Затмение дарвинизма - The eclipse of Darwinism

Джулиан Хаксли использовал фразу «затмение дарвинизма”[а] описать состояние дел до того, что он называл современный синтез, когда эволюция был широко принят в научных кругах, но относительно немногие биологи считали, что естественный отбор был его основным механизмом.[2][3] Историки науки, такие как Питер Дж. Боулер использовали ту же фразу в качестве ярлыка для периода в история эволюционной мысли с 1880-х до примерно 1920 года, когда альтернативы естественный отбор были разработаны и исследованы - поскольку многие биологи считали естественный отбор ошибочным предположением Чарльз Дарвин часть, или, по крайней мере, относительно второстепенная.[4][5] Альтернативный термин, интерфаза дарвинизма, было предложено избежать во многом неверного вывода о том, что предполагаемому затмению предшествовал период энергичных дарвиновских исследований.[1]
Пока было множественные объяснения эволюции включая витализм, катастрофизм, и структурализм В течение XIX века на рубеже XX века действовали четыре основных альтернативы естественному отбору:
- Теистическая эволюция была убеждением, что Бог непосредственно руководит эволюцией.[b]
- Неоламаркизм Идея заключалась в том, что эволюция была вызвана унаследованием характеристик, приобретенных в течение жизни организма.
- Ортогенез была вера в то, что на организмы влияют внутренние силы или законы развития, которые движут эволюцией в определенных направлениях
- Мутационизм Идея заключалась в том, что эволюция в значительной степени является результатом мутаций, которые создают новые формы или виды за один шаг.
К концу XIX века теистическая эволюция в значительной степени исчезла из научной литературы, поскольку прямые призывы к сверхъестественным причинам стали рассматриваться как ненаучные. Другие альтернативы имели значительное распространение еще в 20 веке; основная биология в значительной степени отказалась от них только тогда, когда развитие генетика заставили их казаться все более несостоятельными, и когда развитие популяционная генетика и современный синтез продемонстрировал объяснительную силу естественный отбор. Эрнст Майр писал, что даже в 1930 году большинство учебников по-прежнему подчеркивали такие недарвиновские механизмы.[6]
Контекст
Эволюция получила широкое признание в научных кругах через несколько лет после публикации О происхождении видов, но принятие естественный отбор так как его приводного механизма было намного меньше.[7] В XIX веке этой теории было выдвинуто шесть возражений:[8]

- В Окаменелости был прерывистым, предполагая пробелы в эволюции.[8]
- Физик Лорд Кельвин вычислено в 1862 г., что Земля остыла бы через 100 миллионов лет или меньше от его образования слишком мало времени для эволюции.[c][8]
- Утверждалось, что многие структуры были неадаптивными (лишенными функций), поэтому они не могли развиться при естественном отборе.[8]
- Некоторые структуры, казалось, развивались по регулярному образцу, например, глаза неродственных животных, таких как Кальмар и млекопитающие.[8]
- Утверждалось, что естественный отбор не является творческим, в то время как вариации признавались в основном не имеющими значения.[8]
- Инженер Флиминг Дженкин[d] правильно отметил в 1868 г., рассматривая Происхождение видов, что смешанное наследование одобренный Дарвином будет противодействовать естественному отбору.[e][8][10][11]
И Дарвин, и его ближайший сторонник Томас Генри Хаксли[f] также свободно признал, что отбор не может быть всем объяснением; Дарвин был готов принять некоторую долю ламаркизма, в то время как Хаксли устраивало и то, и другое. внезапное (мутационное) изменение и направленная (ортогенетическая) эволюция.[12]
К концу XIX века критика естественного отбора достигла такой степени, что в 1903 году немецкий ботаник, Эберхард Деннерт , писал, что «мы стоим у смертного одра дарвинизма», а в 1907 г. Стэндфордский Университет энтомолог Вернон Лайман Келлог, который поддерживал естественный отбор, утверждал, что «... правда в том, что теория дарвиновского отбора, рассматриваемая с учетом ее заявленной способности быть независимым и достаточным механическим объяснением происхождения, сегодня серьезно дискредитирована в биологическом мире». [13] Он добавил, однако, что существуют проблемы, препятствующие широкому распространению любой из альтернатив, поскольку большие мутации кажутся слишком редкими, и нет экспериментальных доказательств механизмов, которые могли бы поддерживать либо ламаркизм, либо ортогенез.[14] Эрнст Майр писал, что обзор литературы по эволюции и учебников биологии показал, что еще в 1930 году вера в то, что естественный отбор является наиболее важным фактором эволюции, была точкой зрения меньшинства, и только несколько популяционных генетиков были строгими селекционерами.[6]
Мотивация для альтернатив

Множество различных факторов побудили людей предложить другие эволюционные механизмы в качестве альтернативы естественному отбору, некоторые из них датируются периодом до Дарвина Происхождение видов. Естественный отбор с его упором на смерть и конкуренцию не нравился некоторым натуралистам, потому что они считали его аморальным и оставляли мало места для телеология или концепция прогресса в развитии жизни.[15][16] Некоторые из этих ученых и философов, например Сент-Джордж Джексон Миварт и Чарльз Лайель, которые пришли к согласию с эволюцией, но не любили естественный отбор, высказали религиозные возражения.[17] Другие, такие как Герберт Спенсер, ботаник Джордж Хенслоу (сын наставника Дарвина Джон Стивенс Хенслоу также ботаник), и Сэмюэл Батлер, чувствовал, что эволюция - это изначально прогрессивный процесс, для объяснения которого одного естественного отбора недостаточно. Третьи, в том числе американские палеонтологи Эдвард Дринкер Коуп и Альфеус Хаятт, имел идеалистическую точку зрения и чувствовал, что природа, включая развитие жизни, следовали упорядоченным образцам этот естественный отбор не мог объяснить.[10]
Еще одним фактором стало появление в конце XIX века новой фракции биологов, типичной представленной генетиками. Хьюго ДеВрис и Томас Хант Морган, который хотел преобразовать биологию в экспериментальную лабораторную науку. Они не доверяли работам натуралистов, таких как Дарвин и Альфред Рассел Уоллес в зависимости от полевых наблюдений за изменчивостью, адаптацией и биогеография, считая это слишком анекдотичным. Вместо этого они сосредоточились на таких темах, как физиология, и генетика что можно легко исследовать с контролируемые эксперименты в лаборатории и не учитывал естественный отбор и степень адаптации организмов к окружающей среде, что было нелегко проверить экспериментально.[18]
Антидарвинистские теории во время затмения
Теистическая эволюция

Британская наука развивалась в начале 19 века на основе естественное богословие который видел приспособление фиксированных видов в качестве доказательства того, что они были специально созданы для целенаправленный божественный замысел. Философские концепции Немецкий идеализм вдохновленные концепции упорядоченного плана гармоничного творчества, которые Ричард Оуэн согласовано с естественным богословием как образец гомология показывая доказательства дизайна. По аналогии, Луи Агассис видел теория перепросмотра как символизирующий образец последовательности творений, в которых человечество было целью божественного плана. В 1844 г. Пережитки адаптировал концепцию Агассиса в теистический эволюционизм. Его анонимный автор Роберт Чемберс предложил «закон» божественно упорядоченного прогрессивного развития с трансмутация видов как расширение теории перепросмотра. Это популяризировало идею, но было решительно осуждено научным истеблишментом. Агассис оставался решительным противником эволюции, и после того, как он переехал в Америку в 1846 году, его идеалистический довод о замысле упорядоченного развития стал очень влиятельным.[19] В 1858 году Оуэн осторожно предположил, что это развитие может быть реальным выражением продолжающегося творческого закона, но дистанцировался от трансмутационистов. Два года спустя в своем обзоре Дарвина О происхождении видов Оуэн напал на Дарвина, в то же время открыто поддерживая эволюцию,[20] выражение веры в образец трансмутации законоподобными средствами. Этот идеалистический аргумент от замысла был подхвачен другими натуралистами, такими как Джордж Джексон Миварт, а Герцог Аргайл которые полностью отвергли естественный отбор в пользу законов развития, которые вели эволюцию по предопределенным путям.[21]
Многие сторонники Дарвина приняли эволюцию на том основании, что она может быть согласована с замыслом. Особенно, Аса Грей считал естественный отбор основным механизмом эволюции и стремился примирить его с естественным богословием. Он предположил, что естественный отбор может быть механизмом, в котором проблема зла страдания приносят большее благо адаптации, но признают, что это вызывает трудности, и предполагают, что Бог может влиять на вариации, на которые действует естественный отбор, чтобы направлять эволюцию.[22] Для Дарвина и Томас Генри Хаксли такое всепроникающее сверхъестественное влияние было за пределами научного исследования, и Джордж Фредерик Райт, рукоположенный служитель, который был коллегой Грея по развитию теистической эволюции, подчеркивал необходимость искать вторичные или известные причины, а не прибегать к сверхъестественным объяснениям: «Если мы перестанем соблюдать это правило, наступит конец всей науке и всей здравой науке».[23]
А светский версия этого методологический натурализм приветствовалось молодым поколением ученых, которые стремились исследовать естественные причины органических изменений и отвергали теистическую эволюцию в науке. К 1872 году дарвинизм в его более широком понимании факта эволюции был принят в качестве отправной точки. Примерно в 1890 году лишь несколько пожилых людей придерживались идеи дизайна в науке, и к 1900 году она полностью исчезла из основных научных дискуссий. Все еще оставалось беспокойство по поводу последствий естественного отбора, и те, кто искал цель или направление в эволюции, обратились к неоламаркизм или же ортогенез как дающие естественные объяснения.[24]
Неоламаркизм

Жан-Батист Ламарк первоначально предложил теорию о трансмутация видов в значительной степени это было основано на постепенном стремлении к большей сложности. Ламарк также верил, как и многие другие в то время, что характеристики, приобретенные в течение жизни организма, могут быть унаследованы следующим поколением, и он видел в этом вторичный эволюционный механизм, который приводит к адаптации к окружающей среде. Обычно такие характеристики включают изменения, вызванные использованием или неиспользованием определенного орган. Именно этот механизм эволюционной адаптации через наследование приобретенных характеристик намного позже стал известен как Ламаркизм.[25] Несмотря на то что Альфред Рассел Уоллес полностью отверг концепцию в пользу естественного отбора, Чарльз Дарвин всегда включал то, что он называл Последствия более частого использования и неиспользования частей, контролируемых естественным отбором в О происхождении видов, приводя такие примеры, как у крупных наземных птиц, у которых ноги становятся сильнее благодаря упражнениям, и у более слабых крыльев, когда они не летают, как страус, они вообще не могли летать.[26]
В конце 19 века термин неоламаркизм стали ассоциироваться с позицией натуралисты которые рассматривали наследование приобретенных характеристик как важнейший эволюционный механизм. Сторонниками этой позиции были британский писатель и критик Дарвина. Сэмюэл Батлер, немецкий биолог Эрнст Геккель, то Американец палеонтологи Эдвард Дринкер Коуп и Альфеус Хаятт, а американская энтомолог Альфей Паккард. Они считали ламаркизм более прогрессивным и, следовательно, философски превосходящим дарвиновскую идею естественного отбора, действующего на случайные вариации. Батлер и Коуп считали, что это позволяет организмам эффективно управлять своей собственной эволюцией, поскольку организмы, развившие новое поведение, изменят модели использования своих органов и, таким образом, запустят эволюционный процесс. Вдобавок Коуп и Геккель считали эволюцию прогрессивным процессом. Идея линейного прогресса была важной частью теории Геккеля. теория перепросмотра эволюции, которая считала эмбриологический развитие организма повторяет его эволюционную историю. Коп и Хаятт искали и думали, что нашли закономерности линейного развития Окаменелости.[27][28] Паккард утверждал, что потеря зрения у слепых пещерных насекомых, которых он изучал, лучше всего объясняется ламарковским процессом атрофии из-за неиспользования в сочетании с наследованием приобретенных характеристик.[29] Паккард также написал книгу о Ламарке и его трудах.[27][30]
Многие американские сторонники неоламаркизма находились под сильным влиянием Луи Агассис и некоторые из них, включая Хаятт и Паккард, были его учениками. Агассис имел идеалистический взгляд на природу, связанный с естественное богословие, что подчеркивало важность порядка и узора. Агассис никогда не принимал эволюцию; его последователи сделали это, но они продолжили его программу поиска упорядоченных закономерностей в природе, которые они считали совместимыми с божественным провидением, и предпочитали эволюционные механизмы, такие как неоламаркизм и ортогенез, которые могли бы их породить.[27][30]
В Британии ботаник Джордж Хенслоу, сын наставника Дарвина Джон Стивенс Хенслоу, был важным сторонником неоламаркизма. Он изучал, как экологический стресс влияет на развитие растений, и писал, что изменения, вызванные такими факторами окружающей среды, могут во многом объяснить эволюцию. Историк науки Питер Дж. Боулер пишет, что, как это было типично для многих ламаркистов 19-го века, Хенслоу, похоже, не понимал необходимости продемонстрировать, что такие вызванные окружающей средой вариации будут унаследованы потомками, которые развивались в отсутствие факторов окружающей среды, которые их породили, но просто предполагал, что они было бы.[31]
Поляризация аргумента: зародышевая плазма Вейсмана
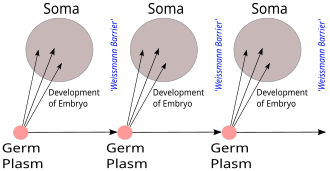
Критики неоламаркизма указывали на то, что никто никогда не приводил убедительных доказательств наследования приобретенных характеристик. Экспериментальная работа немецкого биолога Август Вайсманн привело к зародышевая плазма теория наследования. Это заставило его заявить, что наследование приобретенных характеристик невозможно, поскольку Барьер Вейсмана предотвратит передачу следующих поколений любых изменений, произошедших в организме после рождения. Это эффективно поляризовало спор между дарвинистами и неоламаркианцами, поскольку заставляло людей выбирать, соглашаться или не соглашаться с Вейсманом и, следовательно, с эволюцией путем естественного отбора.[32] Несмотря на критику Вейсмана, неоламаркизм оставался самой популярной альтернативой естественному отбору в конце XIX века и останется позицией некоторых естествоиспытателей даже в XX веке.[28][33]
Эффект Болдуина
Как следствие дискуссии о жизнеспособности неоламаркизма в 1896 г. Джеймс Марк Болдуин, Генри Фэрфилд Осборн и К. Ллойд Морган все независимо друг от друга предложили механизм, при котором новое усвоенное поведение могло вызвать эволюцию новых инстинктов и физических черт посредством естественного отбора, не прибегая к наследованию приобретенных характеристик. Они предположили, что, если особи вида извлекают выгоду из обучения определенному новому поведению, способность изучать это поведение может быть одобрено естественным отбором, а конечным результатом будет развитие новых инстинктов и, в конечном итоге, новых физических адаптаций. Это стало известно как Эффект Болдуина и с тех пор он остается предметом дискуссий и исследований в эволюционной биологии.[34]
Ортогенез

Ортогенез был теорией, согласно которой жизнь имеет врожденную тенденцию к однолинейным изменениям в определенном направлении. Термин был популяризирован Теодор Эймер, немец зоолог в своей книге 1898 г. Об ортогенезе: и о бессилии естественного отбора в формировании видов. Он изучал окраску бабочек и полагал, что обнаружил неадаптивные особенности, которые нельзя объяснить естественным отбором. Эймер также верил в наследование по Ламарку приобретенных характеристик, но он чувствовал, что внутренние законы роста определяют, какие характеристики будут приобретены, и направляют долгосрочное направление эволюции по определенным путям.[35]
У ортогенеза в XIX веке было немало последователей, его сторонниками были российский биолог. Лео С. Берг, а американская палеонтолог Генри Фэрфилд Осборн.[36] Ортогенез был особенно популярен среди некоторых палеонтологов, которые считали, что летопись окаменелостей показывает закономерности постепенных и постоянных однонаправленных изменений. Однако те, кто принимал эту идею, не обязательно соглашались с тем, что механизм, управляющий ортогенезом, был телеологический (целенаправленно). Они действительно считали, что ортогенетические тенденции не адаптивны; на самом деле они чувствовали, что в некоторых случаях они привели к развитию, наносившему вред организму, например, к появлению больших рогов у Ирландский лось что, по их мнению, привело к исчезновению животного.[35]
Поддержка ортогенеза начала снижаться во время современный синтез в 1940-х годах, когда стало очевидно, что ортогенез не может объяснить сложные паттерны ветвления эволюции, выявленные статистическим анализом Окаменелости палеонтологами. Однако некоторые биологи придерживались идеи ортогенеза еще в 1950-х годах, утверждая, что процессы макроэволюция, долговременные тенденции эволюции, были отличны от процессов микроэволюция.[10][11]
Мутационизм

Мутационизм была идея, что новые формы и виды возникают в один шаг в результате больших мутаций. Это рассматривалось как гораздо более быстрая альтернатива дарвиновской концепции постепенного процесса небольших случайных вариаций, на которые воздействует естественный отбор. Он был популярен среди ранних генетиков, таких как Уго де Врис, который вместе с Карл Корренс помог заново открыть Грегор Мендель Наследственные законы России в 1900 году, Уильям Бейтсон британский зоолог, который переключился на генетику и в начале своей карьеры, Томас Хант Морган.[37][38]
Теория эволюции мутаций 1901 года утверждала, что виды переживали периоды быстрых мутаций, возможно, в результате стресса окружающей среды, которые могли вызвать множественные мутации, а в некоторых случаях и совершенно новые виды в одном поколении. Его создателем был голландский ботаник Гуго де Фрис. Де Фрис искал доказательства мутации, достаточно обширной, чтобы произвести новый вид в одном поколении, и подумал, что нашел его в своей работе по разведению вечерней примулы этого рода. Энотера, который он начал в 1886 году. Растения, с которыми работал де Фрис, казалось, постоянно давали новые сорта с поразительными вариациями формы и цвета, некоторые из которых казались новыми видами, потому что растения нового поколения можно было только скрещивать друг с другом. , а не со своими родителями. Сам ДеВриз допускал роль естественного отбора в определении того, какие новые виды выживут, но некоторые генетики, находящиеся под влиянием его работы, в том числе Морган, считали, что естественный отбор вообще не нужен. Идеи де Фриза оказали влияние на первые два десятилетия 20-го века, поскольку некоторые биологи считали, что теория мутаций может объяснить внезапное появление новых форм в летописи окаменелостей; исследования по Энотера распространились по миру. Однако критики, в том числе многие естествоиспытатели, задавались вопросом, почему ни один другой организм не демонстрирует такую быструю мутацию.[39]
Морган был сторонником теории мутации де Вриза и надеялся собрать доказательства в ее пользу, когда начал работать с плодовой мушкой. Drosophila melanogaster в своей лаборатории в 1907 году. Однако исследователь в этой лаборатории, Герман Йозеф Мюллер, которые в 1918 г. определили, что новые сорта, которые де Ври наблюдал во время селекции Энотера были результатом полиплоид гибриды, а не быстрая генетическая мутация.[40][41] Хотя они сомневались в важности естественного отбора, работа генетиков, таких как Морган, Бейтсон, де Фриз и другие, с 1900 по 1915 год установила Менделирующая генетика связан с хромосомная наследственность, который подтвердил критику Августом Вейсманом неоламаркистской эволюции, отказавшись от наследования приобретенных характеристик. Работа в лаборатории Моргана с Дрозофила также подорвали концепцию ортогенеза, продемонстрировав случайный характер мутации.[42]
Конец затмения

В период 1916–1932 гг. Дисциплина популяционная генетика разработаны в основном благодаря работе генетиков Рональд Фишер, J.B.S. Холдейн, и Сьюэлл Райт. В их работе было признано, что подавляющее большинство мутации вызывали небольшие эффекты, которые служили увеличению генетической изменчивости популяции, а не созданию новых видов за один шаг, как предполагали мутационисты. Они смогли создать статистические модели популяционной генетики, которые включали концепцию Дарвина естественный отбор как движущая сила эволюции.[43]
События в генетика убедили естествоиспытателей, таких как Бернхард Ренш и Эрнст Майр отказаться от неоламаркистских идей об эволюции в начале 1930-х годов.[44] К концу 1930-х годов Майр и Феодосий Добжанский синтезировали идеи популяционной генетики со знаниями полевых естествоиспытателей о количестве генетического разнообразия в диких популяциях и важности генетически различных субпопуляций (особенно когда они изолированы друг от друга географическими барьерами) для создания начала 20-го века современный синтез.[45] В 1944 г. Джордж Гейлорд Симпсон интегрированный палеонтология в синтез путем статистического анализа летописи окаменелостей, чтобы показать, что она согласуется с ветвящейся ненаправленной формой эволюции, предсказанной современным синтезом, и, в частности, что линейные тенденции, указанные более ранними палеонтологами в поддержку ламаркизма и ортогенеза, не выдерживают вплоть до тщательного анализа.[46] Майр писал, что к концу синтеза естественный отбор вместе со случайными механизмами, такими как генетический дрейф стали универсальным объяснением эволюционных изменений.[6]
Историография
Концепция затмения предполагает, что дарвиновские исследования были приостановлены, подразумевая, в свою очередь, что предшествующий период активной дарвиновской активности среди биологов был. Однако историки науки, такие как Марк Ларджент, утверждали, что, хотя биологи в целом признали обширные доказательства эволюции, представленные в Происхождение видов, было меньше энтузиазма по поводу естественного отбора как механизма. Вместо этого биологи искали альтернативные объяснения, более соответствующие их мировоззрению, которые включали убеждения, что эволюция должна быть направлена и что она представляет собой форму прогресса. Кроме того, идея периода темного затмения была удобна для таких ученых, как Джулиан Хаксли, которые хотели изобразить современный синтез как новое яркое достижение и, соответственно, изобразить предыдущий период как темный и запутанный. Книга Хаксли 1942 года Эволюция: современный синтез поэтому, утверждал Ларджент, предположил, что так называемый современный синтез начался после длительного периода затмения, продолжавшегося до 1930-х годов, когда менделевцы, неоламаркисты, мутационисты и вейсманисты, не говоря уже о эмбриологах-экспериментаторах и Геккелианский перепросмотрщики вели беговые бои друг с другом.[1] Идея затмения также позволила Хаксли отойти в сторону от неудобной для него ассоциации эволюции с такими аспектами, как социальный дарвинизм, евгеника, империализм, и милитаризм.[1] Аккаунты, такие как Майкл Рус очень большой[1] книга Монада человеку[47] игнорировали, утверждал Ларджент, почти все американские биологи-эволюционисты начала 20 века. Ларджент предложил в качестве альтернативы затмению биологической метафоры, интерфазы дарвинизма, межфазный очевидно спокойный период в цикле деление клеток и рост.[1]
Смотрите также
- Свидетельство окраски для естественного отбора
- История эволюционной мысли
- Возражения против эволюции
- Структурализм
- Витализм
Примечания
- ^ В (Хаксли 1942 С. 22–28). Используется ранее, c. 1925 г., в неопубликованной рукописи автора Дэвид Старр Джордан, чтобы подразумевать, что дарвинизм находится в относительном упадке, а интерес биологов находится в другом месте.[1]
- ^ Это не следует путать с более поздним употреблением термина теистическая эволюция, который относится к теологической вере в совместимость науки и религии.
- ^ Спустя годы открытие радиоактивность предусматривал постоянный источник тепла внутри Земли и возраст в миллиарды лет, что поддерживало позицию Дарвина.[9]
- ^ Дженкин был эрудитом и другом Кельвина.
- ^ От смешанного наследования отказались в пользу Менделирующая генетика в начале 20 века.
- ^ Хаксли был известен как «Бульдог Дарвина».
Рекомендации
- ^ а б c d е ж Ларджент, Марк А. (2009). «Так называемое затмение дарвинизма» (PDF). Произошедший от Дарвина: взгляд на историю эволюционных исследований, 1900–1970 гг.. Американское философское общество.
- ^ (Хаксли 1942, стр. 22–28).
- ^ (Боулер 2003, с. 196, 224).
- ^ (Боулер 1983 )
- ^ (Quammen 2006, стр. 216–223).
- ^ а б c (Mayr & Provine 1998, п. Икс)
- ^ (Quammen 2006, п. 205)
- ^ а б c d е ж грамм (Боулер 1983, стр. 23–26).
- ^ (Боулер 1983, п. 3)
- ^ а б c (Боулер 2003, стр. 196–253).
- ^ а б (Ларсон 2004, стр. 105–129).
- ^ (Боулер 1983, п. 28)
- ^ (Эндерсби 2007, стр. 143,453)
- ^ (Ларсон 2004, п. 128)
- ^ (Боулер 2003, п. 197)
- ^ (Ларсон 2004, стр. 119–120).
- ^ (Quammen 2006, стр. 209–210).
- ^ (Эндерсби 2007, стр. 143–147 182)
- ^ (Боулер 1983, стр. 44–49).
- ^ (Secord 2001, стр. 424, 512).
- ^ (Боулер 1983, стр. 46, 49–50).
- ^ (Боулер 2003, стр. 203–206).
- ^ (Ларсон 2004, стр. 110–111).
- ^ (Боулер 1983, стр. 26–27, 44–45, 54–55).
- ^ (Боулер 2003, стр. 86–95).
- ^ (Дарвин 1872, п.108.)
- ^ а б c (Боулер 2003, стр. 236–244).
- ^ а б (Ларсон 2004, стр. 125–129).
- ^ Паккард, Источник Альфея; Патнэм, Фредерик Уорд (1872 г.). Мамонтова пещера и ее обитатели. Салемское агентство натуралистов. п.12. OCLC 04356215.
- ^ а б (Quammen 2006, стр. 217–219).
- ^ (Боулер 2003, стр. 239–240).
- ^ (Боулер 1983, стр. 41–42).
- ^ (Боулер 2003, стр. 253–255).
- ^ (Боулер 2003, стр. 243, 367).
- ^ а б (Quammen 2006, п. 221)
- ^ (Боулер 2003, п. 249)
- ^ (Боулер 2003, стр. 265–270).
- ^ (Ларсон 2004 С. 127–129, 157–167).
- ^ (Эндерсби 2007, стр. 148–162).
- ^ (Эндерсби 2007, стр. 202–205).
- ^ Рэмси, Джастин; Рэмси, Тара С. (август 2014 г.). «Экологические исследования полиплоидии через 100 лет после ее открытия». Фил. Пер. R. Soc. Лондон. B. 5 (369): 20130352. Дои:10.1098 / rstb.2013.0352. ЧВК 4071525. PMID 24958925.
- ^ (Боулер 2003, стр. 269–272).
- ^ (Mayr & Provine 1998, стр. xi – xii)
- ^ (Mayr & Provine 1998 С. 124–127, 296).
- ^ (Mayr & Provine 1998, стр. xii – xiii)
- ^ (Боулер 2003, п. 337)
- ^ (Русе 1996 )
Источники
- Боулер, Питер Дж. (2003). Эволюция: история идеи. Калифорнийский университет Press. ISBN 0-520-23693-9.CS1 maint: ref = harv (связь)
- Боулер, Питер Дж. (1983). Затмение дарвинизма: антидарвиновские эволюционные теории в период около 1900 года. Издательство Университета Джона Хопкинса. ISBN 978-0-8018-4391-4.CS1 maint: ref = harv (связь)
- Дарвин, Чарльз (1872). Происхождение видов посредством естественного отбора или сохранение избранных рас в борьбе за жизнь (6-е изд.). Лондон: Джон Мюррей. ISBN 1-904633-78-1.CS1 maint: ref = harv (связь)
- Эндерсби, Джим (2007). История биологии морской свинки. Издательство Гарвардского университета. ISBN 978-0-674-02713-8.CS1 maint: ref = harv (связь)
- Хаксли, Джулиан (1942). Эволюция: современный синтез. Лондон: Аллен и Анвин.CS1 maint: ref = harv (связь)
- Ларсон, Эдвард Дж. (2004). Эволюция: замечательная история научной теории. Современная библиотека. ISBN 0-679-64288-9.CS1 maint: ref = harv (связь)
- Майр, Эрнст; Provine, W. B., eds. (1998). Эволюционный синтез: перспективы объединения биологии. Издательство Гарвардского университета. ISBN 0-674-27225-0.CS1 maint: ref = harv (связь)
- Кваммен, Дэвид (2006). Неохотный мистер Дарвин. Книги Атласа. ISBN 0-393-05981-2.CS1 maint: ref = harv (связь)
- Русе, Майкл (1996). От монады к человеку: концепция прогресса в эволюционной биологии. Издательство Гарвардского университета. ISBN 978-0-674-03248-4.CS1 maint: ref = harv (связь)
- Секорд, Джеймс А. (2001). Викторианская сенсация: необычайная публикация, получение и тайное авторство пережитков естественной истории творения. Издательство Чикагского университета. ISBN 978-0-226-74410-0.CS1 maint: ref = harv (связь)