Генетический код - Википедия - Genetic code

В генетический код свод правил, используемых живыми клетки к переведите информация, закодированная в генетическом материале (ДНК или же мРНК последовательности нуклеотидных триплетов, или кодоны) в белки. Перевод осуществляется рибосома, который связывает протеиногенные аминокислоты в порядке, указанном информационная РНК (мРНК), используя переносить РНК (тРНК) для переноса аминокислот и чтения мРНК три нуклеотиды вовремя. Генетический код всех организмов очень похож и может быть выражен в простой таблице с 64 записями.[1]
Код определяет, как кодоны определяют, какая аминокислота будет добавлена следующей во время синтез белка. За некоторыми исключениями,[2] трехнуклеотидный кодон в последовательности нуклеиновой кислоты определяет единственную аминокислоту. Подавляющее большинство гены закодированы по единой схеме (см. Таблица кодонов РНК ). Эту схему часто называют каноническим или стандартным генетическим кодом или просто то генетический код, хотя коды вариантов (например, в человеческом митохондрии ) существовать.
В то время как "генетический код" определяет аминокислотную последовательность белка, другие области генома определяют когда и где производятся эти белки согласно различным "кодам регулирования генов".
История
Попытки понять, как кодируются белки, начались после открытия структуры ДНК в 1953 году. Георгий Гамов постулировал, что наборы из трех оснований должны использоваться для кодирования 20 стандартных аминокислот, используемых живыми клетками для построения белков, что позволило бы использовать максимум 43 = 64 аминокислоты.[3]
Кодоны
В Эксперимент Крика, Бреннера, Барнетта и Уоттса-Тобина впервые продемонстрировал, что кодоны состоят из трех оснований ДНК. Маршалл Ниренберг и Генрих Дж. Маттеи первыми раскрыли природу кодона в 1961 году.[4]
Они использовали бесклеточная система к переведите поли-урацил Последовательность РНК (т.е. UUUUU ...) и обнаружил, что полипептид что они синтезировали, состояла только из аминокислоты фенилаланин.[5] Таким образом, они пришли к выводу, что кодон UUU определяет аминокислоту фенилаланин.
Затем последовали эксперименты в Северо-Очоа лаборатория, которая продемонстрировала, что поли-аденин Последовательность РНК (AAAAA ...), кодирующая полипептидный поли-лизин[6] и что поли-цитозин Последовательность РНК (CCCCC ...), кодируемая для полипептидного поли-пролин.[7] Следовательно, кодон AAA определяет аминокислоту лизин, а кодон CCC указывает аминокислоту пролин. Используя различные сополимеры Затем определяли большинство оставшихся кодонов.
Последующая работа Хар Гобинд Кхорана идентифицировали остальную часть генетического кода. Вскоре после этого, Роберт В. Холли определил структуру переносить РНК (тРНК), адаптерная молекула, которая облегчает процесс трансляции РНК в белок. Эта работа была основана на более ранних исследованиях Очоа, в результате чего последнее Нобелевская премия по физиологии и медицине в 1959 г. для работы над энзимология синтеза РНК.[8]
Продолжая эту работу, Ниренберг и Филип Ледер раскрыл триплетную природу кода и расшифровал его кодоны. В этих экспериментах различные комбинации мРНК были пропущены через фильтр, содержащий рибосомы, компоненты ячеек, переведите РНК в белок. Уникальные триплеты способствовали связыванию специфических тРНК с рибосомой. Ледер и Ниренберг смогли определить в своих экспериментах последовательности 54 из 64 кодонов.[9] Хорана, Холли и Ниренберг получили Нобелевскую премию 1968 года за свою работу.[10]
Три стоп-кодона были названы первооткрывателями Ричардом Эпштейном и Чарльзом Стейнбергом. «Янтарь» был назван в честь их друга Харриса Бернстайна, фамилия которого в переводе с немецкого означает «янтарь».[11] Два других стоп-кодона были названы «охра» и «опал», чтобы сохранить тему «названий цветов».
Расширенные генетические коды (синтетическая биология)
В широкой академической аудитории широко принята концепция эволюции генетического кода от исходного и неоднозначного генетического кода до четко определенного («замороженного») кода с репертуаром из 20 (+2) канонических аминокислот.[12]Однако есть разные мнения, концепции, подходы и идеи, что лучше всего изменить экспериментально. Предлагаются даже модели, которые предсказывают «точки входа» для вторжения синтетических аминокислот в генетический код.[13]
С 2001 года 40 неприродных аминокислот были добавлены в белок путем создания уникального кодона (перекодирования) и соответствующей пары трансфер-РНК: аминоацил - тРНК-синтетаза, чтобы кодировать его с различными физико-химическими и биологическими свойствами, чтобы использовать его в качестве инструмент для изучения структура белка и функционировать или создавать новые или улучшенные белки.[14][15]
Х. Мураками и М. Сисидо расширили некоторые кодоны до четырех и пяти оснований. Стивен А. Беннер построил функционал 65-й (in vivo) кодон.[16]
2015 г. Н. Будиса, Д. Зёлль и коллеги сообщили о полной замене всех 20 899 триптофан остатки (кодоны UGG) с неестественным тиенопирролаланином в генетическом коде бактерии кишечная палочка.[17]
В 2016 году был создан первый стабильный полусинтетический организм. Это была (одноклеточная) бактерия с двумя синтетическими основаниями (называемыми X и Y). Основания пережили деление клеток.[18][19]
В 2017 году исследователи из Южной Кореи сообщили, что они создали мышь с расширенным генетическим кодом, которая может производить белки с неестественными аминокислотами.[20]
В мае 2019 года исследователи объявили о создании нового синтетический (возможно искусственный ) форма жизнеспособный жизнь, вариант бактерии кишечная палочка, уменьшив натуральное число 64 кодоны в бактериальном геном вместо 59 кодонов, чтобы кодировать 20 аминокислоты.[21][22]
Функции
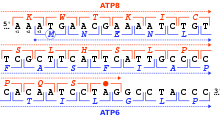
Рамка для чтения
Рамка считывания определяется начальным триплетом нуклеотидов, с которого начинается трансляция. Он устанавливает рамку для серии последовательных, неперекрывающихся кодонов, которая известна как "открытая рамка чтения "(ORF). Например, строка 5'-AAATGAACG-3 '(см. Рисунок) при чтении с первой позиции содержит кодоны AAA, TGA и ACG; при чтении со второй позиции она содержит кодоны AAT и GAA; и если читать с третьей позиции, он содержит кодоны ATG и AAC. Таким образом, каждая последовательность может быть прочитана в своем Направление 5 '→ 3' через три рамки чтения, каждая из которых дает, возможно, отдельную аминокислотную последовательность: в данном примере Lys (K) -Trp (W) -Thr (T), Asn (N) -Glu (E) или Met (M) -Asn (N) соответственно (при трансляции с кодом митохондрий позвоночных). Когда ДНК двухцепочечная, возможны шесть рамки чтения определены: три в прямой ориентации на одной нити и три в обратной ориентации на противоположной нити.[24]:330 Кадры, кодирующие белок, определяются стартовый кодон, обычно это первый кодон AUG (ATG) в последовательности РНК (ДНК).
В эукариоты, ORF в экзоны часто прерываются интроны.
Старт / стоп кодоны
Трансляция начинается с кодона инициации цепи или стартовый кодон. Одного стартового кодона недостаточно для начала процесса. Соседние последовательности, такие как Шайн-Дальгарно последовательность в Кишечная палочка и факторы инициирования также необходимы для начала перевода. Наиболее распространенным стартовым кодоном является AUG, который читается как метионин или, у бактерий, как формилметионин. Альтернативные стартовые кодоны в зависимости от организма включают «GUG» или «UUG»; эти кодоны обычно представляют валин и лейцин соответственно, но как стартовые кодоны они переводятся как метионин или формилметионин.[25]
Три стоп-кодоны есть имена: UAG это Янтарь, UGA - это опал (иногда также называют умбра), а UAA - это охра. Стоп-кодоны также называют «терминирующими» или «бессмысленными» кодонами. Они сигнализируют о высвобождении растущего полипептида из рибосомы, потому что ни одна родственная тРНК не имеет антикодонов, комплементарных этим стоп-сигналам, что позволяет фактор выпуска вместо этого связываться с рибосомой.[26]
Эффект мутаций

В процессе Репликация ДНК, иногда возникают ошибки в полимеризация второй пряди. Эти ошибки, мутации, может повлиять на фенотип, особенно если они встречаются в кодирующей белок последовательности гена. Частота ошибок обычно составляет 1 ошибку на каждые 10–100 миллионов базисов - из-за возможности «корректуры» ДНК-полимеразы.[28][29]
Миссенс-мутации и бессмысленные мутации являются примерами точечные мутации которые могут вызвать генетические заболевания, такие как серповидноклеточная анемия и талассемия соответственно.[30][31][32] Клинически важные миссенс-мутации обычно изменяют свойства кодируемого аминокислотного остатка в основном, кислотном, полярном или неполярном состояниях, тогда как бессмысленные мутации приводят к стоп-кодон.[24]
Мутации, нарушающие последовательность рамки считывания за счет инделы (вставки или же удаления ) нескольких нуклеотидных оснований, не кратных 3, известны как мутации сдвига рамки считывания. Эти мутации обычно приводят к полностью отличному от оригинала переводу и, вероятно, вызывают стоп-кодон to be read, что обрезает белок.[33] Эти мутации могут нарушать функцию белка и поэтому редко встречаются у in vivo белок-кодирующие последовательности. Одна из причин, по которой наследование мутаций сдвига рамки считывания является редким, заключается в том, что, если транслируемый белок необходим для роста под давлением отбора, с которым сталкивается организм, отсутствие функционального белка может привести к смерти до того, как организм станет жизнеспособным.[34] Мутации сдвига рамки считывания могут привести к тяжелым генетическим заболеваниям, таким как Болезнь Тея – Сакса.[35]
Хотя большинство мутаций, изменяющих белковые последовательности, вредны или нейтральны, некоторые мутации имеют преимущества.[36] Эти мутации могут позволить мутантному организму противостоять определенным стрессам окружающей среды лучше, чем дикого типа организмов или размножаются быстрее. В этих случаях мутация, как правило, становится более распространенной в популяции через естественный отбор.[37] Вирусы это использование РНК поскольку их генетический материал имеет высокую скорость мутации,[38] что может быть преимуществом, поскольку эти вирусы, таким образом, быстро развиваются и, таким образом, избегают иммунная система защитные реакции.[39] В больших популяциях организмов, размножающихся бесполым путем, например, Кишечная палочкамогут одновременно происходить множественные полезные мутации. Это явление называется клональное вмешательство и вызывает конкуренцию между мутациями.[40]
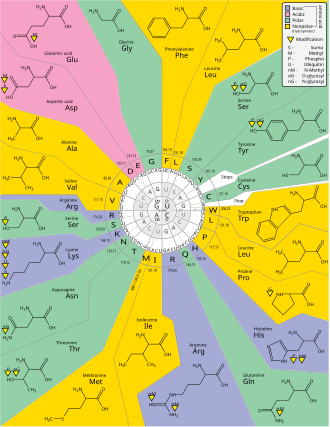
Вырождение

{kind=link}
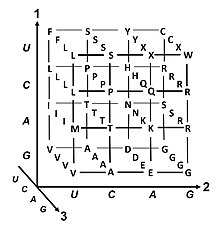
Вырождение - это избыточность генетического кода. Этот термин был дан Бернфилдом и Ниренбергом. Генетический код имеет избыточность, но не неоднозначен (см. таблицы кодонов ниже для полной корреляции). Например, хотя кодоны GAA и GAG определяют глютаминовая кислота (избыточность), ни одна из них не указывает другую аминокислоту (нет двусмысленности). Кодоны, кодирующие одну аминокислоту, могут различаться в любом из трех положений. Например, аминокислота лейцин определяется как YUр или CUN (UUA, UUG, CUU, CUC, CUA или CUG) кодоны (разница в первой или третьей позиции указывается с помощью Обозначение ИЮПАК ), а аминокислота серин указывается UCN или AGY (UCA, UCG, UCC, UCU, AGU или AGC) кодоны (разница в первом, втором или третьем положении).[41] Практическим следствием избыточности является то, что ошибки в третьей позиции триплетного кодона вызывают только молчащую мутацию или ошибку, которая не повлияет на белок, поскольку гидрофильность или же гидрофобность поддерживается эквивалентной заменой аминокислот; например, кодон NUN (где N = любой нуклеотид) имеет тенденцию кодировать гидрофобные аминокислоты. NCN дает аминокислотные остатки небольшого размера и умеренного размера. водность; NAN кодирует гидрофильные остатки среднего размера. Генетический код настолько хорошо структурирован с точки зрения гидропатичности, что математический анализ (Разложение по сингулярным значениям ) 12 переменных (4 нуклеотида х 3 положения) дает замечательную корреляцию (C = 0,95) для предсказания гидропатичности кодируемой аминокислоты непосредственно из триплетной нуклеотидной последовательности, без перевода.[42][43] Обратите внимание на приведенную ниже таблицу, на восемь аминокислот совсем не влияют мутации в третьем положении кодона, тогда как на рисунке выше мутация во втором положении может вызвать радикальное изменение физико-химических свойств кодона. кодируемая аминокислота. Тем не менее, изменения в первом положении кодонов более важны, чем изменения во втором положении в глобальном масштабе.[44] Причина может заключаться в том, что изменение заряда (с положительного на отрицательный или наоборот) может происходить только при мутациях в первом положении определенных кодонов, но не при изменении второго положения любого кодона. Такое изменение заряда может иметь драматические последствия для структуры или функции белка. Предыдущие исследования могли в значительной степени недооценить этот аспект.[44]
Предвзятость использования кодонов
Частота кодонов, также известная как систематическая ошибка использования кодонов, может варьироваться от вида к виду с функциональными последствиями для контроля перевод.
Таблица частот кодонов генома человека | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
Альтернативные генетические коды
Нестандартные аминокислоты
В некоторых белках нестандартные аминокислоты заменяются стандартными стоп-кодонами в зависимости от ассоциированных сигнальных последовательностей в матричной РНК. Например, UGA может кодировать для селеноцистеин и UAG может кодировать для пирролизин. Селеноцистеин стал считаться 21-й аминокислотой, а пирролизин - 22-й.[46] В отличие от селеноцистеина, UAG, кодируемый пирролизином, транслируется при участии специального аминоацил-тРНК синтетаза.[47] И селеноцистеин, и пирролизин могут присутствовать в одном организме.[46] Хотя генетический код обычно фиксируется в организме, ахеальный прокариот Acetohalobium arabaticum может расширить свой генетический код с 20 до 21 аминокислоты (включая пирролизин) в различных условиях роста.[48]
Вариации
Варианты стандартного кода были предсказаны в 1970-х годах.[49] Первый был открыт в 1979 году исследователями, изучавшими митохондриальные гены человека.[50] После этого было обнаружено множество небольших вариантов,[51] включая различные альтернативные митохондриальные коды.[52] Эти второстепенные варианты, например, включают перевод кодона UGA как триптофан в Микоплазма видов, и трансляция CUG в виде серина, а не лейцина у дрожжей «клады CTG» (таких как грибковые микроорганизмы албиканс ).[53][54][55] Поскольку вирусы должны использовать тот же генетический код, что и их хозяева, модификации стандартного генетического кода могут мешать синтезу или функционированию вирусного белка. Однако вирусы, такие как тивирусы адаптировались к модификации генетического кода хозяина.[56] В бактерии и археи, GUG и UUG являются общими стартовыми кодонами. В редких случаях некоторые белки могут использовать альтернативные стартовые кодоны.[51]Удивительно, но вариации в интерпретации генетического кода существуют также в генах, кодируемых ядерным ядром человека: в 2016 году исследователи, изучающие трансляцию малатдегидрогеназы, обнаружили, что примерно в 4% мРНК, кодирующих этот фермент, стоп-кодон естественным образом используется для кодирования аминокислоты триптофан и аргинин.[57] Этот тип перекодирования вызван контекстом стоп-кодона с высокой степенью считывания.[58] и упоминается как функциональное переводческое чтение.[59]
Варианты генетических кодов, используемых организмом, могут быть выведены путем идентификации высококонсервативных генов, кодируемых в этом геноме, и сравнения использования его кодонов с аминокислотами в гомологичных белках других организмов. Например, программа FACIL[60] выводит генетический код, ища, какие аминокислоты в доменах гомологичных белков чаще всего совпадают с каждым кодоном. Полученные вероятности аминокислот для каждого кодона отображаются в логотипе генетического кода, который также показывает поддержку стоп-кодона.
Несмотря на эти различия, все известные природные коды очень похожи. Механизм кодирования одинаков для всех организмов: трехосновные кодоны, тРНК, рибосомы, чтение в одном направлении и перевод отдельных кодонов в отдельные аминокислоты.[61] Самые крайние вариации встречаются у некоторых инфузорий, где значение стоп-кодонов зависит от их положения в мРНК. Находясь близко к 3 ’концу, они действуют как терминаторы, а во внутренних положениях они либо кодируют аминокислоты, как в Кондилостома магнум[62] или триггер рибосомальный сдвиг рамки как в Euplotes.[63]
Источник
Генетический код - ключевая часть история жизни, согласно одной из версий, самовоспроизводящиеся молекулы РНК предшествовали жизни, какой мы ее знаем. Это Гипотеза мира РНК. Согласно этой гипотезе, любая модель возникновения генетического кода тесно связана с моделью передачи от рибозимы (Ферменты РНК) к белкам как основным ферментам в клетках. В соответствии с гипотезой мира РНК, молекулы транспортной РНК, по-видимому, эволюционировали раньше, чем современные аминоацил-тРНК синтетазы, поэтому последний не может быть частью объяснения его закономерностей.[64]
Гипотетический случайно возникший генетический код дополнительно мотивирует биохимическую или эволюционную модель его происхождения. Если бы аминокислоты были случайным образом распределены по триплетным кодонам, получилось бы 1,5 × 1084 возможные генетические коды.[65]:163 Это число определяется путем расчета количества способов, которыми 21 элемент (20 аминокислот плюс одна остановка) может быть помещен в 64 ячейки, при этом каждый элемент используется хотя бы один раз.[66] Однако распределение кодонов в генетическом коде неслучайно.[67] В частности, генетический код группирует определенные назначения аминокислот.
Аминокислоты, которые участвуют в одном и том же пути биосинтеза, обычно имеют одно и то же первое основание в своих кодонах. Это могло быть эволюционным реликтом раннего, более простого генетического кода с меньшим количеством аминокислот, который позже эволюционировал, чтобы кодировать больший набор аминокислот.[68] Он также может отражать стерические и химические свойства, которые оказали другое влияние на кодон во время его эволюции. Аминокислоты с аналогичными физическими свойствами также имеют похожие кодоны,[69][70] уменьшение проблем, вызванных точечными мутациями и ошибочными переводами.[67]
Учитывая неслучайную схему генетического триплетного кодирования, разумная гипотеза происхождения генетического кода могла бы касаться нескольких аспектов таблицы кодонов, таких как отсутствие кодонов для D-аминокислот, вторичные образцы кодонов для некоторых аминокислот, ограничение синонимичных положения к третьему положению, небольшой набор только из 20 аминокислот (вместо числа, приближающегося к 64), и отношение паттернов стоп-кодонов к паттернам кодирования аминокислот.[71]
Три основные гипотезы касаются происхождения генетического кода. Многие модели относятся к одному из них или к гибриду:[72]
- Случайное замораживание: генетический код был создан случайным образом. Например, раннее тРНК -подобные рибозимы могли иметь различное сродство к аминокислотам, при этом кодоны, возникающие из другой части рибозима, демонстрировали случайную изменчивость. Один раз достаточно пептиды были закодированы, любое серьезное случайное изменение генетического кода было бы летальным; следовательно, он стал «замороженным».[73]
- Стереохимическое сродство: генетический код является результатом высокого сродства между каждой аминокислотой и ее кодоном или антикодоном; последний вариант подразумевает, что молекулы пре-тРНК соответствовали своим соответствующим аминокислотам по этому сродству. Позже в ходе эволюции это совпадение было постепенно заменено совпадением аминоацил-тРНК синтетаз.[71][74][75]
- Оптимальность: генетический код продолжал развиваться после своего первоначального создания, так что текущий код максимизирует некоторые фитнес функция, обычно какая-то минимизация ошибок.[71][72]
Гипотезы касались множества сценариев:[76]
- Химические принципы регулируют специфическое взаимодействие РНК с аминокислотами. Эксперименты с аптамеры показали, что некоторые аминокислоты обладают селективным химическим сродством к своим кодонам.[77] Эксперименты показали, что из 8 протестированных аминокислот 6 демонстрируют некоторую ассоциацию триплет-аминокислота РНК.[65][75]
- Биосинтетическое расширение. Генетический код вырос из более простого более раннего кода в процессе «биосинтетического расширения». Первобытная жизнь «открыла» новые аминокислоты (например, как побочные продукты метаболизм ), а позже включил некоторые из них в механизм генетического кодирования.[78] Хотя было найдено много косвенных доказательств того, что в прошлом использовалось меньшее количество типов аминокислот,[79] точные и подробные гипотезы о том, какие аминокислоты и в каком порядке вошли в код, противоречивы.[80][81] Однако несколько исследований показали, что Gly, Ala, Asp, Val, Ser, Pro, Glu, Leu, Thr могут принадлежать к группе аминокислот раннего присоединения, тогда как Cys, Met, Tyr, Trp, His, Phe могут принадлежать к группе аминокислот, добавляемых позже. [82][83][84][85]
- Естественный отбор привел к присвоению кодонов генетического кода, что минимизирует влияние мутации.[86] Недавняя гипотеза[87] предполагает, что триплетный код был получен из кодов, в которых использовались более длинные, чем триплетные кодоны (например, квадруплетные кодоны). Декодирование длиннее, чем триплет, увеличит избыточность кодонов и будет более устойчивым к ошибкам. Эта функция может позволить точное декодирование при отсутствии сложных механизмов перевода, таких как рибосома, например, до того, как клетки начали производить рибосомы.
- Информационные каналы: Информационно-теоретический подходы моделируют процесс трансляции генетического кода в соответствующие аминокислоты как подверженный ошибкам информационный канал.[88] Собственный шум (то есть ошибка) в канале ставит организм перед фундаментальным вопросом: как можно сконструировать генетический код, чтобы противостоять шуму?[89] при этом точно и качественно переводя информацию? Эти "скорость-искажение" модели[90] предполагают, что генетический код возник в результате взаимодействия трех конфликтующих эволюционных сил: потребности в различных аминокислотах,[91] на устойчивость к ошибкам[86] и при минимальных ресурсных затратах. Код возникает при переходе, когда отображение кодонов в аминокислоты становится неслучайным. Появление кода регулируется топология определяется вероятными ошибками и относится к проблема раскраски карты.[92]
- Теория игр: модели на основе сигнальные игры объединить элементы теории игр, естественного отбора и информационных каналов. Такие модели использовались, чтобы предположить, что первые полипептиды, вероятно, были короткими и имели неферментативную функцию. Теоретико-игровые модели предполагают, что организация цепочек РНК в клетках могла быть необходима для предотвращения «обманного» использования генетического кода, то есть предотвращения распространения древнего эквивалента вирусов в мире РНК.[93]
- Стоп-кодоны: кодоны для остановки трансляции также представляют собой интересный аспект проблемы происхождения генетического кода. В качестве примера решения проблемы эволюции стоп-кодонов было высказано предположение, что стоп-кодоны таковы, что они с наибольшей вероятностью прервут трансляцию на ранней стадии в случае сдвиг кадра ошибка.[94] Напротив, некоторые стереохимические молекулярные модели объясняют происхождение стоп-кодонов как «нераспознаваемое».[71]
Утверждалось, что генетический код содержит закономерности и арифметические совпадения, которые очень маловероятны случайно и не возникнут в процессе эволюции. Авторы этого утверждения утверждают, что это в основном сообщение, указывающее на то, что жизнь на Земле была засеяна предыдущей цивилизацией, похожей на панспермия.[95][96]
Смотрите также
Рекомендации
- ^ Шу, Цзянь-Цзюнь (2017). «Новая интегрированная симметричная таблица генетических кодов». Биосистемы. 151: 21–26. arXiv:1703.03787. Bibcode:2017arXiv170303787S. Дои:10.1016 / j.biosystems.2016.11.004. PMID 27887904. S2CID 1121152.
- ^ Туранов А.А., Лобанов А.В., Фоменко Д.Е., Моррисон Х.Г., Согин М.Л., Клобутчер Л.А., Хэтфилд Д.Л., Гладышев В.Н. (январь 2009 г.). «Генетический код поддерживает целенаправленную вставку двух аминокислот одним кодоном». Наука. 323 (5911): 259–61. Дои:10.1126 / science.1164748. ЧВК 3088105. PMID 19131629.
- ^ Крик, Фрэнсис (10 июля 1990 г.). «Глава 8: Генетический код». Какое безумное стремление: личный взгляд на научные открытия. Основные книги. С. 89–101. ISBN 978-0-465-09138-6.
- ^ Янофски, Чарльз (9 марта 2007 г.). «Установление триплетной природы генетического кода». Клетка. 128 (5): 815–818. Дои:10.1016 / j.cell.2007.02.029. Получено 9 октября 2020.
- ^ Ниренберг MW, Matthaei JH (октябрь 1961 г.). «Зависимость внеклеточного синтеза белка в E. coli от природных или синтетических полирибонуклеотидов». Труды Национальной академии наук Соединенных Штатов Америки. 47 (10): 1588–602. Bibcode:1961ПНАС ... 47.1588Н. Дои:10.1073 / pnas.47.10.1588. ЧВК 223178. PMID 14479932.
- ^ Гарднер Р.С., Вахба А.Дж., Базилио С., Миллер Р.С., Лендьель П., Шпейер Дж.Ф. (декабрь 1962 г.). «Синтетические полинуклеотиды и аминокислотный код. VII». Труды Национальной академии наук Соединенных Штатов Америки. 48 (12): 2087–94. Bibcode:1962ПНАС ... 48.2087Г. Дои:10.1073 / пнас.48.12.2087. ЧВК 221128. PMID 13946552.
- ^ Вахба А.Дж., Гарднер Р.С., Базилио С., Миллер Р.С., Шпейер Дж. Ф., Лендьель П. (январь 1963 г.). «Синтетические полинуклеотиды и код аминокислот. VIII». Труды Национальной академии наук Соединенных Штатов Америки. 49 (1): 116–22. Bibcode:1963ПНАС ... 49..116Вт. Дои:10.1073 / pnas.49.1.116. ЧВК 300638. PMID 13998282.
- ^ "Нобелевская премия по физиологии и медицине 1959 г." (Пресс-релиз). Шведская королевская академия наук. 1959 г.. Получено 27 февраля 2010.
Нобелевская премия по физиологии и медицине 1959 г. была присуждена совместно Северо Очоа и Артуру Корнбергу «за открытие механизмов биологического синтеза рибонуклеиновой кислоты и дезоксирибонуклеиновой кислоты».
- ^ Ниренберг М., Ледер П., Бернфилд М., Бримакомб Р., Трупин Дж., Роттман Ф., О'Нил С. (май 1965 г.). «Кодовые слова РНК и синтез белка, VII. Об общей природе кода РНК». Труды Национальной академии наук Соединенных Штатов Америки. 53 (5): 1161–8. Bibcode:1965ПНАС ... 53.1161Н. Дои:10.1073 / pnas.53.5.1161. ЧВК 301388. PMID 5330357.
- ^ "Нобелевская премия по физиологии и медицине 1968 г." (Пресс-релиз). Шведская королевская академия наук. 1968 г.. Получено 27 февраля 2010.
Нобелевская премия по физиологии и медицине 1968 г. была присуждена совместно Роберту У. Холли, Хар Гобинд Хорана и Маршаллу У. Ниренбергу «за их интерпретацию генетического кода и его функции в синтезе белка».
- ^ Эдгар Б. (октябрь 2004 г.). «Геном бактериофага Т4: археологические раскопки». Генетика. 168 (2): 575–82. ЧВК 1448817. PMID 15514035.
- ^ Будиса, Недилько (23 декабря 2005 г.). Книга в онлайн-библиотеке Wiley. Дои:10.1002/3527607188. ISBN 9783527312436.
- ^ Кубышкин, В .; Будиса, Н. (2017). «Синтетическое отчуждение микробных организмов с помощью инженерии генного кода: почему и как?». Биотехнологический журнал. 12 (8): 1600097. Дои:10.1002 / biot.201600097. PMID 28671771.
- ^ Се Дж., Шульц П.Г. (декабрь 2005 г.). «Добавление аминокислот в генетический репертуар». Современное мнение в области химической биологии. 9 (6): 548–54. Дои:10.1016 / j.cbpa.2005.10.011. PMID 16260173.
- ^ Ван К., Пэрриш А. Р., Ван Л. (март 2009 г.). «Расширение генетического кода для биологических исследований». Химия и биология. 16 (3): 323–36. Дои:10.1016 / j.chembiol.2009.03.001. ЧВК 2696486. PMID 19318213.
- ^ Саймон М. (7 января 2005 г.). Новые вычисления: акцент на биоинформатике. Springer Science & Business Media. С. 105–106. ISBN 978-0-387-22046-8.
- ^ Hoesl, M. G .; Oehm, S .; Durkin, P .; Darmon, E .; Peil, L .; Aerni, H.-R .; Rappsilber, J .; Rinehart, J .; Leach, D .; Söll, D .; Будиса, Н. (2015). «Химическая эволюция бактериального протеома». Angewandte Chemie International Edition. 54 (34): 10030–10034. Дои:10.1002 / anie.201502868. ЧВК 4782924. PMID 26136259. NIHMSID: NIHMS711205
- ^ «Создан первый стабильный полусинтетический организм | KurzweilAI». www.kurzweilai.net. 3 февраля 2017 г.. Получено 9 февраля 2017.
- ^ Чжан Ю., Лэмб Б.М., Фельдман А.В., Чжоу А.Х., Лавернь Т., Ли Л., Ромесберг Ф.Э. (февраль 2017 г.). «Полусинтетический организм, созданный для стабильного расширения генетического алфавита». Труды Национальной академии наук Соединенных Штатов Америки. 114 (6): 1317–1322. Дои:10.1073 / pnas.1616443114. ЧВК 5307467. PMID 28115716.
- ^ Хан С., Ян А., Ли С., Ли Х.В., Пак CB, Park HS (февраль 2017 г.). «Расширение генетического кода Mus musculus». Nature Communications. 8: 14568. Bibcode:2017НатКо ... 814568H. Дои:10.1038 / ncomms14568. ЧВК 5321798. PMID 28220771.
- ^ Циммер, Карл (15 мая 2019 г.). «Ученые создали бактерии с синтетическим геномом. Является ли эта жизнь искусственной? - Вехой в синтетической биологии колонии E. coli процветают благодаря ДНК, созданной с нуля людьми, а не природой». Нью-Йорк Таймс. Получено 16 мая 2019.
- ^ Фреденс, Юлий; и другие. (15 мая 2019 г.). «Полный синтез Escherichia coli с перекодированным геномом». Природа. 569 (7757): 514–518. Bibcode:2019Натура.569..514F. Дои:10.1038 / с41586-019-1192-5. ЧВК 7039709. PMID 31092918. S2CID 205571025.
- ^ Homo sapiens митохондрия, полный геном. «Пересмотренная Кембриджская эталонная последовательность (rCRS): номер доступа NC_012920», Национальный центр биотехнологической информации. Проверено 27 декабря, 2017.
- ^ а б King RC, Mulligan P, Stansfield W (10 января 2013 г.). Словарь по генетике. ОУП США. п. 608. ISBN 978-0-19-976644-4.
- ^ Touriol C, Bornes S, Bonnal S, Audigier S, Prats H, Prats AC, Vagner S (2003). «Создание разнообразия изоформ белка путем альтернативной инициации трансляции в кодонах, не относящихся к AUG». Биология клетки. 95 (3–4): 169–78. Дои:10.1016 / S0248-4900 (03) 00033-9. PMID 12867081.
- ^ Малой С. (29 ноября 2003 г.). «Как бессмысленные мутации получили свои названия». Курс микробной генетики. Государственный университет Сан-Диего. Получено 10 марта 2010.
- ^ Ссылки на изображение находятся на странице Wikimedia Commons по адресу: Commons: File: Notable mutations.svg # Ссылки.
- ^ Гриффитс AJ, Миллер JH, Suzuki DT, Lewontin RC, et al., Ред. (2000). «Спонтанные мутации». Введение в генетический анализ (7-е изд.). Нью-Йорк: У. Х. Фриман. ISBN 978-0-7167-3520-5.
- ^ Фрайзингер Э., Гроллман А.П., Миллер Х., Кискер С. (апрель 2004 г.). «Поражение (не) толерантности раскрывает понимание верности репликации ДНК». Журнал EMBO. 23 (7): 1494–505. Дои:10.1038 / sj.emboj.7600158. ЧВК 391067. PMID 15057282.
- ^ (Буа 2006, п. 39)
- ^ Чанг Дж. К., Кан Ю. В. (июнь 1979 г.). "бета 0 талассемия, бессмысленная мутация у человека". Труды Национальной академии наук Соединенных Штатов Америки. 76 (6): 2886–9. Bibcode:1979PNAS ... 76.2886C. Дои:10.1073 / pnas.76.6.2886. ЧВК 383714. PMID 88735.
- ^ Boillée S, Vande Velde C, Cleveland DW (октябрь 2006 г.). «БАС: болезнь мотонейронов и их ненейронных соседей». Нейрон. 52 (1): 39–59. Дои:10.1016 / j.neuron.2006.09.018. PMID 17015226.
- ^ Исбрандт Д., Хопвуд Дж. Дж., Фон Фигура К., Петерс С. (1996). «Две новые мутации сдвига рамки считывания, вызывающие преждевременные стоп-кодоны у пациента с тяжелой формой синдрома Марото-Лами». Человеческая мутация. 7 (4): 361–3. Дои:10.1002 / (SICI) 1098-1004 (1996) 7: 4 <361 :: AID-HUMU12> 3.0.CO; 2-0. PMID 8723688.
- ^ Ворона Дж. Ф. (1993). «Что мы знаем о темпах спонтанных мутаций человека?». Экологический и молекулярный мутагенез. 21 (2): 122–9. Дои:10.1002 / em.2850210205. PMID 8444142.
- ^ Льюис Р. (2005). Генетика человека: концепции и приложения (6-е изд.). Бостон, Массачусетс: Макгроу Хилл. С. 227–228. ISBN 978-0-07-111156-0.
- ^ Сойер С.А., Парш Дж., Чжан З., Хартл Д.Л. (апрель 2007 г.). «Преобладание положительного отбора среди почти нейтральных аминокислотных замен у дрозофилы». Труды Национальной академии наук Соединенных Штатов Америки. 104 (16): 6504–10. Bibcode:2007ПНАС..104.6504С. Дои:10.1073 / pnas.0701572104. ЧВК 1871816. PMID 17409186.
- ^ Мосты КР (2002). «Малярия и красная клетка». Гарвард. Архивировано из оригинал 27 ноября 2011 г.
- ^ Дрейк Дж. У., Холланд Дж. Дж. (Ноябрь 1999 г.). «Скорость мутаций среди РНК-вирусов». Труды Национальной академии наук Соединенных Штатов Америки. 96 (24): 13910–3. Bibcode:1999PNAS ... 9613910D. Дои:10.1073 / пнас.96.24.13910. ЧВК 24164. PMID 10570172.
- ^ Холланд Дж., Шпиндлер К., Городиски Ф., Грабау Э., Никол С., ВандеПол С. (март 1982 г.). «Быстрая эволюция геномов РНК». Наука. 215 (4540): 1577–85. Bibcode:1982Научный ... 215.1577H. Дои:10.1126 / science.7041255. PMID 7041255.
- ^ de Visser JA, Rozen DE (апрель 2006 г.). «Клональное вмешательство и периодический отбор новых полезных мутаций в Escherichia coli». Генетика. 172 (4): 2093–100. Дои:10.1534 / генетика.105.052373. ЧВК 1456385. PMID 16489229.
- ^ Уотсон, Джеймс Д. (2008). Молекулярная биология гена. Пирсон / Бенджамин Каммингс. ISBN 978-0-8053-9592-1. :102–117 :521–522
- ^ Мишель-Байерле, Мария Элизабет (1990). Центры реакции фотосинтезирующих бактерий: Feldafing-II-Meeting. Springer-Verlag. ISBN 978-3-540-53420-4.
- ^ Füllen G, Youvan DC (1994). «Генетические алгоритмы и рекурсивный ансамблевой мутагенез в белковой инженерии». Комплексность Интернэшнл 1.
- ^ а б Фрике, Маркус (2019). «Глобальное значение вторичных структур РНК в последовательностях, кодирующих белки». Биоинформатика. 35 (4): 579–583. Дои:10.1093 / биоинформатика / bty678. ЧВК 7109657. PMID 30101307. S2CID 51968530.
- ^ «Таблица использования кодонов».
- ^ а б Чжан Ю., Баранов П.В., Аткинс Дж. Ф., Гладышев В. Н. (май 2005 г.). «Пирролизин и селеноцистеин используют разные стратегии декодирования». Журнал биологической химии. 280 (21): 20740–51. Дои:10.1074 / jbc.M501458200. PMID 15788401.
- ^ Кшицкий Я.А. (декабрь 2005 г.). «Прямое генетическое кодирование пирролизина». Текущее мнение в микробиологии. 8 (6): 706–12. Дои:10.1016 / j.mib.2005.10.009. PMID 16256420.
- ^ Прат Л., Хайнеман И.Ю., Аэрни Х.Р., Райнхарт Дж., О'Донохью П., Сёлль Д. (декабрь 2012 г.). «Зависимое от источника углерода расширение генетического кода у бактерий». Труды Национальной академии наук Соединенных Штатов Америки. 109 (51): 21070–5. Bibcode:2012PNAS..10921070P. Дои:10.1073 / pnas.1218613110. ЧВК 3529041. PMID 23185002.
- ^ Крик Ф. Х., Оргель Л. Е. (1973). «Направленная панспермия». Икар. 19 (3): 341–6, 344. Bibcode:1973Icar ... 19..341C. Дои:10.1016/0019-1035(73)90110-3.
Немного удивительно, что организмы с несколько разными кодами не сосуществуют.
(Дальнейшее обсуждение ) - ^ Баррелл Б.Г., Банкир А.Т., Друин Дж. (1979). «Другой генетический код в митохондриях человека». Природа. 282 (5735): 189–194. Bibcode:1979Натура.282..189Б. Дои:10.1038 / 282189a0. PMID 226894. S2CID 4335828. ([1] )
- ^ а б Эльзановский А., Остелл Дж. (7 апреля 2008 г.). "Генетические коды". Национальный центр биотехнологической информации (NCBI). Получено 10 марта 2010.
- ^ Jukes TH, Osawa S (декабрь 1990 г.). «Генетический код митохондрий и хлоропластов». Experientia. 46 (11–12): 1117–26. Дои:10.1007 / BF01936921. PMID 2253709. S2CID 19264964.
- ^ Фитцпатрик Д.А., Logue ME, Stajich JE, Butler G (1 января 2006 г.). «Филогения грибов, основанная на 42 полных геномах, полученных из супердерева и комбинированного анализа генов». BMC Эволюционная биология. 6: 99. Дои:10.1186/1471-2148-6-99. ЧВК 1679813. PMID 17121679.
- ^ Сантос М.А., Туите М.Ф. (май 1995 г.). «Кодон CUG расшифровывается in vivo как серин, а не лейцин у Candida albicans». Исследования нуклеиновых кислот. 23 (9): 1481–6. Дои:10.1093 / nar / 23.9.1481. ЧВК 306886. PMID 7784200.
- ^ Батлер Г., Расмуссен М.Д., Лин М.Ф. и др. (Июнь 2009 г.). «Эволюция патогенности и полового размножения в восьми геномах Candida». Природа. 459 (7247): 657–62. Bibcode:2009Натура.459..657Б. Дои:10.1038 / природа08064. ЧВК 2834264. PMID 19465905.
- ^ Тейлор DJ, Баллинджер MJ, Bowman SM, Брюнн JA (2013). «Коэволюция вируса и хозяина в условиях модифицированного ядерного генетического кода». PeerJ. 1: e50. Дои:10.7717 / peerj.50. ЧВК 3628385. PMID 23638388.
- ^ Hofhuis J, Schueren F, Nötzel C, Lingner T, Gärtner J, Jahn O, Thoms S (2016). «Расширение функционального считывания малатдегидрогеназы обнаруживает модификацию генетического кода». Открыть Биол. 6 (11): 160246. Дои:10.1098 / rsob.160246. ЧВК 5133446. PMID 27881739.
- ^ Шуерен Ф., Лингнер Т., Джордж Р., Хофхейс Дж., Гартнер Дж., Томс С. (2014). «Пероксисомальная лактатдегидрогеназа вырабатывается путем считывания трансляции у млекопитающих». eLife. 3: e03640. Дои:10.7554 / eLife.03640. ЧВК 4359377. PMID 25247702.
- ^ Ф. Шуерен и С. Томс (2016). «Функциональное трансляционное чтение: перспектива системной биологии». PLOS Genetics. 12 (8): e1006196. Дои:10.1371 / journal.pgen.1006196. ЧВК 4973966. PMID 27490485.
- ^ Dutilh BE, Jurgelenaite R, Szklarczyk R, van Hijum SA, Harhangi HR, Schmid M, de Wild B, Françoijs KJ, Stunnenberg HG, Strous M, Jetten MS, Op den Camp HJ, Huynen MA (июль 2011 г.). «FACIL: быстрый и точный вывод генетического кода и логотип». Биоинформатика. 27 (14): 1929–33. Дои:10.1093 / биоинформатика / btr316. ЧВК 3129529. PMID 21653513.
- ^ Кубышкин В., Асеведо-Роча К.Г., Будиса Н. (февраль 2018). «Об универсальных событиях кодирования в биогенезе белков». Биосистемы. 164: 16–25. Дои:10.1016 / j.biosystems.2017.10.004. PMID 29030023.
- ^ Хифи С.М., Мариотти М., Гладышев В.Н., Аткинс Дж.Ф., Баранов П.В. (ноябрь 2016 г.). «Новые варианты генетического кода инфузорий, включая переназначение всех трех стоп-кодонов на смысловые кодоны в Большая кондилостома". Молекулярная биология и эволюция. 33 (11): 2885–2889. Дои:10.1093 / molbev / msw166. ЧВК 5062323. PMID 27501944.
- ^ Лобанов А.В., Хифи С.М., Туранов А.А., Геращенко М.В., Пуччарелли С., Деварадж Р.Р. и др. (Январь 2017 г.). "Зависящее от должности увольнение и широко распространенная обязательная смена кадра в Euplotes перевод". Структурная и молекулярная биология природы. 24 (1): 61–68. Дои:10.1038 / nsmb.3330. ЧВК 5295771. PMID 27870834.
- ^ Рибас де Пуплана Л., Тернер Р. Дж., Стир Б. А., Шиммель П. (сентябрь 1998 г.). "Происхождение генетического кода: тРНК старше своих синтетаз?". Труды Национальной академии наук Соединенных Штатов Америки. 95 (19): 11295–300. Bibcode:1998PNAS ... 9511295D. Дои:10.1073 / пнас.95.19.11295. ЧВК 21636. PMID 9736730.
- ^ а б Ярус, Михаил (2010). Жизнь из мира РНК: Предок внутри. Издательство Гарвардского университета. ISBN 978-0-674-05075-4.
- ^ «Функция Mathematica для # возможного расположения элементов в корзинах? - Группы технического обсуждения в Интернете - Сообщество Wolfram». community.wolfram.com. Получено 3 февраля 2017.
- ^ а б Фриланд С.Дж., Херст Л.Д. (сентябрь 1998 г.). «Генетический код - один на миллион». Журнал молекулярной эволюции. 47 (3): 238–48. Bibcode:1998JMolE..47..238F. Дои:10.1007 / PL00006381. PMID 9732450. S2CID 20130470.
- ^ Тейлор Ф.Дж., Коутс Д. (1989). «Код в кодонах». Биосистемы. 22 (3): 177–87. Дои:10.1016/0303-2647(89)90059-2. PMID 2650752.
- ^ Ди Джулио М (октябрь 1989 г.). «Расширение, достигаемое минимизацией полярных расстояний в ходе эволюции генетического кода». Журнал молекулярной эволюции. 29 (4): 288–93. Bibcode:1989JMolE..29..288D. Дои:10.1007 / BF02103616. PMID 2514270. S2CID 20803686.
- ^ Вонг Дж. Т. (февраль 1980 г.). «Роль минимизации химических расстояний между аминокислотами в эволюции генетического кода». Труды Национальной академии наук Соединенных Штатов Америки. 77 (2): 1083–6. Bibcode:1980PNAS ... 77.1083W. Дои:10.1073 / pnas.77.2.1083. ЧВК 348428. PMID 6928661.
- ^ а б c d Эривес А. (август 2011 г.). «Модель ферментов прото-антикодоновой РНК, требующих гомохиральности L-аминокислоты». Журнал молекулярной эволюции. 73 (1–2): 10–22. Bibcode:2011JMolE..73 ... 10E. Дои:10.1007 / s00239-011-9453-4. ЧВК 3223571. PMID 21779963.
- ^ а б Фриланд С.Дж., Найт Р.Д., Ландвебер Л.Ф., Херст Л.Д. (апрель 2000 г.). «Ранняя фиксация оптимального генетического кода». Молекулярная биология и эволюция. 17 (4): 511–18. Дои:10.1093 / oxfordjournals.molbev.a026331. PMID 10742043.
- ^ Крик Ф.Х. (декабрь 1968 г.). «Происхождение генетического кода». Журнал молекулярной эволюции. 38 (3): 367–79. Дои:10.1016/0022-2836(68)90392-6. PMID 4887876.
- ^ Хопфилд Дж. Дж. (1978). «Происхождение генетического кода: проверяемая гипотеза, основанная на структуре, последовательности тРНК и кинетической корректуре». PNAS. 75 (9): 4334–4338. Bibcode:1978PNAS ... 75.4334H. Дои:10.1073 / пнас.75.9.4334. ЧВК 336109. PMID 279919.
- ^ а б Ярус М., Видманн Дж. Дж., Рыцарь Р. (ноябрь 2009 г.). «Связывание РНК-аминокислоты: стереохимическая эра генетического кода». Журнал молекулярной эволюции. 69 (5): 406–29. Bibcode:2009JMolE..69..406Y. Дои:10.1007 / s00239-009-9270-1. PMID 19795157.
- ^ Knight RD, Freeland SJ, Landweber LF (июнь 1999 г.). «Отбор, история и химия: три грани генетического кода» (PDF). Тенденции в биохимических науках. 24 (6): 241–7. Дои:10.1016 / S0968-0004 (99) 01392-4. PMID 10366854.
- ^ Knight RD, Landweber LF (сентябрь 1998 г.). «Рифма или причина: взаимодействия РНК-аргинина и генетический код». Химия и биология. 5 (9): R215–20. Дои:10.1016 / S1074-5521 (98) 90001-1. PMID 9751648.
- ^ Сенгупта С., Хиггс П. Г. (2015). «Пути эволюции генетического кода у древних и современных организмов». Журнал молекулярной эволюции. 80 (5–6): 229–243. Bibcode:2015JMolE..80..229S. Дои:10.1007 / s00239-015-9686-8. PMID 26054480. S2CID 15542587.
- ^ Brooks DJ, Fresco JR, Lesk AM, Singh M (октябрь 2002 г.). «Эволюция частот аминокислот в белках с течением времени: предполагаемый порядок введения аминокислот в генетический код». Молекулярная биология и эволюция. 19 (10): 1645–55. Дои:10.1093 / oxfordjournals.molbev.a003988. PMID 12270892.
- ^ Амирновин Р (май 1997 г.). «Анализ метаболической теории происхождения генетического кода». Журнал молекулярной эволюции. 44 (5): 473–6. Bibcode:1997JMolE..44..473A. Дои:10.1007 / PL00006170. PMID 9115171. S2CID 23334860.
- ^ Роннеберг Т.А., Ландвебер Л.Ф., Фриланд С.Дж. (декабрь 2000 г.). «Проверка биосинтетической теории генетического кода: факт или артефакт?». Труды Национальной академии наук Соединенных Штатов Америки. 97 (25): 13690–5. Bibcode:2000PNAS ... 9713690R. Дои:10.1073 / pnas.250403097. ЧВК 17637. PMID 11087835.
- ^ Трифонов, Эдвард Н. (сентябрь 2009 г.). «Происхождение генетического кода и самых ранних олигопептидов». Исследования в области микробиологии. 160 (7): 481–486. Дои:10.1016 / j.resmic.2009.05.004.
- ^ Хиггс, Пол Дж .; Пудриц, Ральф Э. (июнь 2009 г.). "Термодинамическая основа синтеза пребиотических аминокислот и природа первого генетического кода". Астробиология. 9 (5): 483–490. Дои:10.1089 / аст.2008.0280. ISSN 1531-1074.
- ^ Халиотис, Анаргирос; Властаридис, Панайотис; Мосиалос, Димитрис; Ибба, Майкл; Becker, Hubert D .; Статопулос, Константинос; Амуциас, Григориос Д. (17 февраля 2017 г.). «Сложная эволюционная история аминоацил-тРНК синтетаз». Исследования нуклеиновых кислот. 45 (3): 1059–1068. Дои:10.1093 / нар / gkw1182. ISSN 0305-1048. ЧВК 5388404. PMID 28180287.
- ^ Нтунтуми, Криса; Властаридис, Панайотис; Мосиалос, Димитрис; Статопулос, Константинос; Илиопулос, Иоаннис; Промпонас, Василиос; Оливер, Стивен Дж. Амуциас, Григорис Д. (4 ноября 2019 г.). «Области низкой сложности в белках прокариот выполняют важные функциональные роли и являются высококонсервативными». Исследования нуклеиновых кислот. 47 (19): 9998–10009. Дои:10.1093 / нар / gkz730. ISSN 0305-1048. ЧВК 6821194. PMID 31504783.
- ^ а б Фриланд SJ, Wu T, Keulmann N (октябрь 2003 г.). «Случай для ошибки минимизации стандартного генетического кода». Истоки жизни и эволюция биосферы. 33 (4–5): 457–77. Bibcode:2003 ОЛЕБ ... 33..457Ф. Дои:10.1023 / А: 1025771327614. PMID 14604186. S2CID 18823745.
- ^ Баранов П.В., Венин М, Прован Г (2009). Геммелл, штат Нью-Джерси (ред.). «Уменьшение размера кодона как источник триплетного генетического кода». PLOS ONE. 4 (5): e5708. Bibcode:2009PLoSO ... 4.5708B. Дои:10.1371 / journal.pone.0005708. ЧВК 2682656. PMID 19479032.
- ^ Tlusty T (ноябрь 2007 г.). «Модель возникновения генетического кода как перехода в зашумленном информационном канале». Журнал теоретической биологии. 249 (2): 331–42. arXiv:1007.4122. Дои:10.1016 / j.jtbi.2007.07.029. PMID 17826800. S2CID 12206140.
- ^ Зоннеборн TM (1965). Брайсон В., Фогель Х (ред.). Развивающиеся гены и белки. Нью-Йорк: Academic Press. С. 377–397.
- ^ Tlusty T (февраль 2008 г.). «Сценарий скоростного искажения для возникновения и развития зашумленных молекулярных кодов». Письма с физическими проверками. 100 (4): 048101. arXiv:1007.4149. Bibcode:2008PhRvL.100d8101T. Дои:10.1103 / PhysRevLett.100.048101. PMID 18352335. S2CID 12246664.
- ^ Селла Дж., Арделл Д.Х. (сентябрь 2006 г.). «Совместная эволюция генов и генетических кодов: повторное посещение замороженной аварии Крика». Журнал молекулярной эволюции. 63 (3): 297–313. Bibcode:2006JMolE..63..297S. Дои:10.1007 / s00239-004-0176-7. PMID 16838217. S2CID 1260806.
- ^ Tlusty T (сентябрь 2010 г.). «Красочное происхождение генетического кода: теория информации, статистическая механика и появление молекулярных кодов». Обзоры физики жизни. 7 (3): 362–76. arXiv:1007.3906. Bibcode:2010ФЛРв ... 7..362Т. Дои:10.1016 / j.plrev.2010.06.002. PMID 20558115. S2CID 1845965.
- ^ Джи Дж., Сандстрем А., Мэсси С.Е., Мишра Б. (ноябрь 2013 г.). «Что информационные асимметричные игры могут рассказать нам о контексте« замороженной аварии »Крика?». Журнал Королевского общества, Интерфейс. 10 (88): 20130614. Дои:10.1098 / rsif.2013.0614. ЧВК 3785830. PMID 23985735.
- ^ Ицковиц С, Алон У (2007). «Генетический код почти оптимален для предоставления дополнительной информации в кодирующих белки последовательностях». Геномные исследования. 17 (4): 405–412. Дои:10.1101 / гр.5987307. ЧВК 1832087. PMID 17293451.
- ^ «Биосети - сообщение, которое доходит до адресата».
- ^ Максим Макуков и Владимир Щербак (апрель 2018). «SETI in vivo: проверка гипотезы« мы есть они »». Международный журнал астробиологии. 17 (2): 127–146. arXiv:1707.03382. Bibcode:2018IJAsB..17..127M. Дои:10.1017 / S1473550417000210. S2CID 44826721.
{kind=link}
дальнейшее чтение
- Гриффитс AJ, Миллер JH, Suzuki DT, Левонтин RC, Гилберт WM (1999). Введение в генетический анализ (7-е изд.). Сан-Франциско: W.H. Фримен. ISBN 978-0-7167-3771-1.
- Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П. (2002). Молекулярная биология клетки (4-е изд.). Нью-Йорк: Наука о гирляндах. ISBN 978-0-8153-3218-3.
- Лодиш Х.Ф., Берк А., Зипурски С.Л., Мацудаира П., Балтимор Д., Дарнелл Дж. Э. (2000). Молекулярная клеточная биология (4-е изд.). Сан-Франциско: W.H. Фримен. ISBN 978-0-7167-3706-3.
- Каски CT, Ледер П. (апрель 2014 г.). «Код РНК: Розеттский камень природы». Труды Национальной академии наук Соединенных Штатов Америки. 111 (16): 5758–9. Bibcode:2014PNAS..111.5758C. Дои:10.1073 / pnas.1404819111. ЧВК 4000803. PMID 24756939.
внешняя ссылка
- Генетические коды: таблицы генетических кодов
- В База данных использования кодонов - Таблицы частот кодонов для многих организмов
- История расшифровки генетического кода