Современный синтез (20 век) - Википедия - Modern synthesis (20th century)
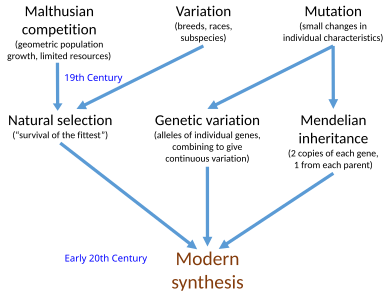

В современный синтез[а] был синтез в начале 20-го века, согласовывающий Чарльз Дарвин теория эволюция и Грегор Мендель Идеи о наследственности в общих математических рамках. Джулиан Хаксли ввел термин в своей книге 1942 года, Эволюция: современный синтез.
Идеи XIX века естественный отбор и Менделирующая генетика были объединены с популяционная генетика, в начале ХХ века. Современный синтез также обратился к взаимосвязи между широкомасштабными изменениями макроэволюция виденный палеонтологи и мелкомасштабные микроэволюция местных население живых организмов. Синтез по-разному определяли его основатели: Эрнст Майр в 1959 г., Г. Ледьярд Стеббинс в 1966 г. и Феодосий Добжанский в 1974 г. предлагает различное количество основных постулатов, хотя все они включают естественный отбор, работающий над наследственными вариациями, вызванными мутациями. Другие важные фигуры в синтезе включали Э. Б. Форд, Бернхард Ренш, Иван Шмальгаузен, и Джордж Гейлорд Симпсон. Ранним событием в современном синтезе было Р. А. Фишер статья 1918 года по математической популяционной генетике, но Уильям Бейтсон, и отдельно Удный Йоль, уже начали показывать, как менделевская генетика могла работать в эволюции в 1902 году.
Последовали различные синтезы, сопровождавшие постепенный распад синтеза начала 20 века, в том числе с социальное поведение в Э. О. Уилсон с социобиология в 1975 г. эволюционная биология развития интеграция эмбриология с генетикой и эволюцией, начиная с 1977 года, и Массимо Пильуччи предлагается расширенный эволюционный синтез 2007 г. По мнению биолога-эволюциониста Евгений Кунин в 2009 году современный синтез будет заменен «постмодернистским» синтезом, который будет включать революционные изменения в молекулярная биология, изучение прокариоты и в результате Дерево жизни, и геномика.[3]
Разработки, приведшие к синтезу

Эволюция Дарвина путем естественного отбора, 1859 г.
Чарльз Дарвин книга 1859 г. О происхождении видов удалось убедить большинство биологов, что эволюция произошло, но не смог убедить их, что естественный отбор был его основным механизмом. В XIX - начале XX веков вариации Ламаркизм (наследование приобретенных характеристик), ортогенез (прогрессивная эволюция), сальтационизм (эволюция скачками) и мутационизм (эволюция, обусловленная мутациями) обсуждались как альтернативы.[4] Альфред Рассел Уоллес выступал за селекционистскую версию эволюции и, в отличие от Дарвина, полностью отвергал ламаркизм.[5] В 1880 году точка зрения Уоллеса была названа неодарвинизм пользователя Samuel Butler.[6][7]

Затмение дарвинизма, 1880-е годы и позже
Начиная с 1880-х годов, среди биологов было широко распространено мнение, что дарвиновская эволюция находится в тяжелом положении. Этот затмение дарвинизма (в Джулиан Хаксли фраза) выросла из слабых мест в описании Дарвина, написанном с неправильным взглядом на наследование. Сам Дарвин верил в смешанное наследование, что означало, что любая новая вариация, даже если она полезна, будет ослабляться на 50% в каждом поколении, поскольку инженер Флиминг Дженкин правильно отмечено в 1868 г.[8][9] Это, в свою очередь, означало, что небольшие вариации не сохранятся достаточно долго, чтобы их можно было выбрать. Таким образом, смешивание будет прямо противодействовать естественному отбору. Вдобавок Дарвин и другие считали вполне возможным ламарковское наследование приобретенных характеристик, а теория Дарвина 1868 г. пангенезис, с вкладом в следующее поколение (геммулы), исходящим от всех частей тела, фактически подразумевает ламаркизм, а также смешение.[10][11][12]
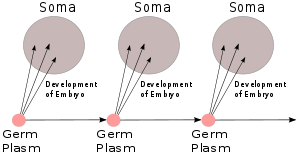
Зародышевая плазма Вейсмана, 1892 г.
Август Вейсманн идея, изложенная в его книге 1892 г. Das Keimplasma: eine Theorie der Vererbung (Зародышевая плазма: теория наследования),[13] был тот наследственный материал, который он назвал зародышевая плазма, а остальная часть тела ( сома ) имела одностороннюю связь: зародышевая плазма формировала тело, но тело не влияло на зародышевую плазму, за исключением косвенного участия в популяции, подверженной естественному отбору. Если это так, то это делает пангенезис Дарвина ошибочным, а наследование по Ламарку невозможным. Его эксперимент на мышах, отрезавший им хвосты и показавший, что у их потомков нормальные хвосты, продемонстрировал, что наследование было «трудным».[b] Он аргументированно и категорично утверждал[15] за дарвинизм и против ламаркизма, поляризующие мнения среди других ученых. Это усилившееся анти-дарвиновское чувство, способствовавшее его затмению.[16][17]
Спорные начала
Генетика, мутационизм и биометрия, 1900–1918 гг.

Проводя селекционные эксперименты по выяснению механизма наследования в 1900 г., Уго де Врис и Карл Корренс независимо открыты заново Грегор Мендель работа. Новости об этом достигли Уильям Бейтсон в Англия, который доложил о статье во время презентации Королевское садоводческое общество в мае 1900 г.[18] В Менделирующее наследование, вклады каждого родителя сохраняют свою целостность, а не смешиваются с вкладом другого родителя. В случае скрещивания двух чистопородных сортов, таких как круглый и морщинистый горох Менделя, все потомки первого поколения одинаковы, а в данном случае - со всех сторон. Позволяя им скрещиваться, исходные характеристики снова проявляются (сегрегация): около 3/4 их потомков круглые, 1/4 морщинистые. Между появлением потомства наблюдается разрыв; де Фрис ввел термин аллель для варианта формы унаследованная характеристика.[19] Это усилило серьезное разделение мысли, существовавшее уже в 1890-х годах, между градуалистами, последовавшими за Дарвином, и сальтационистами, такими как Бейтсон.[20]
Этими двумя школами были менделевцы, такие как Бейтсон и де Вриз, которые поддерживали мутационизм, эволюцию, управляемую мутациями, основанную на генах, аллели которых раздельно сегрегированный как горох Менделя;[21][22] и биометрическая школа во главе с Карл Пирсон и Уолтер Велдон. Биометристы решительно выступили против мутационизма, заявив, что эмпирические данные указывают на то, что вариации у большинства организмов непрерывны, а не дискретны, как, казалось, предсказывал менделизм; они ошибочно полагали, что менделизм неизбежно подразумевает эволюцию прерывистыми скачками.[23][24]

Традиционная точка зрения состоит в том, что биометристы и менделевцы отвергли естественный отбор и 20 лет отстаивали свои отдельные теории, и этот спор разрешился только с развитием популяционной генетики.[23][25]Более поздняя точка зрения состоит в том, что Бейтсон, де Фрис, Томас Хант Морган и Реджинальд Паннетт к 1918 г. сформировал синтез менделизма и мутационизма. Понимание, достигнутое этими генетиками, касалось действия естественного отбора на аллели (альтернативные формы гена), Равновесие Харди – Вайнберга, эволюция постоянно изменяющихся признаков (например, роста) и вероятность того, что новая мутация станет фиксированной. С этой точки зрения первые генетики приняли естественный отбор, но отвергли неменделирующие идеи Дарвина об изменчивости и наследственности, и синтез начался вскоре после 1900 года.[26][27] Традиционное утверждение, что менделевцы отвергают идею непрерывного изменения, ложно; Еще в 1902 году Бейтсон и Сондерс писали, что «если бы было хотя бы так мало, как, скажем, четыре или пять пар возможных аллеломорфов, различные гомо- и гетерозиготные комбинации могли бы при последовательной основе дать такой близкий подход к непрерывной кривой. , что чистота элементов будет неожиданной ".[28] Также в 1902 году статистик Удный Йоль математически показал, что при наличии множества факторов теория Менделя допускает непрерывные вариации. Юл критиковал подход Бейтсона как конфронтационный,[29] но не смог предотвратить ссору между менделевцами и биометристами.[30]
Крысы Касла в капюшонах, 1911 год.
Начиная с 1906 г., Уильям Кастл провели длительное исследование влияния отбора на окрас шерсти в крысы. В пегий или узор с капюшоном был рецессивный к серому дикому типу. Он скрестил крыс с капюшоном как с дикими, так и с «ирландскими» типами, а затем скрестил потомство с чистыми крысами с капюшоном. Темная полоса на спине была больше. Затем он попытался выбрать разные группы для больших или меньших полос для 5 поколений и обнаружил, что можно изменить характеристики, выходящие за пределы начального диапазона вариаций. Это фактически опровергло утверждение де Фриза о том, что непрерывные изменения были вызваны окружающей средой и не могут быть унаследованы. К 1911 году Касл заметил, что результаты могут быть объяснены дарвиновским отбором наследственной вариации достаточного числа менделевских генов.[31][32][33]
Дрозофилы Моргана, 1912 год.
Томас Хант Морган начал свою карьеру в области генетики как сальтационист и начал с попытки продемонстрировать, что мутации могут приводить к появлению новых видов у плодовых мух. Однако экспериментальная работа в его лаборатории с плодовой мушкой, Drosophila melanogaster[c] продемонстрировали, что вместо того, чтобы создавать новые виды за один шаг, мутации увеличивают количество генетических вариаций в популяции.[34] К 1912 году, после многих лет работы над генетикой плодовых мушек, Морган показал, что у этих насекомых есть много небольших менделевских факторов (обнаруженных как мухи-мутанты), над которыми могла работать дарвиновская эволюция, как если бы вариация была полностью непрерывной. Для генетиков был открыт путь к выводу, что менделизм поддерживает дарвинизм. [35]
Препятствие: позитивизм Вудгера, 1929 г.
В биолог-теоретик и философ биологии Джозеф Генри Вудгер привел к введению позитивизм в биологию с его книгой 1929 года Биологические принципы. Он увидел зрелую наука как характеризующиеся рамками гипотезы что может быть подтверждено фактами, установленными эксперименты. Он критиковал традиционные естественная история стиль биология, включая изучение эволюция, как незрелая наука, поскольку она опиралась на повествование.[36] Вудгер решил сыграть для биологии роль Роберт Бойл 1661 год Скептический химик, намереваясь превратить предмет в формальную единую науку, и, в конечном итоге, следуя Венский круг логических позитивистов вроде Отто Нейрат и Рудольф Карнап, уменьшить от биологии к физике и химии. Его усилия стимулировали биолога Дж. Б. С. Холдейн продвигать аксиоматизацию биологии и, оказывая влияние на таких мыслителей, как Хаксли, помог осуществить современный синтез.[36] Позитивистский климат сделал естественную историю немодной, и в Америке исследования и преподавание эволюции на университетском уровне к концу 1930-х годов практически прекратились. Гарвардский физиолог Уильям Джон Крозье сказал своим ученикам, что эволюция - это даже не наука: «Вы не можете экспериментировать с двумя миллионами лет!»[37]
Мнения изменились с принятием математическое моделирование и контролируемое экспериментирование в популяционной генетике, сочетая генетику, экологию и эволюцию в рамках, приемлемых для позитивизма.[38]
События в синтезе
Математическая популяционная генетика Фишера и Холдейна, 1918–1930 гг.
В 1918 г. Р. А. Фишер написал статью "Корреляция между родственниками на основе предположения о менделевской наследственности,"[39] который математически показал, как непрерывное изменение может быть результатом ряда дискретных генетические локусы. В этой и последующих статьях, кульминацией которых стала его книга 1930 г. Генетическая теория естественного отбора,[40] Фишер показал, насколько менделевская генетика согласуется с идеей эволюции, движимой естественным отбором.[41][d]
В течение 1920-х годов серия документов к Дж. Б. С. Холдейн прикладного математического анализа к реальным примерам естественного отбора, таким как эволюция промышленного меланизма у берёзовых бабочек.[41] Холдейн установил, что естественный отбор может работать даже быстрее, чем предполагал Фишер.[43] И рабочие, и другие, такие как Добжанский и Райт, явно намеревались привести биологию в соответствие с философскими стандартами физических наук, сделав ее прочно основанной на математическом моделировании, а ее прогнозы подтверждались экспериментом. Естественный отбор, когда-то считавшийся безнадежно неподдающимся проверке спекуляцией об истории, стал предсказуемым, измеримым и проверяемым.[44]
Эмбриология Де Бир, 1930
Традиционно считается, что биология развития играл небольшую роль в современном синтезе,[45] но в его книге 1930 года Эмбрионы и предки, эволюционный эмбриолог Гэвин де Бир ожидаемый эволюционная биология развития[46] показывая, что эволюция может происходить гетерохрония,[47] например, в сохранение юношеских черт у взрослого.[48] Это, как утверждал де Бир, может вызвать внезапные изменения в Окаменелости, так как зародыши плохо окаменевают. Поскольку пробелы в летописи окаменелостей использовались в качестве аргумента против постепенной эволюции Дарвина, объяснение де Бера поддерживало дарвиновскую позицию.[49]Однако, несмотря на де Бера, современный синтез в значительной степени игнорировал эмбриональное развитие для объяснения формы организмов, поскольку популяционная генетика оказалась адекватным объяснением того, как формы эволюционировали.[50][51][e]
Адаптивный пейзаж Райта, 1932 г.

Популяционный генетик Сьюэлл Райт сосредоточены на комбинациях генов, которые взаимодействуют как комплексы, и на эффектах инбридинг на небольших относительно изолированных популяциях, которые могут подвергаться генетический дрейф. В статье 1932 года он представил концепцию адаптивный ландшафт в которых такие явления, как скрещивание и генетический дрейф в небольших популяциях, могут оттолкнуть их от адаптивных пиков, что, в свою очередь, позволит естественному отбору подтолкнуть их к новым адаптивным пикам.[41][53] Модель Райта понравилась бы естествоиспытателям, таким как Феодосиус Добжанский и Эрнст Майр, которые осознавали важность географической изоляции в популяциях реального мира.[43] Работа Фишера, Холдейна и Райта помогла основать дисциплину теоретической популяционной генетики.[54][55][56]
Эволюционная генетика Добжанского, 1937 г.

Феодосий Добжанский, иммигрант из Советский союз к Соединенные Штаты, который был докторантом в лаборатории Моргана по плодовым мухам, был одним из первых, кто применил генетику к естественным популяциям. Он работал в основном с Drosophila pseudoobscura. Он многозначительно говорит: «Климат России варьируется от арктического до субтропического ... Только лабораторные работники, которые не обладают и не хотят знать никаких знаний о живых существах в природе, были и остаются в меньшинстве».[57] Неудивительно, что были и другие русский генетики с похожими идеями, хотя какое-то время их работы были известны лишь немногим в Запад. Его работа 1937 года Генетика и происхождение видов[58] был ключевым шагом в преодолении разрыва между популяционными генетиками и естествоиспытателями. Он представил выводы, к которым пришли Фишер, Холдейн и особенно Райт в их математических статьях, в форме, легко доступной другим.[41][43] Далее Добжанский утверждал, что в основе эволюции лежат материальные гены, расположенные в виде цепочки на физических наследственных структурах, т.е. хромосомы, и связаны более или менее сильно друг к другу в зависимости от их физического расстояния друг от друга на хромосомах. Как и в случае с Холдейном и Фишером, «эволюционная генетика» Добжанского[59] была настоящей наукой, объединяющей теперь клеточную биологию, генетику, а также микро- и макроэволюцию.[44] В его работе подчеркивалось, что популяции в реальном мире обладают гораздо большей генетической изменчивостью, чем предполагали первые популяционные генетики в своих моделях, и что генетически различные субпопуляции имеют важное значение. Добжанский утверждал, что естественный отбор помогает поддерживать генетическое разнообразие, а также способствует изменениям. На него повлияло то, что в 1920-х годах он познакомился с работами Сергей Четвериков, который изучал роль рецессивных генов в поддержании резервуара генетической изменчивости в популяции до того, как его работа была закрыта из-за роста Лысенковщина в Советский союз.[41][43] К 1937 году Добжанский смог доказать, что мутации были основным источником эволюционных изменений и изменчивости, наряду с хромосомными перестройками, влиянием генов на своих соседей во время развития и полиплоидией. Затем генетический дрейф (он использовал этот термин в 1941 году), отбор, миграция и географическая изоляция могут изменить частоты генов. В-третьих, такие механизмы, как экологическая или сексуальная изоляция и гибридное бесплодие, могут исправить результаты более ранних процессов.[60]
Экологическая генетика Форда, 1940 г.

Э. Б. Форд был экспериментальным натуралистом, который хотел проверить естественный отбор в природе, фактически изобретая область экологическая генетика.[61] Его работа по естественному отбору в диких популяциях бабочек и мотыльков была первой, показавшей, что предсказания Р. А. Фишера верны. В 1940 году он первым описал и дал определение генетический полиморфизм, и предсказать, что полиморфизмы групп крови человека может поддерживаться в популяции, обеспечивая некоторую защиту от болезней.[61][62] Его книга 1949 года Менделизм и эволюция[63] помогли убедить Добжанского изменить акцент в третьем издании его знаменитого учебника. Генетика и происхождение видов от дрейфа к выбору.[64]
Стабилизирующий отбор Шмальгаузена, 1941 г.
Иван Шмальгаузен разработал теорию стабилизирующий отбор, идея о том, что отбор может сохранить определенную ценность, опубликовав статью на русском языке под названием «Стабилизирующий отбор и его место среди факторов эволюции» в 1941 г. и монографию Факторы эволюции: теория стабилизирующего отбора[65] в 1945 году. Он развил это из концепции Дж. М. Болдуина 1902 года, согласно которой изменения, вызванные окружающей средой, в конечном итоге будут заменены наследственными изменениями (включая Эффект Болдуина о поведении), следуя импликациям этой теории к их дарвиновским выводам, и приводя его в противоречие с лысенкоизмом. Шмальгаузен заметил, что стабилизирующий отбор удалит большинство отклонений от нормы, причем большинство мутаций будут вредными.[66][67][68] Добжанский назвал работу «важным недостающим звеном в современном взгляде на эволюцию».[69]
Популяризирующий синтез Хаксли, 1942 г.

В 1942 г. Джулиан Хаксли серьезно, но популяризирующий[70][71] Эволюция: современный синтез[2] ввел название для синтеза и намеренно поставил перед собой цель продвигать «синтетическую точку зрения» на эволюционный процесс. Он представлял себе широкий синтез многих наук: генетики, физиологии развития, экологии, систематики, палеонтологии, цитологии и математического анализа биологии, и предположил, что эволюция будет происходить по-разному в разных группах организмов в зависимости от того, как организован их генетический материал и их стратегии воспроизводства, ведущие к прогрессивным, но меняющимся тенденциям эволюции.[71] Его видение было «эволюционным гуманизмом»,[72] с системой этики и значимым местом для «Человека» в мире, основанным на единой теории эволюции, которая продемонстрирует прогресс, ведущий к человеку на его вершине. По его мнению, естественный отбор был «фактом природы, который можно проверить наблюдением и экспериментом», тогда как «период синтеза» 1920-х и 1930-х годов сформировал «более единую науку»,[72] соперничая с физикой и способствуя «возрождению дарвинизма».[72]
Однако книга не была исследовательским текстом, как казалось. По мнению философа науки Майкл Рус, и, по собственному мнению Хаксли, Хаксли был «универсалом, синтезатором идей, а не специалистом».[70] Руз отмечает, что Хаксли писал так, как будто он добавлял эмпирические доказательства к математической структуре, установленной Фишером и популяционными генетиками, но это не так. Хаксли избегал математики, например, даже не упоминая Фундаментальная теорема естественного отбора Фишера. Вместо этого Хаксли использовал массу примеров, чтобы продемонстрировать мощь естественного отбора и его влияние на менделевские гены. Книга успешно преследовала цель убедить читателей в реальности эволюции, эффективно иллюстрируя такие темы, как островная биогеография, видообразование, и конкуренция. Хаксли далее показал, что появление долгосрочных ортогенетические тенденции - предсказуемые направления эволюции - в летописи окаменелостей легко объяснить как аллометрический рост (так как части связаны между собой). Тем не менее, Хаксли не отвергал ортогенез сразу, а всю жизнь верил в прогресс. Homo sapiens в качестве конечной точки, и с 1912 года он находился под влиянием виталист философ Анри Бергсон, хотя публично он придерживался атеистической позиции в отношении эволюции.[70] Вера Хаксли в прогресс в эволюции и эволюционный гуманизм в различных формах разделялась Добжанским, Майром, Симпсоном и Стеббинсом, и все они писали о «будущем человечества». И Хаксли, и Добжанский восхищались священником-палеонтологом. Пьер Тейяр де Шарден, Хаксли пишет введение к книге Тейяра 1955 года по ортогенезу, Феномен человека. Это видение требовало, чтобы эволюция рассматривалась как центральный и руководящий принцип биологии.[72]
Аллопатрическое видообразование Майра, 1942 г.

Эрнст Майр ключевой вклад в синтез был Систематика и происхождение видов, опубликовано в 1942 году.[73] В нем утверждается важность и цель объяснения популяционной изменчивости в эволюционных процессах, включая видообразование. Он проанализировал, в частности, влияние разнотипный виды, географическая изменчивость и изоляция географическими и другими способами.[74] Майр подчеркнул важность аллопатрическое видообразование, где географически изолированные подгруппы населения расходятся настолько, что репродуктивная изоляция происходит. Он скептически относился к реальности симпатрическое видообразование полагая, что географическая изоляция является предпосылкой для создания внутренних (репродуктивных) изолирующих механизмов. Майр также представил концепция биологических видов это определяло вид как группу скрещивающихся или потенциально скрещивающихся популяций, репродуктивно изолированных от всех других популяций.[41][43][75][76] Прежде чем он ушел Германия для Соединенных Штатов в 1930 году Майр находился под влиянием работ немецкого биолога Бернхард Ренш, который в 1920-х годах проанализировал географическое распространение разнотипных видов, уделяя особое внимание тому, как различия между популяциями коррелируют с такими факторами, как различия в климате.[77][78][79]

Палеонтология Симпсона, 1944 г.
Джордж Гейлорд Симпсон в своей книге 1944 г. продемонстрировал совместимость современного синтеза с палеонтологией. Темп и мода в эволюции. Работа Симпсона имела решающее значение, потому что многие палеонтологи не соглашались, в некоторых случаях категорически, с идеей, что естественный отбор был основным механизмом эволюции. Он показал, что тенденции линейной прогрессии (например, в эволюция лошади ), которые ранее палеонтологи использовали в качестве подтверждения неоламаркизм и ортогенез не выдержал тщательного изучения. Вместо этого Окаменелости соответствовал нерегулярному, ветвящемуся и ненаправленному образцу, предсказанному современным синтезом.[41][43]
Общество изучения эволюции, 1946 г.
Вовремя война Майр редактировал серию бюллетеней Комитета по общим проблемам генетики, палеонтологии и систематики, созданного в 1943 году, в которых сообщалось об обсуждениях «синтетической атаки» на междисциплинарные проблемы эволюции. В 1946 году комитет стал Обществом изучения эволюции, и его первыми подписали Майр, Добжанский и Сьюэлл Райт. Майр стал редактором своего журнала, Эволюция. С точки зрения Майра и Добжанского, как предполагает историк науки Бетти Смоковитис, дарвинизм возродился, эволюционная биология была узаконена, а генетика и эволюция были синтезированы в новую единую науку. В новые рамки уложилось все, кроме «еретиков» вроде Ричард Гольдшмидт который раздражал Майра и Добжанского, настаивая на возможности видообразование путем макромутации, создавая «обнадеживающих монстров». Результатом стали «ожесточенные споры».[52]

Ботаника Стеббинса, 1950 г.
Ботаник Г. Ледьярд Стеббинс расширил синтез, чтобы охватить ботаника. Он описал важное влияние на видообразование из гибридизация и полиплоидия в растениях в его книге 1950 года Вариации и эволюция растений. Это позволяло эволюции происходить быстро, в частности, полиплоидии, очевидно, способной мгновенно эффективно создавать новые виды.[41][80]
Определения учредителей
Современный синтез был по-разному определен его разными основоположниками, с разным количеством основных постулатов, как показано в таблице.
| Компонент | Mayr 1959 | Стеббинс, 1966 | Добжанский, 1974 |
|---|---|---|---|
| Мутация | (1) Случайность во всех случаях, которые приводят к появлению новых генотипов, например мутация [81] | (1) источник изменчивости, но не направление[82] | (1) дает генетическое сырье[83] |
| Рекомбинация | (1) Случайность в рекомбинации, оплодотворение[81] | (2) источник изменчивости, но не направление[82] | |
| Хромосомная организация | (3) влияет на генетическое сцепление, организует вариации в Генофонд[82] | ||
| Естественный отбор | (2) является только определяющим фактором,[81][84] как показано на приспособления к физической и биотической среде[81] | (4) направляет изменения в Генофонд[82] | (2) конструирует эволюционные изменения из генетического сырья[83] |
| Репродуктивная изоляция | (5) ограничивает направление, в котором отбор может направлять популяцию[82] | (3) делает расхождение необратимым в половые организмы[83] |
После синтеза
После синтеза эволюционная биология продолжала развиваться при большом вкладе ученых, включая У. Д. Гамильтона,[85] Джордж К. Уильямс,[86] Э. О. Уилсон,[87] Эдвард Б. Льюис[88] и другие.
Инклюзивный фитнес Гамильтона, 1964 год.
В 1964 г. В. Д. Гамильтон опубликовал две статьи на тему «Генетическая эволюция социального поведения». Эти определенные инклюзивный фитнес как количество потомков, эквивалентных тому, что человек поднимает, спасает или иным образом поддерживает своим поведением. Это контрастировало с личным репродуктивным здоровьем, количеством потомства, которое непосредственно рождает человек. Гамильтон и другие, такие как Джон Мейнард Смит, утверждали, что успех гена заключается в максимальном увеличении количества копий самого себя, либо путем их зачатия, либо косвенного поощрения зачатия родственными индивидами, которые разделяют ген, теория родственный отбор.[85][89]
Геноцентрированная эволюция Уильямса, 1966 г.
В 1966 г. Джордж К. Уильямс опубликовано Адаптация и естественный отбор, обозначил геноцентрический взгляд на эволюцию следуя концепции Гамильтона, оспаривая идею эволюционный прогресс, и нападая на распространенную тогда теорию групповой выбор. Уильямс утверждал, что естественный отбор работает путем изменения частоты аллелей и не может работать на уровне групп.[90][86] Геноцентрированная эволюция была популяризирована Ричард Докинз в его книге 1976 года Эгоистичный ген и развивался в его более технических трудах.[91][92]
Социобиология Вильсона, 1975 г.

В 1975 г. Э. О. Уилсон опубликовал свой скандальный[93] книга Социобиология: новый синтез, подзаголовок, отсылающий к современному синтезу[87] поскольку он попытался включить изучение общества животных в эволюционную ленту. Это казалось совершенно новым, хотя Уилсон следовал за Дарвином, Фишером, Докинзом и другими.[87] Такие критики, как Герхард Ленски отметил, что он следовал подходу Хаксли, Симпсона и Добжанского, который Ленски считал излишне редуцирующим с точки зрения человеческого общества.[94] К 2000 г. предлагаемая дисциплина социобиология превратилась в относительно хорошо принятую дисциплину эволюционная психология.[87]
Гомеотические гены Льюиса, 1978

В 1977 г. рекомбинантная ДНК Технологии позволили биологам начать изучение генетического контроля развития. Рост эволюционная биология развития с 1978 года, когда Эдвард Б. Льюис обнаруженный гомеотический гены, показали, что многие так называемые инструментарий генов действуют, чтобы регулировать развитие, влияя на экспрессию других генов. Также выяснилось, что некоторые из регуляторных генов чрезвычайно древние, поэтому такие разные животные, как насекомые и млекопитающие, имеют общие механизмы контроля; например, Pax6 ген участвует в формировании глаза мышей и плодовых мух. Такой глубокая гомология предоставили убедительные доказательства эволюции и указали пути, по которым пошла эволюция.[88]
Поздние синтезы
В 1982 г. была опубликована историческая справка о серии книг по эволюционной биологии.[f] мог бы заявить без оговорок, что эволюция является центральным организующим принципом биологии. Смоковитис прокомментировал это, что «то, над чем работали архитекторы синтеза, стало к 1982 году фактически», добавив в сноске, что «центральная роль эволюции была таким образом передана. неявное знание, часть получил мудрость профессии ».[95]
Однако к концу 20 века современный синтез показал свой возраст, и свежие синтезы для исправления его дефектов и заполнения пробелов предлагались с разных сторон. К ним относятся такие разнообразные области, как изучение общества,[87] биология развития,[50] эпигенетика[96] молекулярная биология, микробиология, геномика,[3] симбиогенез, и горизонтальный перенос генов.[97] Физиолог Денис Ноубл утверждает, что эти дополнения делают неодарвинизм в смысле современного синтеза начала 20-го века «по крайней мере неполным как теорию эволюции»,[97] и тот, который был сфальсифицирован более поздними биологическими исследованиями.[97]
Майкл Роуз и Тодд Окли отмечают, что эволюционная биология, ранее разделенная на "Балканизированный ", был объединен геномикой. По их мнению, он отверг по крайней мере пять общих допущений современного синтеза, а именно, что геном всегда представляет собой хорошо организованный набор генов; что каждый ген выполняет одну функцию; что виды являются биохимически хорошо адаптированы к своим экологическим нишам; что виды являются прочными единицами эволюции, и для него характерны все уровни от организма до органа, клетки и молекулы внутри вида; и что конструкция каждого организма и клетки является эффективной. Они утверждают, что «новая биология» объединяет геномику, биоинформатика, и эволюционная генетика в универсальный инструментарий для «Постмодернистского синтеза».[54]
Расширенный эволюционный синтез Пильуччи, 2007 г.
В 2007 году, спустя более полувека после современного синтеза, Массимо Пильуччи призвал к расширенный эволюционный синтез включить аспекты биологии, которые не были включены или не существовали в середине 20 века.[98][99] Он пересматривает относительную важность различных факторов, ставит под сомнение допущения, сделанные в современном синтезе, и добавляет новые факторы.[99][100] Такие как многоуровневый отбор, трансгенерационное эпигенетическое наследование, конструкция ниши, и эволюционируемость.[101][96][102]
«Постмодернистский» эволюционный синтез Кунина, 2009 г.
В 2009 году к 200-летию Дарвина Происхождение видов' 150-я и 200-я «раннего эволюционного синтеза» Ламарка,[3] Philosophie Zoologique, эволюционный биолог Евгений Кунин заявил, что, хотя «здание [начала 20 века] современного синтеза рухнуло, по-видимому, не подлежит ремонту»,[3] можно было увидеть новый синтез 21 века. Он утверждал, что в эволюционной биологии произошли три взаимосвязанные революции: молекулярная, микробиологическая и геномная. В молекулярная революция включала нейтральную теорию о том, что большинство мутаций нейтральны и что отрицательный выбор случается чаще, чем положительный формы, и что вся текущая жизнь возникла из один общий предок. В микробиологии синтез расширился, чтобы охватить прокариоты, с помощью рибосомная РНК сформировать Дерево жизни. Ну наконец то, геномика объединил молекулярный и микробиологический синтезы - в частности, горизонтальный перенос генов между бактерии показывает, что прокариоты могут свободно обмениваться генами. Многие из этих замечаний уже были высказаны другими исследователями, такими как Ульрих Кучера и Карл Дж. Никлас.[103]
К замене синтеза
Биологи вместе с учеными, занимающимися историей и философией биологии, продолжали обсуждать необходимость и возможную природу замещающего синтеза. Например, в 2017 году Филипп Хьюнеман и Дени М. Уолш заявили в своей книге Бросая вызов современному синтезу that numerous theorists had pointed out that the disciplines of embryological developmental theory, morphology, and ecology had been omitted. They noted that all such arguments amounted to a continuing desire to replace the modern synthesis with one that united "all biological fields of research related to evolution, adaptation, and diversity in a single theoretical framework."[104] They observed further that there are two groups of challenges to the way the modern synthesis viewed inheritance. The first is that other modes such as epigenetic inheritance, фенотипическая пластичность, то Эффект Болдуина, а материнский эффект allow new characteristics to arise and be passed on and for the genes to catch up with the new adaptations later. The second is that all such mechanisms are part, not of an inheritance system, but a developmental system: the fundamental unit is not a discrete selfishly competing gene, but a collaborating system that works at all levels from genes and cells to organisms and cultures to guide evolution.[105]
Историография
Looking back at the conflicting accounts of the modern synthesis, the historian Betty Smocovitis notes in her 1996 book Объединяющая биология: эволюционный синтез и эволюционная биология that both historians and philosophers of biology have attempted to grasp its scientific meaning, but have found it "a moving target";[106] the only thing they agreed on was that it was a historical event.[106] In her words
"by the late 1980s the notoriety of the evolutionary synthesis was recognized ... So notorious did 'the synthesis' become, that few serious historically minded analysts would touch the subject, let alone know where to begin to sort through the interpretive mess left behind by the numerous critics and commentators".[107]
Смотрите также
- Developmental systems theory
- Gene-centered view of evolution
- История эволюционной мысли
- Neo-Darwinism
- Возражения против эволюции
Примечания
- ^ Also known variously as the New Synthesis, то Modern Evolutionary Synthesis, то Evolutionary Synthesis, а neo-Darwinian Synthesis. These alternative terms are ambiguous as they could possibly include later syntheses, so this article uses Джулиан Хаксли 's 1942 "modern synthesis"[2] throughout.
- ^ Peter Gauthier has however argued that Weismann's experiment showed only that injury did not affect the germplasm. It did not test the effect of Lamarckian use and disuse.[14]
- ^ Morgan's work with fruit flies helped establish the link between Mendelian genetics and the хромосомный theory of inheritance, that the hereditary material was embodied in these bodies within the cell nucleus.[34]
- ^ Fisher also analysed половой отбор in his book, but his work was largely ignored, and Darwin's case for such selection misunderstood, so it formed no substantial part of the modern synthesis.[42]
- ^ Хотя К. Х. Уоддингтон призывал к эмбриология to be added to the synthesis in his 1953 paper "Epigenetics and Evolution".[52]
- ^ In a reissue of Dobzhansky's Генетика и происхождение видов.
Рекомендации
- ^ Гулд 2002, п. 216
- ^ а б Huxley 2010.
- ^ а б c d Koonin, Eugene V. (Ноябрь 2009 г.). "The Origin at 150: is a new evolutionary synthesis in sight?". Тенденции в генетике. 25 (11): 473–475. Дои:10.1016/j.tig.2009.09.007. ЧВК 2784144. PMID 19836100.
- ^ Bowler 2003, pp. 236–256
- ^ Kutschera, Ulrich (Декабрь 2003 г.). "A comparative analysis of the Darwin–Wallace papers and the development of the concept of natural selection". Теория в биологических науках. 122 (4): 343–359. Дои:10.1007/s12064-003-0063-6. S2CID 24297627.
- ^ Батлер, Сэмюэл (1880). Unconscious Memory. David Bogue. п. 280.
I may predict with some certainty that before long we shall find the original Darwinism of Dr. Эразм Дарвин … generally accepted instead of the neo-Darwinism of to-day, and that the variations whose accumulation results in species will be recognised as due to the wants and endeavours of the living forms in which they appear, instead of being ascribed to chance, or, in other words, to unknown causes, as by Mr. Charles Darwin's system
- ^ Beccaloni, George (2013). "On the Terms "Darwinism" and "Neo-Darwinism"". A. R. Wallace Website.
- ^ Bowler 2003, pp. 196–253
- ^ Ларсон 2004, pp. 105–129
- ^ Gayon, Jean (1998). Darwinism's Struggle for Survival: Heredity and the Hypothesis of Natural Selection. Издательство Кембриджского университета. С. 2–3. ISBN 978-0-521-56250-8.
- ^ Дарвин, Чарльз (1868). Изменения животных и растений при одомашнивании. Джон Мюррей. ISBN 978-1-4191-8660-8.
- ^ Holterhoff, Kate (2014). "The History and Reception of Charles Darwin's Hypothesis of Pangenesis". Журнал истории биологии. 47 (4): 661–695. Дои:10.1007/s10739-014-9377-0. PMID 24570302. S2CID 207150548.
- ^ Weismann, August (1892). Das Keimplasma: eine Theorie der Vererbung [The Germ Plasm: A theory of inheritance]. Jena: Fischer.
- ^ Gauthier, Peter (March–May 1990). "Does Weismann's Experiment Constitute a Refutation of the Lamarckian Hypothesis?". BIOS. 61 (1/2): 6–8. JSTOR 4608123.
- ^ Bowler 1989, п. 248.
- ^ Bowler 2003, pp. 253–256
- ^ Bowler 1989, pp. 247–253, 257.
- ^ Ambrose, Mike. "Mendel's Peas". Norwich, UK: Germplasm Resources Unit, Центр Джона Иннеса. Архивировано из оригинал 14 июня 2016 г.. Получено 14 декабря 2017.
- ^ "Reviewing Mendel's Laws". Пирсон. Получено 14 октября 2017.
- ^ Bateson 1894: Mutations (as 'sports') and polymorphisms were well known long before the Mendelian recovery.
- ^ Ларсон 2004, стр. 157–166
- ^ Bowler 1989, стр. 275–276
- ^ а б Grafen & Ridley 2006, п. 69
- ^ Provine 2001, п. 69
- ^ Olby, Robert (September 1989). "The Dimensions of Scientific Controversy: The Biometric-Mendelian Debate". Британский журнал истории науки. 22 (3): 299–320. Дои:10.1017/s0007087400026170. JSTOR 4026898. PMID 11621982.
- ^ Gayon, J. (1988). Darwinism's Struggle for Survival: Heredity and the Hypothesis of Natural Selection. Издательство Кембриджского университета.
- ^ Stoltzfus, Arlin; Cable, Kele (2014). "Mendelian-Mutationism: The Forgotten Evolutionary Synthesis" (PDF). Журнал истории биологии. 47 (4): 501–546. Дои:10.1007/s10739-014-9383-2. PMID 24811736. S2CID 23263558.
- ^ Bateson, William; Saunders, E. R. (1902). "Experimental Studies in the Physiology of Heredity". Королевское общество. Reports to the Evolution Committee.
- ^ Yule, G. Udny (1902). "Mendel's Laws and their probable relations to inter-racial heredity". Новый Фитолог. 1 (10): 226–227. Дои:10.1111/j.1469-8137.1902.tb07336.x.
- ^ Provine 2001 С. 81–82.
- ^ Provine 2001, pp. 109–114.
- ^ Castle, W. E. (7 April 1905). "The Mutation Theory of Organic Evolution, from the Standpoint of Animal Breeding". Наука. 21 (536): 521–525. Bibcode:1905Sci....21..521C. Дои:10.1126/science.21.536.521. ISSN 0036-8075. PMID 17770959.
- ^ Vicedo, Marga (2000). Castle, William Ernest (1867-1962), biologist. Американская национальная биография в Интернете. Издательство Оксфордского университета. Дои:10.1093/anb/9780198606697.article.1302308.
- ^ а б Bowler 2003, стр. 271–272
- ^ Provine 2001 С. 120–121.
- ^ а б Smocovitis 1996, pp. 100–114
- ^ Smocovitis 1996, pp. 114–119
- ^ Smocovitis 1996, стр. 119–122
- ^ Fisher, Ronald A. (January 1919). "XV.—The Correlation between Relatives on the Supposition of Mendelian Inheritance". Сделки Королевского общества Эдинбурга. 52 (2): 399–433. Дои:10.1017 / S0080456800012163. OCLC 4981124. "Paper read by J. Arthur Thomson on July 8, 1918 to the Royal Society of Edinburgh."
- ^ Fisher 1999
- ^ а б c d е ж грамм час Ларсон 2004, pp. 221–243
- ^ Хоскен, Дэвид Дж .; House, Clarissa M. (25 January 2011). "Sexual Selection". Текущая биология. 21 (2): R62–R65. Дои:10.1016/j.cub.2010.11.053. PMID 21256434. S2CID 18470445.
- ^ а б c d е ж Bowler 2003, pp. 325–339
- ^ а б Smocovitis 1996, pp. 122–132
- ^ Smocovitis 1996, п. 192
- ^ Хелд, Льюис И. (2014). Как змея лишилась ног. Curious Tales from the Frontier of Evo-Devo. Издательство Кембриджского университета. п. 67. ISBN 978-1-107-62139-8.
- ^ Gould 1977, стр. 221–222
- ^ Hall, B. K. (2003). "Evo-Devo: evolutionary developmental mechanisms". International Journal of Developmental Biology. 47 (7–8): 491–495. CiteSeerX 10.1.1.113.5158. PMID 14756324.
- ^ Ingo Brigandt (2006). "Homology and heterochrony: the evolutionary embryologist Gavin Rylands de Beer (1899-1972)" (PDF). Журнал экспериментальной зоологии. 306B (4): 317–328. Дои:10.1002/jez.b.21100. PMID 16506229.
- ^ а б Гилберт, С. Ф .; Opitz, J. M.; Raff, R. A. (1996). "Resynthesizing evolutionary and developmental biology". Биология развития. 173 (2): 357–372. Дои:10.1006/dbio.1996.0032. PMID 8605997.
- ^ Adams, M. (1991). Warren, L .; Koprowski, H. (eds.). Through the looking glass: The evolution of Soviet Darwinism. New Perspectives in Evolution. Liss/Wiley. стр.37–63. ISBN 978-0-471-56068-5.
- ^ а б Smocovitis 1996, pp. 153–171
- ^ Wright 1932, стр.356–366
- ^ а б Rose, Michael R.; Oakley, Todd H. (November 24, 2007). "The new biology: beyond the Modern Synthesis" (PDF). Биология Директ. 2 (30): 30. Дои:10.1186/1745-6150-2-30. ЧВК 2222615. PMID 18036242.
- ^ Хаксли, Джулиан (1942). Эволюция: современный синтез. Аллен и Анвин.
- ^ Ridley, Matt (1996). Эволюция (2-е изд.). Blackwell Science. ISBN 978-0632042920.
- ^ Mayr & Provine 1998, п. 231
- ^ Dobzhansky 1937
- ^ Smocovitis 1996, п. 127
- ^ Eldredge, Niles (1985). Unfinished Synthesis: Biological Hierarchies and Modern Evolutionary Thought. Издательство Оксфордского университета. п. 17. ISBN 978-0-19-536513-9.
- ^ а б Ford 1964
- ^ Ford 1975
- ^ Ford, E. B. (1949). Mendelism and Evolution. Метуэн.
- ^ Dobzhansky 1951
- ^ Schmalhausen, Ivan I.; Dordick, Isadore (trans.) (1949). Dobzhansky, Theodosius (ред.). Factors of Evolution. The Theory of Stabilizing Selection. Philadelphia and Toronto: The Blakiston Company.
- ^ Левит, Георгий С .; Хоссфельд, Уве; Олссон, Леннарт (2006). "From the 'Modern Synthesis' to Cybernetics: Ivan Ivanovich Schmalhausen (1884–1963) and his Research Program for a Synthesis of Evolutionary and Developmental Biology". Журнал экспериментальной зоологии. 306B (2006): 89–106. Дои:10.1002 / jez.b.21087. PMID 16419076.
- ^ Adams, M. B. (June 1988). «Недостающее звено в эволюционном синтезе. И. И. Шмальгаузен. Факторы эволюции: теория стабилизирующего отбора». Исида. 79 (297): 281–284. Дои:10.1086/354706. PMID 3049441.
- ^ Glass, Bentley (December 1951). "Reviews and Brief Notices Factors of Evolution. The Theory of Stabilizing Selection. I. I. Schmalhausen, Isadore Dordick, Theodosius Dobzhansky". Quarterly Review of Biology. 26 (4): 384–385. Дои:10.1086/398434.
- ^ Mayr & Provine 1998, п. ix.
- ^ а б c Ruse 1996, pp. 328–338
- ^ а б Ламм, Эхуд. "Review of Julian Huxley, Evolution: The Modern Synthesis – The Definitive Edition, with a new foreword by Massimo Pigliucci and Gerd B. Müller. MIT Press" (PDF). Получено 21 августа 2017.
- ^ а б c d Smocovitis 1996, pp. 138–153
- ^ Mayr 1999
- ^ Hey, Jody; Fitch, Walter M .; Ayala, Francisco J. (2005). "Systematics and the origin of species: An introduction". PNAS. 102 (supplement 1): 6515–6519. Bibcode:2005PNAS..102.6515H. Дои:10.1073/pnas.0501939102. ЧВК 1131868. PMID 15851660.
- ^ Mayr & Provine 1998, стр. 33–34
- ^ Mayr 1982.
- ^ Rensch 1947; Rensch 1959
- ^ Смит, Чарльз Х. "Rensch, Bernhard (Carl Emmanuel) (Germany 1900-1990)". Some Biogeographers, Evolutionists and Ecologists: Chrono-Biographical Sketches. Университет Западного Кентукки. Получено 14 декабря 2017.
- ^ Mayr & Provine 1998, pp. 298–299, 416
- ^ Smocovitis, V. B. (2001). "G. Ledyard Stebbins and the evolutionary synthesis". Ежегодный обзор генетики. 35: 803–814. Дои:10.1146/annurev.genet.35.102401.091525. PMID 11700300.
- ^ а б c d Mayr, E.: Where Are We? Cold Spring Harbor Symposium of Quantitative Biology 24, 1–14, 1959
- ^ а б c d е Stebbins, G.L.: Processes of Organic Evolution, p. 12. Prentice Hall, 1966
- ^ а б c Dobzhansky, T.: In: Ayala, F., Dobzhansky, T. (eds.) Chance and Creativity in Evolution, pp. 307–338. University of California Press, Berkeley and Los Angeles (1974)
- ^ Mayr, E.: In: Mayr, E., Provine, W. (eds.) Some Thoughts on the History of the Evolutionary Synthesis, pp. 1–48. Harvard University Press, 1980
- ^ а б Hamilton, W. D. (1964). "The Genetical Evolution of Social Behaviour I" (PDF). J. Theor. Биол. 7 (1): 1–52. Дои:10.1016/0022-5193(64)90038-4. PMID 5875341.
- ^ а б Уильямс, Джордж К. (1966). Адаптация и естественный отбор. Издательство Принстонского университета. п. 307. ISBN 978-0-691-02615-2.
- ^ а б c d е Yudell, Michael; Desalle, Rob (2000). "Sociobiology. The New Synthesis. by Edward O. Wilson". Журнал истории биологии. 33 (3): 577–584. Дои:10.1023/a:1004845822189. JSTOR 4331614. S2CID 88047081.
- ^ а б Gilbert, S. F. (2000). A New Evolutionary Synthesis. Биология развития. Sinauer Associates.
- ^ Maynard Smith, John (1964). «Групповой отбор и родственный отбор». Природа. 201 (4924): 1145–1147. Bibcode:1964Натура.201.1145S. Дои:10.1038 / 2011145a0. S2CID 4177102.
- ^ Pinker, Steven (1994). Языковой инстинкт. Harper Perennial Modern Classics. п. 294.
- ^ Okasha, Samir (September 22, 2006). "Population Genetics" – via plato.stanford.edu. Цитировать журнал требует
| журнал =(помощь) - ^ Докинз, Р. (1982). Расширенный фенотип. Издательство Оксфордского университета. ISBN 978-0-19-288051-2.
- ^ Fisher, Helen (16 October 1994). "'Wilson,' They Said, 'Your All Wet!'". Нью-Йорк Таймс. Получено 21 июля 2015.
- ^ Lenski, Gerhard (1976). "Sociobiology. The New Synthesis. by Edward O. Wilson". Социальные силы. 55 (2): 530–531. Дои:10.2307/2576242. JSTOR 2576242.
- ^ Smocovitis 1996 С. 186–187.
- ^ а б Пильуччи, Массимо; Finkelman, Leonard (2014). "The Extended (Evolutionary) Synthesis Debate: Where Science Meets Philosophy". Бионаука. 64 (6): 511–516. Дои:10.1093/biosci/biu062.
- ^ а б c Noble, Denis (2011). "Neo-Darwinism, the Modern Synthesis and selfish genes: are they of use in physiology?". Журнал физиологии. 589 (5): 1007–1015. Дои:10.1113/jphysiol.2010.201384. ЧВК 3060581. PMID 21135048.
- ^ Wade, Michael J. (2011). "The Neo-Modern Synthesis: The Confluence of New Data and Explanatory Concepts". Бионаука. 61 (5): 407–408. Дои:10.1525/bio.2011.61.5.10.
- ^ а б John Odling-Smee и другие. "The extended evolutionary synthesis: its structure, assumptions and predictions". Proceedings of the Royal Society B: Biological Sciences, August 2015.
- ^ Wade, Michael J. (2011). "The Neo-Modern Synthesis: The Confluence of New Data and Explanatory Concepts". Бионаука. 61 (5): 407–408. Дои:10.1525/bio.2011.61.5.10.
- ^ Danchin, É.; Charmantier, A.; Champagne, F. A.; Mesoudi, A.; Pujol, B.; Blanchet, S. (2011). "Beyond DNA: integrating inclusive inheritance into an extended theory of evolution". Природа Обзоры Генетика. 12 (7): 475–486. Дои:10.1038/nrg3028. PMID 21681209. S2CID 8837202.
- ^ Laubichler, Manfred D .; Ренн, Юрген (2015). «Расширенная эволюция: концептуальная основа для интеграции регулирующих сетей и создания ниши». Журнал экспериментальной зоологии, часть B: Молекулярная эволюция и эволюция развития. 324 (7): 565–577. Дои:10.1002 / jez.b.22631. ЧВК 4744698. PMID 26097188.
- ^ Kutschera, Ulrich; Niklas, Karl J. (Июнь 2004 г.). "The modern theory of biological evolution: an expanded synthesis". Naturwissenschaften. 91 (6): 255–276. Bibcode:2004NW.....91..255K. Дои:10.1007/s00114-004-0515-y. PMID 15241603. S2CID 10731711.
- ^ Huneman, Philippe; Walsh, Denis M. (2017). Challenging the Modern Synthesis: Adaptation, Development, and Inheritance. Издательство Оксфордского университета. pp. 76 (Chapter 2). ISBN 978-0-19-068145-6.
- ^ Huneman, Philippe; Walsh, Denis M. (2017). Challenging the Modern Synthesis: Adaptation, Development, and Inheritance. Издательство Оксфордского университета. С. Введение. ISBN 978-0-19-068145-6.
- ^ а б Smocovitis 1996, п. 187
- ^ Smocovitis 1996, п. 43
Источники
- Bateson, William (1894). Materials for the Study of Variation Treated with Especial Regard to Discontinuity in the Origin of Species. Лондон; Нью-Йорк: Macmillan. LCCN 07001821. OCLC 1944285.
- Bowler, Peter J. (2003). Evolution: The History of an Idea (3rd completely rev. and expanded ed.). Беркли, Калифорния: Калифорнийский университет Press. ISBN 978-0-520-23693-6.
- Bowler, Peter J. (1989). Evolution: The History of an Idea (2-е изд. Перераб.). Беркли, Калифорния: Калифорнийский университет Press. ISBN 978-0-520-06386-0.
- Dobzhansky, Theodosius (1937). Генетика и происхождение видов. Биологические серии Колумбийского университета. Нью-Йорк: Columbia University Press. LCCN 37033383. OCLC 766405.
- —— (1951). Генетика и происхождение видов. Columbia University Biological Series (3rd revised ed.). Нью-Йорк: издательство Колумбийского университета. LCCN 51014816. OCLC 295774.
- Фишер, Р.А. (1999) [1930]. Генетическая теория естественного отбора. Edited with a foreword and notes by J. H. Bennett (A complete variorum ed.). Оксфорд, Великобритания: Издательство Оксфордского университета. ISBN 978-0-19-850440-5.
- Ford, E. B. (1964). "Ecological Genetics". Развитие науки. Лондон; Нью-Йорк: Methuen Publishing; Джон Уайли и сыновья. 25 (124): 227–35. LCCN 64009038. OCLC 875507052. PMID 5701915.
- Gould, Stephen Jay (1977). Онтогенез и филогения. Belknap Press, издательство Harvard University Press. ISBN 978-0-674-63940-9.
- Gould, Stephen Jay (2002). The Structure of Evolutionary Theory. Belknap Press издательства Гарвардского университета. ISBN 978-0-674-00613-3.
- Grafen, Alan; Ridley, Mark, ред. (2006). Ричард Докинз: Как ученый изменил наше мышление. Издательство Оксфордского университета. ISBN 978-0-19-929116-8.
- Huxley, Julian (2010) [1942]. Эволюция: современный синтез. With a new foreword by Массимо Пильуччи и Герд Б. Мюллер (Окончательный ред.). Кембридж, Массачусетс: MIT Press. ISBN 978-0-262-51366-1.
- Larson, Edward J. (2004). Эволюция: замечательная история научной теории. Modern Library Chronicles. 17. Нью-Йорк: Современная библиотека. ISBN 978-0-679-64288-6.
- Майр, Эрнст (1982). The Growth of Biological Thought: Diversity, Evolution, and Inheritance. Кембридж, Массачусетс: Belknap Press издательства Гарвардского университета. ISBN 978-0-674-36445-5.
- Mayr, Ernst (1999) [1942]. Systematics and the Origin of Species from the Viewpoint of a Zoologist (1st Harvard University Press pbk. Ed.). Издательство Гарвардского университета. ISBN 978-0-674-86250-0.
- Mayr, Ernst; Provine, William B., ред. (1998) [1980]. The Evolutionary Synthesis: Perspectives on the Unification of Biology. With a new preface by Ernst Mayr (1st paperback ed.). Кембридж, Массачусетс: Издательство Гарвардского университета. ISBN 978-0-674-27226-2.
- Provine, W. B. (2001). The Origins of Theoretical Population Genetics, with a new afterword. Издательство Чикагского университета. ISBN 978-0-226-68464-2.
- Rensch, Bernhard (1947). Neuere Probleme der Abstammungslehre. Die transspezifische Evolution [Newer Problems of Evolutionary Theory: The trans-specific Evolution] (на немецком). Stuttgart: Ferdinand Enke Verlag. OCLC 2271422.
- Русе, Майкл (1996). Monad to man: the Concept of Progress in Evolutionary Biology. Издательство Гарвардского университета. ISBN 978-0-674-03248-4.
- Смоковитис, Василики Бетти (1996). Объединяющая биология: эволюционный синтез и эволюционная биология. Журнал истории биологии. 25. Принстон, Нью-Джерси: Издательство Принстонского университета. С. 1–65. Дои:10.1007/bf01947504. ISBN 978-0-691-03343-3. PMID 11623198. S2CID 189833728.
- Райт, Сьюэлл (1932). "The Roles of Mutation, Inbreeding, Crossbreeding and Selection in Evolution". В Jones, Donald F. (ред.). Proceedings of the Sixth International Congress of Genetics. 1. Итака, штат Нью-Йорк: Общество генетиков Америки. OCLC 439596433.
дальнейшее чтение
- Аллен, Гарланд Э. (1978). Thomas Hunt Morgan: The Man and His Science. Принстон, штат Нью-Джерси: Princeton University Press. ISBN 978-0-691-08200-4.
- Докинз, Ричард (1990). The Blind Watchmaker. Penguin Science. Лондон: Книги о пингвинах. ISBN 978-0-14-014481-9.
- Футуйма, Дуглас Дж. (1998). Эволюционная биология (3-е изд.). Sunderland, MA: Sinauer Associates. ISBN 978-0-87893-189-7.
- Холдейн, Дж. Б. С. (1932). The Causes of Evolution. Лондон; Нью-Йорк: Longmans, Green & Co. LCCN 32033284. OCLC 5006266. "This book is based on a series of lectures delivered in January 1931 at the Prifysgol Cymru, Aberystwyth, and entitled 'A re-examination of Darwinism'."
- Хаксли, Джулиан, изд. (1976) [Originally published 1940; Оксфорд, Великобритания: Кларендон Пресс ]. The New Systematics (Перепечатка ред.). St. Clair Shores, MI: Scholarly Press. ISBN 978-0-403-01786-7.
- Levinson, Gene (2020). Rethinking evolution: the revolution that's hiding in plain sight. World Scientific. ISBN 9781786347268.
- Райт, Сьюэлл (March 1, 1931). "Evolution in Mendelian Populations". Генетика. 16 (2): 97–159. ЧВК 1201091. PMID 17246615.
