Эволюция млекопитающих - Википедия - Evolution of mammals

В эволюция из млекопитающие прошел много этапов с момента первого появления своих синапсид предки в Пенсильванский подпериод позднего Каменноугольный период. К серединеТриасовый, было много видов синапсидов, похожих на млекопитающих. Родословная, ведущая к сегодняшним млекопитающим, разделилась на Юрский; синапсиды этого периода включают Dryolestes, более тесно связанные с дошедшими до нас плаценты и сумчатые чем монотремы, а также Амбондро, более тесно связанный с монотремами.[1] Позже евтерский и метатериан родословные разделены; метатерии - животные, более тесно связанные с сумчатыми, а эвтерианы - более близкие к плацентарным. С Джурамая, самый ранний из известных эвтерианцев, живший 160 миллионов лет назад в юрском периоде, это расхождение должно было произойти в тот же период.
После Меловое – палеогеновое вымирание уничтожил нептичьих динозавров (птицы являются единственными выжившими динозаврами) и нескольких групп млекопитающих, плацентарных и сумчатых млекопитающих, появившихся во многих новых формах и экологических нишах по всему миру. Палеоген и Неоген, к концу которого все современные заказы появился.
Млекопитающие - единственные живые синапсиды.[2] Линия синапсидов стала отличаться от сауропсид происхождение в конце каменноугольного периода, между 320 и 315 миллионами лет назад.[3] Сауропсиды - сегодняшние рептилии и птицы, а также все вымершие животные, более тесно связанные с ними, чем с млекопитающими.[3] Это не включает млекопитающие рептилии, группа, более тесно связанная с млекопитающими.
На протяжении Пермский период периода синапсиды включали доминирующие плотоядные животные и несколько важных травоядные животные. Однако в последующий триасовый период ранее неизвестная группа зауропсид, архозавры, стали доминирующими позвоночными. В млекопитающие появились в этот период; их превосходное обоняние, подкрепленное большим мозгом, облегчало проникновение в ночные ниши с меньшим воздействием хищничества архозавров. Ночной образ жизни, возможно, внес большой вклад в развитие таких черт млекопитающих, как эндотермия и волосы. Позже в Мезозойский, после теропод динозавры заменены Rauisuchians как доминирующие хищники, млекопитающие распространились на другие экологические ниши. Например, некоторые стали водный, некоторые были планеры, а некоторые даже питаются молодыми динозаврами.[нужна цитата ]
Большинство доказательств состоит из окаменелости. В течение многих лет окаменелости мезозойских млекопитающих и их непосредственных предков были очень редкими и фрагментарными; но с середины 1990-х годов было сделано много важных новых находок, особенно в Китае. Относительно новые методы молекулярная филогенетика также пролили свет на некоторые аспекты эволюции млекопитающих, оценив сроки важных точек расхождения для современных видов. При осторожном использовании эти методы часто, но не всегда, согласуются с летописью окаменелостей.[нужна цитата ]
Несмотря на то что молочные железы являются характерной чертой современных млекопитающих, мало что известно об эволюции кормление грудью поскольку эти мягкие ткани не часто сохраняются в летописи окаменелостей. Большинство исследований, касающихся эволюции млекопитающих, сосредоточено на форме зубов, наиболее твердых частях тела. четвероногий тело. Другие важные характеристики исследования включают эволюцию кости среднего уха, прямая осанка конечностей, костная вторичная нёбо, мех, волосы и теплокровность.[нужна цитата ]
Определение «млекопитающее»
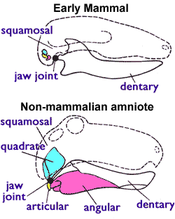
В то время как живые виды млекопитающих могут быть идентифицированы по наличию молочных продуктов. молочные железы у самок требуются другие признаки при классификации окаменелости, потому что молочные железы и другие элементы мягких тканей не видны в окаменелостях.
Одна такая функция доступна для палеонтология, общие для всех живых млекопитающих (включая монотремы ), но не присутствовал ни в одном из ранних Триасовый терапсиды, показан на рисунке 1 (справа), а именно: млекопитающие используют две кости за то, что слышал, что все остальные амниот использовать для еды. У самых ранних амниот был челюстной сустав, состоящий из суставной (небольшая кость в задней части нижней челюсти) и квадратный (небольшая кость в задней части верхней челюсти). Все не млекопитающие четвероногие использовать эту систему, включая амфибии, черепахи, ящерицы, змеи, крокодилы, динозавры (в том числе птицы ), ихтиозавры, птерозавры и терапсиды. Но у млекопитающих другой челюстной сустав, состоящий только из зубной (кость нижней челюсти, несущая зубы) и чешуйчатый (еще одна маленькая кость черепа). В юрском периоде их квадратные и суставные кости эволюционировали в наковальня и молоточек кости в среднее ухо.[4][5] У млекопитающих тоже есть двойная затылочный мыщелок; у них есть два выступа у основания черепа, которые подходят к самому верхнему шейному позвонку, в то время как у других четвероногих есть единственный затылочный мыщелок.[4]
В статье 1981 года Кеннет А. Кермак и его соавторы приводили доводы в пользу проведения границы между млекопитающими и более ранними синапсидами в той точке, где у млекопитающих коренной зуб окклюзия приобретался и появился зубно-чешуйчатый сустав. Они отметили, что выбранный критерий является просто вопросом удобства; их выбор был основан на том факте, что «нижняя челюсть является наиболее вероятным сохранившимся элементом скелета мезозойского млекопитающего».[6] Сегодня большинство палеонтологов считают животных млекопитающими, если они удовлетворяют этому критерию.[7]
Происхождение млекопитающих
| Пеликозавры |
Амниот
Первый полностью земной позвоночные мы амниот - их яйца имели внутренние оболочки, которые позволяли развиваться эмбрион чтобы дышать, но сохраняла воду. Это позволяло амниотам откладывать яйца на суше, в то время как амфибиям обычно нужно откладывать яйца в воде (некоторые амфибии, такие как обыкновенная суринамская жаба, имеют развился другие способы обойти это ограничение). Первые амниоты, по-видимому, возникли посередине Каменноугольный от предков рептилии.[8]
В течение нескольких миллионов лет были выделены две важные линии амниот: млекопитающие синапсид предки и завропсиды, откуда ящерицы, змеи, черепахи / черепахи, крокодилы, динозавры и птицы являются потомками.[3] Самые ранние известные окаменелости синапсидов и сауропсид (например, Археотирис и Гилономус соответственно) датируются примерно 320-315 миллионами лет назад. Трудно узнать время происхождения, потому что окаменелости позвоночных из позднего карбона очень редки, и поэтому фактические первые появления каждого из этих типов животных могли быть значительно раньше чем первое ископаемое.[9]
Синапсиды

Synapsid черепа идентифицируются по отличительному рисунку дыры за каждым глазом, которые служили следующим целям:
- сделал череп легче, не жертвуя силой.
- экономия энергии за счет использования меньшего количества костей.
- Вероятно, предусмотрены точки крепления мышц челюсти. Наличие точек прикрепления, расположенных дальше от челюсти, позволило мышцам стать длиннее и, следовательно, оказывать сильное натяжение в широком диапазоне движений челюсти, не растягиваясь или сокращаясь за пределы их оптимального диапазона.
Синапсид пеликозавры включены крупнейшие наземные позвоночные Ранняя пермь, например, длиной 6 м (20 футов) Cotylorhynchus hancocki. Среди других крупных пеликозавров были Диметродон большой и Edaphosaurus cruciger.
Терапсиды
Терапсиды произошли от пеликозавров в средняя пермь и заняли свое положение в качестве доминирующих наземных позвоночных. Они отличаются от пеликозавров несколькими особенностями черепа и челюстей, в том числе более крупными. височные отверстия и резцы которые равны по размеру.[10]
Линия терапевсидов, которая привела к млекопитающим, прошла ряд стадий, начиная с животных, очень похожих на своих предков-пеликозавров, и заканчивая некоторыми, которых легко можно было принять за млекопитающих:[11]
- постепенное развитие костной вторичной нёбо. Большинство книг и статей интерпретируют это как предпосылку для эволюции высокого уровня развития млекопитающих. скорость метаболизма, потому что он позволял этим животным есть и дышать одновременно. Но некоторые ученые отмечают, что некоторые современные эктотермия используйте мясистое вторичное небо, чтобы отделить ротовую полость от дыхательных путей, и костлявый небо обеспечивает поверхность, на которой язык может манипулировать пищей, облегчая жевание, а не дыхание.[12] Интерпретация костного вторичного неба как вспомогательного средства при жевании также предполагает развитие более быстрого метаболизма, потому что жевание уменьшает размер частиц пищи, доставляемых в желудок, и, следовательно, может ускорить их переваривание. У млекопитающих нёбо образовано двумя специфическими костями, но различные пермские терапсиды имели другие комбинации костей в нужных местах, чтобы функционировать как нёбо.
- то зубной постепенно становится основной костью нижней челюсти.
- прогрессировать в направлении вертикального положения конечностей, что повысит выносливость животных, избегая Ограничение перевозчика. Но этот процесс был беспорядочным и очень медленным - например: у всех травоядных терапсидов сохранились раскидистые конечности (некоторые поздние формы могли иметь полустоячие задние конечности); Пермские плотоядные терапсиды имели раскидистые передние конечности, а у некоторых позднепермских также были полурасположенные задние конечности. На самом деле современные монотремы все еще есть полуразложенные конечности.
Семейное древо терапсид
(упрощено с Palaeos.com[10] ниже описаны только те, которые имеют самое непосредственное отношение к эволюции млекопитающих)
| Терапсиды |
| ||||||||||||||||||||||||||||||||||||||||||
Только дицинодонты, тероцефалы и цинодонты дожили до триаса.
Биармозухия
В Биармозухия были самыми примитивными и похожими на пеликозавров из терапсидов.[13]
Диноцефалы
Диноцефалы («страшные головы») включали как плотоядных, так и травоядных. Они были большими; Антеозавр был до 6 м (20 футов) в длину. У некоторых хищников были полустоячие задние конечности, но у всех диноцефалов передние конечности были распростертыми. Во многих отношениях они были очень примитивными терапсидами; например, у них не было вторичного неба, а их челюсти были скорее «рептильными».[14]
Аномодонты

В аномодонты («аномальные зубы») были одними из самых успешных среди травоядных терапсидов - одна подгруппа, дицинодонты, просуществовал почти до конца триаса. Но аномодонты сильно отличались от современных травоядных млекопитающих, так как их единственными зубами была пара клыков в верхней челюсти, и принято считать, что у них были клювы, как у птиц или птиц. цератопсы.[15]
Териодонты
В териодонты («звериные зубы») и их потомки имели челюстные суставы, в которых нижняя челюсть суставной кость плотно сжимала очень маленький череп квадратная кость. Это дало возможность гораздо шире, и одна группа, хищные горгонопсийцы («лица горгоны»), воспользовались этим для развития «саблезубых зубов». Но шарнир челюсти у териодонта имел более долгосрочное значение - значительно уменьшенный размер квадратной кости был важным шагом в развитии челюстного сустава и среднего уха млекопитающих.
У горгонопсий все еще были некоторые примитивные черты: отсутствие костного вторичного неба (но другие кости в нужных местах для выполнения тех же функций); раздвинутые передние конечности; задние конечности, которые могли работать как в раскинутых, так и в вертикальных позах. Но тероцефалы («звериные головы»), которые, по-видимому, возникли примерно в то же время, что и горгонопсии, имели дополнительные признаки, похожие на млекопитающих, например их кости пальцев рук и ног имели такое же количество фаланг (сегментов), как и у ранних млекопитающих (и такое же количество, что и у них). приматы есть, включая людей).[16]
Цинодонты

В цинодонты, группа териодонтов, возникшая также в конце Пермский период, включая предков всех млекопитающих. Характерные черты цинодонтов, схожие с млекопитающими, включают дальнейшее уменьшение количества костей в нижней челюсти, вторичное костное небо, щечные зубы со сложным рисунком на коронках и мозг, заполняющий эндокраниальную полость.[17]
Обнаружены многокамерные норы, содержащие до 20 скелетов цинодонта раннего триаса. Trirachodon; Считается, что животные были утоплены внезапное наводнение. Обширные общие норы указывают на то, что эти животные были способны к сложному социальному поведению.[18]
Захват триаса
Катастрофический массовое вымирание в конце перми около 252 миллионов лет назад, погибло около 70 процентов земной позвоночное животное виды и большинство наземных растений.
Как результат,[19] экосистемы и пищевые цепи рухнула, и на создание новых стабильных экосистем ушло около 30 миллионов лет. С исчезновением горгонопсийцев, которые были доминирующими хищниками в поздней перми,[20] Основными конкурентами цинодонтов за доминирование в хищных нишах были ранее малоизвестная группа зауропсид, архозавры, в который входят предки крокодилов и динозавров.
Архозавры быстро стали доминирующими хищниками,[20] развитие, которое часто называют «захватом триаса». Их успех мог быть связан с тем, что ранний триас был преимущественно засушливым, и поэтому превосходная водосбережение архозавров дало им решающее преимущество. Все известные архозавры имеют кожу без желез и устраняют азотные отходы в мочевая кислота пасты, содержащей мало воды, в то время как цинодонты, вероятно, выделяли большую часть таких отходов в растворе мочевина, как млекопитающие сегодня; для растворения мочевины требуется значительное количество воды.[21]
Однако эта теория была подвергнута сомнению, поскольку она подразумевает, что синапсиды обязательно имели меньшие преимущества в удержании воды, что уменьшение синапсидов совпадает с изменениями климата или разнообразием архозавров (ни одно из которых не было проверено), и тот факт, что обитающие в пустыне млекопитающие также хорошо адаптированы. в этом отделе как архозавры,[22] и некоторые цинодонты вроде Труцидоцинодон были крупными хищниками.[23]
Захват триаса, вероятно, был жизненно важным фактором в эволюции млекопитающих. Две группы, происходящие от ранних цинодонтов, добились успеха в нишах, которые имели минимальную конкуренцию со стороны архозавров: тритилодонты, которые были травоядные животные, а также млекопитающие, большинство из которых вели небольшой ночной образ жизни. насекомоядные (хотя некоторые, вроде Sinoconodon, были плотоядными животными, которые питались добычей позвоночных, в то время как третьи были травоядными или всеядные ).[24] Как результат:
- Тенденция терапевсида к дифференцировке зубов с точным окклюзия ускоряется, из-за необходимости удерживать захваченный членистоногие и раздавить их экзоскелеты.
- Поскольку длина тела предков млекопитающих упала ниже 50 мм (2 дюймов), прогресс в теплоизоляция и регулирование температуры стал бы необходимым для ночной жизни.[25]
- Обострились острый слух и обоняние.
- Это ускорило развитие среднего уха млекопитающих.
- Увеличение размера обонятельных долей головного мозга увеличивало массу мозга в процентах от общей массы тела.[26] Мозговая ткань требует непропорционально много энергии.[27][28] Потребность в большем количестве пищи для поддержки увеличенного мозга увеличила необходимость улучшения изоляции, регулирования температуры и кормления.
- Вероятно, как побочный эффект ночного образа жизни, млекопитающие потеряли две из четырех колбочек. опсины, фоторецепторы в сетчатка, присутствует в глазах самых ранних амниот. Парадоксально, но это могло улучшить их способность различать цвета при тусклом свете.[29]
Это отступление к ночной роли называется ночное узкое место, и считается, что он объясняет многие особенности млекопитающих.[30]
От цинодонтов до коронных млекопитающих
Окаменелости
Мезозойские синапсиды, которые эволюционировали до состояния челюстного сустава, состоящего из зубных и чешуйчатых костей, сохранились в нескольких хороших окаменелостях, главным образом потому, что они были в основном меньше, чем крысы:
- Они были в основном ограничены средами, которые с меньшей вероятностью окаменелости. Поймы поскольку лучшая земная среда для окаменения дает мало окаменелостей млекопитающих, потому что в них преобладают средние и крупные животные, а млекопитающие не могут конкурировать с архозавры в диапазоне средних и больших размеров. Следы из раннего мела Анголы показывают существование млекопитающих размером с енота 118 миллионов лет назад.[31]
- Их хрупкие кости были уязвимы для разрушения, прежде чем они могли быть окаменелы - мусорщиками (включая грибы и бактерии ) и тем, что по нему наступают.
- Маленькие окаменелости труднее обнаружить и более уязвимы для разрушения в результате выветривания и других природных воздействий, прежде чем они будут обнаружены.
Однако за последние 50 лет количество ископаемых мезозойских млекопитающих значительно увеличилось; например, только 116 родов были известны в 1979 году, а в 2007 году - около 310, с повышением качества, так что «по крайней мере 18 мезозойских млекопитающих представлены почти полными скелетами».[32]
Млекопитающие или формы млекопитающих
Некоторые авторы ограничивают термин «млекопитающее» группа короны млекопитающие, группа, состоящая из самых недавних общих предков монотремы, сумчатые, и плаценты вместе со всеми потомками этого предка. В влиятельной статье 1988 года Тимоти Роу отстаивал это ограничение, утверждая, что «происхождение ... обеспечивает единственное средство для правильного определения таксонов» и, в частности, что отклонение однообразных от животных, более тесно связанных с сумчатыми и плацентарными животными » представляет центральный интерес для любого исследования млекопитающих в целом ».[33] Чтобы учесть некоторые родственные таксоны, не входящие в группу крон, он определил Млекопитающие как включающий «последнего общего предка Morganucodontidae и Mammalia [как он определил последний термин] и всех их потомков». Помимо Morganucodontidae, новый таксон включает Docodonta и Kuehneotheriidae. Хотя харамиидов относят к млекопитающим с 1860-х годов,[34] Роу исключил их из Mammaliaformes как не подпадающих под его определение, поместив их в более крупную кладу, Млекопитающие.
Некоторые авторы приняли эту терминологию, чтобы избежать недоразумений, отметив, что они это сделали. Однако большинство палеонтологов по-прежнему считают, что животные с зубно-чешуйчатым челюстным суставом и молярами, характерными для современных млекопитающих, формально должны быть членами Mammalia.[7]
Там, где двусмысленность в термине «млекопитающее» может сбивать с толку, в этой статье используются термины «млекопитающее» и «коронное млекопитающее».
Генеалогическое древо - цинодонты к млекопитающим группы кроны
(на основе Цинодонтия: Дендрограмма - Палеос )
| Цинодонтия |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

Morganucodontidae
В Morganucodontidae впервые появился в конце триаса, около 205 миллионов лет назад. Они являются прекрасным примером переходных окаменелостей, поскольку имеют как зубно-чешуйчатые, так и суставно-квадратные челюстные суставы.[35] Они также были одними из первых обнаруженных и наиболее тщательно изученных млекопитающие вне коронная группа млекопитающих, поскольку было обнаружено необычно большое количество окаменелостей морганукодонтов.
Докодонты

Докодонты, среди наиболее распространенных юрских млекопитающих, известны изощренностью своих коренных зубов. Считается, что у них были общие полуводные наклонности, с поеданием рыбы. Castorocauda («бобровый хвост»), который жил в серединеЮрский около 164 миллионов лет назад, впервые был обнаружен в 2004 году и описан в 2006 году, являясь наиболее понятным примером. Castorocauda не входил в состав коронной группы млекопитающих, но он чрезвычайно важен для изучения эволюции млекопитающих, потому что первой находкой был почти полный скелет (настоящая роскошь в палеонтологии), и она ломает стереотип «маленькое ночное насекомоядное»:[36]
- Он был заметно больше, чем большинство окаменелостей мезозойских млекопитающих - около 17 дюймов (43 см) от носа до кончика 5-дюймового (130 мм) хвоста и, возможно, весил 500–800 г (18–28 унций).
- Это самые ранние абсолютно достоверные свидетельства наличия волос и меха. Ранее самая ранняя из них была Eomaia, млекопитающее коронной группы, появившееся около 125 миллионов лет назад.
- У него были водные приспособления, включая сплющенные кости копыта и остатки мягких тканей между пальцами задних лап, что позволяет предположить, что они были перепончатыми. Ранее самые ранние известные полуводные формы млекопитающих были из эоцен, примерно 110 миллионов лет спустя.
- Castorocauda 'Мощные передние конечности выглядят приспособленными для рытья. Эта особенность и шпоры на лодыжках делают его похожим на утконос, который тоже плавает и копает.
- Его зубы выглядят приспособленными для поедания рыбы: первые два коренных зуба имели бугорки в прямой ряд, что делало их более подходящими для захвата и разрезания, чем для измельчения; и эти коренные зубы изогнуты назад, чтобы помочь в захвате скользкой добычи.
Адрокодиум
Семейное древо выше показывает Адрокодиум как «тётя» кроновых млекопитающих. Эта форма млекопитающего, датированная примерно 195 миллионами лет назад в самом раннем юрском периоде, демонстрирует некоторые важные особенности:[37]
- Сустав челюсти состоит только из чешуйчатой и зубной костей, а челюсть не содержит меньших костей к задней части зубной кости, в отличие от конструкции терапсида.
- В терапсиды и ранние млекопитающие формируют барабанная перепонка возможно, растянулись над желобом в задней части нижней челюсти. Но Адрокодиум не имел такого желоба, что позволяет предположить, что его ухо было частью череп, как и у млекопитающих группы кроны - и, следовательно, первые суставной и квадратный перекочевал в среднее ухо и стал молоточек и наковальня. С другой стороны, у зубной кости есть «бухта» сзади, которой нет у млекопитающих. Это говорит о том, что Hadrocodium's зубная кость сохранила ту же форму, которую имела бы, если бы суставная и квадратная кости оставались частью челюстного сустава, и поэтому Адрокодиум или очень близкий предок мог быть первым, кто имел полностью среднее ухо млекопитающего.
- У терапсидов и более ранних форм млекопитающих челюстные суставы располагались очень далеко в черепе, отчасти потому, что ухо находилось на заднем конце челюсти, но также должно было быть близко к мозгу. Такое расположение ограничивало размер черепной коробки, потому что заставляло мышцы челюсти бегать по ней. Hadrocodium's черепная коробка и челюсти больше не были связаны друг с другом из-за необходимости поддерживать ухо, и его челюстной сустав был продвинут вперед. У его потомков или у потомков животных с аналогичным расположением корпус мозга мог свободно расширяться, не ограничиваясь челюстью, и челюсть могла свободно меняться, не ограничиваясь необходимостью держать ухо рядом с мозгом - другими словами Теперь у млекопитающих появилась возможность развить большой мозг и приспособить свои челюсти и зубы исключительно к еде.
Древнейшие коронные млекопитающие
В группа короны млекопитающие, которых иногда называют «настоящими млекопитающими», являются сохранившийся млекопитающие и их родственники до их последнего общего предка. Поскольку в этой группе есть живые участники, ДНК Анализ может быть применен в попытке объяснить эволюцию особенностей, которые не появляются в окаменелостях. Это стремление часто включает молекулярная филогенетика, техника, которая стала популярной с середины 1980-х годов.
Генеалогическое древо млекопитающих ранней кроны
Кладограмма по З.-Х Ло.[32] († отмечает вымершие группы)
| Корона группа млекопитающие |
| ||||||||||||||||||||||||||||||||||||||||||
Цветовое зрение
Ранние амниоты имели четыре опсина в колбочках сетчатки, которые использовались для различения цветов: один, чувствительный к красному, один к зеленому и два, соответствующих различным оттенкам синего.[38][39] Зеленый опсин не был унаследован никакими коронными млекопитающими, но все нормальные особи унаследовали красный. Таким образом, у ранних кроновых млекопитающих было три опсина колбочек: красный и оба голубых.[38] Все их нынешние потомки потеряли один из чувствительных к синему опсинов, но не всегда один и тот же: монотремы сохраняют один чувствительный к синему опсин, в то время как сумчатые и плацентарные животные сохраняют другой (кроме китообразные, который позже потерял и другой синий опсин).[40] У некоторых плацентарных и сумчатых животных, включая высших приматов, впоследствии развились опсины, чувствительные к зеленому; подобно ранним коронным млекопитающим, поэтому их зрение трехцветный.[41][42]
Australosphenida и Ausktribosphenidae
Ausktribosphenidae это название группы, которое было дано некоторым довольно загадочным находкам, которые:[43]
- кажется, есть трибосфенические моляры, тип зуба, который иначе известен только у плацентарных и сумчатых животных.[44]
- приходят из серединыМеловой месторождения в Австралии, но Австралия была связана только с Антарктидой, а плаценты возникли в Северном полушарии и оставались там до тех пор, пока Континентальный дрифт сформировали сухопутные связи из Северной Америки в Южную Америку, из Азии в Африку и из Азии в Индию (карта позднего мела здесь показывает, как разделены южные континенты).
- представлены только зубами и фрагментами челюстей, что не очень помогает.
Австралосфенида это группа, которая была определена для включения Ausktribosphenidae и монотремы. Асфальтомилос (средне-поздняя юра, от Патагония ) был интерпретирован как базальный австралосфенид (животное, которое имеет общие черты как с Ausktribosphenidae, так и с монотремами; не имеет черт, присущих Ausktribosphenidae или монотремам; также не имеет черт, которые отсутствуют у Ausktribosphenidae и монотрем), и свидетельствует о том, что австралоосфениды были широко распространены повсюду. Гондвана (старый суперконтинент Южного полушария).[45]
Недавний анализ Тейнолофос, который жил где-то между 121 и 112,5 миллионами лет назад, предполагает, что это была «группа короны» (продвинутая и относительно специализированная) монотремов. Это было воспринято как доказательство того, что базальные (наиболее примитивные) монотремы должны были появиться значительно раньше, но это оспаривается (см. Следующий раздел). Исследование также показало, что некоторые предполагаемые австралосфениды также были монотремами «коронной группы» (например, Стероподон ) и другие предполагаемые австралосфениды (например, Аусктрибосфенос, Епископы, Амбондро, Асфальтомилос) являются более близкими родственниками и, возможно, членами терианских млекопитающих (группа, в которую входят сумчатые и плацентарные, см. ниже).[46]
Монотремы
Тейнолофос из Австралии, является самой ранней известной монотремой. Исследование 2007 г. (опубликовано в 2008 г.) предполагает, что это была не базальная (примитивная, предковая) монотремия, а полноценная утконос, поэтому утконос и ехидна линии разошлись значительно раньше.[46] Однако более недавнее исследование (2009 г.) показало, что, хотя Тейнолофос был разновидностью утконоса, он также был базальной монотремой и предшествовал излучению современных монотрем. Полуводный образ жизни утконосов не позволял им уступить место сумчатым, мигрировавшим в Австралию миллионы лет назад, поскольку джои должны оставаться привязанными к своим матерям и утонуть, если их матери рискнутся в воду (хотя есть исключения, такие как водяной опоссум и лютрин опоссум; однако они оба живут в Южной Америке и поэтому не контактируют с монотремами). Генетические данные определили, что ехидны отделились от линии утконоса не так давно, в 19-48M, когда они совершили переход от полуводного к наземному образу жизни.[47]
Монотремы имеют некоторые особенности, которые могут быть унаследованы от цинодонт предки:
- подобно ящерицам и птицам, они используют одно и то же отверстие для мочеиспускания, дефекации и размножения («монотрем» означает «одно отверстие»).
- они лежат яйца кожистые и некальцинированные, как у ящериц, черепах и крокодилов.
В отличие от других млекопитающих, самки однополых не имеют соски и кормят своих детенышей "потным" молоком из пятен на животе.
Эти особенности не видны в окаменелостях, и основными характеристиками с точки зрения палеонтологов являются:[43]
- стройный зубной кость, в которой венечный отросток маленький или отсутствует.
- наружное отверстие уха лежит у заднего основания челюсти.
- то скуловой кость маленькая или отсутствует.
- примитивный грудной пояс с сильным вентральный элементы: коракоиды, ключицы и межключица. Примечание: терианец у млекопитающих нет межключичного сустава.[48]
- раскидистые или полуразложенные передние конечности.
Мультитуберкулезные
Мультитуберкулезные (назван в честь нескольких бугорки на их "коренные зубы ") часто называют" грызунами мезозоя ", но это пример конвергентная эволюция а не означать, что они тесно связаны с Rodentia. Они существовали около 120 миллионов лет - это самая длинная история окаменелостей среди всех родословных млекопитающих, - но в конечном итоге их вытеснили грызуны, и они вымерли в первые годы жизни. Олигоцен.
Некоторые авторы подвергли сомнению филогению, представленную на приведенной выше кладограмме. Они исключают мультитуберкулятов из коронной группы млекопитающих, считая, что мультитуберкуляты более отдаленно связаны с современными млекопитающими, чем даже Morganucodontidae.[49][50] Мультитуберкуляты похожи на бесспорных коронных млекопитающих тем, что их челюстные суставы состоят только из зубной и чешуйчатый кости - тогда как квадратный и суставной кости являются частью среднего уха; их зубы дифференцированы, закупорены и похожи на зубы млекопитающих. куспиды; у них есть скуловая дуга; и структура таз предполагает, что они родили крошечных беспомощных детенышей, как современные сумчатые.[51] С другой стороны, они отличаются от современных млекопитающих:
- Их «коренные зубы» имеют два параллельных ряда бугорков, в отличие от трибосфеновых (трехостержневых) коренных зубов неоспоримых ранних корончатых млекопитающих.
- В отличается жевательным действие в этом бесспорном крауне млекопитающих жевать с стороны в сторону шлифовального действия, что означает, что коренные зубы, как правило, закупорить только на одну стороны за один раз, в то время как челюсть multituberculates' было неспособны стороны в сторону движения, они жевал, скорее, оттягивая нижние зубы назад к верхним, когда челюсть сомкнулась.
- Передняя (передняя) часть скуловой дуги в основном состоит из верхняя челюсть (верхняя челюсть), а не скуловой, небольшая кость в небольшой прорези в верхнечелюстном отростке (расширение).
- В чешуйчатый не является частью черепная коробка.
- В трибуна (морда) не похожа на морду бесспорных верховых млекопитающих; на самом деле это больше похоже на пеликозавр, Такие как Диметродон. Многотуберкулезный рострум коробчатой формы, с большими плоскими максиллами по бокам, носовой верх и высокий предчелюстная кость впереди.
Терия

Терия («звери») - это клады происходящий от последнего общего предка Евтерия (включая плаценты ) и Метатерия (в том числе сумчатых). Общие функции включают:[52]
- нет межключица.[48]
- коракоидный кости отсутствуют или срослись с лопатка формировать коракоидные отростки.
- тип голеностопный голеностопный сустав, в котором: основной сустав находится между большеберцовая кость и астрагал; то пяточная кость не контактирует с голенью, но образует пятку, к которой могут прикрепляться мышцы. (Другой хорошо известный тип голеностопного сустава голеностопного сустава встречается у крокодилов и работает по-другому - большая часть изгиба в голеностопном суставе находится между пяточной костью и астрагалом).
- трибосфенические моляры.[44]
Метатерия
Живые Метатерия все сумчатые (животные с мешочками). Несколько ископаемых роды, например, монгольский поздний меловой период Азиатерий, могут быть сумчатыми или членами какой-либо другой группы (групп) метатерии.[53][54]
Самый старый известный метатериан - это Sinodelphys, найден в 125-летнем раннемеловом сланец на северо-востоке Китая Провинция Ляонин. Ископаемое почти готово и включает пучки меха и отпечатки мягких тканей.[55]
Дидельфиморфия (опоссумы обыкновенные Западное полушарие ) впервые появились в позднем меловом периоде и до сих пор имеют живых представителей, вероятно потому, что они в основном полу-древесный неспециализированный всеядные.[56]
Самая известная особенность сумчатых - способ размножения:
- Мать развивает своего рода желток мешок в ее утробе, который доставляет питательные вещества к эмбрион. Эмбрионы бандикуты, коалы и вомбаты дополнительно образуют плацентообразные органы, которые соединяют их с матка стенка, хотя плацентоподобные органы меньше, чем у плацентарных млекопитающих, и нет уверенности в том, что они переносят питательные вещества от матери к эмбриону.[57]
- Беременность очень короткая, обычно от четырех до пяти недель. Эмбрион рождается на очень ранней стадии развития, и его длина при рождении обычно составляет менее 2 дюймов (5,1 см). Было высказано предположение, что короткая беременность необходима для снижения риска материнского иммунная система атакует эмбрион.
- Новорожденное сумчатое животное использует передние конечности (с относительно сильными руками), чтобы забраться на сосок, который обычно находится в сумке на животе матери. Мать кормит ребенка, сокращая над ней мышцы молочные железы, так как ребенок слишком слаб, чтобы сосать. Исторически считалось, что потребность новорожденного сумчатого использовать свои передние конечности для восхождения к соску ограничивала метатериальную эволюцию, поскольку предполагалось, что передняя конечность не может стать специализированной внутренней структурой, такой как крылья, копыта или ласты. Однако несколько бандикуты, в первую очередь свиноногий бандикут, имеют настоящие копыта, похожие на копыта плацентарных копытных, и эволюционировали несколько сумчатых планеров.

Хотя некоторые сумчатые очень похожи на некоторые плаценты ( тилацин, "сумчатый тигр" или "сумчатый волк" - хороший пример), у сумчатых скелетов есть некоторые особенности, которые отличают их от плацентарных:[58]
- У некоторых, включая тилацин, четыре коренных зуба; тогда как ни у одной известной плаценты их больше трех.
- У всех есть пара небных отверстий, оконных отверстий в нижней части черепа (в дополнение к меньшим отверстиям для ноздрей).
У сумчатых также есть пара сумчатых костей (иногда их называют "надлобковые кости "), которые поддерживают сумку у самок. Но они не уникальны для сумчатых, поскольку они были обнаружены в окаменелостях мультитуберкулезных, одноплодных и даже эвтерианских особей - так что они, вероятно, являются общей наследственной чертой, которая исчезла в какой-то момент после родословной. живых плацентарных млекопитающих отличились от сумчатых.[59][60]Некоторые исследователи считают, что первоначальная функция надлобковых костей заключалась в том, чтобы помогать передвижению, поддерживая некоторые мышцы, которые тянут бедро вперед.[61]
Евтерия
Время появления первых евтерианцев было предметом споров. С одной стороны, недавно обнаруженные окаменелости Джурамая были датированы 160 миллионами лет назад и классифицированы как эвтерианские.[62] Окаменелости Eomaia from 125 million years ago in the Раннемеловой период have also been classified as eutherian.[63] A recent analysis of phenomic characters, however, classified Eomaia as pre-eutherian and reported that the earliest clearly eutherian specimens came from Maelestes, dated to 91 million years ago.[64] That study also reported that eutherians did not significantly diversify until after the catastrophic extinction at the Граница мела и палеогена, about 66 million years ago.
Eomaia was found to have some features that are more like those of marsupials and earlier metatherians:

- Epipubic bones extending forwards from the pelvis, which are not found in any modern placental, but are found in all other mammals — early mammaliaforms, non-placental eutherians, marsupials, and монотремы — as well as in the цинодонт therapsids that are closest to mammals. Their function is to stiffen the body during locomotion.[65] This stiffening would be harmful in pregnant placentals, whose abdomens need to expand.[66]
- A narrow pelvic outlet, which indicates that the young were very small at birth and therefore pregnancy was short, as in modern marsupials. This suggests that the placenta was a later development.
- Five incisors in each side of the upper jaw. This number is typical of metatherians, and the maximum number in modern placentals is three, except for homodonts, такой как броненосец. Но Eomaia's molar to премоляр ratio (it has more pre-molars than molars) is typical of eutherians, including placentals, and not normal in marsupials.
Eomaia также есть Меккелевская бороздка, a primitive feature of the lower jaw that is not found in modern placental mammals.
These intermediate features are consistent with молекулярная филогенетика estimates that the placentals diversified about 110M years ago, 15M years after the date of the Eomaia ископаемое.
Eomaia also has many features that strongly suggest it was a climber, including several features of the feet and toes; well-developed attachment points for muscles that are used a lot in climbing; and a tail that is twice as long as the rest of the spine.
Placentals' best-known feature is their method of reproduction:
- The embryo attaches itself to the матка via a large плацента via which the mother supplies food and oxygen and removes waste products.
- Pregnancy is relatively long and the young are fairly well-developed at birth. In some species (especially herbivores living on plains) the young can walk and even run within an hour of birth.
It has been suggested that the evolution of placental reproduction was made possible by ретровирусы который:[67]
- make the interface between the placenta and uterus into a синцитий, i.e. a thin layer of cells with a shared external membrane. This allows the passage of oxygen, nutrients and waste products, but prevents the passage of blood and other cells that would cause the mother's иммунная система атаковать плод.
- reduce the aggressiveness of the mother's immune system, which is good for the foetus but makes the mother more vulnerable to infections.
From a paleontologist's point of view, eutherians are mainly distinguished by various features of their teeth,[68] ankles and feet.[69]
Expansion of ecological niches in the Mesozoic


Generally speaking, most species of mammaliaforms did occupy the niche of small, nocturnal insectivores, but recent finds, mainly in Китай, show that some species and especially crown group mammals were larger and that there was a larger variety of lifestyles than previously thought. Например:
- Adalatherium hui is a large sized, erect limbed herbivore from the Меловой из Мадагаскар.[70]
- Castorocauda, a member of Docodonta which lived in the middle Jurassic about 164 million years, was about 42.5 cm (16.7 in) long, weighed 500–800 g (18–28 oz), had a бобр -like tail that was adapted for swimming, limbs adapted for swimming and digging, and teeth adapted for eating fish.[36] Another docodont, Халданодон, also had semi-aquatic habits, and indeed aquatic tendencies were probably common among docodonts based on their prevalence in wetland environments.[71] В eutriconodonts Ляоконодон и Яноконодон have more recently also have been suggested to be freshwater swimmers, lacking Castorocauda's powerful tail but possessing paddle-like limbs;[72] the eutriconodont Астроконодон has similarly been suggested as being semi-aquatic in the past, albeit to less convincing evidence.
- Мультитуберкулезные are allotherians that survived for over 125 million years (from mid-Jurassic, about 160M years ago, to late эоцен, about 35M years ago) are often called the "rodents of the Mesozoic". As noted above, they may have given birth to tiny live neonates rather than laying eggs.
- Фруитафоссор, from the late Jurassic period about 150 million years ago, was about the size of a chipmunk and its teeth, forelimbs and back suggest that it broke open the nest of social insects to prey on them (probably термиты, as ants had not yet appeared).[73]
- Точно так же gobiconodontid Spinolestes possessed adaptations for fossoriality and convergent traits with placental ксенартранс like scutes and xenarthrous vertebrae, so it too might have had anteater like habits. It is also notable for the presence of quills akin to those of modern колючие мыши.
- Волатикотерий, from the boundary the early Cretaceous about 125M years ago, is the earliest-known gliding mammal and had a gliding membrane that stretched out between its limbs, rather like that of a modern белка-летяга. This also suggests it was active mainly during the day.[74] Тесно связанные Арджентоконодон also shows similar adaptations that may also suggest aerial locomotion.[75]
- Репеномам, а эвтриконодонт from the early Cretaceous 130 million years ago, was a stocky, барсук -like predator that sometimes preyed on young dinosaurs. Two species have been recognized, one more than 1 m (39 in) long and weighing about 12–14 kg (26–31 lb), the other less than 0.5 m (20 in) long and weighing 4–6 kg (8.8–13.2 lb).[76][77]
- Schowalteria is a Late Cretaceous species almost as large if not larger than R. giganticus that shows speciations towards herbivory, comparable to those of modern ungulates.
- Zhelestidae is a lineage of Late Cretaceous herbivorous евтерийцы, to the point of being mistaken for stem-копытные.[78]
- По аналогии, mesungulatids are also fairly large sized herbivorous mammals from the Late Cretaceous
- Deltatheroidans мы метатерии that were specialised towards carnivorous habits,[79][80] and possible forms like Oxlestes и Khudulestes might have been among the largest Mesozoic mammals, though their status as deltatheroidans is questionable.
- Ихтиоконодон, a eutriconodont from the Берриасский of Morocco, is currently known from molariforms found in marine deposits. These teeth are sharp-cusped and similar in shape to those of piscivorous mammals, and unlike the teeth of contemporary mammals they do not show degradation, so rather than being carried down by river deposits the animal died на месте or close. This has been taken to mean that it was a морское млекопитающее, likely one of the few examples known from the Mesozoic.[81] Alternatively, its close relations to Волатикотерий и Арджентоконодон might suggest that it was a flying mammal.[75]
- Дидельфодон is a Late Cretaceous riverine species of stagodontid marsupialiform с дурофаг dentition, robust jaws similar to a modern Тасманский дьявол, and a postcranial skeleton very similar in size and shape to an otter. This animal has been lauded as the strongest bite of all Mesozoic mammals. It possibly specialized on eating пресноводные крабы и моллюски.
- Tracks of a raccoon-sized млекопитающее representing the morphofamily Ameghinichnidae are described from the Раннемеловой период (поздно Аптян ) Calonda Formation (Ангола ) к Матеус и другие. (2017), who name a new ichnotaxon Catocapes angolanus.[31]
Evolution of major groups of living mammals
There are currently vigorous debates between traditional палеонтологи и molecular phylogeneticists about how and when the modern groups of mammals diversified, especially the placentals. Generally, the traditional paleontologists date the appearance of a particular group by the earliest known fossil whose features make it likely to be a member of that group, while the molecular phylogeneticists suggest that each lineage diverged earlier (usually in the Cretaceous) and that the earliest members of each group were anatomically very similar to early members of other groups and differed only in their генетика. These debates extend to the definition of and relationships between the major groups of placentals.
Molecular phylogenetics-based family tree of placental mammals
Молекулярная филогенетика uses features of organisms' гены to work out family trees in much the same way as paleontologists do with features of fossils — if two organisms' genes are more similar to each other than to those of a third organism, the two organisms are more closely related to each other than to the third.
Molecular phylogeneticists have proposed a family tree that is both broadly similar to but has notable differences from that of the paleontologists. Like paleontologists, molecular phylogeneticists have differing ideas about various details, but here is a typical family tree according to molecular phylogenetics:[82][83] Note that the diagram shown here omits extinct groups, as one cannot extract DNA from fossils.
| Евтерия |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||



Here are the most significant of the differences between this family tree and the one familiar to paleontologists:
- The top-level division is between Atlantogenata and Boreoeutheria, instead of between Xenarthra and the rest. However, analysis of сменный элемент insertions supports a three-way top-level split between Xenarthra, Afrotheria and Boreoeutheria [84][85] and the Atlantogenata clade does not receive significant support in recent distance-based molecular phylogenetics.[86]
- Afrotheria contains several groups that are only distantly related according to the paleontologists' version: Afroinsectiphilia ("African insectivores"), Tubulidentata (aardvarks, which paleontologists regard as much closer to odd-toed ungulates than to other members of Afrotheria), Macroscelidea (elephant shrews, usually regarded as close to rabbits and rodents). The only members of Afrotheria that paleontologists would regard as closely related are Hyracoidea (hyraxes), Proboscidea (elephants) and Sirenia (manatees, dugongs).
- Insectivores are split into three groups: one is part of Afrotheria and the other two are distinct sub-groups within Boreoeutheria.
- Bats are closer to Carnivora and odd-toed ungulates than to Primates and Dermoptera (colugos).
- Perissodactyla (odd-toed ungulates) are closer to Carnivora and bats than to Artiodactyla (even-toed ungulates).
The grouping together of the Afrotheria has some geological justification. All surviving members of the Afrotheria originate from South American or (mainly) African lineages — even the Индийский слон, which diverged from an African lineage about 7.6 миллион лет назад.[87] В качестве Пангея broke up, Africa and South America separated from the other continents less than 150M years ago, and from each other between 100M and 80M years ago.[88][89] So it would not be surprising if the earliest eutherian immigrants into Africa and South America were isolated there and излученный into all the available экологические ниши.
Nevertheless, these proposals have been controversial. Paleontologists naturally insist that fossil evidence must take priority over deductions from samples of the DNA of modern animals. More surprisingly, these new family trees have been criticised by other molecular phylogeneticists, sometimes quite harshly:[90]
- Митохондриальная ДНК 's mutation rate in mammals varies from region to region — some parts hardly ever change and some change extremely quickly and even show large variations between individuals within the same species.[91][92]
- Mammalian mitochondrial DNA mutates so fast that it causes a problem called "saturation", where random noise drowns out any information that may be present. If a particular piece of mitochondrial DNA mutates randomly every few million years, it will have changed several times in the 60 to 75M years since the major groups of placental mammals diverged.[93]
Timing of placental evolution
Recent molecular phylogenetic studies suggest that most placental заказы diverged late in the Cretaceous period, about 100 to 85 million years ago, but that modern families first appeared later, in the late Eocene and early Миоцен эпохи Кайнозойский период.[94][95] Fossil-based analyses, on the contrary, limit the placentals to the Cenozoic.[96] Many Cretaceous fossil sites contain well-preserved lizards, salamanders, birds, and mammals, but not the modern forms of mammals. It is likely that they simply did not exist, and that the молекулярные часы runs fast during major evolutionary radiations.[97] On the other hand, there is fossil evidence from 85 миллион лет назад из копытный mammals that may be ancestors of modern копытные.[98]
Fossils of the earliest members of most modern groups date from the Палеоцен, a few date from later and very few from the Cretaceous, before the extinction of the dinosaurs. But some paleontologists, influenced by molecular phylogenetic studies, have used statistical methods to экстраполировать назад from fossils of members of modern groups and concluded that приматы arose in the late Cretaceous.[99] However, statistical studies of the fossil record confirm that mammals were restricted in size and diversity right to the end of the Cretaceous, and rapidly grew in size and diversity during the Early Paleocene.[100][101]
Evolution of mammalian features
Jaws and middle ears
Адрокодиум, whose fossils date from the early Jurassic, provides the first clear evidence of fully mammalian jaw joints and middle ears, in which the jaw joint is formed by the dentary и чешуйчатый bones while the суставной и квадратный move to the middle ear, where they are known as the incus и молоточек.
One analysis of the monotreme Тейнолофос suggested that this animal had a pre-mammalian jaw joint formed by the угловатый and quadrate bones and that the definitive mammalian middle ear evolved twice independently, in монотремы И в терианец mammals, but this idea has been disputed.[102] In fact, two of the suggestion's authors co-authored a later paper that reinterpreted the same features as evidence that Тейнолофос was a full-fledged утконос, which means it would have had a mammalian jaw joint and middle ear.[46]
Кормление грудью
It has been suggested that lactation's original function was to keep eggs moist. Much of the argument is based on монотремы (egg-laying mammals):[103][104][105]
- While the amniote egg is usually described as able to evolve away from water, most reptile eggs actually need moisture if they are not to dry out.
- Monotremes do not have nipples, but secrete milk from a hairy patch on their bellies.
- During incubation, monotreme eggs are covered in a sticky substance whose origin is not known. Before the eggs are laid, their shells have only three layers. Afterwards, a fourth layer appears with a composition different from that of the original three. The sticky substance and the fourth layer may be produced by the mammary glands.
- If so, that may explain why the patches from which monotremes secrete milk are hairy. It is easier to spread moisture and other substances over the egg from a broad, hairy area than from a small, bare nipple.
Later research demonstrated that caseins already appeared in the common mammalian ancestor approximately 200–310 million years ago.[106] The question of whether secretions of a substance to keep eggs moist translated into actual lactation in therapsids is open. A small mammaliomorph called Sinocodon, generally assumed to be the sister group of all later mammals, had front teeth in even the smallest individuals. Combined with a poorly ossified jaw, they very probably did not suckle.[107] Thus suckling may have evolved right at the pre-mammal/mammal transition. Тем не мение, tritylodontids, generally assumed to be more basal, show evidence of suckling.[108] Morganucodontans, also assumed to be basal Млекопитающие, also show evidence of lactation.[109]
Пищеварительная система
The evolution of the digestive system has formed a significant influence in mammal evolution. With the emergence of mammals, the digestive system was modified in a variety of ways depending on the animal's diet. For example, cats and most carnivores have simple large intestines, while the horse as a herbivore has a voluminous large intestine.[110] An ancestral feature of жвачные животные is their multi-chambered (usually four-chambered) stomach, which evolved about 50 million years ago.[111] Along with morphology of the gut, gastric acidity has been proposed as a key factor shaping the diversity and composition of microbial communities found in the vertebrate gut. Comparisons of stomach acidity across trophic groups in mammal and bird taxa show that scavengers and carnivores have significantly higher stomach acidities compared to herbivores or carnivores feeding on phylogenetically distant prey such as insects or fish.[112]
Despite the lack of fossilization of the gut, microbial evolution of the gut can be inferred from the interrelationships of existing animals, microbes and probable foodstuffs.[113] Mammals are metagenomic, in that they are composed of not only their own genes, but also those of all of their associated microbes.[114] Микробиота кишечника has co-diversified as mammalian species have evolved. Recent studies indicate that adaptive divergence between mammalian species is shaped in part by changes in the gut microbiota.[115][116] The house mouse may have evolved not only with, but also in response to, the unique bacteria inhabiting its gut.[117]
Hair and fur
The first clear evidence of hair or fur is in fossils of Castorocauda и Мегаконус, from 164M years ago in the mid-Jurassic.[36] As both mammals Мегаконус и Castorocauda have a double coat of hair, with both guard hairs and an undercoat, it may be assumed that their last common ancestor did as well. This animal must have been Triassic as it was an ancestor of the Triassic Тикитериум.[32] More recently, the discovery of hair remnants in Permian coprolites pushes back the origin of mammalian hair much further back in the synapsid line to Палеозой терапсиды.[118]
In the mid-1950s, some scientists interpreted the foramina (passages) in the максиллы (upper jaws) and предчелюстные кости (small bones in front of the maxillae) of цинодонты as channels that supplied blood vessels and nerves to vibrissae (усы ) and suggested that this was evidence of hair or fur.[119][120] It was soon pointed out, however, that foramina do not necessarily show that an animal had vibrissae; the modern lizard Tupinambis has foramina that are almost identical to those found in the non-mammalian cynodont Тринаксодон.[12][121] Popular sources, nevertheless, continue to attribute whiskers to Тринаксодон.[122] А след окаменелости from the Lower Triassic had been erroneously regarded as a cynodont footprint showing hair,[123] but this interpretation has been refuted.[124] A study of cranial openings for facial nerves connected whiskers in extant mammals indicate the Прозостродонтия, small immediate ancestors of mammals, presented whiskers similar to mammals, but that less advanced therapsids would either have immobile whiskers or no whisker at all.[125] Fur may have evolved from whiskers.[126] Whiskers themselves may have evolved as a response to nocturnal and/or burrowing lifestyle.
Ruben & Jones (2000) note that the Железы Хардера, которые выделяют липиды for coating the fur, were present in the earliest mammals like Морганукодон, but were absent in near-mammalian therapsids like Тринаксодон.[127] В Msx2 gene associated with hair follicle maintenance is also linked to the closure of the parietal eye in mammals, indicating that fur and lack of pineal eye is linked. The pineal eye is present in Тринаксодон, but absent in more advanced cynognaths (the Пробаиногнатия ).[125]
Insulation is the "cheapest" way to maintain a fairly constant body temperature, without consuming energy to produce more body heat. Therefore, the possession of hair or fur would be good evidence of homeothermy, but would not be such strong evidence of a high metabolic rate.[128][129]
Erect limbs
Understanding of the evolution of erect limbs in mammals is incomplete — living and fossil монотремы have sprawling limbs. Some scientists think that the parasagittal (non-sprawling) limb posture is limited to the Бореосфенида, a group that contains the therians but not, for example, the multituberculates. In particular, they attribute a parasagittal stance to the therians Sinodelphys и Eomaia, which means that the stance had arisen by 125 million years ago, in the Раннемеловой период. However, they also discuss that earlier mammals had more erect forelimbs as opposed to the more sprawling hindlimbs, a trend still continued to some extent in modern placentals and marsupials.[130]
Warm-bloodedness
"Warm-bloodedness " is a complex and rather ambiguous term, because it includes some or all of the following:
- Эндотермия, the ability to generate heat internally rather than via behaviors such as basking or muscular activity.
- Гомеотермия, maintaining a fairly constant body temperature. Наиболее ферменты have an optimum operating temperature; efficiency drops rapidly outside the preferred range. A homeothermic organism needs only to possess enzymes that function well in a small range of temperatures.
- Tachymetabolism, maintaining a high metabolic rate, particularly when at rest. This requires a fairly high and stable body temperature because of the Q10 эффект: biochemical processes run about half as fast if an animal's temperature drops by 10 °C.
Since scientists cannot know much about the internal mechanisms of extinct creatures, most discussion focuses on homeothermy and tachymetabolism. However, it is generally agreed that endothermy first evolved in non-mammalian synapsids such as дицинодонты, which possess body proportions associated with heat retention,[131] high vascularised bones with Гаверсовские каналы,[132] and possibly hair.[133] More recently, it has been suggested that endothermy evolved as far back as Офиакодон.[134]
Современное монотремы have a low body temperature compared to marsupials and placental mammals, around 32 °C (90 °F).[135] Филогенетический брекетинг suggests that the body temperatures of early crown-group mammals were not less than that of extant monotremes. Есть цитологический evidence that the low metabolism of monotremes is a secondarily evolved trait.[136]
Respiratory turbinates
Modern mammals have respiratory turbinates, convoluted structures of thin bone in the nasal cavity. These are lined with слизистые оболочки that warm and moisten inhaled air and extract heat and moisture from exhaled air. An animal with respiratory turbinates can maintain a high rate of breathing without the danger of drying its lungs out, and therefore may have a fast metabolism. Unfortunately these bones are very delicate and therefore have not yet been found in fossils. But rudimentary ridges like those that support respiratory turbinates have been found in advanced Triassic цинодонты, Такие как Тринаксодон и Диадемодон, which suggests that they may have had fairly high metabolic rates.[119][137][138]
Bony secondary palate
Mammals have a secondary bony palate, which separates the respiratory passage from the mouth, allowing them to eat and breathe at the same time. Secondary bony palates have been found in the more advanced cynodonts and have been used as evidence of high metabolic rates.[119][120][139] But some cold-blooded vertebrates have secondary bony palates (crocodilians and some lizards), while birds, which are warm-blooded, do not.[12]
Диафрагма
A muscular диафрагма helps mammals to breathe, especially during strenuous activity. For a diaphragm to work, the ribs must not restrict the abdomen, so that expansion of the chest can be compensated for by reduction in the volume of the abdomen and наоборот. Diaphragms are known in Идентификатор дела pelycosaurs, indicating an early origin within synapsids, though they were still fairly inefficient and likely required support from other muscle groups and limb motion.[140]
The advanced cynodonts have very mammal-like rib cages, with greatly reduced lumbar ribs. This suggests that these animals had more developed diaphragms, were capable of strenuous activity for fairly long periods and therefore had high metabolic rates.[119][120] On the other hand, these mammal-like rib cages may have evolved to increase agility.[12] However, the movement of even advanced therapsids was "like a wheelbarrow", with the hindlimbs providing all the thrust while the forelimbs only steered the animal, in other words advanced therapsids were not as agile as either modern mammals or the early dinosaurs.[5] So the idea that the main function of these mammal-like rib cages was to increase agility is doubtful.
Limb posture
В therapsids had sprawling forelimbs and semi-erect hindlimbs.[120][141] Это говорит о том, что Ограничение перевозчика would have made it rather difficult for them to move and breathe at the same time, but not as difficult as it is for animals such as lizards, which have completely sprawling limbs.[142] Advanced therapsids may therefore have been significantly less active than modern mammals of similar size and so may have had slower metabolisms overall or else been bradymetabolic (lower metabolism when at rest).
Мозг
Mammals are noted for their large brain size relative to body size, compared to other animal groups. Recent findings suggest that the first brain area to expand was that involved in smell.[143] Scientists scanned the skulls of early mammal species dating back to 190–200 million years ago and compared the brain case shapes to earlier pre-mammal species; they found that the brain area involved in the sense of smell was the first to enlarge.[143] This change may have allowed these early mammals to hunt insects at night when dinosaurs were not active.[143]
Смотрите также
- Эволюция приматов
- Evolution of ungulates
- Геномное разнообразие и эволюция кариотипа млекопитающих
- List of examples of convergent evolution in mammals
- Джурамая
Рекомендации
- ^ ROUGIER, GUILLERMO W.; MARTINELLI, AGUSTÍN G.; FORASIEPI, ANALÍA M.; NOVACEK, MICHAEL J. (2007). "New Jurassic Mammals from Patagonia, Argentina: A Reappraisal of Australosphenidan Morphology and Interrelationships". Американский музей Novitates. 3566 (1): 1. Дои:10.1206/0003-0082(2007)507[1:NJMFPA]2.0.CO;2. HDL:2246/5857.
- ^ Waggoner B (February 2, 1997). "Introduction to the Synapsida". Музей палеонтологии Калифорнийского университета. Получено 28 апреля, 2012.
- ^ а б c White AT (May 18, 2005). "Amniota – Palaeos". Архивировано из оригинал 20 декабря 2010 г.. Получено 23 января, 2012.
- ^ а б Mammalia: Overview – Palaeos В архиве 15 июня 2008 г. Wayback Machine
- ^ а б Cowen R (2000). История Жизни. Оксфорд: Blackwell Science. п. 432. ISBN 978-0-7266-0287-0.
- ^ K. A. Kermack; Frances Mussett; H. W. RIgney (January 1981). "Череп Морганукодон". Зоологический журнал Линнеевского общества. 71 (1): 148. Дои:10.1111/j.1096-3642.1981.tb01127.x.
- ^ а б Kemp TS (2005). Происхождение и эволюция млекопитающих. Издательство Оксфордского университета. п. 3. ISBN 978-0-19-850760-4.
- ^ Carroll R.L. (1991): The origin of reptiles. In: Schultze H.-P., Trueb L., (ed) Origins of the higher groups of tetrapods — controversy and consensus. Ithaca: Cornell University Press, pp 331-353.
- ^ "Synapsida: Varanopseidae – Palaeos". Получено 15 октября 2013.
- ^ а б "Therapsida – Palaeos". Архивировано из оригинал on 2007-04-15.
- ^ Kermack DM, Kermack KA (1984). The evolution of mammalian characters. Крум Шлем. ISBN 978-0709915348.
- ^ а б c d Bennett AF, Ruben JA (1986). "The metabolic and thermoregulatory status of therapsids". In Hotton N, MacLean PD, Roth JJ, Roth EC (eds.). The ecology and biology of mammal-like reptiles. Washington: Smithsonian Institution Press, Washington. pp. 207–218.
- ^ "Therapsida: Biarmosuchia – Palaeos". Получено 16 октября 2013.
- ^ "Therapsida: Dinocephalia". Палеос.
- ^ "Ammodontia – Palaeos". Получено 16 октября 2013.
- ^ "Theriodontia – Paleos". Получено 2013-10-15.
- ^ "Cynodontia Overview – Palaeos".
- ^ GROENEWALD, G. H.; WELMAN, J.; MACEACHERN, J. A. (1 April 2001). "Vertebrate Burrow Complexes from the Early Triassic Cynognathus Zone (Driekoppen Formation, Beaufort Group) of the Karoo Basin, South Africa". ПАЛАИ. 16 (2): 148–160. Bibcode:2001Palai..16..148G. Дои:10.1669/0883-1351(2001)016<0148:VBCFTE>2.0.CO;2.
- ^ "Olenekian Age of the Triassic – Palaeos". Архивировано из оригинал on 2007-04-16. Получено 2007-04-18.
- ^ а б Benton MJ (2004). Палеонтология позвоночных (3-е изд.). Оксфорд: Blackwell Science. ISBN 978-0-632-05637-8.
- ^ Campbell JW (1979). Prosser CL (ed.). Comparative Animal Physiology (3-е изд.). W. B. Sauders. pp. 279–316.
- ^ Даррен Нейш, Episode 38: A Not Too Shabby Podcarts
- ^ Oliveira TV, Soares MB, Schultz CL (2010). "Trucidocynodon riograndensis gen. nov. et sp. nov. (Eucynodontia), a new cynodont from the Brazilian Upper Triassic (Santa Maria Formation)". Zootaxa. 2382: 1–71. Дои:10.11646/zootaxa.2382.1.1.
- ^ Kielan-Jaworowska et al. (2004), p.5
- ^ Ruben, J.A.; Jones, T.D. (2000). «Селективные факторы, связанные с происхождением меха и перьев». Американский зоолог. 40 (4): 585–596. Дои:10.1093 / icb / 40.4.585.
- ^ Rowe TB, Macrini TE, Luo ZX (May 2011). "Fossil evidence on origin of the mammalian brain". Наука. 332 (6032): 955–7. Bibcode:2011Sci...332..955R. Дои:10.1126/science.1203117. PMID 21596988. S2CID 940501.
- ^ Raichle ME, Gusnard DA (August 2002). "Appraising the brain's energy budget". Труды Национальной академии наук Соединенных Штатов Америки. 99 (16): 10237–9. Bibcode:2002PNAS...9910237R. Дои:10.1073/pnas.172399499. ЧВК 124895. PMID 12149485.
- ^ Khamsi, Roxanne (13 September 2006). "Brain power". Новый ученый. Архивировано из оригинал 15 апреля 2009 г.
- ^ Vorobyev, M. (2006). "Evolution of colour vision: The story of lost visual pigments". Восприятие. 35. Архивировано из оригинал на 2014-10-06. Получено 2012-01-26.
- ^ Gerkema MP, Davies WI, Foster RG, Menaker M, Hut RA (August 2013). "The nocturnal bottleneck and the evolution of activity patterns in mammals". Ход работы. Биологические науки. 280 (1765): 20130508. Дои:10.1098/rspb.2013.0508. ЧВК 3712437. PMID 23825205.
- ^ а б Mateus O, Marzola M, Schulp AS, Jacobs LL, Polcyn MJ, Pervov V, Gonçalves AO, Morais ML (2017). "Angolan ichnosite in a diamond mine shows the presence of a large terrestrial mammaliamorph, a crocodylomorph, and sauropod dinosaurs in the Early Cretaceous of Africa". Палеогеография, палеоклиматология, палеоэкология. 471: 220–32. Bibcode:2017PPP...471..220M. Дои:10.1016/j.palaeo.2016.12.049.
- ^ а б c Luo ZX (December 2007). "Transformation and diversification in early mammal evolution". Природа. 450 (7172): 1011–9. Bibcode:2007Natur.450.1011L. Дои:10.1038/nature06277. PMID 18075580. S2CID 4317817.
- ^ Rowe, Timothy (23 September 1988). "Definition, diagnosis, and origin of Mammalia". Журнал палеонтологии позвоночных. 8 (3): 241–264. Дои:10.1080/02724634.1988.10011708.
- ^ "Microlestes rhaeticus Dawkins 1864 (mammal)". База данных палеобиологии. Получено 30 января, 2012.
- ^ "Morganucodontids & Docodonts – Palaeos". Архивировано из оригинал on 2007-04-16.
- ^ а б c Ji Q, Luo ZX, Yuan CX, Tabrum AR (February 2006). "A swimming mammaliaform from the Middle Jurassic and ecomorphological diversification of early mammals". Наука. 311 (5764): 1123–7. Bibcode:2006Sci ... 311.1123J. Дои:10.1126/science.1123026. PMID 16497926. S2CID 46067702. See also the news item at "Jurassic "Beaver" Found; Rewrites History of Mammals".
- ^ Luo ZX, Crompton AW, Sun AL (May 2001). "A new mammaliaform from the early Jurassic and evolution of mammalian characteristics" (PDF). Наука. 292 (5521): 1535–40. Bibcode:2001Sci...292.1535L. Дои:10.1126/science.1058476. PMID 11375489. S2CID 8738213. Архивировано из оригинал (PDF) на 20.08.2013.
- ^ а б Джейкобс, Джеральд Х. (12 октября 2009 г.). «Эволюция цветового зрения у млекопитающих». Философские труды Королевского общества B: биологические науки. 364 (1531): 2957–2967. Дои:10.1098 / rstb.2009.0039. ЧВК 2781854. PMID 19720656.
- ^ Бейлс, Хелена Дж; Дэвис, Уэйн Л; Trezise, Ann EO; Коллин, Шон П. (2007). «Визуальные пигменты в живом ископаемом, австралийской двоякодышащей рыбе Neoceratodus forsteri». BMC Эволюционная биология. 7 (1): 200. Дои:10.1186/1471-2148-7-200. ЧВК 2194722. PMID 17961206.
- ^ Коллин, Шон П. (2010). «Эволюция и экология фоторецепции сетчатки у ранних позвоночных». Мозг, поведение и эволюция. 75 (3): 174–185. Дои:10.1159/000314904. PMID 20733293. S2CID 22974478.
- ^ Dulai, Kanwaljit S .; Дорнум, Миранда фон; Mollon, John D .; Хант, Дэвид М. (1 июля 1999 г.). «Эволюция трихроматического цветового зрения путем дупликации гена опсина у приматов Нового и Старого Света». Геномные исследования. 9 (7): 629–638. Дои:10.1101 / гр. 9.7.629 (неактивно 10.11.2020). PMID 10413401.CS1 maint: DOI неактивен по состоянию на ноябрь 2020 г. (связь)
- ^ Аррезе, Екатерина А .; Харт, Натан С .; Томас, Николь; Бизли, Лин Д .; Шанд, Юлия (апрель 2002 г.). «Трихроматия у австралийских сумчатых». Текущая биология. 12 (8): 657–660. Дои:10.1016 / s0960-9822 (02) 00772-8. PMID 11967153. S2CID 14604695.
- ^ а б «Млекопитающие - Палеос». Архивировано из оригинал на 2007-04-12.
- ^ а б Джейкобс Л.Л., Винклер Д.А., Марри П.А. (июль 1989 г.). «Происхождение современных млекопитающих: ступени эволюции в раннем меловом периоде Северной Америки». Труды Национальной академии наук Соединенных Штатов Америки. 86 (13): 4992–5. Bibcode:1989PNAS ... 86.4992J. Дои:10.1073 / pnas.86.13.4992. JSTOR 34031. ЧВК 297542. PMID 2740336.
- ^ Раухут О.В., Мартин Т., Ортис-Хорегуисар Э., Пуэрта П. (март 2002 г.). «Юрское млекопитающее из Южной Америки». Природа. 416 (6877): 165–8. Bibcode:2002Натура416..165р. Дои:10.1038 / 416165a. PMID 11894091. S2CID 4346804.
- ^ а б c Rowe T, Rich TH, Vickers-Rich P, Springer M, Woodburne MO (январь 2008 г.). «Самый старый утконос и его влияние на время расхождения клад утконоса и ехидны». Труды Национальной академии наук Соединенных Штатов Америки. 105 (4): 1238–42. Bibcode:2008ПНАС..105.1238Р. Дои:10.1073 / pnas.0706385105. ЧВК 2234122. PMID 18216270.
- ^ Филлипс MJ, Беннетт TH, Ли MS (октябрь 2009 г.). «Молекулы, морфология и экология указывают на недавнее амфибийное происхождение ехидн». Труды Национальной академии наук Соединенных Штатов Америки. 106 (40): 17089–94. Bibcode:2009PNAS..10617089P. Дои:10.1073 / pnas.0904649106. ЧВК 2761324. PMID 19805098.
- ^ а б "Аппендикулярного скелета".
- ^ Батлер, П. М. (2000). «Обзор ранних аллотерийских млекопитающих» (PDF). Acta Palaeontologica Polonica. 45 (4): 317–342.
- ^ Уайт, А. Т. (21 мая 2005 г.). "Mammaliaformes - Palaeos". Архивировано из оригинал 20 декабря 2010 г.. Получено 20 января, 2012.
- ^ Kielan-Jaworowska et al. (2004), стр. 299
- ^ «Млекопитающие: Spalacotheroida и Cladotheria - Palaeos».
- ^ «Метатерия - Палеос».
- ^ Салай Ф.С., Трофимов Б.А. (1996). «Монгольский позднемеловой азиатерий, ранняя филогения и палеобиогеография метатерии». Журнал палеонтологии позвоночных. 16 (3): 474–509. Дои:10.1080/02724634.1996.10011335. Архивировано из оригинал 5 марта 2001 г.
- ^ «Древнейшее ископаемое сумчатое животное, найденное в Китае». National Geographic News. 2003-12-15.
- ^ «Дидельфиморфия - Палеос». Получено 2013-10-15.
- ^ «Семейство Peramelidae (бандикуты и эхимиперы)».
- ^ «Вид такой же, как и вид… Часть II». 2005-12-12.
- ^ «Сумчатые». Архивировано из оригинал 5 апреля 2003 г.
- ^ Новачек MJ, Rougier GW, Wible JR, McKenna MC, Dashzeveg D, Horovitz I (октябрь 1997 г.). «Надлобковые кости у здоровых млекопитающих из позднего мела Монголии». Природа. 389 (6650): 483–6. Bibcode:1997Натура.389..483Н. Дои:10.1038/39020. PMID 9333234. S2CID 205026882.
- ^ Белый TD (август 1989 г.). «Анализ функции надлобковой кости у млекопитающих с использованием теории масштабирования». Журнал теоретической биологии. 139 (3): 343–57. Дои:10.1016 / S0022-5193 (89) 80213-9. PMID 2615378.
- ^ Луо ZX, Юань CX, Мэн QJ, Цзи Q (август 2011). «Юрское эвтериальное млекопитающее и расхождение сумчатых и плацентарных» (PDF). Природа. 476 (7361): 442–5. Bibcode:2011Натура.476..442L. Дои:10.1038 / природа10291. PMID 21866158. S2CID 205225806.
- ^ "Eomaia scansoria: открытие старейшего известного плацентарного млекопитающего".
- ^ О'Лири М.А., Блох Дж. И., Флинн Дж. Дж., Годен Т. Дж., Джалломбардо А., Джаннини Н. П. и др. (Февраль 2013). «Предок плацентарных млекопитающих и пост-K-Pg-излучение плаценты». Наука. 339 (6120): 662–7. Bibcode:2013Наука ... 339..662O. Дои:10.1126 / science.1229237. HDL:11336/7302. PMID 23393258. S2CID 206544776.
- ^ Рейли С.М., Белый TD (январь 2003 г.). «Гипаксиальные двигательные паттерны и функция надлобковых костей у примитивных млекопитающих». Наука. 299 (5605): 400–2. Bibcode:2003Наука ... 299..400р. Дои:10.1126 / science.1074905. PMID 12532019. S2CID 41470665.
- ^ Новачек MJ, Rougier GW, Wible JR, McKenna MC, Dashzeveg D, Horovitz I (октябрь 1997 г.). «Надлобковые кости у здоровых млекопитающих из позднего мела Монголии». Природа. 389 (6650): 483–6. Bibcode:1997Натура.389..483Н. Дои:10.1038/39020. PMID 9333234. S2CID 205026882.
- ^ Фокс Д (1999). «Почему мы не откладываем яйца». Новый ученый.
- ^ «Евтерия - Палеос».
- ^ Джи Кью, Луо ZX, Юань С.Х., Wible JR, Чжан Дж. П., Георгий Дж. А. (апрель 2002 г.) «Самое раннее известное млекопитающее эвтерианской породы». Природа. 416 (6883): 816–22. Bibcode:2002Натура.416..816J. Дои:10.1038 / 416816a. PMID 11976675. S2CID 4330626.
- ^ Краузе, Дэвид В .; Хоффманн, Симона; Ху, Яомин; Wible, John R .; Rougier, Guillermo W .; Кирк, Э. Кристофер; Groenke, Joseph R .; Роджерс, Раймонд Р .; Росси, Джеймс Б.; Schultz, Julia A .; Эванс, Алистер Р .; фон Кенигсвальд, Вигхарт; Рахантарисоа, Лидия Дж. (29 апреля 2020 г.). «Скелет мелового млекопитающего с Мадагаскара отражает длительную замкнутость». Природа. 581 (7809): 421–427. Bibcode:2020Натура.581..421K. Дои:10.1038 / с41586-020-2234-8. PMID 32461642. S2CID 216650606.
- ^ Палеонтология и геология верхнеюрской свиты Моррисон: Бюллетень 36.
- ^ Чен, Мэн; Уилсон, Грегори П. (24 февраля 2015 г.). «Многофакторный подход к выводу о локомоторных режимах у мезозойских млекопитающих». Палеобиология. 41 (2): 280–312. Дои:10.1017 / pab.2014.14. S2CID 86087687.
- ^ Луо ZX, Wible JR (апрель 2005 г.). «Позднеюрское роющее млекопитающее и разнообразие ранних млекопитающих». Наука. 308 (5718): 103–7. Bibcode:2005Наука ... 308..103Л. Дои:10.1126 / science.1108875. PMID 15802602. S2CID 7031381.
- ^ Мэн Дж, Ху Й, Ван И, Ван Х, Ли Ц (декабрь 2006 г.). «Мезозойское летающее млекопитающее из Северо-Восточного Китая». Природа. 444 (7121): 889–93. Bibcode:2006Натура 444..889М. Дои:10.1038 / природа05234. PMID 17167478. S2CID 28414039.
- ^ а б Гаэтано Л.К., Ружье Г.В. (2011). «Новые материалы Argentoconodon fariasorum (Mammaliaformes, Triconodontidae) из юрского периода Аргентины и его влияние на филогению триконодонтов». Журнал палеонтологии позвоночных. 31 (4): 829–843. Дои:10.1080/02724634.2011.589877. HDL:11336/68497. S2CID 85069761.
- ^ Li, J .; Wang, Y .; Wang, Y .; Ли, К. (2000). «Новое семейство примитивных млекопитающих из мезозоя западного Ляонина, Китай». Китайский научный бюллетень. 46 (9): 782–785. Bibcode:2001ЧСБУ..46..782Л. Дои:10.1007 / BF03187223. S2CID 129025369. аннотация, на английском языке
- ^ Ху И, Мэн Дж, Ван И, Ли Ц (январь 2005 г.). «Крупные мезозойские млекопитающие питаются молодыми динозаврами» (PDF). Природа. 433 (7022): 149–52. Bibcode:2005Натура.433..149H. Дои:10.1038 / природа03102. PMID 15650737. S2CID 2306428.[постоянная мертвая ссылка ]
- ^ Майкл Дж. Бентон, Михаил А. Шишкин, Дэвид М. Анвин, Эпоха динозавров в России и Монголии
- ^ КРИСТИАН ДЕ МЮЙЗОН и БРИДЖИТ ЛАНЖ-БАДРЕ, Хищные стоматологические адаптации у трибосфеновых млекопитающих и филогенетическая реконструкция, Статья впервые опубликована в Интернете: 29 марта 2007 г. Дои:10.1111 / j.1502-3931.1997.tb00481
- ^ Зофья Киелан-Яворовская; Ричард Л. Чифелли; Чжэ-Си Ло (2004). «Глава 12: Метатерииане». Млекопитающие эпохи динозавров: происхождение, эволюция и строение. Нью-Йорк: издательство Колумбийского университета. С. 425–262. ISBN 978-0-231-11918-4.
- ^ Сигоньо-Рассел, Д. (1995). «Два, возможно, водных триконодонта из раннего мела Марокко». Acta Palaeontologica Polonica. 40 (2): 149–162.
- ^ Мерфи WJ, Eizirik E, O'Brien SJ, Madsen O, Scally M, Douady CJ, et al. (Декабрь 2001 г.). «Разрешение раннего излучения плаценты млекопитающих с использованием байесовской филогенетики». Наука. 294 (5550): 2348–51. Bibcode:2001Научный ... 294.2348M. Дои:10.1126 / science.1067179. PMID 11743200. S2CID 34367609.
- ^ Кригс Дж.О., Чураков Г., Кифманн М., Джордан У., Брозиус Дж., Шмитц Дж. (Апрель 2006 г.). «Ретропозиционные элементы как архив эволюционной истории плацентарных млекопитающих». PLOS Биология. 4 (4): e91. Дои:10.1371 / journal.pbio.0040091. ЧВК 1395351. PMID 16515367. (pdf версия )
- ^ Нисихара Х., Маруяма С., Окада Н. (март 2009 г.). «Анализ ретропозонов и недавние геологические данные свидетельствуют о почти одновременном расхождении трех надотрядов млекопитающих». Труды Национальной академии наук Соединенных Штатов Америки. 106 (13): 5235–40. Bibcode:2009ПНАС..106.5235Н. Дои:10.1073 / pnas.0809297106. ЧВК 2655268. PMID 19286970.
- ^ Чураков Г., Кригс Дж., Бэрч Р., Земанн А., Брозиус Дж., Шмитц Дж. (Май 2009 г.). "Мозаичные схемы вставки ретропозона у плацентарных млекопитающих". Геномные исследования. 19 (5): 868–75. Дои:10.1101 / гр.090647.108. ЧВК 2675975. PMID 19261842.
- ^ Мередит Р.В., Янечка Ю.Е., Гейтси Дж., Райдер О.А., Фишер К.А., Тилинг ЕС и др. (Октябрь 2011 г.). «Влияние меловой земной революции и исчезновения КПГ на диверсификацию млекопитающих». Наука. 334 (6055): 521–4. Bibcode:2011Наука ... 334..521М. Дои:10.1126 / science.1211028. PMID 21940861. S2CID 38120449.
- ^ «Ученые составляют карту эволюции слонов». Новости BBC. 2007-07-24. Получено 2008-08-11.
- ^ Историческая перспектива (Dynamic Earth, USGS)
- ^ Меловая карта
- ^ Обзор насекомоядных - Palaeos В архиве 2007-07-15 на Wayback Machine
- ^ Springer MS, Douzery E (октябрь 1996 г.). «Вторичная структура и закономерности эволюции среди молекул митохондриальной 12S рРНК млекопитающих». Журнал молекулярной эволюции. 43 (4): 357–73. Bibcode:1996JMolE..43..357S. Дои:10.1007 / BF02339010. PMID 8798341. S2CID 12317859.
- ^ Спрингер М.С., Холлар Л.Дж., Бурк А. (ноябрь 1995 г.). «Компенсаторные замены и эволюция гена митохондриальной 12S рРНК у млекопитающих». Молекулярная биология и эволюция. 12 (6): 1138–50. Дои:10.1093 / oxfordjournals.molbev.a040288. PMID 8524047.
- ^ Ли WH (1997). Молекулярная эволюция. Sinauer Associates. ISBN 978-0-87893-266-5.
- ^ Бининда-Эмондс OR, Cardillo M, Jones KE, MacPhee RD, Beck RM, Grenyer R и др. (Март 2007 г.). «Отсроченный рост современных млекопитающих». Природа. 446 (7135): 507–12. Bibcode:2007Натура.446..507Б. Дои:10.1038 / природа05634. PMID 17392779. S2CID 4314965. Архивировано из оригинал на 2008-01-25. Получено 2007-06-24.
- ^ Тарвер Дж. Э., Дос Рейс М., Мирараб С., Моран Р. Дж., Паркер С., О'Рейли Дж. Э. и др. (Январь 2016 г.). «Взаимоотношения плацентарных млекопитающих и пределы филогенетического вывода». Геномная биология и эволюция. 8 (2): 330–44. Дои:10.1093 / gbe / evv261. ЧВК 4779606. PMID 26733575.
- ^ Wible JR, Rougier GW, Novacek MJ, Asher RJ (июнь 2007 г.). «Меловые эвтерианы и лавразийское происхождение плацентарных млекопитающих вблизи границы К / Т» (PDF). Природа. 447 (7147): 1003–6. Bibcode:2007 Натур.447.1003W. Дои:10.1038 / природа05854. PMID 17581585. S2CID 4334424. Архивировано из оригинал (PDF) 23 августа 2012 г.
- ^ Бентон MJ (декабрь 1999 г.). «Раннее происхождение современных птиц и млекопитающих: молекулы против морфологии». BioEssays. 21 (12): 1043–51. Дои:10.1002 / (SICI) 1521-1878 (199912) 22: 1 <1043 :: AID-BIES8> 3.0.CO; 2-B. PMID 10580989.
- ^ Арчибальд Дж. Д. (май 1996 г.). «Ископаемые свидетельства позднемелового происхождения« копытных »млекопитающих». Наука. 272 (5265): 1150–3. Bibcode:1996Научный ... 272.1150A. Дои:10.1126 / science.272.5265.1150. PMID 8662448. S2CID 33510594.
- ^ Мартин Р. Д., Солиго С., Таваре С. (2007). «Происхождение приматов: последствия мелового происхождения». Folia Primatologica; Международный журнал приматологии. 78 (5–6): 277–96. Дои:10.1159/000105145. PMID 17855783. S2CID 6651479. - аналогичная статья этих авторов доступна бесплатно в Интернете по адресу Новый взгляд на даты происхождения и расхождения приматов В архиве 2007-07-20 на Wayback Machine
- ^ Алрой Дж (март 1999 г.). «Летопись окаменелостей североамериканских млекопитающих: свидетельство эволюционной радиации палеоцена». Систематическая биология. 48 (1): 107–18. Дои:10.1080/106351599260472. PMID 12078635.
- ^ Арчибальд Дж. Д., Дойчман Д. Д. (июнь 2001 г.). «Количественный анализ времени возникновения и диверсификации существующих порядков плаценты». Журнал эволюции млекопитающих. 8 (2): 107–124. Дои:10.1023 / А: 1011317930838. S2CID 15581162.
- ^ Rich TH, Hopson JA, Musser AM, Flannery TF, Vickers-Rich P (февраль 2005 г.). «Независимое происхождение костей среднего уха у монотремов и терианов». Наука. 307 (5711): 910–4. Bibcode:2005Наука ... 307..910R. Дои:10.1126 / science.1105717. PMID 15705848. S2CID 3048437. Другие мнения см. В разделе «Технические комментарии» на той же веб-странице.
- ^ Офтедал ОТ (июль 2002 г.). «Молочная железа и ее происхождение в процессе эволюции синапсидов» (PDF). Журнал биологии и неоплазии молочных желез. 7 (3): 225–52. Дои:10.1023 / А: 1022896515287. PMID 12751889. S2CID 25806501. Архивировано из оригинал (PDF) на 2011-12-03. Получено 2012-06-26.
- ^ Офтедал ОТ (июль 2002 г.). «Происхождение лактации как источника воды для яиц с пергаментной скорлупой» (PDF). Журнал биологии и неоплазии молочных желез. 7 (3): 253–66. Дои:10.1023 / А: 1022848632125. PMID 12751890. S2CID 8319185.
- ^ Кормление на яйцах В архиве 14 апреля 2009 г. Wayback Machine
- ^ Borders R, Робертсон WO (июнь 1993 г.). «Вашингтонская панель по пестицидам: процесс и продукт». Ветеринария и токсикология человека. 35 (3): 258–9. PMID 8351802.
- ^ Кромптон, Альфред В .; Ло, Чжэси (1993). Филогения млекопитающих. Нью-Йорк, Нью-Йорк: Springer New York. С. 30–44. DOI: 10.1007 / 978-1-4615-7381-4_4. ISBN 9781461573838.
- ^ Ху Й, Мэн Дж, Кларк Дж. М. (2009). «Новый тритилодонтид из верхней юры Синьцзяна, Китай». Acta Palaeontologica Polonica. 54 (3): 385–391. Дои:10.4202 / приложение.2008.0053.
- ^ Панцироли Э, Бенсон РБ, Уолш С (2017). «Зубной камень Wareolestes rex (Megazostrodontidae): новый образец из Шотландии и его значение для замены морганукодонтального зуба». Статьи по палеонтологии. 3 (3): 373–386. Дои:10.1002 / spp2.1079.
- ^ «Эволюционные тенденции в пищеварительной системе млекопитающих». campus.murraystate.edu. Получено 2019-09-30.
- ^ Hackmann TJ, Испания JN (апрель 2010 г.). «Приглашенный обзор: экология и эволюция жвачных животных: перспективы, полезные для исследований и производства жвачных животных». Журнал молочной науки. 93 (4): 1320–34. Дои:10.3168 / jds.2009-2071. PMID 20338409.
- ^ Бисли Д.Е., Кольц А.М., Ламберт Дж. Э., Фирер Н., Данн Р. Р. (2015-07-29). «Эволюция кислотности желудка и ее значение для микробиома человека». PLOS ONE. 10 (7): e0134116. Bibcode:2015PLoSO..1034116B. Дои:10.1371 / journal.pone.0134116. ЧВК 4519257. PMID 26222383.
- ^ Хьюм И.Д., Warner AC (1980). «Эволюция микробного пищеварения у млекопитающих». В Ruckebusch Y, Thivend P (ред.). Физиология пищеварения и метаболизм у жвачных животных. Физиология пищеварения и метаболизм у жвачных животных: материалы 5-го Международного симпозиума по физиологии жвачных животных, состоявшегося в Клермон-Ферран, 3–7 сентября 1979 г.. Springer Нидерланды. С. 665–684. Дои:10.1007/978-94-011-8067-2_32. ISBN 9789401180672.
- ^ Мёллер А.Х., Сузуки Т.А., Фифер-Рикси М., Нахман М.В. (октябрь 2018 г.). «Способы передачи микробиоты кишечника млекопитающих». Наука. 362 (6413): 453–457. Bibcode:2018Научный ... 362..453M. Дои:10.1126 / science.aat7164. PMID 30361372.
- ^ Moeller AH, Gomes-Neto JC, Mantz S, Kittana H, Segura Munoz RR, Schmaltz RJ, et al. (Июль 2019). "Экспериментальные данные по адаптации к специфической кишечной микробиоте у домашних мышей". мСфера. 4 (4): e00387–19. Дои:10,1128 / мСфера.00387-19. ЧВК 6620377. PMID 31292233.
- ^ Шарптон ТиДжей (24.04.2018). «Роль кишечного микробиома в эволюции позвоночных». mSystems. 3 (2): e00174–17. Дои:10.1128 / mСистем.00174-17. ЧВК 5881020. PMID 29629413.
- ^ «Исследование предполагает, что кишечные бактерии помогли сформировать эволюцию млекопитающих». Phys.org. Получено 2019-09-30.
- ^ П. Байдек, Микробиота и пищевые остатки, включая возможные доказательства наличия волос до млекопитающих в копролитах верхней перми из России, 2015 г., Дои:10.1111 / лет.12156
- ^ а б c d Бринк А.С. (1955). "Исследование скелета Диадемодон". Palaeontologia Africana. 3: 3–39.
- ^ а б c d Кемп Т.С. (1982). Млекопитающие рептилии и происхождение млекопитающих. Лондон: Academic Press. п. 363. ISBN 978-0-12-404120-2.
- ^ Эстес Р. (1961). «Анатомия черепа цинодонтовой рептилии. Thrinaxodon liorhinus". Вестник Музея сравнительной зоологии. 125 (1253): 165–180.
- ^ "Тринаксодон: Новое млекопитающее ". National Geographic Daily News. 11 февраля 2009 г.. Получено 26 августа, 2012.
- ^ Элленбергер, Пол (1976). "Трасса по следам сои épaisses dans le Trias inférieur a moyen de Lodève (Эро, Франция): Cynodontipus polythrix nov. ген., ноя. sp. les Cynodontes en France ". Geobios. 9 (6): 769–787. Дои:10.1016 / S0016-6995 (76) 80078-2.
- ^ Ольсен, Пол Э. (2012): Цинодонтип: Нора проколофонид, а не след волосатых цинодонтов (средний-поздний триас: Европа, Марокко, восточная часть Северной Америки). В архиве 2016-03-09 в Wayback Machine. Программа Северо-восточной секции - 47-е ежегодное собрание (18–20 марта 2012 г.) Хартфорд, Коннектикут, Геологическое общество Америки
- ^ а б Бенуа Дж., Manger PR, Rubidge BS (май 2016 г.). «Палеоневрологические ключи к эволюции определения характеристик мягких тканей млекопитающих». Научные отчеты. 6: 25604. Bibcode:2016НатСР ... 625604Б. Дои:10.1038 / srep25604. ЧВК 4860582. PMID 27157809.
- ^ Свитек, Брайан. «К корням меха». Национальная география. Получено 24 ноября 2014.
- ^ Рубен Дж. А., Джонс Т. Д. (2000). «Селективные факторы, связанные с происхождением меха и перьев» (PDF). Американский зоолог. 40 (4): 585–596. Дои:10.1093 / icb / 40.4.585.
- ^ Шмидт-Нильсен К. (1975). Физиология животных: адаптация и окружающая среда. Кембридж: Издательство Кембриджского университета. п.699. ISBN 978-0-521-38196-3.
- ^ Холка ПК (1992). Сравнительная физиология животных. Форт-Уэрт: Колледж Сондерса. п. 949. ISBN 978-0-03-012847-9.
- ^ Kielan-Jaworowska Z, Hurum JH (2006). «Поза конечностей у ранних млекопитающих: раскидистая или парасагиттальная» (PDF). Acta Palaeontologica Polonica. 51 (3): 10237–10239. Получено 2008-09-24.
- ^ Баккер 1975
- ^ ДЖЕННИФЕР БОТА-БРИНК и КЕННЕТ Д. АНДЖЕЛЧИК, Объясняют ли необычайно высокие темпы роста пермо-триасовых дицинодонтов (Therapsida, Anomodontia) их успех до и после конца пермского вымирания ?, Версия записи онлайн: 26 июля 2010 г. Дои:10.1111 / j.1096-3642.2009.00601.x
- ^ Микробиота и пищевые остатки, включая возможные свидетельства наличия волос до млекопитающих в копролитах верхней перми из России Петр Байдек1, Мартин Кварнстрём2, Кшиштоф Овоцкий3, Томаш Сулей3, Андрей Геннадьевич Сенников4,5, Валерий К. Голубев4,5 и Гжегожский Недёвид впервые опубликованы в Интернете: 25 ноя.2015 Дои:10.1111 / лет.12156
- ^ «Выявлена родословная« теплокровность »млекопитающих». www.sciencedaily.com. Общество палеонтологии позвоночных. 29 октября 2015 г.. Получено 29 октября, 2015.
- ^ Пол Г.С. (1988). Хищные динозавры мира. Нью-Йорк: Саймон и Шустер. п.464. ISBN 978-0-671-61946-6.
- ^ Дж. М. Уотсон; ВАРЕНЬЕ. Могилы (1988). «Монотремные клеточные циклы и эволюция гомеотермии». Австралийский зоологический журнал. 36 (5): 573–584. Дои:10.1071 / ZO9880573.
- ^ Хиллениус WH (1992). «Эволюция носовых раковин и эндотермия млекопитающих». Палеобиология. 18 (1): 17–29. Дои:10.1017 / S0094837300012197. JSTOR 2400978.
- ^ Рубен Дж (1995). «Эволюция эндотермии у млекопитающих и птиц: от физиологии к окаменелостям». Ежегодный обзор физиологии. 57: 69–95. Дои:10.1146 / annurev.ph.57.030195.000441. PMID 7778882.
- ^ Макнаб Б.К. (1978). «Эволюция эндотермии в филогенезе млекопитающих». Американский натуралист. 112 (983): 1–21. Дои:10.1086/283249. S2CID 84070652.
- ^ Маркус Ламбертц и др., Пример эволюции гомолога диафрагмы среди самых ранних синапсидов, Анналы Нью-Йоркской академии наук (2016). Дои:10.1111 / няс.13264
- ^ Дженкинс Ф.А. младший (1971). «Посткраниальный скелет африканских цинодонтов». Бюллетень музея естественной истории Пибоди (36): 1–216.
- ^ Pough FH, Heiser JS, McFarland WN (1996). Позвоночные животные. Нью-Джерси: Прентис-Холл. п.798. ISBN 978-0-02-396370-4.
- ^ а б c Виктория Гилл (20 мая 2011 г.). "Большой мозг млекопитающих развил обоняние". Новости BBC. Получено 22 мая 2011.
дальнейшее чтение
- Роберт Л. Кэрролл, Палеонтология и эволюция позвоночных, W.H. Freeman and Company, Нью-Йорк, 1988 г. ISBN 0-7167-1822-7. Главы с XVII по XXI
- Николас Хоттон III, Пол Д. Маклин, Ян Дж. Рот и Э. Кэрол Рот, редакторы, Экология и биология млекопитающих рептилий, Smithsonian Institution Press, Вашингтон и Лондон, 1986 г. ISBN 0-87474-524-1
- Т. С. Кемп, Происхождение и эволюция млекопитающих, Oxford University Press, Нью-Йорк, 2005 г. ISBN 0-19-850760-7
- Зофья Киелан-Яворовская, Ричард Л. Чифелли и Чжэ-Си Луо, Млекопитающие эпохи динозавров: происхождение, эволюция и структура, Columbia University Press, Нью-Йорк, 2004 г. ISBN 0-231-11918-6. Всесторонний охват от первых млекопитающих до времени Меловое – палеогеновое вымирание.
- Луо ZX (декабрь 2007 г.). «Трансформация и диверсификация в ранней эволюции млекопитающих» (PDF). Природа. 450 (7172): 1011–9. Bibcode:2007 Натур.450.1011L. Дои:10.1038 / природа06277. PMID 18075580. S2CID 4317817. Архивировано из оригинал (PDF) на 24.11.2012. Обзорная статья с 98 ссылками на научную литературу.
внешняя ссылка
- Цинодонтия охватывает несколько аспектов эволюции цинодонтов в млекопитающих с большим количеством ссылок.
- Млекопитающие, Дискуссия BBC Radio 4 с Ричардом Корфилдом, Стивом Джонсом и Джейн Фрэнсис (В наше время, 13 октября 2005 г.)

{kind=link}