Фитнес-пейзаж - Fitness landscape

В эволюционная биология, фитнес-пейзажи или же адаптивные ландшафты (виды эволюционные пейзажи ) используются для визуализации взаимосвязи между генотипы и репродуктивный успех. Предполагается, что каждый генотип имеет четко определенную скорость репликации (часто называемую фитнес ). Этот фитнес - «высота» пейзажа. Сходные генотипы считаются «близкими» друг к другу, тогда как очень разные генотипы «далеки» друг от друга. Набор всех возможных генотипов, их степень сходства и связанные с ними значения пригодности затем называют ландшафтом пригодности. Идея фитнес-ландшафта - это метафора, помогающая объяснить ошибочные формы в эволюции естественный отбор, включая эксплойты и сбои у животных, например их реакцию на сверхнормальные стимулы.
Идея изучения эволюции путем визуализации распределения значений приспособленности как некоего ландшафта была впервые представлена Сьюэлл Райт в 1932 г.[1]
В эволюционная оптимизация проблемы, фитнес-пейзажи - это оценки фитнес-функция для всех возможных решений (см. ниже).
В биологии

Во всех сферах фитнеса рост представляет и является визуальной метафорой для фитнес. Есть три различных способа описания других измерений, хотя в каждом случае расстояние представляет и является метафорой степени несходства.[2]
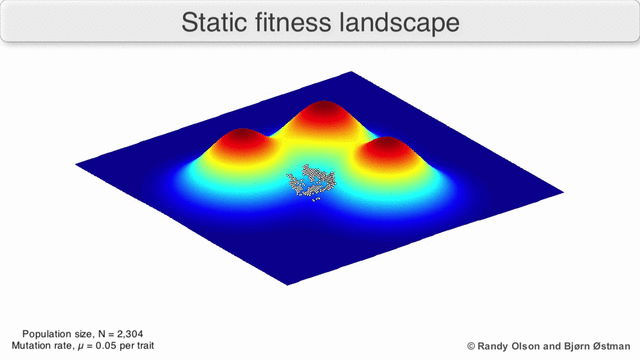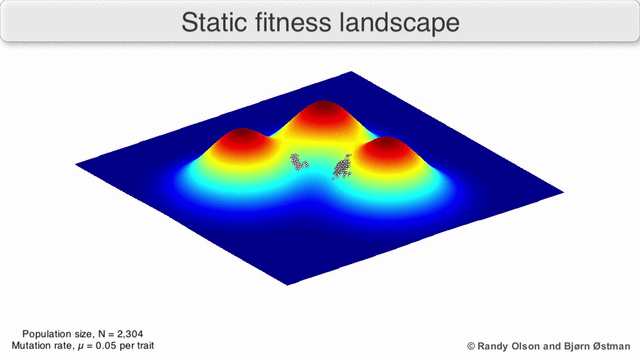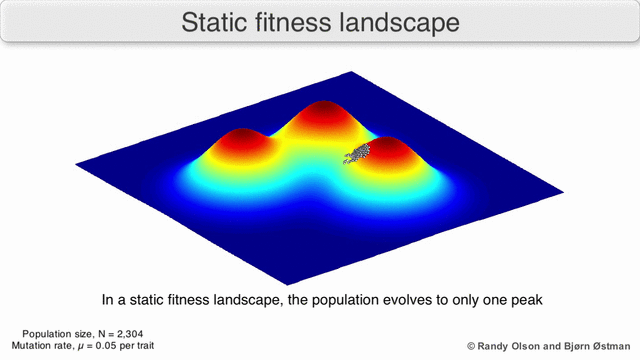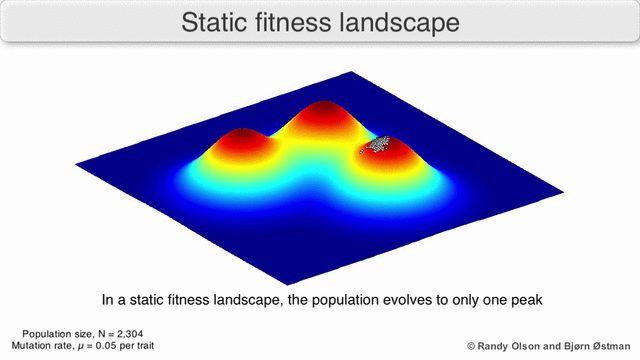
Фитнес-пейзажи часто представляют собой горные хребты. Существуют локальные вершины (точки, от которых все пути идут вниз, то есть к более низкой пригодности) и впадины (регионы, от которых многие пути ведут в гору). Фитнес-ландшафт с множеством местных вершин, окруженных глубокими долинами, называется скалистым. Я упал генотипы имеют одинаковую скорость репликации, с другой стороны, фитнес-ландшафт считается плоским. Развивающийся численность населения обычно поднимается в гору в фитнес-ландшафте за счет серии небольших генетических изменений, пока - за бесконечное время - не будет достигнут локальный оптимум.
Обратите внимание, что локальный оптимум не всегда может быть найден даже во время эволюции: если локальный оптимум может быть найден за разумный промежуток времени, то ландшафт пригодности называется «легким», а если требуемое время экспоненциально, то ландшафт пригодности называется « жесткий".[3] Твердые ландшафты характеризуются свойством лабиринта, благодаря которому аллель, который когда-то был полезен, становится вредным, заставляя эволюцию отступить. Однако наличие свойства лабиринта в ландшафтах фитнеса, вдохновленных биофизикой, может быть недостаточным для создания жесткого ландшафта.[4]
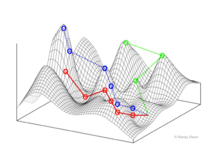
Генотип фитнес-ландшафтов
Райт визуализировал пространство генотипов как гиперкуб.[1] Никакого непрерывного «измерения» генотипа не определено. Вместо этого сеть генотипов связана мутационными путями.
Стюарт Кауфман с Модель НК попадает в эту категорию фитнес-ландшафта. Новые методы сетевого анализа, такие как построение графиков притяжения, взвешенных по выбору (SWAG), также используют безразмерное пространство генотипов.[5]
Частота аллелей для фитнес-ландшафтов
Математическая работа Райта описала приспособленность как функцию частот аллелей.[2] Здесь каждое измерение описывает частоту аллеля в другом гене и находится в диапазоне от 0 до 1.
Фенотип фитнес-ландшафтов
В третьем виде ландшафта фитнеса каждое измерение представляет разные фенотипические признаки.[2] Исходя из предположений количественной генетики, эти фенотипические параметры можно сопоставить с генотипами. См. Визуализации ниже, чтобы увидеть примеры фенотипа для фитнес-ландшафтов.
В эволюционной оптимизации
Помимо области эволюционной биологии, концепция фитнес-ландшафта также приобрела значение в эволюционная оптимизация такие методы как генетические алгоритмы или же стратегии эволюции. При эволюционной оптимизации пытаются решить реальные проблемы (например, инженерное дело или же логистика задач), имитируя динамику биологической эволюции. Например, грузовик для доставки с несколькими адресами назначения может следовать по большому количеству различных маршрутов, но только очень немногие из них приведут к короткому времени в пути.
Чтобы использовать эволюционную оптимизацию, нужно определить для всех возможных решение s к интересующей проблеме (то есть к каждому возможному маршруту в случае грузовика для доставки), насколько он «хорош». Это делается путем введения скаляр -значен функция f (s) (скалярное значение означает, что f (s) простое число, например 0,3, а s может быть более сложным объектом, например, списком адресов назначения в случае грузовика), который называется фитнес-функция.
Высота f (s) подразумевает, что s хорошее решение. В случае грузовика для доставки, f (s) может быть количество доставок в час на маршруте s. Лучшее или, по крайней мере, очень хорошее решение затем находится следующим образом: сначала создается совокупность случайных решений. Затем решения видоизменяются и отбираются для тех, которые имеют более высокую пригодность, пока не будет найдено удовлетворительное решение.
Методы эволюционной оптимизации особенно полезны в ситуациях, когда легко определить качество отдельного решения, но трудно перебрать все возможные решения одно за другим (легко определить время вождения для определенного маршрута грузовика доставки. , но практически невозможно проверить все возможные маршруты, когда количество пунктов назначения вырастет до более чем нескольких).
Концепция скалярной функции приспособленности f (s) также соответствует понятию потенциального или функция энергии в физика. Эти две концепции отличаются только тем, что физики традиционно думают о минимизации потенциальной функции, в то время как биологи предпочитают представление о том, что физическая форма максимальна. Следовательно, обратное преобразование потенциальной функции превращает ее в функцию пригодности, и наоборот.
Предостережения и ограничения
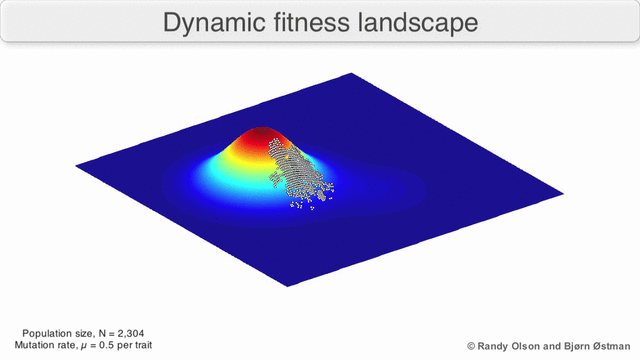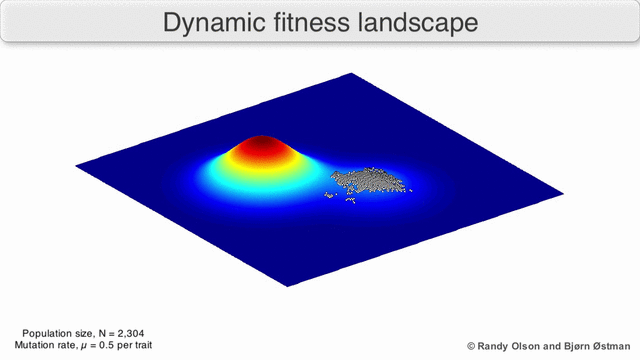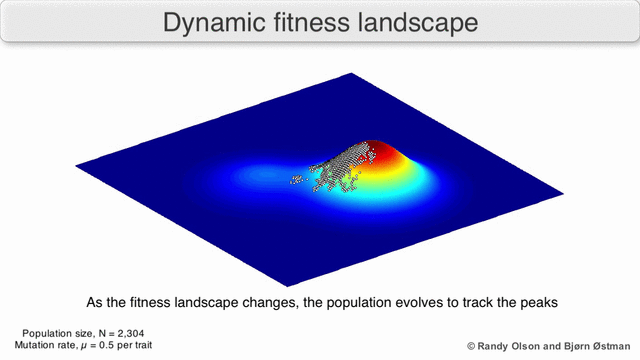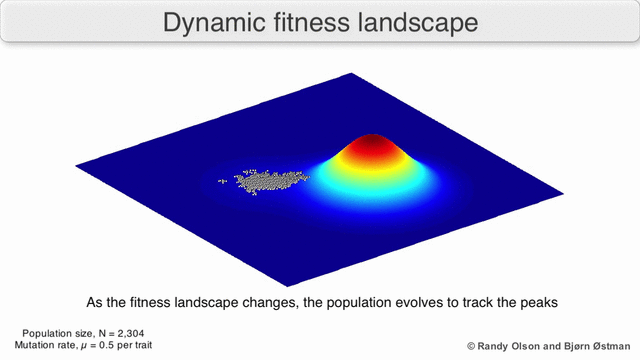
Есть несколько важных оговорок. Поскольку человеческий разум изо всех сил пытается мыслить в более чем трех измерениях, трехмерные топологии могут вводить в заблуждение при обсуждении многомерных ландшафтов фитнеса.[6][7] В частности, неясно, действительно ли пики в естественных ландшафтах биологической пригодности когда-либо действительно разделяются долинами пригодности в таких многомерных ландшафтах, или они связаны чрезвычайно длинными нейтральными хребтами.[8][9] Кроме того, фитнес-ландшафт не статичен во времени, а зависит от меняющейся среды и эволюции других генов.[5] Следовательно, это больше похоже на морской пейзаж,[10] дальнейшее влияние на то, насколько фактически могут быть разделены адаптивные пики. Кроме того, важно принять во внимание, что ландшафт в целом не абсолютная, а относительная функция.[11] Наконец, поскольку при обсуждении ферментов обычно используется функция в качестве показателя приспособленности, любые беспорядочные связи существуют как перекрывающиеся ландшафты, которые вместе определяют окончательную приспособленность организма, подразумевая разрыв между различными сосуществующими относительными ландшафтами.[12]
Помня об этих ограничениях, фитнес-ландшафт может быть поучительным способом размышлять об эволюции. Принципиально возможно измерить (даже если не визуализировать) некоторые параметры неровности ландшафта и количества пиков, высоты, разделения и кластеризации. Затем можно использовать упрощенные трехмерные ландшафты относительно друг друга для визуального представления соответствующих объектов. Кроме того, можно экспериментально построить и визуализировать фитнес-ландшафты небольших подмножеств эволюционных путей, потенциально выявляя такие особенности, как пики и спады приспособленности.[5] Пейзажи приспособленности эволюционных путей указывают на возможные этапы эволюции и конечные точки среди наборов индивидуальных мутаций.
Смотрите также
- Приближение фитнеса
- Функция фитнеса
- Эволюция
- Генетический алгоритм
- Среда обитания (экология)
- скалолазание
- Модель НК
- Возможная функция
- Самоорганизованная критичность
- Телеология в биологии
Рекомендации
- ^ а б Райт, Сьюэлл (1932). «Роли мутации, инбридинга, скрещивания и отбора в эволюции» (PDF). Материалы Шестого Международного конгресса по генетике. 1 (8): 355–66.
- ^ а б c Provine, Уильям Б. (1986). Сьюэлл Райт и эволюционная биология. Издательство Чикагского университета.[страница нужна ]
- ^ Казначеев, Артем (2019). "Вычислительная сложность как главное ограничение эволюции". Генетика. 212 (1): 245–265. Дои:10.1534 / генетика.119.302000. ЧВК 6499524. PMID 30833289.
- ^ Бертрам, Джейсон; Масел, Джоанна (апрель 2020 г.). «Эволюция быстро оптимизирует стабильность и агрегацию в решеточных белках, несмотря на широко распространенные ландшафтные долины и лабиринты». Генетика. 214 (4): 1047–1057. Дои:10.1534 / генетика.120.302815. ЧВК 7153934. PMID 32107278.
- ^ а б c Стейнберг, Б. Остермайер, М (2016). «Изменения окружающей среды соединяют эволюционные долины». Достижения науки. 2 (1): e1500921. Bibcode:2016SciA .... 2E0921S. Дои:10.1126 / sciadv.1500921. ЧВК 4737206. PMID 26844293.
- ^ Маккэндлиш, Дэвид М (2011). «Визуализация фитнес-ландшафтов». Эволюция. 65 (6): 1544–58. Дои:10.1111 / j.1558-5646.2011.01236.x. ЧВК 3668694. PMID 21644947.
- ^ МакГи, Джордж Р. (2006). Геометрия эволюции: адаптивные пейзажи и теоретические морфопространства. ISBN 978-1-139-45995-2.[страница нужна ]
- ^ Гаврилец, С. (2004). Фитнес-пейзажи и происхождение видов. Издательство Принстонского университета. ISBN 978-0-691-11983-0.[страница нужна ]
- ^ Каплан, Джонатан (2008). «Конец адаптивной ландшафтной метафоры?». Биология и философия. 23 (5): 625–38. Дои:10.1007 / s10539-008-9116-z. S2CID 170649453.
- ^ Мустонен, Вилле; Лессиг, Майкл (2009). «От фитнес-ландшафтов к морским пейзажам: неравновесная динамика отбора и адаптации». Тенденции в генетике. 25 (3): 111–9. Дои:10.1016 / j.tig.2009.01.002. PMID 19232770.
- ^ Вудкок, Гленн; Хиггс, Пол Г. (1996). «Эволюция популяции на мультипликативном пиковом ландшафте пригодности». Журнал теоретической биологии. 179 (1): 61–73. Дои:10.1006 / jtbi.1996.0049. PMID 8733432.
- ^ Диас Очоа, Хуан Джи (2017). «Упругие многомасштабные механизмы: вычисления и биологическая эволюция». Журнал молекулярной эволюции. 86 (1): 47–57. Bibcode:2018JMolE..86 ... 47D. Дои:10.1007 / s00239-017-9823-7. PMID 29248946. S2CID 22624633.
внешняя ссылка
- Примеры визуализированных фитнес-ландшафтов
- Видео: Использование фитнес-ландшафтов для визуализации эволюции в действии
- Блог BEACON - Evolution 101: фитнес-пейзажи
- Блог Pleiotropy - интересное обсуждение работ Сергея Гаврильца
- Эволюция щенков - Калифорнийский университет в Дэвисе
- Эволюция 101 - Теория смещения баланса (рисунок внизу страницы)
- Наложение эволюционных траекторий на фитнес-ландшафты в виртуальной реальности
- дальнейшее чтение
- Противовес: эволюция как движение в фитнес-ландшафте - интересное (хотя и ошибочное) обсуждение эволюции и фитнес-ландшафтов.
- Пример использования эволюционных ландшафтов в размышлениях и разговорах об эволюции
- Хендрик Рихтер; Андрис П. Энгельбрехт (2014). Последние достижения в теории и применении фитнес-ландшафтов. ISBN 978-3-642-41888-4.
- Беренвинкель, Нико; Пахтер, Лиор; Штурмфельс, Бернд (2007). «Эпистаз и формы фитнес-ландшафтов». Statistica Sinica. 17 (4): 1317–42. arXiv:q-bio.PE/0603034. Bibcode:2006q.bio ..... 3034B. МИСТЕР 2398598.
- Ричард Докинз (1996). Восхождение на маунт Невероятно. ISBN 0-393-03930-7.
- Сергей Гаврилец (2004). Фитнес-пейзажи и происхождение видов. ISBN 978-0-691-11983-0.
- Стюарт Кауфман (1995). Дома во Вселенной: поиск законов самоорганизации и сложности. ISBN 978-0-19-511130-9.
- Мелани Митчелл (1996). Введение в генетические алгоритмы (PDF). ISBN 978-0-262-63185-3.
- Langdon, W. B .; Поли, Р. (2002). "Глава 2 Фитнес-пейзажи". Основы генетического программирования. ISBN 3-540-42451-2.
- Стюарт Кауфман (1993). Истоки порядка. ISBN 978-0-19-507951-7.
- Poelwijk, Франк Дж; Кивиет, Дэниел Дж; Weinreich, Daniel M; Загар, Сандер Дж (2007). «Пейзажи эмпирической пригодности открывают доступные эволюционные пути». Природа. 445 (7126): 383–6. Bibcode:2007Натура.445..383П. Дои:10.1038 / природа05451. PMID 17251971. S2CID 4415468.