Амеба - Amoeba

An амеба (/əˈмябə/; реже пишется амеба или же амеба; множественное число am (o) ebas или же am (o) ebae /əˈмябя/),[1] часто называют амебовидный, это тип клетка или же одноклеточный организм который имеет способность изменять свою форму, в первую очередь за счет выдвижения и втягивания ложноножки.[2] Амебы не образуют единого таксономическая группа; вместо этого они встречаются во всех основных родословная из эукариотический организмы. Амебоидные клетки встречаются не только среди простейшие, но и в грибы, водоросли, и животные.[3][4][5][6][7]
Микробиологи часто используют термины «амебоид» и «амеба» как синонимы для любого организма, который проявляет амебовидное движение.[8][9]
В старых системах классификации большинство амеб помещалось в учебный класс или же подтип Саркодина, группировка одноклеточные организмы которые обладают ложноножками или передвигаются протоплазматический поток. Тем не мение, молекулярный филогенетический исследования показали, что Sarcodina не монофилетический группа, участники которой разделяют общее происхождение. Следовательно, амебоидные организмы больше не объединяются в одну группу.[10]
Самый известный амебоид протисты находятся Хаос Каролиненс и Амеба протей, оба из которых широко культивировались и изучались в классах и лабораториях.[11][12] Другие хорошо известные виды включают так называемую амебу, поедающую мозг. Naegleria fowleri кишечного паразита Entamoeba histolytica, что приводит к амебной дизентерией, а многоклеточная «социальная амеба» или слизь Dictyostelium discoideum.
Форма, движение и питание

У амеб нет клеточных стенок, что позволяет им свободно передвигаться. Амебы передвигаются и питаются с помощью псевдопод, которые представляют собой выпуклости цитоплазма сформированный согласованным действием актин микрофиламенты выталкивая плазматическая мембрана что окружает клетку.[13] Внешний вид и внутреннее строение ложноножек используются для того, чтобы отличать группы амеб друг от друга. Амебозойный виды, например, принадлежащие к роду Амеба, как правило, имеют луковичные (лопастные) ложноножки, закругленные на концах и примерно трубчатые в поперечном сечении. Cercozoan амебоиды, такие как Эуглифа и Громия, имеют тонкие нитевидные (филозные) ложноножки. Фораминиферы испускают тонкие ветвящиеся псевдоподы, которые сливаются друг с другом, образуя сетчатые (ретикулезные) структуры. Некоторые группы, такие как Радиолярии и Heliozoa, имеют жесткие, игольчатые, расходящиеся аксоподия (актиноподы) поддерживаются изнутри пучками микротрубочки.[3][14]


Свободноживущие амебы могут быть "завещать "(заключенный в твердую оболочку) или" голый "(также известный как гимнастика, без твердого покрытия). Оболочки семенниковых амеб могут состоять из различных веществ, в том числе: кальций, кремнезем, хитин, или склеивания найденных материалов, таких как мелкие песчинки и панцири из диатомеи.[15]
Регулировать осмотическое давление, большинство пресноводных амеб обладают сократительной вакуоль который удаляет лишнюю воду из клетки.[16] Этот органелла необходимо, потому что пресная вода имеет более низкую концентрацию растворенные вещества (например, соль), чем внутренние жидкости амебы (цитозоль ). Потому что окружающая вода гипотонический Что касается содержимого клетки, вода переносится через клеточную мембрану амебы посредством осмос. Без сократительной вакуоли клетка заполнится избытком воды и, в конце концов, лопнет. Морские амебы обычно не обладают сократительной вакуолью, поскольку концентрация растворенных веществ внутри клетки находится в равновесии с концентрацией растворенных веществ в клетке. тонус окружающей воды.[17]
Рацион питания

Источники питания амеб различны. Некоторые амебы являются хищниками и питаются бактериями и другими протисты. Некоторые детритофаги и есть мертвые органические вещества.
Амебы обычно глотают пищу фагоцитоз, расширяющие псевдоножки, чтобы окружать и поглощать живую добычу или частицы извлеченного материала. Амебоидные клетки не имеют рта или цитостом, и на клетке нет фиксированного места, в котором обычно происходит фагоцитоз.[18]
Некоторые амебы также питаются пиноцитоз, впитывая растворенные питательные вещества через пузырьки образуется внутри клеточной мембраны.[19]
Диапазон размеров

Размер амебоидных клеток и видов очень разнообразен. Морской амебоид Massisteria voersi всего 2,3 к 3 микрометры в диаметре,[20] в пределах размера многих бактерий.[21] С другой стороны, раковины глубоководных ксенофиофоры может достигать 20 см в диаметре.[22] Большинство свободноживущих пресноводных амеб, которые обычно встречаются в пруду, канавах и озерах, являются микроскопический, но некоторые виды, например, так называемые «гигантские амебы» Pelomyxa palustris и Хаос Каролиненс, может быть достаточно большим, чтобы увидеть его невооруженным глазом.
| Виды или тип клеток | Размер в микрометрах |
|---|---|
| Massisteria voersi[20] | 2.3–3 |
| Naegleria fowleri[23] | 8–15 |
| Нейтрофил (лейкоцит)[24] | 12–15 |
| Акантамеба[25] | 12–40 |
| Entamoeba histolytica[26] | 15–60 |
| Арчелла обыкновенная[27] | 30–152 |
| Амеба протей[28] | 220–760 |
| Хаос Каролиненс[29] | 700–2000 |
| Pelomyxa palustris[30] | до 5000 |
| Syringammina fragilissima[22] | вплоть до 200000 |
Амебы как специализированные клетки и стадии жизненного цикла

Немного многоклеточные организмы имеют амебоидные клетки только на определенных этапах жизни или используют амебоидные движения для выполнения специализированных функций. В иммунной системе человека и других животных амебовидный белые кровяные клетки преследовать вторгшиеся организмы, такие как бактерии и патогенные простейшие, и поглощать их фагоцитоз.[31]
Амебоидные стадии также встречаются у многоклеточных грибовидных протистов, так называемых слизевые формы. Обе формы плазмодиальной слизи, в настоящее время классифицируемые по классу Миксогастрия, и ячеистые слизевики групп Acrasida и Диктиостелиида, живут как амебы на стадии питания. Амебоидные клетки первого объединяются, образуя гигантский многоядерный организм,[32] в то время как клетки последних живут отдельно до тех пор, пока не закончится еда, и в это время амебы объединяются, образуя многоклеточный мигрирующий «слизень», который функционирует как единый организм.[8]
Другие организмы также могут представлять амебоидные клетки на определенных этапах жизненного цикла, например, гаметы некоторых зеленых водорослей (Zygnematophyceae )[33] и пеннат диатомеи,[34] споры (или фазы распространения) некоторых Мезомицетозое,[35][36] и спороплазма этап Myxozoa и из Аскетоспорея.[37]
Амебы как организмы
Ранняя история и происхождение Саркодины
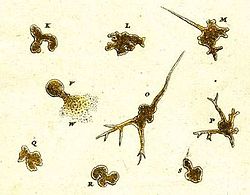
Самая ранняя запись об амебоидном организме была произведена в 1755 г. Август Иоганн Рёзель фон Розенхоф, который назвал свое открытие «Der Kleine Proteus» («Маленький Протеус»).[38] На иллюстрациях Рёзеля изображена неидентифицируемая пресноводная амеба, внешне похожая на обычный вид, известный сейчас как Амеба протей.[39] Термин «Proteus animalcule» оставался в употреблении на протяжении XVIII и XIX веков как неофициальное название любого крупного свободно живущего амебоида.[40]
В 1822 г. род Амиба (от Греческий ἀμοιβή амойб, что означает «изменение») был построен французским натуралистом Бори де Сент-Винсент.[41][42] Современник Бори, К. Г. Эренберг, принял этот род в своей классификации микроскопических существ, но изменил написание на Амеба.[43]
В 1841 г. Феликс Дюжарден ввел термин "саркод"(от греческого σάρξ саркс, "плоть" и εἶδος эйдос, «форма») для «густого, клейкого, однородного вещества», которое заполняет тела клеток простейших.[44] Хотя этот термин первоначально относился к протоплазме любого простейшего, вскоре он стал использоваться в ограниченном смысле для обозначения студенистого содержимого амебоидных клеток.[10] Тридцать лет спустя австрийский зоолог Людвиг Карл Шмарда использовал «саркод» в качестве концептуальной основы для своего подразделения Sarcodea, филюм -уровневая группа, состоящая из «нестабильных, изменчивых» организмов, тела которых в основном состоят из «саркода».[45] Более поздние работники, в том числе влиятельный систематик Отто Бютчли, исправили эту группу, чтобы создать класс Sarcodina,[46] а таксон которые оставались широко используемыми на протяжении большей части 20 века.
В пределах традиционной Sarcodina амебы обычно подразделялись на морфологические категории, исходя из формы и структуры их ложноножки. Амебы с псевдоножками, поддерживаемые регулярными массивами микротрубочки (например, пресноводный Heliozoa и морской Радиолярии ) были классифицированы как Актиноподы; тогда как те, у кого ложные ножки не поддерживались, были классифицированы как Ризоподы.[47] В соответствии с морфологией их ложноногих, Rhizopods были подразделены на лобозные, филозовые и ретикулезные амебы.
Демонтаж Саркодины
В последнее десятилетие 20-го века серия молекулярных филогенетических анализов подтвердила, что Sarcodina не была монофилетический группа. Ввиду этих результатов от старой схемы отказались, и амебы Sarcodina были рассредоточены среди многих других таксономических групп высокого уровня. Сегодня большинство традиционных саркодинов помещено в два эукариотических тела. супергруппы: Амебозоа и Ризария. Остальные были распределены между раскапывает, опистоконцы, и страменопилы. Некоторые, как Centrohelida, еще не попали ни в одну супергруппу.[10][48]
Классификация
Согласно недавней классификации различные роды амебоидов разделены на следующие группы:
| Супергруппы | Основные группы и роды | Морфология |
|---|---|---|
| Амебозоа |
|
|
| Ризария |
| |
| Экскавата |
| |
| Гетероконта |
|
|
| Альвеолаты |
| |
| Опистоконта |
|
|
| Разгруппирован/ неизвестный |
|
Некоторые из упомянутых групп амебоидов (например, часть хризофиты, часть ксантофиты, хлорарахниофиты ) не были традиционно включены в Саркодину, будучи классифицированными как водоросли или же бичеванный простейшие.
Патогенные взаимодействия с другими организмами

Некоторые амебы могут заражать другие организмы патогенно, вызывающие заболевание:[52][53][54][55]
- Entamoeba histolytica является причиной амебиаз, или амебная дизентерия.
- Naegleria fowleri («амеба, поедающая мозг») - это обитающий в пресной воде вид, который может быть смертельным для человека при попадании через нос.
- Акантамеба может вызвать амебный кератит и энцефалит в людях.
- Balamuthia mandrillaris является причиной (часто со смертельным исходом) гранулематозный амебный менингоэнцефалит.
- Было обнаружено, что амеба собирать урожай и расти бактерии, причастные к чума.
- Амебы также могут быть хозяевами микроскопических организмов, которые являются патогенными для людей и помогают распространять такие микробы. Бактериальные возбудители (например, Легионелла ) может препятствовать поглощению пищи при поедании амебами.[56]
- В настоящее время наиболее широко используемыми и наиболее изученными амебами, которые являются хозяевами других организмов, являются Acanthamoeba castellanii и Dictyostelium discoideum.[57]
- Микроорганизмы, которые могут преодолевать охрану одноклеточных животных, увеличивают укрытие, в котором они могут размножаться, где они защищены от неблагоприятных внешних условий их случайными хозяевами.
Мейоз
Последние данные показывают, что несколько линий Amoebozoa претерпевают мейоз.
Ортологи генов, используемых в мейоз сексуального эукариоты были недавно обнаружены в Акантамеба геном. Эти гены включали Spo11, Mre11, Rad50, Rad51, Rad52, Mnd1, Dmc1, Мш и Mlh.[58] Это открытие предполагает, что «Acanthamoeba» способны к некоторой форме мейоза и могут подвергаться половому размножению.
Специфический для мейоза рекомбиназа, Dmc1, необходим для эффективного мейотического гомологичная рекомбинация, и Dmc1 выражается в Entamoeba histolytica.[59] Очищенный Dmc1 от E. histolytica формы пресинаптический волокна и катализаторы АТФ -зависимый гомологичное спаривание ДНК и обмен цепями ДНК более чем на несколько тысяч пар оснований.[59] Реакции спаривания ДНК и обмена цепей усиливаются эукариотическим мейоз-специфическим дополнительным фактором рекомбинации (гетеродимером) Hop2-Mnd1.[59] Эти процессы являются центральными для мейотической рекомбинации, предполагая, что E. histolytica претерпевает мейоз.[59]
Исследования Entamoeba invadens обнаружил, что при преобразовании из тетраплоид безъядерный трофозоит к четырехъядерной кисте, гомологичная рекомбинация усиливается.[60] Экспрессия генов с функциями, связанными с основными этапами мейотической рекомбинации, также увеличивается во время инцистирования.[60] Эти выводы в E. invadensв сочетании с данными исследований E. histolytica указывают на наличие мейоза в Entamoeba.
Dictyostelium discoideum в супергруппе Амебозоа может подвергнуться спариванию и половое размножение включая мейоз при недостатке пищи.[61][62]
Поскольку амебозоа рано отошли от эукариотический генеалогическое древо, эти результаты предполагают, что мейоз присутствовал на ранних этапах эволюции эукариот. Кроме того, эти результаты согласуются с предложением Lahr et al.[63] что большинство амебоидных клонов имеют древнюю половую принадлежность.
Рекомендации
- ^ «Амеба» В архиве 22 ноября 2015 г. Wayback Machine на Oxford Commandings.com
- ^ Синглтон, Пол (2006). Словарь по микробиологии и молекулярной биологии, 3-е издание, переработанное. Чичестер, Великобритания: John Wiley & Sons. стр.32. ISBN 978-0-470-03545-0.
- ^ а б Дэвид Дж. Паттерсон. «Амебы: протисты, которые передвигаются и питаются псевдоподиями». Интернет-проект "Древо жизни". В архиве из оригинала 15 июня 2010 г.. Получено 21 сентября 2009.
- ^ «Амебы». Эдинбургский университет. Архивировано из оригинал 10 июня 2009 г.
- ^ Вим ван Эгмонд. «Солнечные звери и амебы». Микроскопия-Великобритания. В архиве с оригинала 4 ноября 2005 г.. Получено 23 октября 2005.
- ^ Флор-Парра, Игнасио; Бернал, Мануэль; Журинский, Яков; Дага, Рафаэль Р. (17 декабря 2013 г.). «Миграция и деление клеток в амебоидных делящихся дрожжах». Биология Открыть. 3 (1): 108–115. Дои:10.1242 / bio.20136783. ISSN 2046-6390. ЧВК 3892166. PMID 24357230.
- ^ Friedl, P .; Borgmann, S .; Бреккер, Э. Б. (1 октября 2001 г.). «Амебоидные лейкоциты, пролезающие через внеклеточный матрикс: уроки парадигмы движения клеток Dictyostelium». Журнал биологии лейкоцитов. 70 (4): 491–509. ISSN 0741-5400. PMID 11590185.
- ^ а б Мари, Афанасий FM; Хогевег, Полин (2001). «Как амебоиды самоорганизуются в плодовое тело: координация многоклеточных организмов у Dictyostelium discoideum». Труды Национальной академии наук. 98 (7): 3879–3883. Дои:10.1073 / pnas.061535198. ЧВК 31146. PMID 11274408.
- ^ Mackerras, M. J .; Эрколе, К. Н. (1947). «Наблюдения за действием палудрина на малярийных паразитов». Труды Королевского общества тропической медицины и гигиены. 41 (3): 365–376. Дои:10.1016 / s0035-9203 (47) 90133-8. PMID 18898714.
- ^ а б c Ян Павловски: Сумерки Саркодины: молекулярная перспектива полифилетического происхождения амебоидных протистов. Protistology, Band 5, 2008, S. 281–302. (pdf, 570 кБ) В архиве 14 июня 2013 г. Wayback Machine
- ^ Тан; и другие. (2005). «Простая массовая культура амебы Chaos carolinense: повторение» (PDF). Протистология. 4: 185–90. В архиве (PDF) из оригинала 29 сентября 2017 г.. Получено 28 сентября 2017.
- ^ «Отношения с людьми». Амеба протей. 12 апреля 2013 г. В архиве из оригинала 29 сентября 2017 г.. Получено 28 сентября 2017.
- ^ Alberts Eds .; и другие. (2007). Молекулярная биология клетки 5-е издание. Нью-Йорк: Наука Гарланд. п. 1037. ISBN 9780815341055.
- ^ Маргулис, Линн (2009). Королевства и домены. Академическая пресса. стр.206 –7. ISBN 978-0-12-373621-5.
- ^ Огден, К. Г. (1980). Атлас пресноводных семенников амебы. Оксфорд, Лондон и Глазго: Издательство Оксфордского университета, Британский музей (естественная история). С. 1–5. ISBN 978-0198585022.
- ^ Alberts Eds .; и другие. (2007). Молекулярная биология клетки 5-е издание. Нью-Йорк: Наука Гарланд. п. 663. ISBN 9780815341055.
- ^ Кудо, Ричард Роксабро. «Протозоология». Протозоология 4-е издание (1954). п. 83
- ^ Торп, Джеймс Х. (2001). Экология и классификация пресноводных беспозвоночных Северной Америки. Сан-Диего: академический. п. 71. ISBN 0-12-690647-5.
- ^ Чон, Кван В. (1973). Биология амебы. Нью-Йорк: Academic Press. стр.100.
- ^ а б Мыльников, Александр П .; Вебер, Феликс; Юргенс, Клаус; Wylezich, Claudia (1 августа 2015 г.). «У Massisteria marina есть сестра: Massisteria voersi sp. Nov., Редкий вид, изолированный из прибрежных вод Балтийского моря». Европейский журнал протистологии. 51 (4): 299–310. Дои:10.1016 / j.ejop.2015.05.002. ISSN 1618-0429. PMID 26163290.
- ^ «Размер, форма и расположение бактериальных клеток». classes.midlandstech.edu. Архивировано из оригинал 9 августа 2016 г.. Получено 21 августа 2016.
- ^ а б Gooday, A.J .; Аранда да Силва, А .; Павловски, Дж. (1 декабря 2011 г.). «Xenophyophores (Rhizaria, Foraminifera) из каньона Назаре (окраина Португалии, северо-восточная Атлантика)». Deep-Sea Research Part II: Актуальные исследования в океанографии. Геология, геохимия и биология подводных каньонов к западу от Португалии. 58 (23–24): 2401–2419. Bibcode:2011DSRII..58.2401G. Дои:10.1016 / j.dsr2.2011.04.005.
- ^ «Амеба, поедающая мозг (Naegleria Fowleri): причины и симптомы». В архиве с оригинала 21 августа 2016 г.. Получено 21 августа 2016.
- ^ «Атласы анатомии: Атлас микроскопической анатомии. Раздел 4: Кровь». www.anatomyatlases.org. В архиве с оригинала 19 августа 2016 г.. Получено 21 августа 2016.
- ^ "Акантамеба | Микромир ». www.arcella.nl. Архивировано из оригинал 18 августа 2016 г.. Получено 21 августа 2016.
- ^ "Микроскопия Entamoeba histolytica". msu.edu. В архиве из оригинала 5 октября 2016 г.. Получено 21 августа 2016.
- ^ "Арчелла обыкновенная | Микромир ». www.arcella.nl. В архиве с оригинала 18 августа 2016 г.. Получено 21 августа 2016.
- ^ "Амеба протей | Микромир ». www.arcella.nl. В архиве с оригинала 18 августа 2016 г.. Получено 21 августа 2016.
- ^ "Хаос | Микромир ». www.arcella.nl. В архиве из оригинала 12 октября 2016 г.. Получено 21 августа 2016.
- ^ "Pelomyxa palustris | Микромир ». www.arcella.nl. В архиве с оригинала 18 августа 2016 г.. Получено 21 августа 2016.
- ^ Фридл, Питер; Боргманн, Стефан; Ева-Б, Брекер (2001). «Амебовидные лейкоциты, ползающие через внеклеточный матрикс: уроки парадигмы движения клеток Dictyostelium». Журнал биологии лейкоцитов. 70 (4): 491–509. PMID 11590185.
- ^ Накагаки; и другие. (2000). «Интеллект: решение лабиринта амебоидным организмом». Природа. 407 (6803): 470. Bibcode:2000Натура.407..470Н. Дои:10.1038/35035159. PMID 11028990. S2CID 205009141.
- ^ Вер, Джон Д. (2003). Пресноводные водоросли Северной Америки. Сан-Диего и Лондон: Academic Press. стр.353. ISBN 978-0-12-741550-5.
- ^ «Мир водорослей: пол и жизненные циклы диатомовых водорослей». Мир водорослей. Королевский ботанический сад Эдинбурга. В архиве из оригинала 23 сентября 2014 г.. Получено 1 марта 2015.
- ^ Валле, Л. (2014). «Новые виды Paramoebidium (trichomycetes, Mesomycetozoea) из Средиземного моря, с комментариями об амебоидных клетках у Amoebidiales». Микология. 106 (3): 481–90. Дои:10.3852/13-153. PMID 24895422. S2CID 3383757.
- ^ Тейлор, Дж. У. и Берби, М. Л. (2014). Грибы от ПЦР до геномики: распространяющаяся революция в эволюционной биологии. В: Систематика и эволюция. Springer Berlin Heidelberg. п. 52, [1] В архиве 30 июня 2015 г. Wayback Machine
- ^ Корлисс, Дж. О. (1987). «Филогения и эукариогенез протистана». Международный обзор цитологии. 100: 319–370. Дои:10.1016 / S0074-7696 (08) 61703-9. ISBN 9780080586373. PMID 3549607.
- ^ Розенхоф, Р. (1755). Monatlich herausgegebene Insektenbelustigungen, т. 3, стр. 621, г. [2] В архиве 13 июля 2015 г. Wayback Machine.
- ^ Чон, Кван В. (1973). Биология амебы. Нью-Йорк: Academic Press. стр. 2–3, [3].
- ^ Макальпайн, Дэниел (1881). Биологический атлас: руководство по практическому изучению растений и животных. Эдинбург и Лондон: W. & A. K. Johnston. стр.17.
- ^ Бори де Сен-Винсент, Ж. Б. Г. М. «Essai d'une классификации микроскопических животных». Agasse, Париж (1826 г.). Стр. 28
- ^ МакГрат, Кимберли; Блэчфорд, Стейси, ред. (2001). Gale Encyclopedia of Science Vol. 1: Трубкозуб-катализатор (2-е изд.). Гейл Групп. ISBN 978-0-7876-4370-6. OCLC 46337140.
- ^ Эренберг, Кристиан Готфрид. Организация, систематика и географическая верхняя часть: Zwei vorträge, in der Akademie der wissenschaften zu Berlin gehalten in den jahren 1828 и 1830. Druckerei der Königlichen akademie der wissenschaften, 1832. 59
- ^ Дюжарден, Феликс (1841). Histoire Naturelle des Zoophytes Infusoires. Париж: Librarie Encyclopedique de Roret. стр.26.
- ^ Шмарда, Людвиг Карл (1871). Зоология. В. Браумюллер. стр.156.
- ^ Бючли, Отто (1882). Klassen und Ordnungen des Thier-Reichs I. Abteilung: Sarkodina und Sporozoa. Paleontologische Entwicklung der Rhisopoda von C. Scwager. п. 1.
- ^ Калкинс, Гэри Н. (1909). Протозоология. Нью-Йорк: Леа и Фебигер. стр.38 –40.
- ^ Adl, Sina M .; и другие. (2012). «Пересмотренная классификация эукариот». Журнал эукариотической микробиологии. 59 (5): 429–93. Дои:10.1111 / j.1550-7408.2012.00644.x. ЧВК 3483872. PMID 23020233.
- ^ а б Park, J. S .; Simpson, A.G.B .; Brown, S .; Чо, Б. С. (2009). «Ультраструктура и молекулярная филогения двух гетеролобозных амеб, Euplaesiobystra hypersalinica gen. Et sp. Nov. и Tulamoeba peronaphora gen. Et sp. Nov., изолированные из чрезвычайно гиперсоленой среды обитания». Протист. 160 (2): 265–283. Дои:10.1016 / j.protis.2008.10.002. PMID 19121603.
- ^ Отт, Дональд В., Карла К. Олдхэм-Отт, Наталия Рыбалка и Томас Фридл. 2015. Ксантофиты, эустигматофиты и рафидофиты. В: Wehr, J.D., Sheath, R.G., Kociolek, J.P. (Eds.) Пресноводные водоросли Северной Америки: экология и классификация, 2-е изд. Academic Press, Амстердам, стр. 483–534, [4] В архиве 22 января 2017 г. Wayback Machine.
- ^ Паттерсон, Д. Дж .; Simpson, A.G.B .; Роджерсон, А. (2000). «Амебы неопределенного родства». В: Lee, J. J .; Leedale, G.F .; Брэдбери, П. Иллюстрированный путеводитель по простейшим, 2-е изд., Т. 2, стр. 804-827. Лоуренс, Канзас: Общество протозоологов / Allen Press. [5] В архиве 8 марта 2016 г. Wayback Machine. Роды, которые в 2000 г. считались несгруппированными / неизвестными этим источником, но с тех пор стали классифицированными, были перемещены в эти классификации в Википедии.
- ^ Casadevall A (2008) Эволюция внутриклеточных патогенов. Анну Рев Микробиол 62: 19–33. 10.1146 / annurev.micro.61.080706.093305 [PubMed] [CrossRef] [Google Scholar]
- ^ Guimaraes AJ, Gomes KX, Cortines JR, Peralta JM, Peralta RHS (2016) Acanthamoeba spp. в качестве универсального хозяина для патогенных микроорганизмов: один мост от вирулентности окружающей среды к хозяину. Микробиологические исследования 193: 30–38. 10.1016 / j.micres.2016.08.001 [PubMed] [CrossRef] [Google Scholar]
- ^ Hilbi H, Weber SS, Ragaz C, Nyfeler Y, Urwyler S (2007) Экологические хищники как модели бактериального патогенеза. Экологическая микробиология 9: 563–575. 10.1111 / j.1462-2920.2007.01238.x [PubMed] [CrossRef] [Google Scholar]
- ^ Greub, G; Рауль, Д. (2004). «Микроорганизмы, устойчивые к свободноживущим амебам». Обзоры клинической микробиологии. 17 (2): 413–433. Дои:10.1128 / CMR.17.2.413-433.2004. ЧВК 387402. PMID 15084508.
- ^ Видьясагар, Апарна (апрель 2016 г.). "Что такое амеба?". livescience.com. Получено 8 ноября 2020.
- ^ Тевес, Саша; Солдати, Тьерри; Эйхингер, Людвиг (2019). "От редакции: амебы как модели-хозяева для изучения взаимодействия с патогенами". Границы клеточной и инфекционной микробиологии. 9: 47. Дои:10.3389 / fcimb.2019.00047. ЧВК 6433779. PMID 30941316.
- ^ Хан Н.А., Сиддики Р. (2015). «Есть ли доказательства полового размножения (мейоза) у акантамебы?». Pathog Glob Health. 109 (4): 193–5. Дои:10.1179 / 2047773215Y.0000000009. ЧВК 4530557. PMID 25800982.
- ^ а б c d Келсо А.А., Сэй А.Ф., Шарма Д., Ледфорд Л.Л., Турчик А., Саски К.А., Кинг А.В., Аттавей С.К., Темесвари Л.А., Сехорн М.Г. (2015). «Entamoeba histolytica Dmc1 катализирует гомологичное спаривание ДНК и обмен цепей, который стимулируется кальцием и Hop2-Mnd1». PLOS ONE. 10 (9): e0139399. Bibcode:2015PLoSO..1039399K. Дои:10.1371 / journal.pone.0139399. ЧВК 4589404. PMID 26422142.
- ^ а б Сингх Н., Бхаттачарья А., Бхаттачарья С. (2013). «Гомологичная рекомбинация происходит у Entamoeba и усиливается во время стресса роста и конверсии стадий». PLOS ONE. 8 (9): e74465. Bibcode:2013PLoSO ... 874465S. Дои:10.1371 / journal.pone.0074465. ЧВК 3787063. PMID 24098652.
- ^ Флауэрс Дж. М., Ли С. И., Статос А., Саксер Дж., Островски Е. А., Квеллер Д. К., Штрассманн Дж. Е., Пуругганан М. Д. (2010). «Вариации, пол и социальное сотрудничество: молекулярная популяционная генетика социальной амебы Dictyostelium discoideum». PLOS Genet. 6 (7): e1001013. Дои:10.1371 / journal.pgen.1001013. ЧВК 2895654. PMID 20617172.
- ^ О'Дей Д.Х., Кесей А. (2012). «Сигнализация и секс у социальных амеб». Биол Рев Камб Филос Соц. 87 (2): 313–29. Дои:10.1111 / j.1469-185X.2011.00200.x. PMID 21929567. S2CID 205599638.
- ^ Лар DJ, Парфри Л.В., Митчелл Е.А., Кац Л.А., Лара Е. (2011). «Целомудрие амеб: переоценка доказательств секса у амебоидных организмов». Proc. Биол. Наука. 278 (1715): 2081–90. Дои:10.1098 / rspb.2011.0289. ЧВК 3107637. PMID 21429931.
дальнейшее чтение
- Валочник Дж. И Аспек Х. (2007). Amöben: Paradebeispiele für Probleme der Phylogenetik, Klassifikation und Nomenklatur. Денис 20: 323–350. (На немецком)
- Амебы: протисты, которые передвигаются и питаются псевдоподиями в веб-проекте "Древо жизни"
- Павловски Дж. И Бурки Ф. (2009). Распутывая филогению амебоидных протистов. Журнал эукариотической микробиологии 56.1: 16–25.
внешняя ссылка
- Сименсма, Ф. Микромир: мир амебоидных организмов.
- Фёлькер, Э. и Клаус, С. Визуальный ключ к амебоидным морфотипам. Penard Labs.
- Амебы веб-сайт лаборатории Макивера Эдинбургского университета, объединяет информацию из опубликованных источников.
- Цифровая видеогалерея молекулярных выражений: Жизнь в пруду - Амеба (простейшие) - информативные видео амебы