Парамеций - Paramecium
| Парамеций | |
|---|---|
 | |
| Парамеций аурелия | |
| Научная классификация | |
| Clade: | SAR |
| Infrakingdom: | Альвеолаты |
| Тип: | Цилиофора |
| Класс: | Олигогименофорея |
| Заказ: | Peniculida |
| Семья: | Parameciidae |
| Род: | Парамеций Мюллер, 1773 |
| Разновидность | |
См. Текст | |
| Синонимы | |
| |
Парамеций (также Парамеций, /ˌпærəˈмяʃ(я)əм/, PARR-ə-MEE-sh (ee-) əm, /-sяəм/, -См-м )[1] это род одноклеточный инфузории, обычно изучается как представитель группы инфузорий. Парамеция широко распространены в пресная вода, солоноватый, и морской среды и часто очень многочисленны в стоячих бассейнах и прудах. Поскольку некоторые виды легко культивируются и их легко заставить сопрягать и разделять, он широко использовался в классах и лабораториях для изучения биологические процессы. Его полезность как модельный организм заставил одного исследователя инфузорий охарактеризовать его как "белая крыса "типа Цилиофора.[2]
Историческое прошлое


Парамеция были одними из первых инфузорий, которых заметили микроскописты, в конце 17 века. Вероятно, они были известны голландскому пионеру протозоология, Антони ван Левенгук, и были ясно описаны его современником Кристиан Гюйгенс в письме 1678 г.[3] Самая ранняя из известных иллюстраций парамециума была анонимно опубликована в Философские труды Королевского общества, в 1703 г.[4]
В 1718 году французский учитель математики и микроскопист Луи Джобло опубликовал описание и иллюстрацию микроскопического Пуассон (рыба), которую он обнаружил в настой коры дуба в воде. Джоблот дал этому существу имя "Шоссон", или «тапочка», а фраза «тапочка анималкула» оставалась в употреблении как разговорный эпитет для Парамеций, на протяжении 18 и 19 веков.[5]
Название «Парамеций» - построено из Греческий παραμήκης (paramēkēs, "продолговатый") - придуман в 1752 году английским микроскопистом. Джон Хилл, который применил это название в целом к «Анималкулам, у которых нет видимых конечностей или хвостов, и они имеют неправильную продолговатую форму».[6] В 1773 г. О. Ф. Мюллер, первый исследователь, поместивший род в Линнеевский система таксономия принял название Парамеций, но изменил написание на Парамеций. К. Г. Эренберг, в крупном исследовании инфузория опубликованное в 1838 году, восстановило первоначальное написание Хилла названия рода, и большинство исследователей последовали его примеру.[7]
Описание
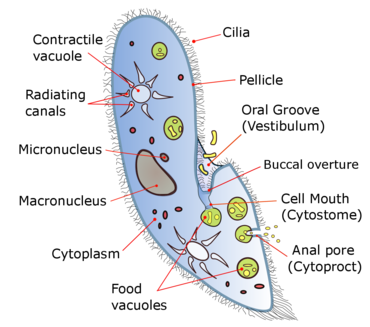
Виды Парамеций Диапазон размеров от 50 до 330 микрометров (от 0,0020 до 0,0130 дюйма) в длину. Клетки обычно имеют яйцевидную, удлиненную, форму стопы или сигару.
Тело клетки окружено жесткой, но эластичной структурой, называемой пленка. Он состоит из внешнего клеточная мембрана (плазматическая мембрана), слой уплощенных мембраносвязанных мешочков, называемых альвеолы, и внутренняя мембрана, называемая эпиплазма. Пленка не гладкая, а текстурированная с шестиугольными или прямоугольными углублениями. Каждый из этих многоугольников перфорирован центральным отверстием, через которое выступает единственная ресничка. Между альвеолярными мешочками пленки большинство видов Парамеций имеют близкорасположенные веретенообразные трихоцисты, взрывчатые органеллы, выделяющие тонкие нетоксичные волокна, часто используемые в оборонительных целях.[8][9]
Обычно анальная пора (цитопрокт) находится на вентральный поверхность, в задний половина клетки. У всех видов имеется глубокая оральная бороздка, идущая от передний ячейки до ее середины. Он покрыт незаметными ресничками, которые непрерывно бьются, втягивая пищу внутрь клетки.[10] Парамеция жить в основном гетеротрофия, питаясь бактерии и другие мелкие организмы. Несколько видов миксотрофы, получая некоторые питательные вещества из эндосимбиотический водоросли (хлорелла ) осуществляется в цитоплазма ячейки.[11]
Осморегуляция осуществляется сократительные вакуоли, которые активно вытесняют воду из клетки, чтобы компенсировать поглощение жидкости осмос из его окрестностей.[12] Количество сократительных вакуолей варьируется от одной до многих, в зависимости от вида.[10]
Движение
А Парамеций продвигается хлыстовыми движениями ресничек, которые расположены плотно расположенными рядами вокруг внешней части тела. Биение каждой реснички имеет две фазы: быстрый «эффективный ход», во время которого ресничка относительно жесткая, за которым следует медленный «ход восстановления», во время которого ресничка свободно изгибается в одну сторону и движется вперед против часовой стрелки. мода. Плотно расположенные реснички движутся скоординированным образом, волны активности движутся по «цилиарному ковру», создавая эффект, иногда напоминающий ветер, дующий через зерновое поле.[13]
В Парамеций движется по спирали в воде. Когда случается столкновение с препятствием, «эффективный ход» его ресничек меняется на противоположный, и организм на короткое время плывет назад, прежде чем возобновить свое продвижение вперед. Это называется реакция избегания. Если он снова сталкивается с твердым объектом, он повторяет этот процесс, пока не сможет пройти мимо объекта.[14]
Было подсчитано, что Парамеций тратит более половины своей энергии на продвижение по воде.[15] Эффективность этого цилиарного метода передвижения составляет менее 1%. Этот низкий процент, тем не менее, близок к максимальной теоретической эффективности, которая может быть достигнута организмом, оснащенным ресничками такой же длины, как и у представителей Парамеций.[16]
Сбор еды

Парамеция питаются микроорганизмами, такими как бактерии, водоросли и дрожжи. Чтобы собрать еду, Парамеций совершает движения с ресничками для захвата организмов-жертв вместе с небольшим количеством воды через ротовую борозду (преддверие или преддверие) в клетку. Пища проходит из ротовой бороздки, выстланной ресничками, в более узкую структуру, известную как ротовая полость (глотка). Оттуда частицы пищи проходят через небольшое отверстие, называемое цитостом, или устье клетки, и переместитесь внутрь клетки. Когда пища попадает в клетку, она собирается в пищу. вакуоли, которые периодически закрываются и выпускаются в цитоплазма, где они начинаются циркулирует по телу клетки. Пищевые вакуоли циркулируют за счет потокового движения содержимого клеток, этот процесс называется циклоз или цитоплазматический поток. По мере продвижения пищевой вакуоли, ферменты из цитоплазмы попасть в нее, чтобы переварить содержимое. По мере ферментативного пищеварения содержимое вакуолей становится более кислым. В течение пяти минут после образования вакуоли pH его содержимого снижается с 7 до 3.[17] Когда переваренные питательные вещества попадают в цитоплазму, вакуоль сжимается. Когда вакуоль с ее полностью переваренным содержимым достигает анальной поры, она разрывается, вытесняя отходы в окружающую среду за пределы клетки.[18][19][20]
Симбиоз
Некоторые виды Парамеций форма мутуалистический отношения с другими организмами. Paramecium bursaria и Paramecium chlorelligerum содержат эндосимбиотические зеленые водоросли, из которых получают питательные вещества и обладают определенной степенью защиты от хищников, таких как Didinium nasutum.[21][22] Многочисленные бактериальные эндосимбионты были идентифицированы у видов Парамеций.[23] Некоторые внутриклеточные бактерии, известные как Каппа-частицы, дать Парамеция которые обладают способностью убивать другие штаммы Парамеций что не хватает Каппы.[23]
Геном
В геном вида Paramecium tetraurelia был упорядочен, обеспечивая доказательство трех целыхдупликации генома.[24]
У некоторых инфузорий, например Стилонихия и Парамеций, только UGA декодируется как стоп-кодон, в то время как UAG и UAA переназначены как смысловые кодоны (то есть, когда стандартная аминокислота «кодируется» стоп-кодоном), кодирование аминокислота глютаминовая кислота.[25]
Учусь
Вопрос в том, Парамеция обучение экспонентам было предметом множества экспериментов, которые дали неоднозначные результаты. Однако исследование, опубликованное в 2006 году, похоже, показывает, что Paramecium caudatum можно обучить, применив 6.5 вольт электрический ток, чтобы различать уровни яркости.[26] Этот эксперимент был назван возможным примером клеточной памяти или эпигенетическое обучение в организмах без нервная система.[27] Однако другое исследование, проведенное в 2017 году, показало, что Парамеция может только научиться связывать яркую сторону своей плавательной среды с электрическим током, а не темную сторону.[28] В том же исследовании был предложен молекулярный механизм обучения в Парамеция.[29][28]
Размножение и сексуальные явления
Как и все инфузории, Парамеций имеет двойную ядерный аппарат, состоящий из полиплоид макронуклеус, и один или несколько диплоид микроядра. Макронуклеус контролирует нерепродуктивные функции клеток, выражая гены необходим для повседневного функционирования. Микроядро - генеративное, или зародышевый ядро, содержащее генетический материал, который передается от одного поколения к другому.[30]
Парамеций воспроизводит бесполым путем, к двойное деление. Во время размножения макронуклеус расщепляется по типу амитоз, и микроядра подвергаются митоз. Затем клетка делится поперечно, и каждая новая клетка получает копию микронуклеуса и макронуклеуса.[2]
Деление может происходить спонтанно, в ходе вегетативного клеточный цикл. При определенных условиях ему может предшествовать самооплодотворение (автогамия ),[31] или это может последовать спряжение, сексуальный феномен, при котором Парамеций совместимых типов спаривания временно сливаются и обмениваются генетическим материалом. Во время конъюгации микроядра каждого конъюганта делятся на мейоз и гаплоидный гаметы переходят из одной клетки в другую. Затем гаметы каждого организма сливаются, образуя диплоид микроядра. Старые макронуклеары разрушаются, а новые развиваются из новых микроядер.[30]
Автогамия или конъюгация могут быть вызваны нехваткой пищи в определенных точках тела. Парамеций жизненный цикл.[32]
Старение
В фазе роста бесполого деления, во время которой деление клеток происходит путем митоза, а не мейоза, происходит клональное старение, ведущее к постепенной потере жизнеспособности. У некоторых видов, например у хорошо изученных Paramecium tetraurelia, асексуальная линия клонального старения Парамеция теряет жизнеспособность и умирает примерно через 200 делений, если клетки не могут подвергнуться автогамии или конъюгации. Основа клонального старения была разъяснена трансплантация эксперименты Ауфдерхайда в 1986 году.[33] Когда макронуклеары клонально молодые Парамеция были введены в Парамеция стандартного клонального возраста продолжительность жизни (клональные деления) реципиента была продлена. Напротив, передача цитоплазма из клонально молодых Парамеция не продлил жизнь получателю. Эти эксперименты показали, что за клональное старение отвечает скорее макронуклеус, чем цитоплазма. Другие эксперименты Смита-Зоннеборна,[34] Холмс и Холмс,[35] и Гилли и Блэкберн[36] продемонстрировали, что во время клонального старения Повреждение ДНК резко увеличивается.[37] Таким образом, повреждение ДНК в макронуклеусе, по-видимому, является причиной старения у П. тетраурелия. В этом одноклеточном протист, старение, похоже, происходит так же, как и в многоклеточных эукариоты, как описано в Теория повреждений ДНК старения.
Мейоз и омоложение
При клональном возрасте П. тетраурелия стимулируются к мейозу в связи с конъюгацией или автомиксис, генетические потомки омолаживаются и могут иметь гораздо больше митотических бинарных делений. Во время любого из этих процессов микроядра клетки (ов) подвергаются мейозу, старое макронуклеус распадается, а новое макронуклеус формируется путем репликации микроядерной ДНК, которая недавно подверглась мейозу. Очевидно, что в новом макронуклеусе повреждения ДНК незначительны, если они вообще есть. Эти находки еще больше подтверждают, что клональное старение в значительной степени связано с прогрессирующим накоплением повреждений ДНК; и что омоложение происходит за счет восстановления этого повреждения микроядра во время мейоза. Мейоз, по-видимому, является адаптацией к восстановлению и омоложению ДНК в П. тетраурелия.[нужна цитата ]
Видео галерея
Paramecium bursaria, вид с симбиотическими водорослями
Paramecium putrinum
Парамеций двойное деление
Парамеций в спряжении
Paramecium caudatum


Список видов
Парамеций аурелия видовой комплекс:
- Paramecium primaurelia
- Paramecium biaurelia
- Paramecium triaurelia
- Paramecium tetraurelia
- Paramecium pentaurelia
- Paramecium sexaurelia
- Paramecium septaurelia
- Paramecium octaurelia
- Paramecium novaurelia
- Paramecium decaurelia
- Paramecium undecaurelia
- Парамеций додекаурелия
- Paramecium tredecaurelia
- Paramecium quadecaurelia
- Paramecium sonneborni
Другие виды:
- Paramecium buetschlii
- Paramecium bursaria
- Paramecium calkinsi
- Paramecium caudatum
- Paramecium chlorelligerum
- Paramecium duboscqui
- Paramecium grohmannae
- Paramecium jenningsi
- Paramecium multimicronucleatum
- Paramecium nephridiatum
- Paramecium polycaryum
- Paramecium putrinum
- Paramecium schewiakoffi
- Paramecium woodruffi
Рекомендации
- ^ "парамеций". Словарь Merriam-Webster.
- ^ а б Линн, Денис (2008). Ресничные простейшие: характеристика, классификация и руководство по литературе. Springer Science & Business Media. п. 279. ISBN 9781402082399.
- ^ Добелл, Клиффорд (1932). Энтони ван Левенгук и его «Зверюшки» (Изд. 1960). Нью-Йорк: Дувр. С. 164–165. ISBN 978-0-486-60594-4.
- ^ Долан, Джон Р. (1 августа 2019 г.). "Разоблачение" Старшего сына отца протозоологии ": Чарльз Кинг". Протист. 170 (4): 374–384. Дои:10.1016 / j.protis.2019.07.002. ISSN 1434-4610. PMID 31479910.
- ^ Джоблот, Луи (1718 г.). Описание и использование микроскопов Plusieurs Nouveaux, простое составление (На французском). 2. Париж: Жак Колломбат. п. 79.
- ^ Хилл, Джон (1752). История животных. Париж: Томас Осборн. п.5.
- ^ Вудрафф, Лоранд Лосс (сентябрь 1921 г.). «Структура, история жизни и внутриродовые отношения Paramecium calkinsi, sp. Nov». Биологический бюллетень. 41 (3): 171–180. Дои:10.2307/1536748. JSTOR 1536748.
- ^ Линн, Денис (2008). Ресничные простейшие: характеристика, классификация и руководство по литературе (3-е изд.). Springer Нидерланды. ISBN 9781402082382.
- ^ Вихтерман, Р. (2012-12-06). Биология Paramecium. Springer Science & Business Media. ISBN 9781475703726.
- ^ а б Творог, Colin R .; Гейтс, Майкл; Робертс, Дэвид МакЛ. (1983). Британские и другие пресноводные ресничные простейшие. 2. Издательство Кембриджского университета. п. 126.
- ^ Эстебан, Геновева Ф .; Фенчел, Том; Финли, Блэнд Дж. (2010). «Миксотрофия инфузорий». Протист. 161 (5): 621–641. Дои:10.1016 / j.protis.2010.08.002. PMID 20970377.
- ^ Рис, Джейн Б. (2011). Кэмпбелл Биология. Сан-Франциско: образование Пирсона. п.134. ISBN 9780321558237.
- ^ Блейк, Джон Р .; Сани, Майкл А. (февраль 1974 г.). «Механика цилиарного движения». Биологические обзоры. 49 (1): 85–125. Дои:10.1111 / j.1469-185x.1974.tb01299.x. PMID 4206625. S2CID 41907168.
- ^ Огура А. и К. Такахаши. «Искусственная децилиляция вызывает потерю зависимых от кальция ответов у Paramecium» (1976): 170–172.
- ^ Кацу-Кимура, Юмико; и другие. (2009). «Существенные затраты энергии на передвижение инфузорий подтверждены одновременным измерением скорости потребления кислорода и скорости плавания». Журнал экспериментальной биологии. 212 (12): 1819–1824. Дои:10.1242 / jeb.028894. PMID 19482999.
- ^ Остерман, Натан; Вильфан, Андрей (20 сентября 2011 г.). «Поиск модели биения ресничек с оптимальной эффективностью» (PDF). Труды Национальной академии наук. 108 (38): 15727–15732. arXiv:1107.4273. Bibcode:2011PNAS..10815727O. Дои:10.1073 / pnas.1107889108. ЧВК 3179098. PMID 21896741.
- ^ Вихтерман Р. (1986). Биология Paramecium. Springer США. С. 200–1. ISBN 9781475703740.
- ^ Вихтерман, Ральф (1985). Биология Paramecium. Нью-Йорк: Пленум Пресс. С. 88–90. ISBN 978-1-4757-0374-0.
- ^ Рис, Джейн Б.; и другие. (2011). Кэмпбелл Биология. Сан-Франциско: образование Пирсона. п.584. ISBN 9780321558237.
- ^ Маст, С. О. (февраль 1947 г.). «Пищевая вакуоль в Paramecium». Биологический бюллетень. 92 (1): 31–72. Дои:10.2307/1537967. JSTOR 1537967. PMID 20284992.
- ^ Бергер, Жак (1980). "Кормление Didinium nasutum на Paramecium bursaria с нормальными или апохлоротическими зоохлореллами ». Журнал общей микробиологии. 118 (2): 397–404. Дои:10.1099/00221287-118-2-397.
- ^ Крейц, Мартин; Стоук, Торстен; Фойсснер, Вильгельм (2012). «Морфологическая и молекулярная характеристика Парамеций (Виридопарамеций ноя подген.) хлорилгерум Каль (Цилиофора) ". Журнал эукариотической микробиологии. 59 (6): 548–563. Дои:10.1111 / j.1550-7408.2012.00638.x. ЧВК 3866650. PMID 22827482.
- ^ а б Преер, Джон Р. Младший; Преер, Луиза Б.; Джуранд, Артур (июнь 1974 г.). «Каппа и другие эндосимбионты в Парамеций аурелия". Бактериологические обзоры. 38 (2): 113–163. Дои:10.1128 / MMBR.38.2.113-163.1974. ЧВК 413848. PMID 4599970.
- ^ Ори, Жан-Марк; Джайон, Оливер; Винкер, Патрик; и другие. (Ноябрь 2006 г.). "Глобальные тенденции полногеномных дупликаций инфузорий Paramecium tetraurelia". Природа. 444 (7116): 171–8. Bibcode:2006Натура.444..171А. Дои:10.1038 / природа05230. PMID 17086204.
- ^ Лекомцев, Сергей; Колосов, Петр; Биду, Лауре; Фролова Людмила; Руссе, Жан-Пьер; Киселев, Лев (26 июня 2007 г.). «Различные способы ограничения стоп-кодонов Стилонихия и Парамеций Факторы завершения трансляции eRF1 ". Труды Национальной академии наук Соединенных Штатов Америки. 104 (26): 10824–9. Bibcode:2007ПНАС..10410824Л. Дои:10.1073 / pnas.0703887104. ЧВК 1904165. PMID 17573528.
- ^ Armus, Harvard L .; Montgomery, Amber R .; Джеллисон, Дженни Л. (осень 2006 г.). "Обучение дискриминации в парамеции (P. caudatum)". Психологический отчет. 56 (4): 489–498. Дои:10.1007 / BF03396029. S2CID 142785414.
- ^ Гинзбург, Симона; Яблонька, Ева (2009). «Эпигенетическое обучение у ненейронных организмов». Журнал биологических наук. 34 (4): 633–646. Дои:10.1007 / s12038-009-0081-8. PMID 19920348. S2CID 15507569.
- ^ а б Алипур, Абольфазл; Дорваш, Мохаммадреза; Еганех, Ясаман; Хатам, Голамреза (27 ноября 2017 г.). «Обучение парамециуму: новые идеи и модификации». Дои:10.1101/225250. Цитировать журнал требует
| журнал =(Помогите) - ^ Алипур, Абольфазл; Дорваш, Мохаммадреза; Еганех, Ясаман; Хатам, Голамреза; Серадж, Сейед Хасан (22 мая 2017 г.). «Возможные молекулярные механизмы обучения парамеций». Журнал перспективных медицинских наук и прикладных технологий. 3 (1): 39. Дои:10.18869 / nrip.jamsat.3.1.39. ISSN 2423-5903.
- ^ а б Прескотт, Д. М .; и другие. (1971). «ДНК мерцательных простейших». Хромосома. 34 (4): 355–366. Дои:10.1007 / bf00326311. S2CID 5013543.
- ^ Бергер, Джеймс Д. (октябрь 1986 г.). «Автогамия в Парамеций специфическая приверженность стадии клеточного цикла к мейозу ". Экспериментальные исследования клеток. 166 (2): 475–485. Дои:10.1016/0014-4827(86)90492-1. PMID 3743667.
- ^ Бил, Джеффри; Преер, Джон Р. младший (2008). Парамеций: генетика и эпигенетика. CRC. ISBN 9780203491904.
- ^ Ауфдерхайде, Карл Дж. (1986). "Клональное старение в Paramecium tetraurelia. II. Доказательства функциональных изменений макронуклеуса с возрастом ». Механизмы старения и развития. 37 (3): 265–279. Дои:10.1016/0047-6374(86)90044-8. PMID 3553762. S2CID 28320562.
- ^ Смит-Зоннеборн, Дж. (1979). «Восстановление ДНК и обеспечение долголетия Paramecium tetraurelia». Наука. 203 (4385): 1115–1117. Bibcode:1979Научный ... 203.1115С. Дои:10.1126 / science.424739. PMID 424739.
- ^ Холмс, Джордж Э .; Холмс, Норрин Р. (июль 1986 г.). «Накопление повреждений ДНК при старении. Paramecium tetraurelia". Молекулярная и общая генетика MGG. 204 (1): 108–114. Дои:10.1007 / bf00330196. PMID 3091993. S2CID 11992591.
- ^ Гилли, Дэвид; Блэкберн, Элизабет Х. (1994). "Отсутствие укорочения теломер во время старения у Парамеций" (PDF). Труды Национальной академии наук Соединенных Штатов Америки. 91 (5): 1955–1958. Bibcode:1994ПНАС ... 91.1955Г. Дои:10.1073 / пнас.91.5.1955. ЧВК 43283. PMID 8127914.
- ^ Bernstein, H; Бернштейн, C (1991). Старение, секс и восстановление ДНК. Сан-Диего: Academic Press. С. 153–156. ISBN 978-0120928606.