome bryozoans. Линнеевское общество Лондона. п. 7. ISBN 978-90-04-07697-6. Получено 9 августа 2009.
- ^ а б МакКинни; Фрэнк К; Джереми. «Эволюция мшанок». Бостон: Анвин и Хайман, 1989.
- ^ Torsvik, T.H .; Райан, Пол Д .; Тренч, Аллан; Харпер, Дэвид А. (Январь 1991 г.). «Кембрийско-ордовикская палеогеография Балтики». Геология. 19 (1): 7–10. Bibcode:1991Гео .... 19 .... 7Т. Дои:10.1130 / 0091-7613 (1991) 019 <0007: COPOB> 2.3.CO; 2.
- ^ а б c d е Dewel, R.A .; Winston, J.E .; Маккинни, Ф.Дж. (2002). «Деконструкция byozoans: происхождение и последствия уникального строения тела». In Wyse Jacksdon, P.E .; Buttler, C.E .; Спенсер Джонс, M.E. (ред.). Мшанология 2001: материалы Двенадцатой Международной конференции по мшанкам. M.E. Lisse: Swets and Zeitlinger. С. 93–96. ISBN 978-90-5809-388-2. Получено 13 августа 2009.
- ^ Олемпская, Э. (2012). «Исключительная сохранность мягких тканей в сверлящих ктеностомных мшанках и связанных с ними« грибковых »скважинах из раннего девона Подолии, Украина». Acta Palaeontologica Polonica. 57 (4): 925–940. Дои:10.4202 / app.2011.0200.
- ^ МакКинни, Ф.К. (1994). «Сто миллионов лет конкурентных взаимодействий между кладами мшанок: асимметричные, но не нарастающие». Биологический журнал Линнеевского общества. 56 (3): 465–481. Дои:10.1111 / j.1095-8312.1995.tb01105.x.
- ^ Вуд Р. (1999). Эволюция рифов. Издательство Оксфордского университета. С. 235–237. ISBN 978-0-19-857784-3. Получено 11 августа 2009.
- ^ а б c Wood, T.S .; Лор М. (2005). «Высшая филогения phylactolaemate bryozoans, выведенная из последовательностей 18S рибосомной ДНК» (PDF). In Moyano, H.I .; Cancino, J.M .; Уайз-Джексон, П. (ред.). Исследования мшанок 2004: Труды 13-й Международной ассоциации мшанок.. Лондон: Taylor & Francis Group. стр. 361–367. Получено 24 августа 2009.
- ^ Поховски, Р. (1978). «Сверкающие ctenostomate bryozoa: систематика и палеобиология на основе полостей в известняковых субстратах». Бюллетени американской палеонтологии. 73: 192 стр.
- ^ Garey, J.R .; Шмидт-Реза, Андреас (1998). «Существенная роль« второстепенного »типа в молекулярных исследованиях эволюции животных». Американский зоолог. 38 (6): 907–917. Дои:10.1093 / icb / 38.6.907.
- ^ а б c Нильсен, К. (2001). «Тип Ectoprocta». Эволюция животных: взаимоотношения живых типов (2-е изд.). Издательство Оксфордского университета. С. 244–264. ISBN 978-0-19-850681-2. Получено 14 августа 2009.
- ^ "Введение в Hemichordata". Музей палеонтологии Калифорнийского университета. Архивировано из оригинал 1 февраля 2019 г.. Получено 22 сентября 2008.
- ^ а б c Helmkampf, M .; Брухгауз, Ирис; Хаусдорф, Бернхард (2008). «Филогеномный анализ лофофоратов (брахиопод, форонид и мшанок) подтверждает концепцию Lophotrochozoa». Труды Королевского общества B: биологические науки. 275 (1645): 1927–1933. Дои:10.1098 / rspb.2008.0372. ЧВК 2593926. PMID 18495619.
- ^ Нильсен, К; Worsaae, K (сентябрь 2010 г.). «Строение и встречаемость личинок cyphonautes (Bryozoa, Ectoprocta)». Журнал морфологии. 271 (9): 1094–1109. Дои:10.1002 / jmor.10856. PMID 20730922. S2CID 11453241.
- ^ Шэнь Х, Тянь М, Мэн Х, Лю Х, Ченг Х, Чжу Ц., Чжао Ф. (сентябрь 2012 г.). "Полный митохондриальный геном Membranipora grandicella (Bryozoa: Cheilostomatida), определенный с помощью секвенирования следующего поколения: первый представитель подотряда Malacostegina ". Сравнительная биохимия и физиология, часть D: геномика и протеомика. 7 (3): 248–253. Дои:10.1016 / j.cbd.2012.03.003. PMID 22503287.
- ^ а б c Callaghan, T.P .; Р., Карлсон (июнь 2002 г.). «Летний покой как убежище от гибели пресноводных мшанок». Plumatella emarginata". Oecologia. 132 (1): 51–59. Bibcode:2002Oecol.132 ... 51C. Дои:10.1007 / s00442-002-0946-0. PMID 28547286. S2CID 19925846.
- ^ а б c Пратт, М. (2008). «Жизнь там, где течет правильный поток: как поток влияет на питание мшанок». Интегративная и сравнительная биология. 48 (6): 808–822. Дои:10.1093 / icb / icn052. PMID 21669834.
- ^ Райланд, Дж. (1967). «Дыхание у полизоа (эктопрокта)». Природа. 216 (5119): 1040–1041. Bibcode:1967Натура.216.1040R. Дои:10.1038 / 2161040b0. S2CID 4207120.
- ^ Стратманн, Р.Р. (март 2006 г.). «Универсальное цилиарное поведение при захвате частиц личинкой мшанки cyphonautes». Acta Zoologica. 87 (1): 83–89. Дои:10.1111 / j.1463-6395.2006.00224.x.
- ^ а б Wood, T.S .; Окамура, Бет (декабрь 1998 г.). "Asajirella gelatinosa в Панаме: расширение ареала мшанок в Западном полушарии ". Гидробиология. 390 (1–3): 19–23. Дои:10.1023 / А: 1003502814572. S2CID 1525771.
- ^ О'Ди, Джексон, Тейлор, Родригес. «Способы размножения современных и ископаемых купуладриидных мшанок». Палеонтология.CS1 maint: использует параметр авторов (связь)[постоянная мертвая ссылка ]
- ^ Эмилиани, К. (1992). «Палеозой». Планета Земля: космология, геология и эволюция жизни и окружающей среды. Издательство Кембриджского университета. стр.488–490. ISBN 978-0-19-503652-7. Получено 11 августа 2009.
- ^ а б c d Джонс, Р. В. (2006). «Основные группы ископаемых». Прикладная палеонтология. Издательство Кембриджского университета. п. 116. ISBN 978-0-521-84199-3. Получено 11 августа 2009.
- ^ Куклински, П .; Бадер, Беате (2007). «Сравнение ассоциаций мшанок из двух контрастирующих регионов арктического шельфа». Эстуарии, прибрежные районы и шельфовые науки. 73 (3–4): 835–843. Bibcode:2007 ECSS ... 73..835K. Дои:10.1016 / j.ecss.2007.03.024.
- ^ Пелагический мшанок из Антарктиды | SpringerLink
- ^ Мэтт МакГрат (16 июня 2014 г.). "'Сорняк процветает, пока теплые берега Антарктиды ». Новости BBC. Получено 16 июн 2014.
- ^ Brusca, R; Бруска, Г. «Беспозвоночные (2-е издание)». Сандерленд, Массачусетс: Sinauer Associates.
- ^ Рамель, Г. "Тип Bryozoa (Bryozoa)". Earthlife. Отсутствует или пусто
| url =(помощь) - ^ а б Маргулис, Л .; Шварц К.В. (1998). «Мшанки». Пять королевств: иллюстрированный справочник по типам жизни на Земле. Эльзевир. п.335. ISBN 978-0-7167-3027-9. Получено 20 августа 2009.
- ^ Iyengar, E.V .; Харвелл, CD (2002). «Специфика сигналов, вызывающих защитные шипы у мшанок Membraniporambranacea». Серия "Прогресс морской экологии". 225: 205–218. Bibcode:2002MEPS..225..205I. Дои:10,3354 / meps225205. Получено 18 августа 2009.
- ^ Hayward, P.J .; Райланд, Дж. (1985). «Хищники». Cyclostome bryozoans: определители и примечания для определения вида. Brill Archive. п. 27. ISBN 978-90-04-07697-6. Получено 18 августа 2009.
- ^ Day, R.W .; Осман, Р. В. (январь 1981 г.). "Хищничество Патирия миниата (Asteroidea) на мшанках ». Oecologia. 51 (3): 300–309. Bibcode:1981Oecol..51..300D. Дои:10.1007 / BF00540898. PMID 28310012. S2CID 19976956.
- ^ McKinney, F.K .; Taylor, P.D .; Лидгард, С. (2003). «Хищничество мшанок и его отражение в летописи окаменелостей». В Kelley, P.H .; Ковалевски, М .; Хансен, Т. (ред.). Взаимодействие хищника и жертвы в летописи окаменелостей. Springer. С. 239–246. ISBN 978-0-306-47489-7. Получено 18 августа 2009.
- ^ Вуд, Т. (Октябрь 2006 г.). «Пресноводные мшанки Таиланда (Ectoprocta и Entoprocta)» (PDF). Журнал естественной истории университета Чулалонгкорн. 6 (2): 83–119. Получено 24 августа 2009.
- ^ Вуд, Т. (Май 2006 г.). "Интенсивное истребление пресноводных мшанок золотой яблочной улиткой, Pomacea canaliculata" (PDF). Журнал естествознания Чулалонгкорнского университета. 6 (1): 31–36. Архивировано из оригинал (PDF) 6 октября 2011 г.. Получено 18 августа 2009.
- ^ Пьюс, С. (2007). «Симбиотические отношения между гидроидами и мшанками». Конгресс Международного общества симбиоза номер 5. 44 (1–3): 137–143. Получено 18 августа 2009.
- ^ Gray, C.A .; McQuaid, CD; Дэвис-Коулман, MT (декабрь 2005 г.). «Симбиотические мшанки, покрытые оболочкой, обеспечивают сублиторальных щенков химической защитой от каменных лобстеров». Африканский журнал морских наук. 27 (3): 549–556. Дои:10.2989/18142320509504115. S2CID 84531235.
- ^ Кликпера, Андре; Тейлор, Пол Д .; Вестфаль, Хильдегард (30 июля 2013 г.). «Бриолиты, созданные мшанками в симбиотических ассоциациях с раками-отшельниками в тропической гетерозойной карбонатной системе, Гольф-д'Арген, Мавритания». Морское биоразнообразие. 43 (4): 429–444. Дои:10.1007 / s12526-013-0173-4. S2CID 15841444.
- ^ а б Андерсон, С .; Canning, E.U .; Окамура, Б. (1999). «Молекулярные данные указывают на то, что мшанки являются хозяевами для PKX (Phylum Myxozoa), и идентифицируют кладу мшаночных паразитов в составе Myxozoa». Паразитология. 119 (6): 555–561. Дои:10.1017 / S003118209900520X. PMID 10633916. Получено 18 августа 2009.
- ^ Клин, Б. (2008). «Профессиональная светочувствительная экзема рыбаков при контакте с мшанками: инвалидность профессионального дерматоза» (PDF). Международное морское здравоохранение. 59 (1–4): 1–4. PMID 19227737. Получено 18 августа 2009.
- ^ Wood, T.S .; Марш, Терренс Дж. (Февраль 1999 г.). «Биообрастание очистных сооружений пресноводной мшанкой, Plumatella vaihiriae". Водные исследования. 33 (3): 609–614. Дои:10.1016 / S0043-1354 (98) 00274-7.
- ^ «Бриостатин 1». 19 июня 2006 г. Архивировано с оригинал 9 мая 2007 г.. Получено 20 августа 2009.
- ^ «Исследование безопасности, эффективности, фармакокинетики и фармакодинамики бриостатина 1 у пациентов с болезнью Альцгеймера». Национальные институты здоровья. 19 августа 2009 г.. Получено 20 августа 2009.
- ^ Nelsen et. др., JT Влияние бриостатина на когнитивные функции и PKCɛ в фазе IIa болезни Альцгеймера и в исследованиях расширенного доступа J. Alzheimers Dis. 2017; 58 (2): 521–535. Доступ: 27 декабря 2017 г.
- ^ Wender, P.A .; Барыза, JL; Беннетт, CE; Би, ФК; Brenner, SE; Кларк, Миссури; Horan, JC; Кан, С; и другие. (20 ноября 2002 г.). «Практический синтез нового и сильнодействующего аналога Бриостатина». Журнал Американского химического общества. 124 (46): 13648–13649. Дои:10.1021 / ja027509 +. PMID 12431074.
дальнейшее чтение
- Холл, С.Р .; Тейлор, PD; Davis, SA; Манн, S (2002). «Электронографические исследования известковых скелетов мшанок». Журнал неорганической биохимии. 88 (3–4): 410–419. Дои:10.1016 / S0162-0134 (01) 00359-2. PMID 11897358.
- Хейворд, П.Г., Дж. Райланд и П.Д. Тейлор (ред.), 1992. Биология и палеобиология мшанок, Olsen and Olsen, Фреденсборг, Дания.
- Уинстон, Дж. Э. (2010). "Жизнь в колониях: изучение чужеродных путей колониальных организмов". Интегративная и сравнительная биология. 50 (6): 919–33. Дои:10.1093 / icb / icq146. PMID 21714171.
- Робисон, Р. (ред.), 1983. Трактат по палеонтологии беспозвоночных, Часть G, Bryozoa (исправлено). Геологическое общество Америки и Университет Канзаса Press.
- Шарп, JH; Уинсон, МК; Портер, JS (2007). «Метаболиты мшанок: экологическая перспектива» (PDF). Отчеты о натуральных продуктах. 24 (4): 659–73. Дои:10.1039 / b617546e. HDL:2160/3792. PMID 17653353.
- Тейлор, П; Уилсон, М.А. (2003). «Палеоэкология и эволюция морских сообществ твердого субстрата» (PDF). Обзоры наук о Земле. 62 (1): 1–103. Bibcode:2003ESRv ... 62 .... 1Т. Дои:10.1016 / S0012-8252 (02) 00131-9. Архивировано из оригинал (PDF) 25 марта 2009 г.
- Винн О., Уилсон М.А., Мытус М.-А. и Тоом, У. (2014). «Самый ранний мшаночный паразит: средний ордовик (дарривиль) острова Осмуссаар, Эстония». Палеогеография, палеоклиматология, палеоэкология. 414: 129–132. Bibcode:2014ППП ... 414..129В. Дои:10.1016 / j.palaeo.2014.08.021. Получено 9 января 2014.CS1 maint: использует параметр авторов (связь)
- Woollacott, R.M. и Р.Л. Циммер (ред.), 1977. Биология мшанок, Academic Press, Нью-Йорк.
внешняя ссылка
- Указатель Bryozoa Домашняя страница Bryozoa, была в RMIT; сейчас bryozoa.net
- Другие ресурсы по мшанкам в Интернете
- Международная ассоциация бриозоологов Официальный веб-сайт
- Неоген Bryozoa Британии
- Мшанки Введение
- Тип Ectoprocta (Bryozoa)
- Тип Bryozoa в Wikispecies
- Мшанки в Река Коннектикут
- Информационный бюллетень Bryozoa