Эволюционный пейзаж - Evolutionary landscape
An эволюционный ландшафт это метафора[1] или конструкция, используемая для размышления и визуализации процессов эволюция (например. естественный отбор и генетический дрейф ) действующие на биологический объект[2] (например, ген, белок, Население, или виды ).[3] Эту сущность можно рассматривать как ищущую или перемещающуюся по пространству поиска. Например, область поиска гена будет включать все возможные нуклеотидные последовательности. Пространство поиска - лишь часть эволюционного ландшафта. Последний компонент - «ось Y», которая обычно фитнес. Каждое значение в пространстве поиска может привести к высокой или низкой пригодности объекта.[1] Если небольшие перемещения в пространстве поиска вызывают относительно небольшие изменения в приспособленности, то ландшафт считается гладким. Гладкие ландшафты возникают, когда большинство фиксированных мутаций практически не влияют на приспособленность, чего и следовало ожидать от нейтральная теория молекулярной эволюции. Напротив, если небольшие движения приводят к большим изменениям в физической форме, тогда ландшафт считается неровным.[1] В любом случае движение, как правило, направлено на области более высокой физической подготовки, хотя обычно это не глобальный оптимум.
Что именно составляет «эволюционный ландшафт», в литературе часто неясно; этот термин часто используется как синонимы «адаптивный ландшафт» и «фитнес-ландшафт», хотя некоторые авторы имеют разные определения адаптивного ландшафта и ландшафта фитнеса. Кроме того, существуют большие разногласия относительно того, следует ли использовать концепцию эволюционного ландшафта в качестве визуальной метафоры, не связанной с лежащей в основе математики, инструмента для оценки моделей эволюции или модели самой по себе, используемой для генерации гипотез и предсказаний.
История
Пре-Райт
Согласно Маккою (1979), первый эволюционный ландшафт был представлен Арманом Жане из Тулона, Франция, в 1895 году.[4] В эволюционном ландшафте Джанет вид представлен как точка или область на многомерной поверхности фенотипов, которая для простоты сокращена до двух измерений. Размер популяции пропорционален количеству вариаций внутри популяции.[5] Естественный отбор (влияние внешних особенностей) представлен вектором. В отличие от эволюционных ландшафтов тех, кто последует за ними, в концепции Джанет естественный отбор подтягивает виды к минимумам, а не к максимумам. Это потому, что ось Y представляет не приспособленность, а стабильность.[4] Одним из важных аспектов эволюционного ландшафта Джанет (по сравнению с ландшафтом Райта) является то, что ландшафт меняется по мере изменения окружающей среды.[5]
Райтовские пейзажи
Заслуга первого эволюционного ландшафта обычно достается Сьюэлл Райт, и его идея, возможно, имела гораздо более широкую аудиторию и большее влияние на область эволюционной биологии, чем любое другое сопоставимое понимание «эволюционного ландшафта».[5] В своей статье 1932 года Райт представляет концепцию эволюционного ландшафта, состоящего из многомерного массива генов (аллель ) или частоты генотипов и ось приспособленности, которая служила визуальной метафорой для объяснения его теория смещения баланса. Как и Джанет, Райт чувствовал, что для простоты пейзаж можно уменьшить до двух измерений. (Это одна из самых серьезных критических замечаний, которые обсуждаются ниже.) Популяции представлены территориями, размер которых соответствует количеству генетического разнообразия в популяции. Естественный отбор подталкивает популяции к максимумам, в то время как дрейф представляет собой блуждание и потенциально может вызвать смещение пика. Движение по ландшафту отражало изменения частот генов.[1] Этот пейзаж был представлен в виде ряда контурных линий, очень похожих на топографическую карту; в то время как отбор сохранял или перемещал биологическую сущность к пику, генетический дрейф позволял исследовать разные пики.[1]
В 1944 году Симпсон расширил ландшафт Райта, включив фенотипы.[6] В модели Симпсона ландшафт - это средство визуализации «взаимосвязи между выбором, структурой и адаптацией».[5][6] В отличие от Райта, Симпсон использовал ландшафт, чтобы представить как естественный отбор, так и генетический дрейф. Подъемы связаны с положительный выбор, а спуски связаны с отрицательный выбор. Размер и форма пика указывали на относительную специфичность отбора; т.е. острый и высокий пик указывает на очень специфический выбор. Еще одно различие между пейзажами Симпсона и Райта - это уровень, на котором действует эволюция. Для Райта популяционный генетик, были показаны только популяции вида. На рисунках Симпсона нарисованные круги представляют всех конных.[6] Самая важная разница заключается в том, что в модели Симпсона пейзаж может меняться со временем, тогда как в модели Райта пейзаж был статичным. Райт сделал обзор работы Симпсона (Темп и мода в эволюции) и не возражал против использования Симпсоном эволюционных ландшафтов.[5] В более поздних работах Симпсон называл пики адаптивными зонами.[7]
В серии статей Рассел Ланде разработал математическую модель фенотипического ландшафта Симпсона.[7][8] Ланде согласовал точку зрения Райта на популяционном уровне с использованием Симпсоном более высоких таксономических уровней.[7] Ланде считает, что пики приспособленности определяются окружающей средой и, таким образом, представляют собой экологические ниши или адаптивные зоны для популяции. Кластеры пиков, населенные фенотипически сходными популяциями, можно рассматривать как более высокие таксономические уровни.[7]
Молекулярная эра
Представления об эволюционных ландшафтах снова изменились по мере появления современного понимания молекулярной эволюции. Утверждается, что Мейнард Смит (1970) был первым, кто визуализировал эволюцию белков как сеть белков, находящуюся на расстоянии одного мутационного шага от других.[5] Однако для этого должны быть пути между функциональными белками. Выражая признательность за работу Кимуры, Кинга и Джукса (нейтральная теория молекулярной эволюции ), Мейнард Смит понял, что белки на таких путях могут иметь одинаковую функциональность или быть нейтральными. Другими словами, не все шаги в эволюции идут «в гору».[5] В 1984 году Гиллеспи адаптировал концепцию эволюционных ландшафтов к нуклеотидным последовательностям и таким образом визуализировал «мутационный ландшафт», согласно которому все нуклеотидные последовательности находятся на расстоянии одного мутационного шага от другого,[5] который удивительно похож, но в то же время фундаментально отличается от первоначальной концепции Райта. Этот концептуальный сдвиг, наряду с развитием огромных вычислительных мощностей, позволил эволюционным ландшафтам превратиться из простой визуальной метафоры в рабочую модель эволюции. Как и следовало ожидать, это вызвало резкую критику и вызвало много исследований.[9]
Критика
Одна из первых критических замечаний (или, по крайней мере, трудностей) в отношении эволюционных ландшафтов - их размерность. Райт признал, что настоящие пейзажи могут иметь тысячи измерений, но он также чувствовал, что уменьшение этих размеров до двух было приемлемым, поскольку его целью было просто передать сложную идею.[1] В качестве визуальной метафоры это могло бы быть правильным сокращением; однако работа Гаврилеца показала, что учет большой размерности эволюционных ландшафтов может иметь значение.[10] В многомерной структуре пики и впадины исчезают и заменяются гиперобъемными областями высокой и низкой пригодности, которые можно визуализировать как изогнутые поверхности и отверстия в трехмерном ландшафте.[10] Пока это не влияет на визуализацию ландшафта. как таковой (т.е. дыры эквивалентны впадинам), это действительно влияет на лежащую в основе математическую модель и прогнозируемые результаты. [[[
Файл: EvoLandscape2.png | thumb | Гипотетический эволюционный ландшафт. Горизонтальные оси представляют собой измеряемые биологические параметры (т. Е. Компонент фенотипа, генотипа, комбинации нуклеотидных последовательностей), а вертикальная ось представляет приспособленность. Этот пейзаж - преувеличенный образец дырявого пейзажа Гаврильца.]]
Работа Гаврильца, наряду с другими проблемами, побудила Каплана (2008) предложить отказаться от метафоры эволюционных ландшафтов (которые он называет адаптивными или приспособленными ландшафтами).[10] Каплан (2008) выдвигает шесть основных критических замечаний по поводу метафоры: (1) у нее нет объяснительной силы; (2) отсутствует соответствующая математическая модель; (3) он не выполняет эвристической роли; (4) неточно; (5) это больше сбивает с толку, чем объясняет; и (6) больше нет причин думать в 2D или 3D, когда у нас есть вычислительные мощности для рассмотрения более высокой размерности. Другие считают, что критика Каплана необоснованна, потому что он (и другие) хотят, чтобы эволюционные ландшафты соответствовали стандартам математической модели; однако метафора пейзажа - это всего лишь метафора. Он имеет эвристическое значение как метафорический инструмент, позволяющий визуализировать и оценивать общее ядро предположений в эволюционной модели.[11]
Хотя Каплан (2008) желает полностью отказаться от идеи пейзажей, Массимо Пильуччи менее радикальный. Он выделяет четыре категории ландшафтов: фитнес-ландшафты, адаптивные ландшафты, фитнес-поверхности и морфопространства. Фитнес-ландшафты похожи на те, что предложил Райт (1932) (ниже они называются адаптивными и фитнес-ландшафтами). Адаптивные ландшафты - это фенотипические ландшафты, предложенные Симпсоном (1944), а фитнес-поверхности - это фенотипические ландшафты с примененными к ним математическими моделями Ланде. Морфопространства, впервые предложенные Раупом (1966), представляют собой фенотипические ландшафты, развитые априори с использованием математических моделей, на которые наносятся наблюдаемые измерения. У них отсутствует ось приспособленности, и они используются для отображения занятых областей в пределах потенциального фенотипического пространства. Пильуччи предлагает отказаться от рисованных фитнес-ландшафтов. Адаптивные ландшафты и фитнес-поверхности можно использовать с осторожностью, т. Е. С пониманием того, что они не являются фенотипическими версиями исходной концепции Райта и чреваты потенциально ошибочными предположениями. Наконец, Пильуччи призывает к дальнейшим исследованиям морфопространств из-за их эвристической ценности, а также их способности генерировать понятные и проверяемые гипотезы.[12]
Типы эволюционных ландшафтов
Адаптивные пейзажи
Адаптивные ландшафты представляют популяции (биологических объектов) в виде единой точки, а оси соответствуют частотам аллели или генотипы и средняя приспособленность популяции.[1][10]
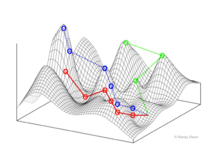
Фитнес-пейзажи
Фитнес-пейзажи представляют популяции (биологических объектов) в виде кластеров точек, каждая из которых представляет уникальный генотип. Оси соответствуют локусам этих генотипов и результирующей средней приспособленности популяции.[10]
Фенотипические пейзажи
Фенотипические ландшафты представляют популяции или виды в виде кластеров точек, каждая из которых представляет собой фенотип. Оси соответствуют частотам фенотипов и средней приспособленности популяции.[10] См. Визуализации ниже для примеров фенотипических ландшафтов.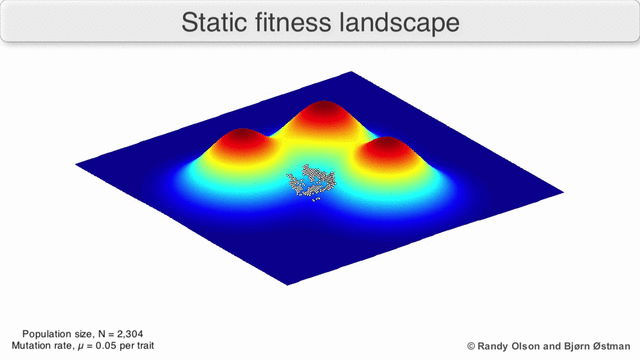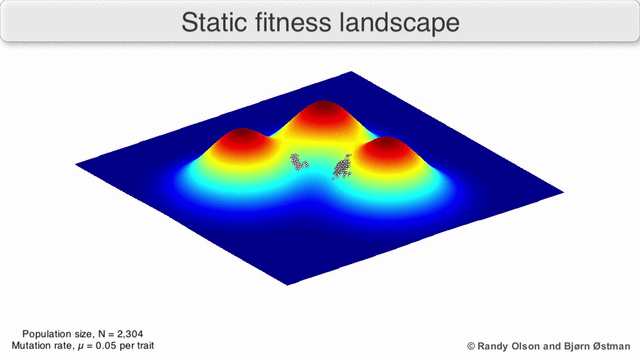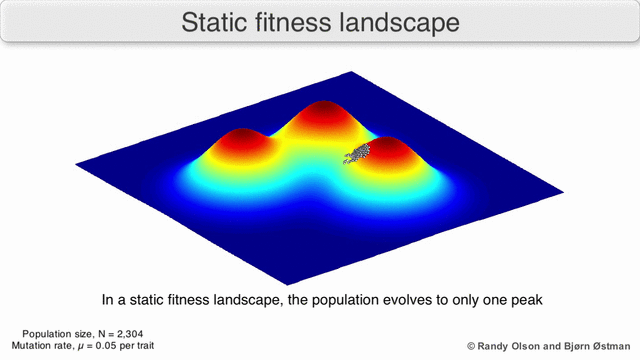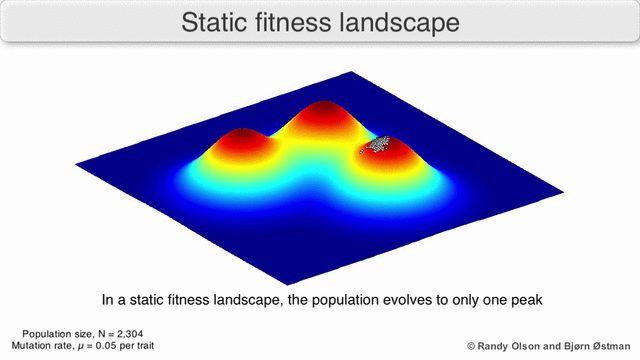
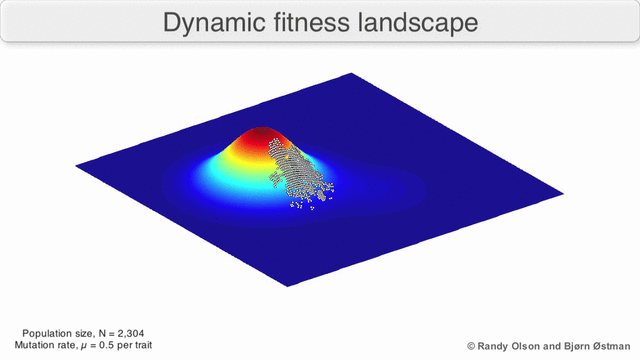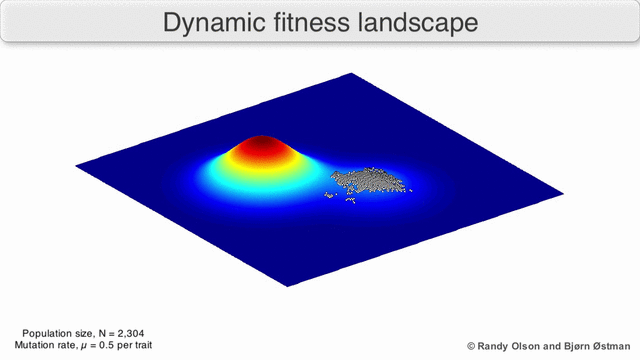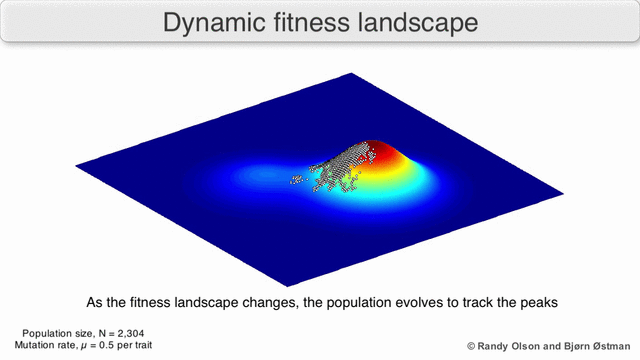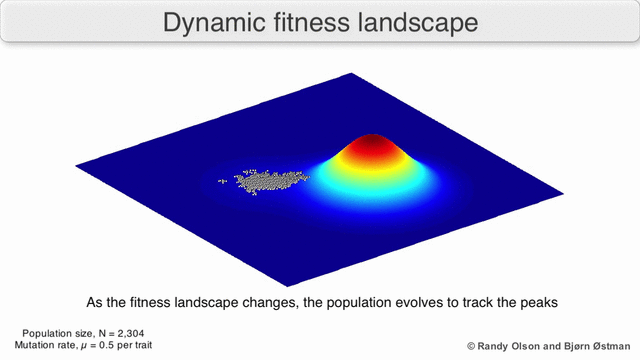
График притяжения, взвешенный по выбору
Взвешенный по выбору график привлекательности (SWAG) использует сетевое построение графиков, ориентированных по силе, для визуализации ландшафтов фитнеса.[13] В этой визуализации генотипы представлены узлами, которые притягиваются друг к другу пропорционально относительному изменению приспособленности между ними (узлы будут располагаться ближе, если в среднем между ними будет более сильная сила отбора). Кроме того, значения пригодности могут быть присвоены оси z, чтобы создать эмпирическую трехмерную модель ландшафта и отобразить пики и спады приспособленности. Кластеры на этом изображении могут представлять локальные пики пригодности.
Фенотипическая пластичность пейзажа
Пейзажи с фенотипической пластичностью отличаются от других ландшафтов тем, что они не используют среднюю приспособленность популяции. Вместо этого эта ось представляет персонажи (фенотипические признаки) а другие оси представляют основные факторы, влияющие на персонажа.[10]
Эпигенетические пейзажи
Эпигенетические пейзажи «используются для описания модальной тенденции развития и основных отклонений» с «пространством абстрактных переменных». [10]
Морфопространства
Морфопространствам также не хватает измерения приспособленности. Вместо этого их оси представляют собой математические модели фенотипических признаков, разработанные априори к наблюдательным измерениям. Затем на полученную поверхность наносятся результаты наблюдений, чтобы указать области возможного фенотипического пространства, занятые рассматриваемыми видами.[12]
Приложения к геномике
Недавнее увеличение вычислительной мощности и простоты построения последовательности позволило превратить концепцию эволюционных ландшафтов из чисто концептуальной метафоры в нечто, что можно моделировать и исследовать. Особенно это касается области геномика. Хорошим примером является исследовательская статья «Эволюционный ландшафт цитозольных микробных сенсоров у людей».[14] В своем исследовании Vasseur et al. интересовались эволюцией врожденная иммунная система; в частности, они хотели составить карту генетического разнообразия - занятого эволюционного ландшафта - и моделей отбора и диверсификации - движений, совершаемых и совершаемых в этом ландшафте - Семейство NOD-подобных рецепторов (NLR) из рецепторы распознавания образов (PRR) которые управляют врожденным иммунным ответом. Эти гены отвечают за обнаружение паттернов / химических веществ (например, хитин, окислительный стресс ) связанные с вторжением микробы, повреждение тканей и стресс. Для этого они секвенировали 21 ген от 185 человек и использовали несколько статистических методов для изучения моделей отбора и диверсификации. Семейство NLR можно разделить на два подсемейства - подсемейство NALP и подсемейство NOD / IPAF. Авторы обнаружили, что семья NALP находилась под сильным очищающий отбор и показали низкое генетическое и функциональное разнообразие. Они предполагают, что это связано с тем, что эти гены играют жизненно важные, неизбыточные роли. Доказательства в пользу этой гипотезы исходят из независимого открытия редких аллелей с мутациями в двух генах, ведущих к тяжелому воспалительному заболеванию и осложнениям беременности. Подсемейство NOD / IPAF, по-видимому, эволюционировало при ослабленном отборе и демонстрирует изрядное генетическое и функциональное разнообразие. Авторы также нашли доказательства положительный выбор. Ген с наиболее сильным положительным отбором - NLRP1, который имеет два гаплотипы проходящий выборочные зачистки. Первый гаплотип - семь аминокислоты в сильном нарушение равновесия по сцеплению. Этот гаплотип глобален и, похоже, движется к фиксации, которая началась в Азии и все еще встречается в Европе и Африке. Второй гаплотип ограничен Европой и не находится в неравновесном сцеплении с глобальным гаплотипом. Этот европейский гаплотип несет в себе мутацию, связанную с аутоиммунными заболеваниями. Авторы предполагают, что отбор действует на другую мутацию в гаплотипе, и эта вредная мутация просто путешествует вместе с ней.[14]
Второй пример взят из статьи под названием «Гены-синонимы исследуют различные эволюционные ландшафты».[15] Авторы этой статьи в целом заинтересованы в способности белка развиваться. Они специально хотели знать, как синонимичные замены повлияли на эволюционный ландшафт белка. Для этого они использовали программу под названием Evolutionary Landscape Printer для разработки синонимичной версии устойчивость к антибиотикам ген aac (6 ') - IB. Синонимичный белок имеет одинаковую аминокислотную последовательность, но разные нуклеотидные последовательности. Таким образом, синонимичный белок имеет ту же функцию и ценность пригодности, но другой окружающий эволюционный ландшафт. По сути, это способ прыгать через вершины, фактически не двигаясь.[15] Ландшафт исходного белка и синонимичной копии был исследован с помощью вычислений. Моделирование Монте-Карло и подвержен ошибкам ПЦР. Затем продукты ПЦР вставляли в компетентный Кишечная палочка клетки и проверены на устойчивость к новым антибиотикам. Они обнаружили, что каждый из двух белков дает начало совершенно разному новому фенотипу, который теоретически недостижим для другого. На основании своих результатов авторы пришли к выводу, что синонимичные кодоны позволяют более широко изучить местный эволюционный ландшафт, и что используемый ими метод увеличивает шансы на обнаружение полезной мутации, что полезно для прогнозирования того, как может измениться популяция, и для лучшего проектирования. организмы для промышленности.[15]
использованная литература
- ^ а б c d е ж г Райт, Сьюолл (1932) Роль мутации, инбридинга, скрещивания и отбора в эволюции. Материалы Шестого Международного конгресса генетиков 1: 356–366
- ^ Райт, Сьюолл (1988). Поверхности выборочной ценности. Американский натуралист 131(1):115-123
- ^ Ли, Кэрол Э. и Гелебюк, Грегори В. (2008) Эволюционное происхождение инвазивных популяций. «Эволюционные приложения» 1: 427–448.
- ^ а б Маккой Дж. Винн. 1979. Происхождение концепции «Адаптивный ландшафт». Американский натуралист 113 (4): 610-613.
- ^ а б c d е ж г час Дитрих М.Р. и шкипер Р.А. 2012 "Меняющийся ландшафт: Краткая история адаптивного ландшафта". В кн .: Адаптивный ландшафт в эволюционной биологии. ред. Эрик Свенссон и Райан Калсбек Oxford Press.
- ^ а б c Симпсон, Г.Г. 1944 г. Темп и мода в эволюции Издательство Колумбийского университета, Нью-Йорк.
- ^ а б c d Ланде, Р. (1976) Естественный отбор и случайный генетический дрейф в фенотипической эволюции. Evolution 30: 314-334.
- ^ Ланде, Р. (1979) Эффективные размеры демов во время долгосрочной эволюции, рассчитанные на основе темпов хромосомной перестройки. Эволюция 33: 234–251
- ^ Адаптивный ландшафт в эволюционной биологии. Ред. Эрик Свенссон и Райан Калсбек. 2012 г.
- ^ а б c d е ж г час Каплан Дж. (2008) Конец метафоры адаптивного ландшафта? Bio Philos 23:625-638
- ^ Шкипер RA и Дитрих MR. 2012 "Адаптивный ландшафт Сьюэлла Райта: философские размышления об эвристической ценности". В кн .: Адаптивный ландшафт в эволюционной биологии. ред. Эрик Свенссон и Райан Калсбек Oxford Press.
- ^ а б Pigliucci M 2012. «Пейзажи, поверхности и морфопространства: для чего они нужны?» В кн .: Адаптивный ландшафт в эволюционной биологии. ред. Эрик Свенссон и Райан Калсбек Oxford Press.
- ^ Стейнберг, Барретт; Остермайер, Марк (01.01.2016). «Изменения окружающей среды мостят эволюционные долины». Достижения науки. 2 (1): e1500921. Bibcode:2016SciA .... 2E0921S. Дои:10.1126 / sciadv.1500921. ISSN 2375-2548. ЧВК 4737206. PMID 26844293.
- ^ а б Вассер Э., М. Бониотто, Э. Патен, Г. Лаваль, Х. Квач, Дж. Манри, Б. Кроу-Рой и Л. Кинтана-Мурси. 2012. Эволюционный ландшафт цитозольных микробных сенсоров у людей. Американский журнал генетики человека 91: 27-37.
- ^ а б c Cambray G&D Mazel. 2008. Синонимичные гены исследуют различные эволюционные ландшафты. PLoS Genetics 4 (11).
дальнейшее чтение
- Противовес: эволюция как движение в фитнес-ландшафте - интересное (хотя и ошибочное) обсуждение эволюции и фитнес-ландшафтов.
- Пример использования эволюционных ландшафтов в размышлениях и разговорах об эволюции
внешние ссылки
Примеры визуализированных эволюционных ландшафтов:
- Видео: Использование фитнес-ландшафтов для визуализации эволюции в действии
- Блог BEACON - Эволюция 101: Фитнес-пейзажи
- Блог Pleiotrophy - интересное обсуждение работ Сергея Гаврилеца
- Эволюция щенков рыбы - Калифорнийский университет в Дэвисе
- Evolution 101 - теория смещения баланса (рисунок внизу страницы)