Доказательства общего происхождения - Evidence of common descent

Доказательства общего происхождения жизни организмы был обнаружен учеными, проводившими исследования в различных дисциплинах на протяжении многих десятилетий, демонстрируя, что вся жизнь на Земле произошла от одного предка. Это составляет важную часть доказательства на котором зиждется эволюционная теория, демонстрирует, что эволюция действительно происходит, и иллюстрирует процессы, которые создали Землю биоразнообразие. Он поддерживает современный эволюционный синтез -электрический ток научная теория это объясняет, как и почему жизнь меняется со временем. Эволюционные биологи документируют доказательства общего происхождения, начиная с древних времен. последний универсальный общий предок путем разработки проверяемых прогнозов, проверки гипотез и построения теорий, которые иллюстрируют и описывают его причины.
Сравнение Генетические последовательности ДНК организмов показал, что организмы, которые филогенетически близкие имеют более высокую степень сходства последовательностей ДНК, чем организмы, которые филогенетически далеки. Генетические фрагменты, такие как псевдогены, регионы ДНК которые ортологичный гену в родственном организм, но больше не активны и, по-видимому, претерпевают устойчивый процесс дегенерации из-за кумулятивных мутаций, что подтверждает общее происхождение наряду с универсальной биохимической организацией и паттернами молекулярной дисперсии, обнаруженными у всех организмов. Дополнительная генетическая информация убедительно подтверждает взаимосвязь жизни и позволила ученым (с момента открытия ДНК) разработать филогенетические деревья: конструкцию эволюционной взаимосвязи организмов. Это также привело к развитию молекулярные часы методы датирования времени расхождения таксонов и их калибровки с летописью окаменелостей.
Окаменелости важны для оценки того, когда в геологическое время. Поскольку окаменелость - редкое явление, обычно требующее твердых частей тела и смерти вблизи места, где отложения депонируются, палеонтологическая летопись дает лишь скудную и непостоянную информацию об эволюции жизни. Свидетельства существования организмов до развития твердых частей тела, таких как раковины, кости и зубы, особенно скудны, но существуют в виде древних микрофоссилий, а также оттиски различных мягкотелых организмов. Сравнительное исследование анатомия групп животных показывает структурные особенности, которые фундаментально похожи (гомологичны), демонстрируя филогенетические и наследственные отношения с другими организмами, особенно по сравнению с окаменелостями древних вымерший организмы. Рудиментарные структуры и сравнения в эмбриональное развитие в значительной степени способствуют анатомическому сходству в соответствии с общим происхождением. поскольку метаболический процессы не оставляют окаменелостей, исследования эволюции основных клеточных процессов в основном проводятся путем сравнения существующих организмов. физиология и биохимия. Многие линии разошлись на разных этапах развития, поэтому можно определить, когда возникли определенные метаболические процессы, сравнив черты потомков общего предка.
Свидетельства из окраска животных был собран некоторыми современниками Дарвина; камуфляж, мимикрия, и предупреждающая окраска все легко объясняются естественный отбор. Особые случаи, такие как сезонные изменения в оперении куропатка, маскируя это против снега зимой и против коричневой вересковой пустоши летом убедительно свидетельствуют о том, что отбор работает. Дальнейшие доказательства получены из области биогеография потому что эволюция по общему происхождению обеспечивает лучшее и наиболее полное объяснение множества фактов, касающихся географического распределения растений и животных по всему миру. Это особенно очевидно в области островная биогеография. В сочетании с хорошо известной геологической теорией тектоника плит общее происхождение дает возможность объединить факты о текущем распространении видов с данными из летописи окаменелостей, чтобы обеспечить логически последовательное объяснение того, как распределение живых организмов изменилось с течением времени.
Развитие и распространение устойчивый к антибиотикам бактерии свидетельствуют о том, что эволюция в результате естественного отбора - это непрерывный процесс в мире природы. Естественный отбор применяется повсеместно во всех исследованиях, касающихся эволюции, с учетом того, что все следующие примеры в каждом разделе статьи документируют этот процесс. Наряду с этим наблюдаются случаи разделения популяций видов на множества новых видов (видообразование ). Видообразование было наблюдается в лаборатории и в природе. Множественные их формы описаны и задокументированы в качестве примеров для отдельных способов видообразования. Более того, доказательства общего происхождения исходят из прямых лабораторных экспериментов с селекция организмов - исторически и в настоящее время - и другие контролируемые эксперименты, затрагивающие многие темы статьи. Эта статья суммирует различные дисциплины, которые предоставляют доказательства эволюции и общего происхождения всей жизни на Земле, сопровождаемые многочисленными специализированными примерами, указывающими на убедительные соответствие доказательств.
Данные сравнительной физиологии и биохимии
Генетика
Одно из самых убедительных доказательств общего происхождения исходит от последовательностей генов. Сравнительный анализ последовательности исследует взаимосвязь между последовательностями ДНК разных видов,[1] приведение нескольких линий доказательств, подтверждающих первоначальную гипотезу Дарвина об общем происхождении. Если гипотеза об общем происхождении верна, то виды, имеющие общего предка, унаследовали последовательность ДНК этого предка, а также мутации, уникальные для этого предка. Более близкородственные виды имеют большую долю идентичных последовательностей и общих замен по сравнению с более отдаленно родственными видами.

Самое простое и убедительное доказательство предоставлено филогенетическая реконструкция. Такие реконструкции, особенно когда они выполняются с использованием медленно развивающихся белковых последовательностей, часто бывают довольно надежными и могут использоваться для реконструкции значительной части эволюционной истории современных организмов (и даже в некоторых случаях эволюционной истории вымерших организмов, таких как восстановленные генные последовательности мамонты или Неандертальцы ). Эти реконструированные филогении резюмируют отношения, установленные в результате морфологических и биохимических исследований.[2] Наиболее детальные реконструкции были выполнены на основе митохондриальных геномов, общих для всех эукариотический организмы[3] которые короткие и легко поддаются последовательности; самые широкие реконструкции были выполнены либо с использованием последовательностей нескольких очень древних белков, либо с использованием рибосомальная РНК последовательность.[нужна цитата ]
Филогенетические отношения распространяются на широкий спектр нефункциональных элементов последовательности, включая повторы, транспозоны, псевдогены, и мутации в кодирующих белки последовательностях, которые не изменяют аминокислотную последовательность. Хотя впоследствии может быть обнаружено, что меньшинство этих элементов выполняет функцию, в совокупности они демонстрируют, что идентичность должна быть продуктом общего происхождения, а не общей функции.[4]
Универсальная биохимическая организация и паттерны молекулярной дисперсии
Все известные сохранившийся (выжившие) организмы основаны на одних и тех же биохимических процессах: генетическая информация, закодированная как нуклеиновая кислота (ДНК, или РНК для многих вирусов), транскрибируется в РНК, затем переведен на белки (то есть полимеры аминокислоты ) очень консервативными рибосомы. Возможно, наиболее показательно то, что Генетический код («таблица трансляции» между ДНК и аминокислотами) одинакова почти для всех организмов, а это означает, что часть ДНК в бактерия кодирует ту же аминокислоту, что и у человека ячейка. АТФ используется как валюта энергии всей существующей жизнью. Более глубокое понимание биология развития показывает, что общая морфология на самом деле является продуктом общих генетических элементов.[5] Например, хотя считается, что глаза, похожие на камеру, развивались независимо во многих отдельных случаях,[6] они имеют общий набор светочувствительных белков (опсины ), предлагая общую точку происхождения для всех зрячих существ.[7][8] Другой пример - знакомый план тела позвоночного, строение которого контролируется семейство генов гомеобокса (Hox).
Секвенирование ДНК
Сравнение последовательностей ДНК позволяет группировать организмы по сходству последовательностей, и в результате филогенетический деревья обычно сочетаются с традиционными таксономия, и часто используются для усиления или исправления таксономической классификации. Сравнение последовательностей считается мерой, достаточно надежной, чтобы исправить ошибочные предположения в филогенетическом дереве в случаях, когда других доказательств мало. Например, нейтральные последовательности ДНК человека примерно на 1,2% отличаются (на основе замен) от последовательностей их ближайшего генетического родственника, шимпанзе, 1,6% от гориллы, и 6,6% от бабуины.[9][10] Таким образом, доказательства генетической последовательности позволяют делать выводы и количественно определять генетическое родство между людьми и другими людьми. обезьяны.[11][12] Последовательность 16S рибосомная РНК ген, жизненно важный ген, кодирующий часть рибосома, был использован для обнаружения широких филогенетических связей между всеми существующими живыми существами. Анализ Карл Вёзе привело к трехдоменная система, утверждая, что на раннем этапе эволюции жизни произошли два основных раскола. Первый раскол привел к современному Бактерии и последующий раскол привел к современному Археи и Эукариоты.
Некоторые последовательности ДНК являются общими для очень разных организмов. Теория эволюции предсказала, что различия в таких последовательностях ДНК между двумя организмами должны примерно напоминать биологическое различие между ними в зависимости от их анатомия и время, прошедшее с тех пор, как эти два организма разделились в ходе эволюции, как показано на ископаемое доказательства. Скорость накопления таких изменений должна быть низкой для некоторых последовательностей, а именно тех, которые кодируют критические РНК или белки и высокий для других, которые кодируют менее важные РНК или белки; но для каждой конкретной последовательности скорость изменения должна быть примерно постоянной во времени. Эти результаты подтверждены экспериментально. Двумя примерами являются последовательности ДНК, кодирующие рРНК, который является высококонсервативным, и последовательности ДНК, кодирующие фибринопептиды (аминокислота цепочки, которые отбрасываются при образовании фибрин ), которые очень неконсервативны.[13]
Белки
Протеомный доказательства также подтверждают универсальное происхождение жизни. Жизненно важно белки, такой как рибосома, ДНК-полимераза, и РНК-полимераза, встречаются во всем: от самых примитивных бактерий до самых сложных млекопитающих. Основная часть белка сохраняется во всех линиях жизни и выполняет аналогичные функции. Высшие организмы развили дополнительные белковые субъединицы, в значительной степени влияя на регулирование и белок-белковое взаимодействие ядра. Другие общие сходства между всеми линиями существующих организмов, такими как ДНК, РНК, аминокислоты и липидный бислой, поддержите теорию общего происхождения. Филогенетический анализ белковых последовательностей различных организмов дает похожие деревья взаимоотношений между всеми организмами.[14] В хиральность ДНК, РНК и аминокислот сохраняется на протяжении всей известной жизни. Поскольку у правой или левой молекулярной хиральности нет функционального преимущества, простейшая гипотеза состоит в том, что выбор был сделан ранними организмами случайным образом и передан всей существующей жизни через общее происхождение. Дальнейшие свидетельства реконструкции родословных предков получены из мусорная ДНК такие как псевдогены, «мертвые» гены, которые неуклонно накапливают мутации.[15]
Псевдогены
Псевдогены, также известные как некодирующая ДНК, представляют собой дополнительные ДНК в геноме, которые не транскрибируются в РНК для синтеза белков. Некоторые из этих некодирующих ДНК обладают известными функциями, но большая их часть не имеет известной функции и называется «мусорной ДНК».[16][17][18][19] Это пример пережитка, поскольку репликация этих генов требует энергии, что во многих случаях делает ее пустой тратой. Псевдоген может образовываться, когда кодирующий ген накапливает мутации, которые препятствуют его транскрипции, что делает его нефункциональным.[16] Но поскольку она не транскрибируется, она может исчезнуть, не влияя на приспособленность, если только она не обеспечила какую-либо полезную функцию в качестве некодирующей ДНК. Нефункциональные псевдогены могут передаваться более поздним видам, тем самым маркируя более поздние виды как потомки более ранних видов.
Прочие механизмы
Большой объем молекулярных доказательств поддерживает множество механизмов больших эволюционных изменений, в том числе: геном и дупликация гена, который способствует быстрой эволюции за счет предоставления значительных количеств генетического материала при слабых селективных ограничениях или без них; горизонтальный перенос генов, процесс передачи генетического материала в другую клетку, которая не является потомком организма, позволяющий видам приобретать полезные гены друг от друга; и рекомбинация, способный переупорядочивать большое количество различных аллели и создания репродуктивная изоляция. В эндосимбиотическая теория объясняет происхождение митохондрии и пластиды (в том числе хлоропласты ), которые органеллы эукариотических клеток, как включение древних прокариотический клетка в древний эукариотический ячейка. Вместо того, чтобы развиваться эукариотический органеллы Постепенно эта теория предлагает механизм внезапного эволюционного скачка за счет включения генетического материала и биохимического состава отдельных видов. Доказательства, подтверждающие этот механизм, были найдены в протист Хатена: как хищник он охватывает зеленые водоросли ячейка, которая впоследствии ведет себя как эндосимбионт, питательный Хатена, который, в свою очередь, теряет свой питающий аппарат и ведет себя как автотроф.[20][21]
поскольку метаболический процессы не оставляют окаменелостей, исследования эволюции основных клеточных процессов проводятся в основном путем сравнения существующих организмов. Многие линии разошлись, когда появились новые метаболические процессы, и теоретически возможно определить, когда появились определенные метаболические процессы, сравнив черты потомков общего предка или обнаружив их физические проявления. Например, появление кислород в земная атмосфера связано с эволюцией фотосинтез.
Конкретные примеры из сравнительной физиологии и биохимии
Хромосома 2 у человека
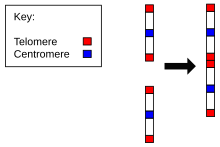
Свидетельства эволюции Homo sapiens от общего предка с шимпанзе обнаруживается по количеству хромосом у людей по сравнению со всеми другими членами Гоминиды. Все гоминиды имеют 24 пары хромосом, за исключением человека, у которого всего 23 пары. Хромосома 2 человека является результатом полного слияния двух наследственных хромосом.[22][23]
Доказательства этого включают:
- Соответствие хромосомы 2 двум хромосомам обезьяны. Ближайший родственник человека, обыкновенный шимпанзе, имеет почти идентичные последовательности ДНК с хромосомой 2 человека, но они находятся в двух отдельных хромосомах. То же самое и с более далекими горилла и орангутанг.[24][25]
- Наличие рудиментарный центромера. Обычно хромосома имеет только одну центромеру, но в хромосоме 2 есть остатки второй центромеры.[26]
- Наличие рудиментарного теломеры. Обычно они находятся только на концах хромосомы, но в хромосоме 2 есть дополнительные последовательности теломер в середине.[27]
Таким образом, хромосома 2 представляет убедительные доказательства в пользу общего происхождения людей и других обезьяны. Согласно J. W. Ijdo, «мы пришли к выводу, что локус, клонированный в космидах c8.1 и c29B, является пережитком древнего слияния теломер-теломер и отмечает точку, в которой слились две предковые хромосомы обезьяны, давая начало хромосоме 2 человека».[27]
Цитохром c и b
Классическим примером биохимического доказательства эволюции является вариация повсеместного (то есть она есть у всех живых организмов, потому что она выполняет очень основные жизненные функции) белок Цитохром с в живых клетках. Дисперсия цитохрома с у разных организмов измеряется количеством различных аминокислот, каждая из которых является результатом определенного базовая пара замена, а мутация. Если каждая различающаяся аминокислота считается результатом замены одной пары оснований, то можно рассчитать, как давно два вида разошлись, умножив количество замен пар оснований на расчетное время, необходимое для замещенной пары оснований гена цитохрома c. быть успешно переданным. Например, если среднее время, необходимое для мутации пары оснований гена цитохрома с, составляет N лет, количество аминокислот, составляющих белок цитохрома с, у обезьян отличается на единицу от такового у людей, это приводит к выводу что два вида разошлись N лет назад.
Первичная структура цитохрома с состоит из цепочки примерно из 100 аминокислоты. Многие организмы более высокого порядка обладают цепочкой из 104 аминокислот.[28]
Молекула цитохрома с была тщательно изучена на предмет того, что она дает в эволюционной биологии. И то и другое курица и индейки имеют идентичную гомологию последовательностей (аминокислота для аминокислоты), как и свиньи, коровы и овец. И то и другое люди и шимпанзе разделяют идентичную молекулу, а макаки резус разделяют все аминокислоты, кроме одной:[29] 66-я аминокислота изолейцин в бывшем и треонин в последнем.[28]
Что делает это гомологичное сходство особенно наводящим на мысль об общем происхождении в случае цитохрома c, помимо того факта, что полученные от них филогении очень хорошо соответствуют другим филогениям, так это высокая степень функциональной избыточности молекулы цитохрома c. Различные существующие конфигурации аминокислот существенно не влияют на функциональность белка, что указывает на то, что замены пар оснований не являются частью направленного дизайна, а являются результатом случайных мутаций, которые не подлежат отбору.[30]
Кроме того, цитохром b обычно используется как область митохондриальная ДНК определить филогенетический отношения между организмами из-за изменчивости последовательности. Считается наиболее полезным при определении отношений внутри семьи и роды. Сравнительные исследования с участием цитохрома b привели к новым схемам классификации и были использованы для отнесения вновь описанных видов к роду, а также для углубления понимания эволюционных взаимоотношений.[31]
Эндогенные ретровирусы
Эндогенные ретровирусы (или ERV) - это остаточные последовательности в геноме, оставшиеся от древних вирусных инфекций в организме. Ретровирусы (или вирогены) всегда прошло следующему поколению того организма, который заразился. В результате вироген остается в геноме. Поскольку это событие является редким и случайным, обнаружение идентичных хромосомных положений вирогена у двух разных видов предполагает общее происхождение.[30] Кошки (Felidae ) представляют собой замечательный пример последовательностей вирогенов, демонстрирующих общее происхождение. Стандартное филогенетическое дерево для кошачьих имеет кошек меньшего размера (Felis chaus, Фелис сильвестрис, Felis nigripes, и Felis catus ) расходящиеся с более крупными кошками, такими как подсемейство Пантеры и другие плотоядные животные. Тот факт, что маленькие кошки имеют ERV, тогда как более крупные кошки не предполагают, что ген был вставлен в предка маленьких кошек после того, как большие кошки разошлись.[32] Другой пример - люди и шимпанзе. Люди содержат множество ERV, которые составляют значительный процент генома. Источники различаются, но 1%[33] до 8%[34] было предложено. У людей и шимпанзе есть семь разных вирогенов, в то время как все приматы имеют сходные ретровирусы, соответствующие филогении.[35][36]
Недавнее африканское происхождение современного человека
Математические модели эволюции, впервые разработанные такими авторами, как Сьюэлл Райт, Рональд Фишер и Дж. Б. С. Холдейн и продлен через теория диффузии от Мотоо Кимура, позволяют делать прогнозы о генетической структуре развивающихся популяций. Прямое изучение генетической структуры современных популяций с помощью секвенирования ДНК позволило проверить многие из этих предсказаний. Например, Из Африки теория происхождения человека, которая утверждает, что современные люди развились в Африке, и небольшая часть населения мигрировала (претерпев узкое место населения ), подразумевает, что современное население должно демонстрировать признаки этой модели миграции. В частности, популяции, пережившие «узкое место» (европейцы и азиаты), должны демонстрировать более низкое общее генетическое разнообразие и более равномерное распределение частот аллелей по сравнению с африканским населением. Оба этих прогноза подтверждаются фактическими данными ряда исследований.[37]
Данные сравнительной анатомии
Сравнительное изучение анатомии групп животных или растений показывает, что некоторые структурные особенности в основном похожи. Например, базовая структура всех цветы состоит из чашелистики, лепестки, стигма, стиль и яичник; еще размер, цвет, количество Состав частей и видовая структура различны для каждого отдельного вида. Нервную анатомию окаменелых останков также можно сравнить с использованием передовых методов визуализации.[38]
Атавизмы
Когда-то считавшиеся опровержением эволюционной теории, атавизмы «теперь рассматриваются как мощное свидетельство того, сколько генетического потенциала сохраняется ... после того, как определенная структура исчезла из вида».[40] «Атавизмы - это повторное появление утраченного персонажа, типичного для далеких предков, которого не видно у родителей или недавних предков ...»[41] и являются «[указанием] на пластичность эмбриона в развитии ...»[41] Атавизмы возникают потому, что гены для ранее существовавших фенотипических признаков часто сохраняются в ДНК, даже если гены не экспрессируются в некоторых или большинстве организмов, обладающих ими.[42] Многочисленные примеры документально подтверждают появление атавизмов наряду с экспериментальными исследованиями, инициирующими их формирование. Из-за сложности и взаимосвязанности факторов, участвующих в развитии атавизмов, и биологи, и медицинские работники считают «трудно, если не невозможно, отличить [их] от пороков развития».[43]
Некоторые примеры атавизмов, найденные в научной литературе, включают:
- Задние конечности в киты.[41] (см. рисунок 2а)
- Повторное появление конечностей у безногие позвоночные.[40][41][44]
- Задняя пара ласт на бутылконосый Дельфин.[45]
- Дополнительные пальцы современной лошади.[41][46][47]
- Человеческие хвосты (не псевдо-хвосты)[43][48] и лишние соски у людей.[41]
- Реэволюция сексуальности из партеногенеза в орбитальные клещи.[49]
- Зубы у кур.[50]
- Dewclaws у собак.[41]
- Повторное появление крыльев у бескрылых палочников.[51] и уховертки.[41]
- Атавистические мышцы у нескольких птиц[52][53] и млекопитающие, такие как гончая[54] и тушканчик.[52]
- Дополнительные пальцы в морские свинки.[41][55]
Эволюционная биология развития и эмбриональное развитие
Эволюционная биология развития - это биологическая область, которая сравнивает процесс развития разных организмов, чтобы определить родовые отношения между видами. Большое разнообразие геномов организмов содержит малая доля генов которые контролируют развитие организмов. Hox-гены являются примером этих типов почти универсальных генов в организмах, указывающих на происхождение от общего предка. Эмбриологические доказательства получены в результате развития организмов на эмбриологическом уровне при сравнении сходства эмбрионов разных организмов. Остатки наследственных признаков часто появляются и исчезают на разных стадиях эмбриологического развития.
Вот некоторые примеры:
- Рост и выпадение волос (лануго ) во время человеческого развития.[56]
- Развитие и дегенерация желточный мешок.
- Земные лягушки и саламандры проходят через личиночную стадию внутри яйца - с чертами типично водных личинок - но вылупляются готовыми к жизни на суше;[57]
- Появление жаберных структур (глоточная дуга ) в развитии эмбриона позвоночных. Обратите внимание, что у рыб дуги продолжают развиваться как жаберные дуги в то время как у людей, например, они вызывают разнообразие конструкций внутри головы и шеи.
Гомологические структуры и дивергентная (адаптивная) эволюция
Если широко разделенные группы организмов произошли от общего предка, ожидается, что они будут иметь некоторые общие черты. Степень сходства между двумя организмами должна указывать на то, насколько тесно они связаны в эволюции:
- Предполагается, что группы, у которых мало общего, отошли от общий предок гораздо раньше в геологической истории, чем группы с большим количеством общего;
- Решая, насколько тесно связаны два животных, сравнительный анатом ищет структуры которые принципиально похожи, хотя они могут выполнять разные функции в взрослый. Такие структуры описываются как гомологичный и предположить общее происхождение.
- В случаях, когда подобные структуры выполняют разные функции у взрослых, может потребоваться проследить их происхождение и эмбриональное развитие. Подобное происхождение предполагает, что они имеют одну и ту же структуру и, следовательно, вероятно, произошли от общего предка.
Когда группа организмов разделяет гомологичную структуру, которая специализируется на выполнении множества функций для адаптации к различным условиям окружающей среды и образу жизни, это называется адаптивное излучение. Постепенное распространение организмов с помощью адаптивной радиации известно как дивергентная эволюция.
Вложенные иерархии и классификация
Таксономия основан на том факте, что все организмы связаны друг с другом во вложенных иерархиях на основе общих характеристик. Большинство существующих видов можно довольно легко организовать во вложенные иерархическая классификация. Это очевидно из схемы классификации Линнея. На основе общих производных признаков близкородственные организмы могут быть помещены в одну группу (например, род), несколько родов могут быть сгруппированы в одно семейство, несколько семейств могут быть сгруппированы вместе в порядок и т. Д.[58] Существование этих вложенных иерархий было признано многими биологами до Дарвина, но он показал, что его теория эволюции с ее ветвящимся паттерном общего происхождения может их объяснить.[58][59] Дарвин описал, как общее происхождение может служить логической основой для классификации:[60]
Все вышеизложенные правила и вспомогательные средства, а также трудности в классификации объясняются, если я не сильно обманываю себя, на той точке зрения, что естественная система основана на происхождении с модификацией; что натуралисты считают, что признаки истинного родства между любыми двумя или более видами - это те, которые унаследованы от общего родителя, и до сих пор вся истинная классификация является генеалогической; это сообщество происхождения является скрытой связью, которую натуралисты бессознательно искали ...
— Чарльз Дарвин, О происхождении видов, стр. 577
Эволюционные деревья
An эволюционное дерево (Амниоты, например, последнего общего предка млекопитающих и рептилий, и всех их потомков) иллюстрируют начальные условия, вызывающие сходство эволюционных паттернов (например, все Амниоты производят яйцо, которое обладает амниос ) и паттерны дивергенции между линиями (например, млекопитающие и рептилии, ответвленные от общего предка в Amniota). Эволюционные деревья предоставляют концептуальные модели развивающихся систем, которые когда-то считались ограниченными в области построения прогнозов на основе теории.[61] Однако метод филогенетический брекетинг используется для получения прогнозов с гораздо большей вероятностью, чем грубые предположения. Например, палеонтологи используют этот метод, чтобы делать прогнозы о несохраняемых чертах у ископаемых организмов, таких как пернатые динозавры, а молекулярные биологи используют этот метод для создания прогнозов о метаболизме РНК и функциях белков.[62][63] Таким образом, эволюционные деревья - это эволюционные гипотезы, которые относятся к конкретным фактам, таким как характеристики организмов (например, чешуя, перья, мех), обеспечивая доказательства закономерностей происхождения и причинное объяснение модификаций (например, естественный отбор или нейтральный дрейф). ) в любой данной линии (например, Амниота). Биологи-эволюционисты проверяют эволюционную теорию, используя филогенетические систематические методы, которые измеряют, насколько гипотеза (конкретный образец ветвления в эволюционном дереве) увеличивает вероятность доказательства (распределение признаков по родословным).[64][65][66] Серьезность проверки теории возрастает, если предсказания «с наименьшей вероятностью будут соблюдены, если причинное событие не произошло».[67] «Проверяемость - это мера того, насколько гипотеза увеличивает вероятность доказательства».[68]
Рудиментарные структуры
Доказательства общего происхождения исходят из существования рудиментарных структур.[69] Эти рудиментарные структуры часто гомологичны структурам, которые соответствуют родственным или предковым видам. Существует широкий спектр структур, таких как мутировавшие и нефункционирующие гены, части цветка, мышцы, органы и даже формы поведения. Это разнообразие встречается во многих различных группах видов. Во многих случаях они дегенерированы или недоразвиты. Существование рудиментарных органов можно объяснить изменениями в окружающей среде или образе жизни вида. Эти органы, как правило, функциональны у предков, но сейчас они либо полуфункциональны, либо нефункциональны, либо используются повторно.
Научной литературы по рудиментарным структурам предостаточно. В одном исследовании было собрано 64 примера рудиментарных структур, обнаруженных в литературе по широкому кругу дисциплин в 21 веке.[70] Следующий неполный список резюмирует Senter et al. наряду с другими примерами:
- Наличие остатка митохондрии (митосомы ), утратившие способность синтезировать АТФ в Entamoeba histolytica, Trachipleistophora hominis, Криптоспоридиум парвум, Blastocystis hominis, и Лямблии кишечные.[71]
- Остаток хлоропласт органеллы (лейкопласты ) у нефотосинтезирующих видов водорослей (Плазмодий falciparum, Toxoplasma gondii, Аспазия длинная, Anthophysa Vegetans, Настой цилиофриса, Pteridomonas danica, Парафизомонады, Спумелла и Эпифаг американский.[72]
- Отсутствует тычинки (без сосудов стаминодии ) на Gilliesia и Gethyum цветы.[73]
- Не работает андроцей в женских цветках и нефункционирующих гинецей в мужских цветках видов кактусов Consolea спиносиссима.[74]
- Остатки тычинок на женских цветках Fragaria virginiana;[75] все виды в роду Schiedea;[76] и дальше Penstemon centranthifolius, P. rostriflorus, П. эллиптический, и P. palmeri.[77]
- Рудиментарный пыльники на Немофила Menziesii.[78]
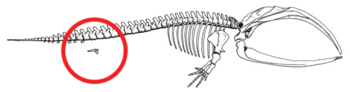
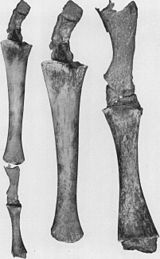
- Уменьшение задних конечностей и тазовый пояс встроены в мышцы сохранившихся киты (см. рисунок 2b).[79][80][81][82] Иногда гены, кодирующие более длинные конечности, заставляют современного кита развивать ноги. 28 октября 2006 г. был пойман и изучен четырехплавный дельфин-афалина из-за лишнего набора задних конечностей.[83] Эти ножки Китообразные показать пример атавизма, предсказанного по их общей родословной.
- Неработающие задние крылья в Карабус Сольери[84] и другие жуки.[80]
- Остаточные глаза (и глазные структуры) у животных, потерявших зрение, таких как слепые пещерные рыбы (например, Astyanax mexicanus ),[85] землекопы, змеи, пауки, саламандры, креветки, раки и жуки.[86][87]
- Рудиментарный глаз в сохранившихся Rhineura floridana и остаток скуловой в вымерших Инкубаторий Rhineura (реклассифицировано как Protorhineura hatcherii ).[88][89]
- Бесполезные крылья нелетающих птиц, таких как страусы, киви, казуары и эму.[90][91]
- Наличие plica semilunaris в человеческом глазу - рудиментарный остаток мигательная перепонка.[92]
- Железа Хардера у приматов.[93]
- Уменьшение структур задних конечностей и тазового пояса у безногие ящерицы, сцинки,[94] амфисбайцы, и несколько змей.[95][96]
- Снижение или отсутствие обонятельного аппарата у китов, которые все еще обладают рудиментарными субгеномами обонятельных рецепторов.[97]
- Рудиментарные зубы в нарвал.[98]
- Рудиментарные цифры Ателес Жоффройи, Колобус гереза, и Perodicticus potto.[99]
- Рудиментарные зачатки зубов в структуре эмбриональных зубов мышей.[100]
- Снижено или отсутствует вомероназальный орган у людей и обезьян Старого Света.[101][102]
- Наличие у людей нефункциональных волосяных мышц пазух, используемых для движения усов.[103]
- Дегенерирующий длинная ладонная мышца мышцы у человека.[104]
- Костистые рыбы, антропоидные приматы (Обезьяны ), морские свинки, некоторые виды летучих мышей, а некоторые Воробьиные утратили способность синтезировать витамин С (аскорбиновая кислота ), но все еще обладают задействованными генами. Эта неспособность связана с мутациями L-гулоно-γ-лактоноксидазы (GLO) ген - а у приматов, костистых рыб и морских свинок он необратим.[105]
- Остаточные сегменты брюшка у усоногих (ракушки ).[106]
- Эмбрионы позвоночных, не являющихся млекопитающими, зависят от питательных веществ из желточный мешок. Геномы человека и других млекопитающих содержат сломанные, нефункционирующие гены, которые кодируют производство желтка. наряду с наличием пустого желточного мешка с эмбрионом.[107][108][109]
- Зародыши зачатков конечностей дельфина.[110]
- Листообразование у некоторых видов кактусов.[111]
- Наличие рудиментарного эндосимбионта Лепидодиниум вирид внутри динофлагелляты Gymnodinium chlorophorum.[112]
- Виды Долабрифера долабрифера имеет чернильную железу, но «неспособен производить чернила или связанные с ними белки против хищников».[113]
Конкретные примеры из сравнительной анатомии
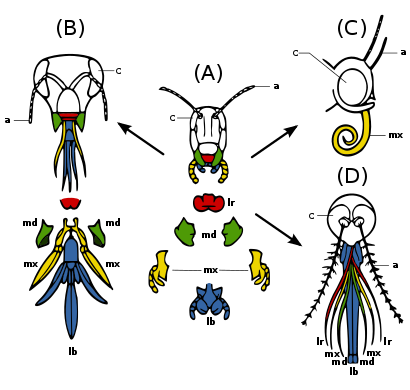
(A) Первобытное состояние - кусание и жевание: например кузнечик. Сильные нижние и верхние челюсти для манипуляций с пищей.
(B) Тикинг и укус: например пчела. Губа долго ласкать нектар; челюсти жевать пыльца и плесень воск.
(C) Сосание: например бабочка. Нижняя губа снижена; нижние челюсти потеряны; maxillae длинные, образующие сосательную трубку.
(D) пирсинг и сосание, например. женский комар. Верхняя и нижняя губа образуют трубку; нижние челюсти образуют пронзительные стилеты; верхняя губа с бороздками для удержания других частей.

Ротовой аппарат и придатки насекомых
У многих разных видов насекомых ротовой аппарат произошел от одних и тех же эмбриональных структур, что указывает на то, что ротовой аппарат является модификацией исходных черт общего предка. К ним относятся верхняя губа (верхняя губа), пара челюсти, а гипофаринкс (дно рта), пара максиллы, а губа. (Рис. 2c) Эволюция вызвала увеличение и изменение этих структур у некоторых видов, в то время как она вызвала их уменьшение и потерю у других видов. Модификации позволяют насекомым использовать различные пищевые материалы.
Ротовой аппарат и усики насекомых считаются гомологами ног насекомых. Параллельное развитие наблюдается в некоторых паукообразные: Передняя пара ног может быть изменена как аналоги усиков, особенно у кнут скорпионов, которые ходят на шести ногах. Эти разработки подтверждают теорию о том, что сложные модификации часто возникают в результате дублирования компонентов, причем дубликаты изменяются в разных направлениях.
Строение таза динозавров
Подобно пентадактильной конечности у млекопитающих, самые ранние динозавры разделены на два отдельных отряда - заурисхия и орнитишия. Они классифицируются как те или иные в соответствии с тем, что демонстрируют окаменелости. Рисунок 2d показывает, что ранние заурисхи напомнил рано орнитисхианы. Образец таз у всех видов динозавров это пример гомологичных структур. У каждого отряда динозавров кости таза немного отличаются, что свидетельствует об их общем происхождении. Кроме того, современные птицы показать сходство с древними заурисхий структуры таза, указывающие на эволюция птиц от динозавров. Это также можно увидеть на рисунке 5c как Авес ответвление от Тероподы подотряд.
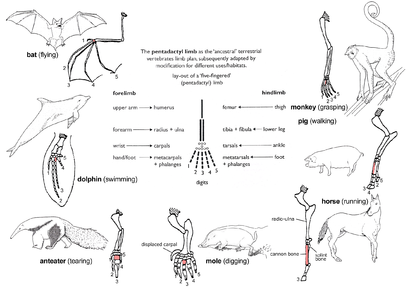
Пентадактильная конечность
Узор костей конечностей называется пентадактильная конечность является примером гомологичных структур (рис. 2д). Встречается во всех классах четвероногие (т.е. от амфибии к млекопитающие ). Это даже можно проследить до плавники определенных ископаемые рыбы от которого произошли первые амфибии, такие как тиктаалик. На конечности имеется единственная проксимальная кость (плечевая кость ), две дистальные кости (радиус и локтевая кость ), серии запястья (запястье кости), за которыми следуют пять серий пястных костей (Пальма кости) и фаланги (цифры). У всех четвероногих фундаментальные структуры конечностей пентадактилей одинаковы, что указывает на их происхождение от общего предка. Но в ходе эволюции эти фундаментальные структуры претерпели изменения. Они стали внешне разными и не связанными друг с другом структурами, выполняющими разные функции при адаптации к разным условиям и образу жизни. Это явление проявляется на передних конечностях млекопитающих. Например:
- В обезьяны, передние конечности сильно удлинены, образуя хватательную руку, используемую для лазания и раскачивания между деревьями.
- Свиньи потеряли свою первую цифру, а вторая и пятая цифры уменьшились. Остальные два пальца длиннее и толще остальных и имеют копыто для поддержки тела.
- У лошадей передние конечности хорошо приспособлены к силе и опоре. Быстрый бег на длинные дистанции возможен благодаря значительному удлинению третьего пальца, несущего копыто.
- В моль имеет пару коротких лопатообразных передних конечностей для роющий.
- Муравьеды использовать увеличенный третий палец для разрыва муравей и термит гнезда.
- У китообразных передние конечности становятся ласты для управления и поддержания равновесия во время плавания.
- В летучие мыши, передние конечности сильно изменились и превратились в функционирующие крылья. Четыре пальца стали удлиненными, а крючковидный первый палец остался свободным и используется для захвата.
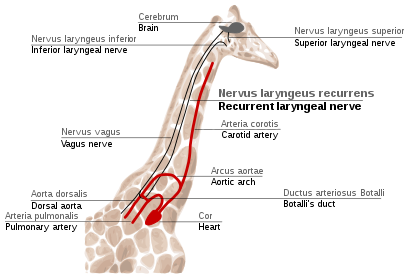
Возвратный гортанный нерв у жирафов
В возвратный гортанный нерв это четвертая ветвь блуждающий нерв, что является черепно-мозговой нерв. У млекопитающих его путь необычайно длинный. Являясь частью блуждающего нерва, он выходит из головного мозга, проходит через шею вниз к сердцу, огибает дорсальная аорта и возвращается к гортань снова через шею. (Рис. 2f)
Этот путь неоптимален даже для человека, но для жирафы он становится еще более неоптимальным. Из-за длины их шеи возвратный гортанный нерв может быть до 4 м (13 футов) в длину, несмотря на то, что его оптимальный маршрут составляет всего несколько дюймов.
Непрямой путь этого нерва - результат эволюции млекопитающих от рыб, у которых не было шеи и был относительно короткий нерв, который иннервировал одну жаберную щель и проходил около жаберной дуги. С тех пор иннервируемая им жабра стала гортани, а жаберная дуга - спинной аортой у млекопитающих.[114][115]
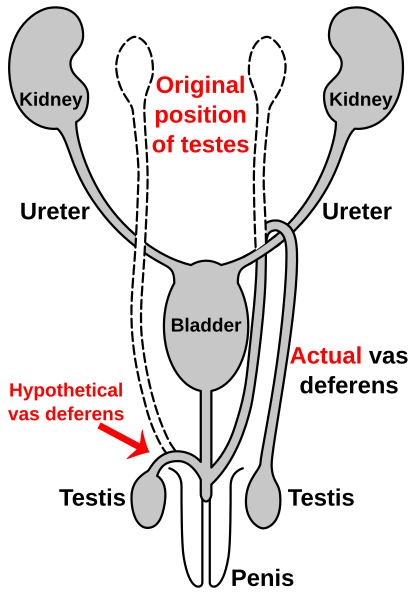
Путь семявыносящего протока
Подобно гортанному нерву у жирафов, семявыносящий проток является частью мужская анатомия из многих позвоночные; транспортирует сперму из придаток яичка в ожидании эякуляция. У людей семявыносящий проток поднимается вверх от яичко, перебирая мочеточник, и обратно к уретра и пенис. Было высказано предположение, что это связано с опусканием яичек в ходе эволюции человека, что, вероятно, связано с температурой. Когда яички опускались, семявыносящие протоки удлинялись, чтобы приспособиться к случайному «зацепу» за мочеточник.[115][116]
Свидетельства палеонтологии





Когда организмы умирают, они часто разлагать быстро или потребляются мусорщики, не оставляя никаких постоянных доказательств их существования. Однако иногда некоторые организмы сохраняются. Остатки или следы организмов из прошлого геологический возраст встроенный в горные породы естественными процессами называются окаменелости. Они чрезвычайно важны для понимания эволюционная история жизни на Земле, поскольку они предоставляют прямые доказательства эволюции и подробную информацию о происхождении организмов. Палеонтология - это изучение прошлой жизни, основанное на записях окаменелостей и их связи с различными геологическими периодами времени.
Чтобы произошло окаменение, следы и останки организмов должны быть быстро захоронены, чтобы выветривание и разложения не происходит. Скелетные структуры или другие твердые части организмов являются наиболее часто встречающейся формой окаменелых останков. Есть также некоторые следы "окаменелостей", показывающие формы, слепки или отпечатки некоторых предыдущих организмов.
Когда животное умирает, органические материалы постепенно разлагаются, так что кости становятся пористыми. Если животное впоследствии захоронят в грязь, минеральная соли проникают в кости и постепенно заполняют поры. Кости превращаются в камни и сохраняются в виде окаменелостей. Этот процесс известен как окаменение. Если мертвые животные накрыты ветром песок, и если песок впоследствии превратится в грязь тяжелыми дождь или наводнения, может произойти тот же процесс минеральной инфильтрации. Помимо окаменения, мертвые тела организмов могут хорошо сохраняться в лед, в закаленном смола из хвойный деревья (рисунок 3а), в смоле или в анаэробной среде, кислый торф. Иногда окаменелость может быть следом, отпечатком формы. Примеры включают листья и следы, окаменелости которых сделаны слоями, которые затем затвердевают.
Окаменелости
Можно расшифровать, как развивалась определенная группа организмов, расположив ее летопись окаменелостей в хронологической последовательности. Такую последовательность можно определить, поскольку окаменелости в основном встречаются в осадочная порода. Осадочная порода образована слоями ил или грязь друг на друга; Таким образом, полученная порода содержит серию горизонтальных слоев, или слои. Каждый слой содержит окаменелости, характерные для определенного временной период когда они образовались. Самые низкие пласты содержат самые старые породы и самые ранние окаменелости, в то время как самые высокие пласты содержат самые молодые породы и более поздние окаменелости.
Последовательность животных и растений также можно увидеть по находкам окаменелостей. Изучая количество и сложность различных окаменелостей в разных стратиграфический На уровнях было показано, что более старые породы, содержащие окаменелости, содержат меньше типов окаменелых организмов и все они имеют более простую структуру, тогда как более молодые породы содержат большее разнообразие окаменелостей, часто со все более сложной структурой.[118]
В течение многих лет геологи могли лишь приблизительно оценить возраст различных слоев и найденных окаменелостей. Они сделали это, например, оценивая время образования осадочных пород слой за слоем. Сегодня, измерив пропорции радиоактивный и стабильный элементы в данной породе ученые могут более точно датировать возраст окаменелостей. Этот метод известен как радиометрическое датирование.
На протяжении всей летописи окаменелостей многие виды, которые появляются на раннем стратиграфическом уровне, исчезают на более позднем уровне. С точки зрения эволюции это интерпретируется как указание времени, когда виды возникли и вымерли. Географические регионы и климатические условия менялись повсюду. История Земли. Поскольку организмы адаптированы к определенной среде, постоянно меняющиеся условия благоприятствовали видам, которые адаптировались к новой среде посредством механизма естественный отбор.
Объем летописи окаменелостей
Несмотря на относительную редкость подходящих условий для окаменелости, было названо около 250 000 ископаемых видов.[119] Число отдельных окаменелостей, которые это представляет, сильно варьируется от вида к виду, но многие миллионы окаменелостей были восстановлены: например, более трех миллионов окаменелостей от последнего Ледниковый период были восстановлены из La Brea Tar Pits в Лос-Анжелес.[120] Еще больше окаменелостей все еще находится в земле в различных геологических формациях, которые, как известно, содержат высокую плотность окаменелостей, что позволяет сделать оценки общего содержания окаменелостей в формации. Пример этого происходит в Южной Африке. Формация Бофорта (часть Кару Супергруппа, который охватывает большую часть Южной Африки), который богат окаменелостями позвоночных, в том числе терапсиды (рептилии-млекопитающие переходные формы ).[121] Было подсчитано, что это образование содержит 800 миллиардов окаменелостей позвоночных.[122] Палентологи задокументировали множество переходных форм и составили «удивительно исчерпывающий отчет о ключевых переходах в эволюции животных».[123] Проведя обзор палеонтологической литературы, можно было бы обнаружить, что существует «множество свидетельств того, как все основные группы животных связаны между собой, большая часть из которых представлена в виде превосходных переходных окаменелостей».[123]
Ограничения
Летопись окаменелостей является важным источником для ученых при отслеживании эволюционной истории организмов. Однако из-за ограничений, присущих описанию, между родственными группами видов нет мелких масштабов промежуточных форм. Отсутствие в летописи непрерывных окаменелостей является основным ограничением при отслеживании происхождения биологических групп. Когда переходные окаменелости обнаружены, что показывают промежуточные формы в том, что раньше было пробелом в знаниях, их часто в народе называют «недостающими звеньями».
Существует промежуток примерно в 100 миллионов лет между началом Кембрийский период и конец Ордовик период. Ранний кембрийский период был периодом, когда многочисленные окаменелости губки, книдарийцы (например, медуза ), иглокожие (например, эокриноиды ), моллюски (например, улитки ) и членистоногие (например, трилобиты ) найдены. Первое животное, обладавшее типичными чертами позвоночные, то Арандаспис, датируется более поздним ордовиком. Таким образом, мало окаменелостей промежуточного типа между беспозвоночные и позвоночные, хотя вероятные кандидаты включают Burgess Shale животное Pikaia gracilens,[124] и это Маотяньшаньские сланцы родные, Myllokunmingia, Юннанозоон, Haikouella lanceolata,[125] и Haikouichthys.[126]
Некоторые из причин неполной летописи окаменелостей:
- В общем, вероятность того, что организм окаменет, очень мала;
- Некоторые виды или группы с меньшей вероятностью станут ископаемыми, потому что они имеют мягкое тело;
- Некоторые виды или группы с меньшей вероятностью станут окаменелостями, потому что они живут (и умирают) в условиях, неблагоприятных для окаменения;
- Многие окаменелости были уничтожены в результате эрозии и тектонических движений;
- Большинство окаменелостей фрагментарны;
- Некоторое эволюционное изменение происходит в популяциях на границах экологического ареала вида, и, поскольку эти популяции, вероятно, малы, вероятность окаменения ниже (см. прерывистое равновесие );
- Точно так же, когда изменяются условия окружающей среды, популяция вида, вероятно, значительно сократится, так что любые эволюционные изменения, вызванные этими новыми условиями, с меньшей вероятностью останутся окаменевшими;
- Большинство окаменелостей несут информацию о внешней форме, но мало о том, как функционировал организм;
- Используя современные биоразнообразие в качестве ориентира это говорит о том, что раскопанные окаменелости представляют собой лишь небольшую часть большого числа видов организмов, которые жили в прошлом.
Конкретные примеры из палеонтологии
Эволюция лошади

Из-за почти полной летописи окаменелостей, обнаруженных в осадочных отложениях Северной Америки с раннего эоцен до настоящего времени лошадь представляет собой один из лучших примеров эволюционной истории (филогения ).
Эта эволюционная последовательность начинается с маленького животного по имени Гиракотерий (обычно называют Эогиппус), который жил в Северной Америке около 54 миллионов лет назад, затем распространился на Европа и Азия. Ископаемые останки Гиракотерий показать, что он отличался от современной лошади в трех важных отношениях: это было маленькое животное (размером с лиса ), легкая конструкция и приспособленность для бега; конечности были короткими и тонкими, а лапы удлиненными, так что пальцы были почти вертикальными, с четырьмя пальцами в передние конечности и три цифры в Задние конечности; и резцы были маленькими, коренные зубы с низкими коронками с закругленными куспиды покрыто эмаль.[127]
Вероятный ход развития лошадей от Гиракотерий к Equus (современная лошадь) задействовано не менее 12 роды и несколько сотен виды. Основные тенденции, наблюдаемые в развитии лошади к изменяющимся условиям окружающей среды, можно резюмировать следующим образом:
- Увеличение размера (с 0,4 м до 1,5 м - с 15 до 60 дюймов);
- Удлинение конечностей и стоп;
- Уменьшение боковых цифр;
- Увеличение длины и толщины третьего пальца;
- Увеличение ширины резцы;
- Замена премоляры от коренные зубы; и
- Увеличивается длина зуба, высота коронки коренных зубов.
Ископаемые растения, обнаруженные в разных пластах, показывают, что болотистый, лесная страна, в которой Гиракотерий жилые стали постепенно суше. Выживание теперь зависело от того, чтобы голова находилась в приподнятом положении, чтобы хорошо видеть окружающую сельскую местность, и от высокой скорости для побега. хищники, отсюда увеличение размера и замена растопыренной ступни копытной. Более сухая и твердая почва сделает исходную растопыренную ногу ненужной для поддержки. Изменения в зубах можно объяснить, если предположить, что диета изменилась с мягкой. растительность к трава. Доминирующий род из каждого геологического периода был выбран (см. Рисунок 3e), чтобы показать медленное изменение линии происхождения лошади от ее предковой к ее современной форме.[128]
Переход от рыб к амфибиям
До 2004 года палеонтологи находили окаменелости земноводных с шеей, ушами и четырьмя ногами в породах возрастом не более 365 миллионов лет. В скалах возрастом более 385 миллионов лет они могли найти только рыбу без этих характеристик амфибий. Эволюционная теория предсказывала, что, поскольку земноводные произошли от рыб, промежуточная форма должна быть найдена в породах, датируемых 365–385 миллионами лет назад. Такая промежуточная форма должна обладать многими рыбоподобными характеристиками, сохранившимися с 385 миллионов лет назад или более, но также иметь многие характеристики амфибий. В 2004 году экспедиция на острова в канадской Арктике в поисках этой ископаемой формы в породах возрастом 375 миллионов лет обнаружила окаменелости Тиктаалик.[129] Однако несколько лет спустя ученые Польша найдены доказательства окаменелости четвероногий следы, предшествующие Тиктаалик.[130]
Данные биогеографии
Данные о наличии или отсутствии видов на различных континенты и острова (биогеография ) может предоставить доказательства общего происхождения и пролить свет на закономерности видообразование.
Континентальное распределение
Все организмы в большей или меньшей степени адаптированы к окружающей среде. Если абиотические и биотические факторы в пределах место обитания способны поддерживать определенный вид в одной географической области, тогда можно предположить, что тот же вид может быть найден в аналогичной среде обитания в аналогичной географической области, например в Африка и Южная Америка. Это не вариант. Виды растений и животных распространены по всему миру прерывисто:
- Африка имеет Обезьяны Старого Света, обезьяны, слоны, леопарды, жирафы, и птицы-носороги.
- Южная Америка Обезьяны Нового Света, пумы, ягуары, ленивцы, ламы, и туканы.
- В пустынях Северной и Южной Америки есть кактусы, но в пустынях Африки, Азии и Австралии сочный (Помимо Rhipsalis baccifera )[131] которые являются родными молочай напоминающие кактусы, но очень разные.
Еще большие различия можно найти, если Австралия учитывается, хотя занимает ту же широта столько же Южной Америки и Африки. Сумчатые любить кенгуру, бандикуты, и Quolls составляют около половины аборигенных видов млекопитающих Австралии.[132] Напротив, сумчатые в настоящее время полностью отсутствуют в Африке и составляют меньшую часть фауны млекопитающих Южной Америки, где опоссумы, опоссумы землеройки, а Монито дель Монте происходят. Единственные ныне живущие представители первобытных млекопитающих-откладывающих яйца (монотремы ) являются ехидны и утконос. Короткоклювая ехидна (Акулеатус тахиглосс ) и его подвиды населяют Австралию, Тасмания, Новая Гвинея, и Остров Кенгуру а длинноклювая ехидна (Zaglossus bruijni ) живет только в Новой Гвинее. Утконос обитает в водах восточной Австралии. Их познакомили с Тасманией, King Island, и остров Кенгуру. Эти Monotremes полностью отсутствуют в остальном мире.[133] С другой стороны, в Австралии отсутствуют многие группы плацентарный млекопитающие, которые распространены на других континентах (плотоядные, парнокопытные, землеройки, белки, зайцеобразные ), хотя есть и коренные летучие мыши и мышиный грызуны; многие другие плаценты, такие как кролики и лисы, были завезены туда людьми.
Другие примеры распространения животных включают медведи, расположенный на всех континентах, за исключением Африки, Австралии и Антарктиды, а белый медведь - только за Полярным кругом и прилегающими массивами суши.[134] Пингвины встречаются только вокруг Южного полюса, несмотря на аналогичные погодные условия на Северном полюсе. Семьи сирены распределены по земным водам, где ламантины расположены только в водах Западной Африки, Северной Южной Америки и Вест-Индии, в то время как родственная семья, дюгони, расположены только в Океанический воды к северу от Австралии и побережья, окружающие Индийский океан. Теперь вымершие Стеллерова морская корова проживал в Берингово море.[135]
Такие же окаменелости были обнаружены в районах, которые, как известно, прилегают друг к другу в прошлом, но которые в результате Континентальный дрифт, сейчас находятся в самых разных географических точках. Например, окаменелости тех же типов древних земноводных, членистоногих и папоротников обнаружены в Южной Америке, Африке, Индии, Австралии и Антарктиде, которые можно датировать Палеозой Эра, когда эти регионы были объединены в единый массив, называемый Гондвана.[136] Иногда потомки этих организмов могут быть идентифицированы и обнаруживают безошибочное сходство друг с другом, даже несмотря на то, что теперь они населяют очень разные регионы и климатические условия.
Биогеография острова

Типы видов, обитающих на островах
Свидетельства из островная биогеография сыграл важную историческую роль в развитии эволюционная биология. Для целей биогеография, острова делятся на два класса. Континентальные острова подобны островам Великобритания, и Япония которые когда-то были частью континента. Океанические острова, такие как Гавайские острова, то Галапагосские острова и Св. Елены, с другой стороны, это острова, которые образовались в океане и никогда не были частью какого-либо континента. Океанические острова имеют несбалансированное распределение местных растений и животных, что отличает их от биоты встречается на континентах или континентальных островах. На океанских островах нет местных наземных млекопитающих (иногда встречаются летучие мыши и тюлени), амфибий или пресноводных рыб. В некоторых случаях у них есть наземные рептилии (например, игуаны и гигантские черепахи с Галапагосских островов), но часто (например, на Гавайях) их нет. И это несмотря на то, что когда такие виды, как крысы, козы, свиньи, кошки, мыши и тростниковые жабы, завезены на такие острова людьми, на которых они часто процветают. Начиная с Чарльз Дарвин Многие ученые проводили эксперименты и наблюдения, которые показали, что виды животных и растений, обнаруженные и не найденные на таких островах, согласуются с теорией о том, что эти острова были случайно колонизированы растениями и животными, которые смогли добраться до них. Такая случайная колонизация может происходить по воздуху, например, семена растений, переносимые перелетными птицами, или летучие мыши и насекомые, уносимые ветром над морем, или плывущие по морю с континента или другого острова (например, некоторыми видами семена растений, такие как кокосы, которые могут выдержать погружение в соленую воду), и рептилии, которые могут выжить в течение длительного времени на плотах с растительностью, уносимых в море штормами.[137]
Эндемизм
Многие виды, обитающие на отдаленных островах, являются эндемичный к определенному острову или группе островов, то есть они не встречаются больше нигде на Земле. Примеры видов, эндемичных для островов, включают множество нелетающих птиц Новая Зеландия, лемуры из Мадагаскар, то Дракон Комодо из Комодо,[138] Кровавое дерево дракона Сокотра,[139] Туатара Новой Зеландии,[140][141] и другие. Однако многие такие эндемичные виды связаны с видами, обитающими на других близлежащих островах или континентах; хорошо известным примером является родство животных, обитающих на Галапагосских островах, с животными, обитающими в Южной Америке.[137] Все эти факты, типы растений и животных, встречающиеся на океанических островах, большое количество эндемичных видов, обитающих на океанических островах, и связь таких видов с теми, что живут на ближайших континентах, легче всего объяснить, если бы острова были колонизированы. видами с близлежащих континентов, которые превратились в эндемичных видов, которые сейчас встречаются здесь.[137]
Другие типы эндемизма не обязательно должны включать острова в строгом смысле слова. Острова могут означать изолированные озера или отдаленные и изолированные районы. Примеры этого включают высокогорье Эфиопия, озеро Байкал, финбос из Южная Африка, леса Новая Каледония, и другие. Примеры эндемичных организмов, обитающих в изолированных районах, включают кагу Новой Каледонии,[142] облачные крысы из Тропические сосновые леса Лусона из Филиппины,[143][144] дерево буджум (Fouquieria columnaris ) из Полуостров Нижняя Калифорния,[145] то Байкальский тюлень[146] и омуль озера Байкал.
Адаптивные излучения
Океанические острова часто населены скоплениями близкородственных видов, которые заполняют множество экологические ниши, часто ниши, которые заполняются самыми разными видами на континентах. Такие скопления, как зяблики Галапагосских островов, Гавайские медоносы, представители семейства подсолнечных на Архипелаг Хуана Фернандеса а лесных долгоносиков на острове Св. Елены называют адаптивные излучения потому что они лучше всего объясняются тем, что один вид колонизирует остров (или группу островов), а затем диверсифицируется, чтобы заполнить имеющиеся экологические ниши. Такие излучения могут быть впечатляющими; 800 видов семейства плодовых мух Дрозофила, почти половина мирового населения, являются эндемиками Гавайских островов. Другой показательный пример с Гавайев - это альянс серебряных мечей, который представляет собой группу из тридцати видов, обитающих только на этих островах. Члены варьируются от серебряные мечи которые эффектно цветут на высоких вулканических склонах, деревьях, кустарниках, виноградных лозах и циновках, которые встречаются на разных высотах от вершины горы до уровня моря, а также в гавайских средах обитания, которые варьируются от пустынь до тропических лесов. Их ближайшие родственники за пределами Гавайев, согласно молекулярным исследованиям, являются тарвиды найден на западном побережье Северной Америки. Эти тарвики имеют липкие семена, которые облегчают распространение перелетными птицами.[147] Кроме того, почти все виды на острове могут быть скрещены, а гибриды часто плодовиты.[57] и они были экспериментально гибридизированы с двумя видами тарвика западного побережья.[148] Континентальные острова имеют менее выраженную биоту, но те, которые давно отделены от любого континента, также имеют эндемичные виды и адаптивную радиацию, такие как 75 лемур виды Мадагаскар, и одиннадцать вымерших моа виды Новая Зеландия.[137][149]
Кольцевые виды

Кольцевой вид - это связанная серия популяций, каждая из которых может скрещиваться со своими соседями, по крайней мере, с двумя «конечными» популяциями, которые слишком отдаленно связаны с межпородными скрещиваниями, хотя и имеют потенциал для скрещивания. поток генов между всеми народами.[150] Кольцевые виды представляют видообразование и были цитированы как свидетельство эволюции. Они иллюстрируют, что происходит с течением времени, когда популяции генетически расходятся, особенно потому, что они представляют, в живых популяциях, что обычно происходит с течением времени между популяциями давно умерших предков и живыми популяциями, в которых промежуточные звенья стали вымерший. Ричард Докинз говорит, что виды колец «только показывают нам в пространственном измерении то, что всегда должно происходить во временном измерении».[151]
Конкретные примеры из биогеографии
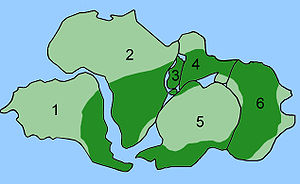

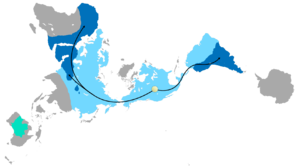
Распространение Глоссоптерис
Сочетание дрейфа континентов и эволюции иногда можно использовать для предсказания того, что будет найдено в летописи окаменелостей. Глоссоптерис вымерший вид семенной папоротник растения из Пермский период. Глоссоптерис появляется в летописи окаменелостей в начале пермского периода на древнем континенте Гондвана.[152] Дрейф континентов объясняет текущую биогеографию дерева. Сегодняшний день Глоссоптерис окаменелости найдены в пермских слоях на юго-востоке Южной Америки, юго-восточной Африке, на всем Мадагаскаре, на севере Индии, во всей Австралии, во всей Новой Зеландии и разбросаны по южным и северным краям Антарктиды. В пермский период эти континенты были соединены в Гондвану (см. Рисунок 4c) в соответствии с магнитной полосой, другими распределениями окаменелостей и ледниковыми царапинами, указывающими в сторону от умеренного климата Южного полюса во время пермского периода.[153][154]
Распределение метатериана
История метатерии (клады, содержащие сумчатые и их вымершие, примитивные предки) представляет собой пример того, как эволюционная теория и движение континентов могут быть объединены, чтобы делать прогнозы в отношении стратиграфии и распределения окаменелостей. В древнейшие окаменелости метатерии находятся в настоящее время Китай.[155] Метатерии распространились на запад в современную Северную Америку (все еще присоединенную к Евразии), а затем в Южную Америку, которая была связана с Северной Америкой примерно до 65 млн лет назад. Сумчатые достигли Австралии через Антарктиду примерно через 50 миллионов лет назад, вскоре после отделения Австралии, что позволяет предположить, что произошло единственное событие рассеяния только одного вида.[156] Эволюционная теория предполагает, что австралийские сумчатые произошли от более старых, обитающих в Америке. Геологические данные свидетельствуют о том, что между 30 и 40 миллионами лет назад Южная Америка и Австралия все еще были частью суперконтинента Южного полушария. Гондвана и что они были связаны сушей, которая сейчас является частью Антарктиды. Таким образом, комбинируя модели, ученые могли предсказать, что сумчатые животные мигрировали из современной Южной Америки через Антарктиду, а затем в современную Австралию между 40 и 30 миллионами лет назад. Первый сумчатое животное окаменелость вымершего семейства Полидолопиды был найден на Остров Сеймур на Антарктический полуостров в 1982 г.[157] Впоследствии были обнаружены другие окаменелости, в том числе представители сумчатых отрядов. Дидельфиморфия (опоссум) и Микробиотерия,[158] а также копытные и член загадочного вымершего ордена Гондванатерия возможно Sudamerica ameghinoi.[159][160][161]
Миграция, изоляция и распространение верблюда
История создания верблюд представляет собой пример того, как ископаемые остатки можно использовать для реконструкции миграции и последующей эволюции. Летопись окаменелостей указывает на то, что эволюция верблюды зародились в Северной Америке (см. рисунок 4e), откуда шесть миллионов лет назад они мигрировали через Берингов пролив в Азию, а затем в Африку, а 3,5 миллиона лет назад через Панамский перешеек в Южную Америку. Будучи изолированными, они развивались по своему собственному пути, давая начало Двугорбый верблюд и верблюд в Азии и Африке и лама и ее родственники В Южной Америке. Затем верблюды вымерли в Северной Америке в конце последнего Ледниковый период.[162]
Доказательства отбора
Примеры свидетельств эволюции часто происходят из непосредственного наблюдения естественный отбор в полевых условиях и в лаборатории. Этот раздел уникален тем, что предоставляет более узкий контекст, касающийся процесса отбора. Все примеры, приведенные до этого, описывали доказательства того, что эволюция произошла, но не предоставляли основной основной механизм: естественный отбор. В этом разделе явно представлены доказательства того, что естественный отбор имеет место, был воспроизведен искусственно и может быть воспроизведен в лабораторных экспериментах.
Ученые наблюдали и задокументировали множество событий, в которых действует естественный отбор. Наиболее известные примеры - устойчивость к антибиотикам в области медицины, а также более известные лабораторные эксперименты, документирующие возникновение эволюции. Естественный отбор равносилен общему происхождению в том смысле, что длительное возникновение и давление отбора могут привести к разнообразию жизни на Земле, которое существует сегодня. Все адаптации - задокументированные и недокументированные изменения - вызваны естественным отбором (и некоторыми другими второстепенными процессами). Хорошо известно, что «... естественный отбор - повсеместная часть видообразования ...»,[163] и является основной движущей силой видообразования;[164] поэтому следующие примеры естественного отбора и видообразования часто взаимозависимы или соответствуют друг другу. Приведенные ниже примеры - это лишь небольшая часть реальных экспериментов и наблюдений.
Искусственный отбор и экспериментальная эволюция

Искусственный отбор демонстрирует разнообразие, которое может существовать среди организмов, имеющих относительно недавнего общего предка. При искусственном отборе один вид селективно разводится в каждом поколении, позволяя воспроизводиться только тем организмам, которые демонстрируют желаемые характеристики. Эти характеристики становятся все более развитыми в следующих поколениях. Искусственный отбор был успешным задолго до того, как наука открыла генетическую основу. Примеры искусственного отбора включают: разведение собак, генетически модифицированные продукты, цветоводство и выращивание таких продуктов, как дикая капуста,[165] и другие.
Экспериментальная эволюция использует контролируемые эксперименты для проверки гипотез и теорий эволюции. В одном из ранних примеров Уильям Даллинджер Незадолго до 1880 года поставил эксперимент, подвергая микробы воздействию тепла с целью вызвать адаптивные изменения. Его эксперимент длился около семи лет, и его опубликованные результаты были высоко оценены, но он не возобновил эксперимент после того, как аппарат вышел из строя.[166]
Крупномасштабный пример экспериментальной эволюции: Ричард Ленски с эксперимент нескольких поколений с кишечная палочка. Ленский заметил, что некоторые штаммы Кишечная палочка развил новую сложную способность, способность метаболизировать цитрат через десятки тысяч поколений.[167][168] Биолог-эволюционист Джерри Койн выступил с критикой креационизма, сказав: «Больше всего мне нравится то, что вы можете получить эти сложные черты развивается в результате сочетания маловероятных событий. Креационисты говорят, что этого не может быть ».[167] В дополнение к метаболическим изменениям было обнаружено, что различные популяции бактерий расходились как по морфологии (общий размер клетки), так и по приспособленности (которая измерялась в конкуренции с предками).[169] В Кишечная палочка долгосрочный эволюционный эксперимент, начатый в 1988 году, все еще продолжается и показал адаптации, включая эволюцию штамма Кишечная палочка который мог расти на лимонной кислоте в питательной среде - черта, отсутствующая у всех других известных форм Кишечная палочка, включая начальную деформацию.
Беспозвоночные
Историческая толерантность к свинцу в Дафния
Изучение видов Дафния и загрязнение свинцом в 20 веке предсказало, что увеличение загрязнения свинцом приведет к строгому выбору толерантности к свинцу. Исследователи смогли использовать «экологию воскрешения», выведенную десятилетиями давности. Дафния яйца из тех времен, когда озера были сильно загрязнены свинцом. Птенцов в исследовании сравнивали с нынешними. Дафния, и продемонстрировали «резкие различия в приспособленности между старыми и современными фенотипами при столкновении с широко распространенным историческим стрессором окружающей среды». По сути, современный Дафния были неспособны противостоять или переносить высокие уровни свинца (это связано с огромным сокращением загрязнения свинцом в озерах 21 века). Однако старые птенцы были способны переносить высокое загрязнение свинцом. Авторы пришли к выводу, что «используя методы экологии воскрешения, мы смогли показать явные фенотипические изменения за десятилетия ...».[170]
Моль перечная
Классическим примером было изменение фенотипа, адаптация цвета от светлого к темному, в моль перечная, из-за загрязнения Индустриальная революция в Англии.
Микробы
Устойчивость к противомикробным препаратам
Развитие и распространение антибиотик -устойчивый бактерии свидетельствует о процессе эволюции видов. Таким образом, появление ванкомицин -устойчивый Золотистый стафилококк, и опасность, которую он представляет для пациентов больниц, является прямым результатом эволюции путем естественного отбора. Подъем Шигелла штаммы, устойчивые к синтетическим антибиотикам класса сульфаниламиды также демонстрирует генерацию новой информации как эволюционный процесс.[171] Точно так же появление ДДТ сопротивление в различных формах Анофелес комаров, а появление миксоматоз устойчивость в популяциях размножающихся кроликов в Австралии, оба являются свидетельством существования эволюции в ситуациях эволюционного давление отбора у видов, у которых поколения происходят быстро.
Устойчивость развивается у всех классов микробов: в том числе у грибов (противогрибковый сопротивление), вирусы (противовирусное средство сопротивление), простейшие (противопротозойный сопротивление), и бактерии (антибиотик сопротивление). Этого следовало ожидать, если учесть, что вся жизнь проявляет универсальный генетический код и, следовательно, подвержена процессу эволюции через свои различные механизмы.
Бактерии, поедающие нейлон
Еще один пример адаптации организмов к антропогенным условиям: Бактерии, поедающие нейлон: штамм Флавобактерии которые способны переваривать определенные побочные продукты нейлон 6 изготовление. Существует научный консенсус в отношении того, что способность синтезировать нейлоназу, скорее всего, возникла в результате одноступенчатой мутации, которая выжила, потому что улучшила приспособленность бактерий, обладающих мутацией. Это рассматривается как хороший пример эволюции через мутации и естественный отбор, которые наблюдались по мере их возникновения и не могли произойти до тех пор, пока люди не начали производство нейлона.[172][173][174][175]
Растения и грибы
Обезьянье излучение
Оба подвида Mimulus aurantiacus puniceus (с красными цветками) и Mimulus aurantiacus australis (желтоцветковые) обезьяньих цветов изолированы из-за предпочтений их опылителей колибри и ястреба. Излучение М. aurantiacus подвиды преимущественно желтого цвета; однако оба М. а. ssp. Puniceus и М. а. ssp. Flemingii красные. Филогенетический анализ предполагает два независимых происхождения красных цветов, возникших из-за СНГ-регуляторные мутации в гене MaMyb2 что присутствует во всех М. aurantiacus подвид. Дальнейшие исследования показали, что не произошло двух независимых мутаций, а произошла одна MaMyb2 аллель был перенесен путем интрогрессивной гибридизации.[176] Это исследование представляет собой пример пересечения исследований в различных дисциплинах. Выделение генов и СНГ-регулирующие функции; филогенетический анализ; географическое положение и предпочтения опылителей; а гибридизация и видообразование видов - это лишь некоторые из областей, в которых можно получить данные, подтверждающие наличие эволюции.
Радиотрофные грибы
Как и в случае с треской, антропогенное загрязнение может принимать различные формы. Радиотрофные грибы является прекрасным примером естественного отбора, происходящего после химической аварии. Радиотрофные грибы используют пигмент. меланин преобразовать гамма-излучение в химическую энергию для роста[177][178] и были впервые обнаружены в 2007 году как черные формы растет внутри и вокруг Чернобыльская АЭС.[177] Исследования в Медицинский колледж Альберта Эйнштейна показали, что три меланинсодержащих гриба, Cladosporium sphaerospermum, Wangiella dermatitidis, и Криптококк neoformans, увеличилось в биомасса и накопил ацетат быстрее в среде, в которой радиация уровень был в 500 раз выше, чем в обычной среде.
Позвоночные
Гуппи
Во время учебы гуппи (Poecilia reticulata) в Тринидад биолог Джон Эндлер обнаружил, что селекция воздействует на популяции рыб. Чтобы исключить альтернативные возможности, Эндлер поставил строго контролируемый эксперимент, чтобы имитировать естественную среду обитания, построив десять прудов в лабораторной теплице в Университет Принстона. В каждом пруду был гравий, точно такой же, как в естественных прудах. После отлова случайной выборки гуппи из прудов в Тринидад, он вырастил и смешал их, чтобы создать похожие генетически разнообразные популяции, и измерил каждую рыбу (длину пятна, высоту пятна, площадь пятна, относительную длину пятна, относительную высоту пятна, общую площадь пятна и стандартную длину тела). Для эксперимента он добавил Crenicichla альта (P. reticulata 'главный хищник) в четырех прудах, Ривулус Hartii (рыба, не являющаяся хищником) в четырех прудах, а оставшиеся два пруда остались пустыми, оставив только гуппи.После 10 поколений были проведены сравнения популяций гуппи в каждом пруду, и измерения были проведены снова. Эндлер обнаружил, что в популяциях резко различались цветовые узоры в контрольной группе и пуле без хищников, а также тусклые цветовые узоры в пуле хищников. Давление хищников привело к выделению на фоне гравия.[179]

Параллельно с этим во время этого эксперимента Эндлер провел полевой эксперимент на Тринидаде, где он поймал гуппи из прудов, где у них были хищники, и переселил их в пруды выше по течению, где хищники не жили. Спустя 15 поколений Эндлер обнаружил, что перемещенные гуппи развили драматические и красочные узоры. По сути, оба эксперимента показали конвергенцию из-за схожего давления отбора (т. Е. Отбор хищников против контрастных цветовых узоров и половой отбор для контрастных цветовых узоров).[179]
В более позднем исследовании Дэвида Резника полевую популяцию изучили 11 лет спустя после того, как Эндлер перевел гуппи в высокие реки. Исследование показало, что популяции эволюционировали по-разному: яркие цветовые узоры, позднее созревание, большие размеры, меньшие размеры помета и более крупное потомство в помете.[180] Дальнейшие исследования P. reticulata и их хищники в ручьях Тринидада указали, что различные способы отбора посредством хищничества не только изменили цветовые узоры, размеры и поведение гуппи, но и их истории жизни и образцы истории жизни.[181]
Люди
Естественный отбор наблюдается в современных популяциях людей, и недавние результаты показывают, что население, подверженное риску тяжелого изнурительного заболевания Куру имеет значительную избыточную представленность иммунного варианта прионный белок ген G127V в сравнении с неиммунными аллелями. Ученые постулируют одну из причин быстрого отбора этого генетический вариант - летальность заболевания у лиц без иммунитета.[182][183] Другие известные эволюционные тенденции в других популяциях включают удлинение репродуктивного периода, снижение уровня холестерина, глюкозы в крови и артериального давления.[184]
Хорошо известным примером селекции, происходящей в человеческих популяциях, является толерантность к лактозе. Непереносимость лактозы это неспособность метаболизировать лактоза, из-за отсутствия необходимого фермента лактаза в пищеварительной системе. Нормальное состояние млекопитающих заключается в том, что молодые особи вида испытывают снижение выработки лактазы в конце периода кормления. отлучение от груди период (период времени, зависящий от вида). У людей в обществах, не потребляющих молочные продукты, выработка лактазы обычно падает примерно на 90% в течение первых четырех лет жизни, хотя точное падение со временем сильно варьируется.[185] Сохранение активности лактазы у взрослых связано с двумя факторами: полиморфизмы: C / T 13910 и G / A 22018, расположенные в MCM6 ген.[186] Это различие генов устраняет остановку производства лактазы, позволяя членам этих групп населения без проблем продолжать потребление сырого молока и других свежих и ферментированных молочных продуктов на протяжении всей жизни. Это, кажется, эволюционно недавнее (около 10 000 лет назад [и 7 500 лет назад в Европе][187]) адаптация к потреблению молочных продуктов,[188] и произошел независимо как в Северной Европе, так и в Восточной Африке в популяциях с исторически скотоводческим образом жизни.[189][190]
Итальянские настенные ящерицы
В 1971 г. десять взрослых особей Podarcis sicula (итальянская стенная ящерица) были перевезены из хорватский от острова Под Копиште до острова Под Мрчару (около 3,5 км к востоку). Оба острова лежат в Адриатическое море около Ластово, где ящерицы основали новый узкое место Население.[191][192] Два острова имеют одинаковый размер, высота, микроклимат, и вообще отсутствие наземных хищников[192] и P. sicula расширилась на десятилетия без вмешательства человека, даже превзойдя (ныне вымершие на местном уровне[191]) Podarcis melisellensis Население.[193]
В 1990-х годах ученые вернулись в Под Мрчару и обнаружили, что ящерицы там сильно отличаются от ящериц Копиште. В то время как митохондриальная ДНК анализы подтвердили, что P. sicula в настоящее время на Мрчару генетически очень похожи на исходную популяцию Копиште,[191] новое население Мрчару P. sicula имели больший средний размер, более короткие задние конечности, более низкую максимальную скорость спринта и измененную реакцию на симулированные нападения хищников по сравнению с исходной популяцией Копиште.[192] Эти изменения были приписаны «ослаблению интенсивности хищничества» и большей защите от растительности на Мрчару.[192]
В 2008 году дальнейший анализ показал, что популяция мрчару P. sicula имеют существенно отличающуюся морфологию головы (более длинную, широкую и более высокую голову) и повышенную силу укуса по сравнению с исходной популяцией Копиште.[191] Это изменение формы головы соответствовало изменению диеты: Копиште P. sicula в первую очередь насекомоядный, но те, кто на Мрчару, едят значительно больше растительной пищи.[191] Изменения в стиле кормодобывания могли способствовать увеличению плотности популяции и снижению территориального поведения популяции мрчару.[191]
Еще одно различие, обнаруженное между двумя популяциями, заключалось в обнаружении у ящериц Мрчару слепые клапаны, которые замедляют прохождение пищи и предоставляют бродильные камеры, позволяя комменсальный микроорганизмы для преобразования целлюлоза питательным веществам, усваиваемым ящерицами.[191] Кроме того, исследователи обнаружили, что нематоды были обычны в кишках ящериц Мрчару, но отсутствовали в Копиште. P. sicula, у которых нет слепых клапанов.[191] Клапаны слепой кишки, которые встречаются менее чем у 1 процента всех известных видов чешуйчатых рептилий,[191] были описаны как «адаптивная новинка, совершенно новая особенность, отсутствующая у предков и недавно появившаяся у этих ящериц».[194]
Устойчивость к ПАУ у киллиф
Аналогичное исследование было проведено и в отношении полициклические ароматические углеводороды (ПАУ), загрязняющие воды реки Элизабет в Портсмуте, штат Вирджиния. Это химическое вещество является продуктом креозот, разновидность дегтя. Атлантический киллфиш (Fundulus гетероклит) развил устойчивость к ПАУ с участием гена AHR (того же гена, что и у котов). Это конкретное исследование было сфокусировано на устойчивости к «острой токсичности и сердечному тератогенезу», вызываемой ПАУ, которые мутировали у котов в реке Гудзон.[195]
Устойчивость к ПХБ у трески
Примером прямого наблюдения за модификацией генов из-за давления отбора является устойчивость трески к ПХБ. После General Electric сброшен полихлорированные бифенилы (Печатных плат) в река Гудзон с 1947 по 1976 год, Tomcods (Microgadus tomcod), живущие в реке, развили повышенную устойчивость к токсическим эффектам соединения.[196] Устойчивость к токсинам обусловлена изменением кодирующей части конкретного гена. Генетические образцы были взяты у трески из 8 различных рек в регионе Новой Англии: реки Св. Лаврентия, реки Мирамичи, реки Маргари, реки Сквамскотт, реки Ниантик, реки Шиннекок Бейсик, реки Гудзон и реки Хакенсак. Генетический анализ показал, что в популяции котиков четырех самых южных рек ген AHR2 (рецептор арильных углеводородов 2) присутствует как аллель с разницей в две аминокислотные делеции.[197] Эта делеция придавала устойчивость к ПХБ у видов рыб и была обнаружена у 99% котов реки Гудзон, 92% в реке Хакенсак, 6% в реке Ниантик и 5% в заливе Шиннекок.[197] Эта картина вдоль отобранных водоемов предполагает прямую корреляцию селективных давлений, ведущих к эволюции устойчивости к ПХБ у атлантических рыб.[197]
Городская дикая природа
Городская дикая природа представляет собой широкий и легко наблюдаемый случай антропогенного давления отбора на дикая природа. С ростом среды обитания человека различные животные приспособились к выживанию в этих городских условиях. Эти типы окружающей среды могут оказывать давление отбора на организмы, часто приводя к новым приспособлениям. Например, сорняк Crepis sancta, найденный во Франции, имеет два типа семян: тяжелые и пушистые. Тяжелые семена приземляются рядом с родительским растением, а пушистые семена летят дальше по ветру. В городских условиях далеко летающие семена часто приземляются на неплодородный бетон. Примерно через 5–12 поколений сорняк эволюционирует и дает значительно более тяжелые семена, чем его сельские родственники.[198][199] Другие примеры городской дикой природы: голуби и виды ворон, адаптирующиеся к городской среде по всему миру; Африканские пингвины в Саймонов город; бабуины в Южная Африка; и различные насекомые, обитающие в человеческих жилищах. Были проведены исследования, которые выявили поразительные изменения в поведении животных (в частности, млекопитающих) и физическом размере мозга из-за их взаимодействия с окружающей средой, созданной человеком.[200][201]
Ящерицы Белых Песков

Животные, выставляющие экотональный вариации позволяют исследовать механизмы, поддерживающие дифференциацию населения. Обширная информация о естественном отборе, генотипических и фенотипических вариациях;[202][203] адаптация и экоморфология;[204] и социальная сигнализация[205] был получен в результате исследований трех видов ящериц, обитающих в пустыне Белые пески в Нью-Мексико. Holbrookia maculata, Aspidoscelis inornatus, и Sceloporus undulatus демонстрируют экотональные популяции, которые соответствуют как темным почвам, так и белым пескам в регионе. Исследования, проведенные на этих видах, обнаружили значительные фенотипические и генотипические различия между темными и светлыми популяциями из-за сильного давления отбора. Например, H. maculata демонстрирует самое сильное фенотипическое различие (лучше всего соответствует субстрату) светлой популяции, что соответствует наименьшему количеству потока генов между популяциями и наибольшим генетическим различиям по сравнению с двумя другими видами ящериц.[202]
Нью-Мексико белый песок являются недавним геологическим образованием (приблизительно 6000 лет[205] возможно до 2000 лет[202]). Это недавнее происхождение этих гипсовых песчаных дюн предполагает, что виды, демонстрирующие вариации более светлого цвета, эволюционировали за относительно короткий период времени. Было обнаружено, что три ранее упомянутых вида ящериц демонстрируют различную социальную сигнальную окраску при сосуществовании со своими экотональными вариантами.[205] Мало того, что три вида конвергентно развили свои более легкие варианты из-за давления отбора со стороны окружающей среды, они также развили экоморфологические различия: морфологию, поведение (в данном случае - поведение побега) и производительность (в данном случае скорость спринта). коллективно.[204] Работа Роше обнаружила удивительные результаты в побеге. H. maculata и S. undulatus. Когда темные морфы были помещены на белый песок, их испуганная реакция значительно уменьшилась. Этот результат может быть связан с различными факторами, связанными с температурой песка или остротой зрения; однако, независимо от причины, «… неспособность несоответствующих ящериц к спринту может привести к дезадаптации при столкновении с хищником».[204]
Свидетельства видообразования
Видообразование - это эволюционный процесс, в результате которого возникают новые биологические виды. Биологи изучают виды, используя различные теоретические основы того, что составляет вид (см. проблема вида и видовой комплекс ), и ведутся споры относительно разграничения.[206] Тем не менее, большая часть текущих исследований предполагает, что «... видообразование - это процесс зарождающейся генеалогической отличимости, а не разрыв, затрагивающий все гены одновременно»[207] а в аллопатрии (наиболее распространенной форме видообразования) «репродуктивная изоляция является побочным продуктом эволюционных изменений в изолированных популяциях и, таким образом, может считаться эволюционной случайностью».[208] В результате последнего происходит видообразование (аллопатрия); однако задокументировано множество различных агентов, которые часто определяются и классифицируются в различных формах (например, перипатрические, парапатрические, симпатрические, полиплоидизация, гибридизация и т. д.). Примеры видообразования наблюдались как в природе, так и в лаборатории. А.-Б. Флорин и А. Один отмечают, что «убедительные лабораторные доказательства аллопатрического видообразования отсутствуют ...»; однако, в отличие от лабораторных исследований (сфокусированных конкретно на моделях аллопатрического видообразования), «видообразование наиболее определенно имеет место; [и] огромное количество свидетельств природы делает неразумным утверждать иное».[209] Койн и Орр составили список из 19 лабораторных экспериментов на Дрозофила представляя примеры аллопатрического видообразования посредством дивергентного отбора, делая вывод, что «репродуктивная изоляция в аллопатрии может развиваться как побочный продукт дивергентного отбора».[210]
Исследования, подтверждающие видообразование, многочисленны. Биологи задокументировали многочисленные примеры видообразования в природе, причем в результате эволюции появилось гораздо больше видов, чем любой наблюдатель сочтет необходимым. Например, существует более 350 000 описанных видов жуков.[211] Примеры видообразования взяты из наблюдений островной биогеографии и процесса адаптивной радиации, оба объясненных ранее. Доказательства общего происхождения также можно найти в палеонтологических исследованиях видообразования в геологических пластах. Примеры, описанные ниже, представляют различные способы видообразования и служат убедительным доказательством общего происхождения. Важно признать, что не во всех исследованиях видообразования наблюдается прямое расхождение от «начала до конца». Это происходит из-за разграничения в исследованиях и неоднозначности определений, и иногда исследования приводят к историческим реконструкциям. В свете этого примеров предостаточно, и нижеследующее ни в коем случае не является исчерпывающим и составляет лишь небольшую часть наблюдаемых случаев. Еще раз обратите внимание на установленный факт, что «... естественный отбор - это повсеместная часть видообразования ...»,[163] и является основной движущей силой видообразования,[164] так; в дальнейшем примеры видообразования часто взаимозависимы и соответствуют отбору.
Окаменелости
При рассмотрении концепции того, что составляет вид, существуют ограничения в летописи окаменелостей. Палеонтологи в основном полагаются на другую основу: концепцию морфологического вида.[212] Из-за отсутствия такой информации, как репродуктивное поведение или генетический материал в окаменелостях, палеонтологи различают виды по их фенотипическим различиям.[212] Обширное исследование палеонтологической летописи привело к появлению множества теорий, касающихся видообразования (в контексте палеонтологии), причем многие исследования предполагают, что застой, пунктуация, и ветвление по линии распространены. В 1995 г. Д. Х. Эрвин и др. опубликовал большую работу -Новые подходы к видообразованию в летописи окаменелостей- который собрал 58 исследований видообразования окаменелостей (между 1972 и 1995 гг.), Обнаружив большинство примеров, указывающих на застой (включая анагенез или пунктуация) и 16 исследований, предполагающих видообразование.[212] Несмотря на то, что на первый взгляд стазис кажется преобладающим выводом, в этом конкретном метаисследовании было проведено более глубокое исследование и сделан вывод о том, что «... ни одна модель не кажется доминирующей ...» с «... преобладанием исследований, иллюстрирующих и то и другое застой и постепенность в истории одной линии ».[213] Многие из проведенных исследований используют донные отложения, которые могут предоставить значительный объем данных, касающихся планктонный микрофоссилий.[212] Последовательность окаменелостей в стратиграфии может использоваться для определения тенденций эволюции ископаемых организмов. Кроме того, случаи видообразования можно интерпретировать на основе данных, и были проведены многочисленные исследования, документирующие как морфологическую эволюцию, так и видообразование.
Глобороталия
Обширные исследования планктонных фораминифер Глобороталия truncatulinoides предоставил представление о палеобиогеографических и палеоэкологических исследованиях, а также о взаимосвязи между окружающей средой и эволюцией. В обширном исследовании палеобиогеографии G. truncatulinoides, исследователи нашли доказательства, указывающие на формирование нового вида (через структуру симпатрического видообразования). Керны, взятые из осадка, содержащего три вида G. crassaformis, G. tosaensis, и G. truncatulinoides обнаружил, что до 2,7 млн лет только G. crassaformis и G. tosaensis существовал. В это время произошло событие видообразования, при котором промежуточные формы существовали довольно долго. В конце концов G. tosaensis исчезает из записи (предполагая исчезновение), но существует как промежуточное звено между сохранившимися G. crassaformis и G. truncatulinoides. Эта запись окаменелостей также соответствовала уже существующей филогении, построенной по морфологическим признакам трех видов.[214] См. Рисунок 6а.
Радиолярии
В большом исследовании пяти видов радиолярии (Calocycletta caepa, Птерокан призматический, Псевдокулезная вема, Eucyrtidium calvertense, и Eucyrtidium matuyamai ), исследователи задокументировали значительные эволюционные изменения в каждой линии. Наряду с этим, тенденции с близкородственными видами E. calvertense и Э. матуямаи показал, что около 1,9 млн лет назад E. calvertense вторглись в новый регион Тихого океана, изолировавшись от основного населения. Стратиграфия этого вида ясно показывает, что эта изолированная популяция превратилась в Э. Матуямай. Затем он заново овладел областью все еще существующих и статичных E. calvertense популяции, в результате чего произошло внезапное уменьшение размеров тела. В конце концов захватчик Э. матуямаи исчезли из слоя (предположительно из-за исчезновения), что совпало с отказом от уменьшения размера E. calvertense Население. С этого момента изменение размера стало постоянным. Авторы предлагают смещение персонажа.[215][216]
Ризосоления
Исследователи провели измерения на 5000 Ризосоления (планктонный диатомовые водоросли ) образцы из восьми осадочных кернов Тихого океана. Возраст образцов керна составлял два миллиона лет, и они были хронологизированы с использованием измерений изменения направления магнитного поля в осадках. Все образцы керна показали аналогичную картину дивергенции: с одной линией (R. bergonii), происходящие до 3.1 Mya и двух морфологически различных линий (дочерние виды: R. praebergonii) появляется после. Параметры, использованные для измерения образцов, были одинаковыми для каждого керна.[217] Дополнительное изучение дочерних видов R. praebergonii обнаружили, что после расхождения он вторгся в Индийский океан.[212][218]
Турбороталия
Недавнее исследование было проведено с участием планктонных фораминифер Турбороталия. Авторы извлекли "51 стратиграфически упорядоченную пробу с участка в океанографически стабильной тропической зоне. Северный тихоокеанский круговорот ". Двести отдельных видов были исследованы с использованием десяти конкретных морфологических признаков (размер, индекс сжатия, соотношение сторон камеры, расширение камеры, соотношение сторон апертуры, высота испытания, испытательное расширение, угол шлангокабеля, направление наматывания и количество камер в конечном оборот). Используя многомерные статистические методы кластеризации, исследование показало, что вид продолжал развиваться ненаправленно в пределах эоцен от 45 до 36 млн лет. Однако в период от 36 до примерно 34 млн лет в стратиграфических слоях были обнаружены два отдельных кластера со значительно определяющими характеристиками, отличающими друг друга от одного вида. Авторы пришли к выводу, что должно было произойти видообразование и что два новых вида были предками предшествующих видов.[219] Как и в большинстве случаев эволюционной биологии, этот пример представляет междисциплинарный характер области и необходимый сбор данных из различных областей (например, океанографии, палеонтологии) и интеграцию математического анализа (например, биометрии).
Позвоночные
Несмотря на ограничения, наложенные летописью окаменелостей, существуют свидетельства видообразования позвоночных. Были проведены исследования, подтверждающие аналогичные закономерности, наблюдаемые у морских беспозвоночных.[212] Например, обширные исследования, подтверждающие скорость морфологических изменений, эволюционные тенденции и образцы видообразования у мелких млекопитающих.[220] внес значительный вклад в научную литературу; еще раз, демонстрируя, что эволюция (и видообразование) происходила в прошлом и подтверждает общее происхождение.
Исследование четырех родов млекопитающих: Hyopsodus, Пеликодус, Гапломил (три из эоцена), и Плезиадапис (из палеоцена) обнаружил, что - благодаря большому количеству стратиграфических слоев и выборок - каждая группа демонстрировала «постепенную филетическую эволюцию, общее увеличение размера, итеративную эволюцию мелких видов и расхождение признаков после происхождения каждой новой линии».[221] Авторы этого исследования пришли к выводу, что видообразование можно различить. В другом исследовании, посвященном морфологическим тенденциям и темпам эволюции, было обнаружено, что европейские арвиколид грызун распространился на 52 различные линии за период времени в 5 миллионов лет, документируя примеры филетической постепенности, пунктуации и стазиса.[222]
Беспозвоночные
Drosophila melanogaster

Уильям Р. Райс и Джордж В. Солт обнаружили экспериментальные доказательства симпатрическое видообразование в обыкновенная плодовая муха. Они собрали популяцию Drosophila melanogaster от Дэвис, Калифорния и поместили куколок в лабиринт среды обитания. Новорожденным мухам пришлось исследовать лабиринт в поисках пищи. У мух было три варианта поиска пищи. Светлый и темный (фототаксис ), вверх и вниз (геотаксис ), и запах ацетальдегид и запах этанола (хемотаксис ) были три варианта. Это в конечном итоге разделило мух на 42 пространственно-временных среды обитания.
Затем они культивировали два сорта, которые выбрали противоположные места обитания. Один из штаммов появился рано и тут же взлетел вверх в темноте, привлеченный ацетальдегид. Другой штамм появился поздно и сразу же полетел вниз, привлеченный светом и этанолом. Затем куколок двух штаммов помещали вместе в лабиринт и давали возможность спариваться на месте кормления. Затем они были собраны. Избирательное наказание было применено к самкам мух, меняющих местообитания. Это означало, что ни один из их гаметы перейдет к следующему поколению. После 25 поколений этого теста спаривания он показал репродуктивную изоляцию между двумя штаммами. Они повторили эксперимент снова, не создавая штрафов за смену среды обитания, и результат был тем же; репродуктивная изоляция.[223][224][225]
Галловые осы
Исследование желчеобразующая оса виды Belonocnema Treatae обнаружили, что популяции, населяющие разные растения-хозяева (Quercus geminata и Q. virginiana ) выставлялись разного размера тела и желчь морфология наряду с сильным выражением сексуальной изоляции. Исследование выдвинуло гипотезу о том, что B. Treatae популяции, населяющие разные растения-хозяева, покажут доказательства расходящийся отбор способствуя видообразованию. Исследователи взяли образцы желчных ос и местонахождения дубов, измерили размер тела (правая рука большеберцовая кость каждой осы) и подсчитали номера желчных камер. Помимо измерений, они провели анализы спаривания и статистический анализ. Генетический анализ также проводился на двух мтДНК сайтов (416 пар оснований из цитохрома С и 593 пары оснований из цитохромоксидазы) для «контроля смешивающего воздействия времени с момента расхождения между аллопатрические популяции ".[226]
В дополнительном исследовании ученые изучили два вида галловых ос. B. Treatae и Disholcaspis quercusvirens и обнаружили сильные морфологические и поведенческие различия среди популяций, связанных с хозяином. Это исследование еще больше запутало предпосылки к видообразованию.[227]
Боярышник
Одним из примеров действующей эволюции является случай боярышника, Rhagoletis pomonella, также известная как муха яблочного личинки, которая, кажется, подвергается симпатрическое видообразование.[228] Разные популяции боярышника питаются разными фруктами. Особая популяция появилась в Северной Америке в XIX веке спустя некоторое время после яблоки, неместный вид. Эта популяция, питающаяся яблоками, обычно питается только яблоками, а не исторически предпочитаемыми фруктами. боярышник. Нынешняя популяция боярышника обычно не питается яблоками. Некоторые доказательства, такие как тот факт, что шесть из тринадцати аллозим loci различны, мухи боярышника созревают позже и созревают дольше, чем яблони; и то, что существует мало свидетельств межпородного скрещивания (исследователи задокументировали уровень гибридизации 4–6%), предполагает, что имеет место видообразование.[229][230][231][232][233]
Лондонский подземный комар
В Лондонский подземный комар это разновидность комар в роду Culex найдено в Лондонское метро. Он произошел от наземных видов Culex pipiens. Этот комар, хотя впервые был обнаружен в системе лондонского метрополитена, был обнаружен в подземных системах по всему миру. Предполагается, что он, возможно, адаптировался к искусственным подземным системам с прошлого века из местных наземных источников. Culex pipiens,[234] хотя более свежие данные свидетельствуют о том, что это южный сорт комаров, связанный с Culex pipiens адаптированный к теплым подземным пространствам северных городов.[235]
У этих двух видов очень разное поведение,[236] очень сложно спариваться,[234] и с другой частотой аллелей, соответствующей генетическому дрейфу во время событие учредителя.[237] Точнее, этот комар, Culex pipiens Mostus, размножается круглый год, не переносит холода и кусает крыс, мышей и людей, в отличие от наземных видов. Culex pipiens морозоустойчив, зимой впадает в спячку, кусает только птиц. При скрещивании двух разновидностей яйца оказались бесплодными, что свидетельствует о репродуктивной изоляции.[234][236]
Генетические данные показывают, что пристанище Форма москита в лондонском метро, по-видимому, имеет общее происхождение, а не популяция на каждой станции, относящаяся к ближайшей надземной популяции (т. е. pipiens форма). Рабочая гипотеза Бирна и Николса заключалась в том, что адаптация к подземной среде произошла локально в Лондоне только однажды. Эти сильно разделенные популяции отличаются очень незначительными генетическими различиями, которые позволяют предположить, что форма молестуса возникла: единственное различие мтДНК, характерное для подземных популяций десяти российских городов;[238] один фиксированный микроспутник разница в населении, охватывающая Европу, Японию, Австралию, Ближний Восток и острова Атлантики.[235]
Креветки и панамский перешеек
Существуют дебаты о том, когда Панамский перешеек закрыто. Большая часть доказательств поддерживает закрытие примерно от 2,7 до 3,5 млн лет, используя «... несколько линий доказательств и независимых опросов».[239] Однако недавнее исследование предполагает, что более ранний переходный мостик существовал от 13 до 15 млн лет назад.[240] Независимо от того, когда приблизится перешеек, биологи могут изучать виды, обитающие на берегах Тихого океана и Карибского моря, в рамках того, что было названо «одним из величайших естественных экспериментов в области эволюции».[239] Исследования креветок этого рода Алфей предоставили прямые свидетельства событий аллопатрического видообразования,[241] и внес свой вклад в литературу, касающуюся скорости молекулярной эволюции.[242] Филогенетические реконструкции с использованием «мультилокусных наборов данных и аналитических методов, основанных на коалесценции» подтверждают взаимосвязи видов в группе.[239] и методы молекулярных часов поддерживают разделение 15 пар Алфей вид между 3 и 15 миллионами лет назад.[242]
Растения
Ботаник Верн Грант стал пионером в области видообразования растений благодаря своим исследованиям и крупным публикациям по этой теме.[243] Как указывалось ранее, многие биологи полагаются на концепцию биологических видов, а некоторые современные исследователи используют концепция филогенетических видов. Споры ведутся о том, какие рамки следует применять в исследовании.[243] Несмотря на это, репродуктивная изоляция является основной ролью в процессе видообразования и широко изучается биологами в их соответствующих дисциплинах.
Как гибридизация, так и полиплоидия также являются основными факторами видообразования растений.[244] С появлением молекулярных маркеров «гибридизация [происходит] значительно чаще, чем считалось ранее».[243] В дополнение к этим двум способам, ведущим к видообразованию, предпочтение и изоляция опылителей, хромосомные перестройки и дивергентный естественный отбор стали критическими для видообразования растений. Кроме того, недавние исследования показывают, что половой отбор, эпигенетический драйверов, и создание несовместимых комбинаций аллелей, вызванных балансирующий выбор также способствуют образованию новых видов.[244] Примеры этих режимов были исследованы как в лаборатории, так и в природе. Исследования также показали, что из-за «сидячей природы растений ... [это увеличивает] относительную важность экологического видообразования ...»[245]
Гибридизация между двумя разными видами иногда приводит к различному фенотипу. Этот фенотип также может быть более подходящим, чем родительская линия, и поэтому естественный отбор может тогда благоприятствовать этим особям. В конце концов, если репродуктивная изоляция будет достигнута, это может привести к появлению отдельного вида. Однако репродуктивной изоляции между гибридами и их родителями добиться особенно трудно, и поэтому гибридное видообразование считается редким событием. Однако гибридизация, приводящая к репродуктивной изоляции, считается важным средством видообразования растений.[246] поскольку полиплоидия (имеющая более двух копий каждой хромосомы) переносится растениями легче, чем животными.[247][248]
Полиплоидия важен для гибридов, поскольку позволяет воспроизводить два разных набора хромосом, каждый из которых может спариваться с идентичным партнером во время мейоза.[249] Полиплоиды также обладают большим генетическим разнообразием, что позволяет им избегать инбридинговая депрессия в небольших популяциях.[250] Гибридизация без изменения числа хромосом называется видообразованием гомоплоидного гибрида. Считается очень редким, но был показан в Геликоний бабочки[251] и подсолнухи. Полиплоидное видообразование, которое включает изменение числа хромосом, является более распространенным явлением, особенно у видов растений.
Полиплоидия - это механизм, который вызвал множество событий быстрого видообразования в симпатрии, потому что потомство, например, от тетраплоидных х диплоидных скрещиваний часто приводит к триплоидному стерильному потомству.[252] Не все полиплоиды репродуктивно изолированы от родительских растений, и поток генов все еще может происходить, например, через триплоидные гибридные х диплоидные спаривания, которые производят тетраплоиды, или спаривания между мейотически нередуцированный гаметы из диплоидов и гаметы из тетраплоидов. Было высказано предположение, что многие из существующих видов растений и животных подверглись полиплоидизации в своей эволюционной истории.[249][253] Размножение успешных полиплоидных видов иногда бывает бесполым путем партеногенез или апомиксис, так как по неизвестным причинам многие бесполые организмы полиплоидны. Известны редкие случаи полиплоидных млекопитающих, но чаще всего они приводят к внутриутробной смерти.
Исследователи считают репродуктивную изоляцию ключом к видообразованию.[254] Важным аспектом исследования видообразования является определение природы барьеров, препятствующих размножению. Ботаники часто считают зоологические классификации презиготических и постзиготных барьеров неадекватными.[254] Приведенные ниже примеры дают представление о процессе видообразования.
Mimulus peregrinus
Создание нового аллополиплоидного вида обезьянника (Mimulus peregrinus ) наблюдался на берегу Shortcleuch Water - реки в Ледхиллс, Южный Ланаркшир, Шотландия. От скрещивания двух видов Mimulus guttatus (содержащий 14 пар хромосом) и Mimulus luteus (содержащий 30-31 пару от дупликации хромосомы), М. peregrinus имеет шесть копий своих хромосом (вызванных дупликацией стерильного гибридного триплоида). Из-за природы этих видов они обладают способностью к самооплодотворению. Из-за количества хромосом он не может спариваться с М. guttatus, М. luteus, или их бесплодное триплоидное потомство. М. peregrinus либо умрет, не производя потомства, либо будет эффективно воспроизводиться, что приведет к появлению нового вида.[255][256]
Raphanobrassica
Raphanobrassica включает все межродовые гибриды между родами Raphanus (редис) и Brassica (капуста и др.).[257][258] В Raphanobrassica является аллополиплоид помесь редис (Raphanus sativus) и капуста (Brassica oleracea). Растения этого происхождения теперь известны как радиколь. Две другие плодородные формы Raphanobrassica известны. Рапарадиш, аллополиплоидный гибрид между Raphanus sativus и Brassica rapa выращивается как кормовая культура. "Raphanofortii" представляет собой аллополиплоидный гибрид между Brassica tournefortii и Raphanus caudatus. В Raphanobrassica - интересное растение, потому что (несмотря на гибридную природу) оно не стерильно. Это привело некоторых ботаников к предположению, что случайная гибридизация цветка с пыльцой другого вида в природе может быть механизмом видообразования, обычным для высших растений.
Сенесио (земля)
Валлийский сульчик - это аллополиплоид, растение, которое содержит наборы хромосом двух разных видов. Его предком был Senecio × baxteri, бесплодный гибрид, который может возникнуть спонтанно, когда близкородственный наземный сель (Senecio vulgaris ) и оксфордский крестовник (Senecio squalidus ) растут рядом друг с другом. Где-то в начале 20 века случайное удвоение числа хромосом в S. × baxteri растение привело к образованию нового плодородного вида.[259][260]
В York Groundsel (Senecio eboracensis) - гибридный вид несовместимый с самим собой Senecio squalidus (также известный как оксфордский крестовник) и самосовместимый Senecio vulgaris (также известный как обыкновенная земляная земля). подобно S. vulgaris, S. eboracensis самосовместим; тем не менее, он демонстрирует незначительное естественное скрещивание со своим родительским видом или вовсе его не скрещивает и, следовательно, репродуктивно изолирован, что указывает на наличие сильных породных барьеров между этим новым гибридом и его родителями. Это произошло в результате обратное скрещивание из F1 гибрид своих родителей S. vulgaris. S. vulgaris родом из Великобритании, а S. squalidus был завезен из Сицилии в начале 18 века; следовательно, S. eboracensis произошел от этих двух видов за последние 300 лет.
Известны и другие гибриды от тех же двух родителей. Некоторые бесплодны, например С. Икс Baxteri. Также известны другие фертильные гибриды, в том числе S. vulgaris var. спячка, теперь распространено в Британии, и аллогексаплоид С. cambrensis, которые, согласно молекулярным данным, вероятно, возникали независимо, по крайней мере, три раза в разных местах. Морфологические и генетические данные подтверждают статус S. eboracensis отдельно от других известных гибридов.[261]
Тале кресс

Кирстен Бомблис и другие. от Институт биологии развития Макса Планка обнаружил два гена в кресс-салате, Arabidopsis thaliana. Когда оба гена наследуются индивидуумом, это вызывает реакцию в гибридном растении, которая настраивает против него его собственную иммунную систему. У родителей гены не были вредными, но они развивались отдельно, чтобы реагировать дефектно при объединении.[262] Чтобы проверить это, Бомбли скрестили 280 генетически различных штаммов Арабидопсис 861 различными способами и обнаружили, что 2 процента полученных гибридов были некротическими. Наряду с одинаковыми показателями, 20 растений также имели сопоставимый набор генетической активности в группе из 1080 генов. Почти во всех случаях Бомблис обнаружил, что только два гена должны были вызвать аутоиммунный ответ. Бомбли детально изучил один гибрид и обнаружил, что один из двух генов принадлежит NB-LRR класс, общая группа генов устойчивости к болезням, участвующих в распознавании новых инфекций. Когда Бомбли удалил проблемный ген, гибриды развились нормально.[262] В течение последующих поколений эта несовместимость могла приводить к разделению между различными штаммами растений, уменьшая их шансы на успешное спаривание и превращая разные сорта в отдельные виды.[263]
Трагопогон (козлобородник)

Трагопогон это один из примеров, когда наблюдалось гибридное видообразование. В начале 20 века люди завезли в Северную Америку три вида сальсифа. Эти виды, западные сальсификации (Трагопогон дубиус), луговая сальсифа (Tragopogon pratensis), а устричное растение (Tragopogon porrifolius), теперь обычные сорняки на городских пустырях. В 1950-х годах ботаники обнаружили два новых вида в регионах Айдахо и Вашингтон, где пересекались три уже известных вида. Один новый вид, Tragopogon miscellus, это тетраплоид гибрид T. dubius и T. pratensis. Другой новый вид, Трагопогонный вирус, также является аллополиплоидом, но его предки были T. dubius и T. porrifolius. Эти новые виды обычно называют «гибридами Оунбей» по имени ботаника, впервые описавшего их. В T. mirus популяция растет в основном за счет воспроизводства собственных представителей, но дополнительные эпизоды гибридизации продолжают увеличивать T. mirus Население.[264]
T. dubius и T. pratensis спаривались в Европе, но так и не смогли скрещиваться. Исследование, опубликованное в марте 2011 года, показало, что, когда эти два растения были завезены в Северную Америку в 1920-х годах, они спарились и удвоили количество хромосом в гибриде. Tragopogon miscellus позволяя «перезагрузить» его гены, что, в свою очередь, допускает большее генетическое разнообразие. Профессор Дуг Солтис из Университет Флориды сказал: «Мы уловили эволюцию в действии… Новые и разнообразные модели экспрессии генов могут позволить новым видам быстро адаптироваться в новой среде».[265][266] Это наблюдаемое событие видообразования посредством гибридизации дополнительно подтверждает доказательства общего происхождения организмов и временные рамки, в которых новый вид возник в новой среде. Гибридизации воспроизводились искусственно в лабораториях с 2004 г. по настоящее время.
Позвоночные
Blackcap
Виды птиц, Сильвия атрикапилла, обычно называемые чернокожими, обитают в Германии и летят на юго-запад, в Испанию, тогда как небольшая группа летает зимой на северо-запад в Великобританию. Грегор Ролсхаузен из Фрайбургский университет обнаружили, что генетическое разделение двух популяций уже происходит. Обнаруженные различия возникли примерно в 30 поколениях. С помощью секвенирования ДНК людей можно отнести к правильной группе с точностью 85%. Стюарт Беархоп из Эксетерский университет сообщили, что птицы, зимующие в Англии, обычно спариваются только между собой, а не обычно с птицами, зимующими в Средиземном море.[267] По-прежнему остается вывод, что популяции станут двумя разными видами, но исследователи ожидают этого из-за продолжающегося генетического и географического разделения.[268]
Моллинезии
Молли короткоперый (Poecilia mexicana ) - небольшая рыбка, обитающая в Серные пещеры Мексики. Годы изучения этого вида показали, что две различные популяции моллинезий - темная внутренняя рыба и светлая поверхностная рыба - становятся все более генетически расходящимися.[269] У населения нет очевидного барьера, разделяющего их; однако было обнаружено, что на молли охотятся большие водяные клопы (Белостома виды ). Тоблер собрал жука и оба вида молли, поместил их в большие пластиковые бутылки и положил обратно в пещеру. Через день было обнаружено, что на свету адаптированные к пещерам рыбы пострадали больше всего: четыре из каждых пяти ножевых ранений были нанесены водяными клопами острым ротовым аппаратом. В темноте ситуация была противоположной. Чувства моллинезий могут обнаружить угрозу хищника в своей среде обитания, но не в других. Переезд из одного места обитания в другое значительно увеличивает риск смерти. Тоблер планирует дальнейшие эксперименты, но считает, что это хороший пример появления нового вида.[270]
Полярный медведь
Естественный отбор, географическая изоляция и происходящее видообразование иллюстрируются взаимоотношениями между Полярный медведь (Ursus maritimus) и бурый медведь (Ursus arctos). Считаются отдельными видами во всем их ареале;[271] однако документально подтверждено, что они обладают способностью скрещиваться и производить плодовитое потомство. Эта интрогрессивная гибридизация встречался как в дикой природе, так и в неволе и был задокументирован[272] и проверено тестированием ДНК.[273] Самые старые известные ископаемые останки белых медведей датируются примерно 130 000–110 000 лет назад;[274] однако молекулярные данные показали разные оценки времени расхождения. Анализ митохондриальной ДНК дал оценку 150 000 лет назад.[274] в то время как анализ ядерного генома показал приблизительное расхождение 603 000 лет назад.[275] Недавние исследования с использованием полных геномов (а не мтДНК или частичных ядерных геномов) устанавливают расхождение полярных и бурых медведей между 479-343 тысячами лет назад.[276] Несмотря на различия в скорости дивергенции, молекулярные исследования показывают, что сестринские виды претерпели очень сложный процесс видообразования и смешения между ними.[277]
Белые медведи приобрели значительные анатомические и физиологические отличия от бурого медведя, которые позволяют ему с комфортом выживать в условиях, которые бурый медведь, вероятно, не мог бы. Известные примеры включают способность плавать шестьдесят миль или более за один раз в ледяной воде, мех, который смешивается со снегом, и оставаться в тепле в арктической среде, удлиненная шея, которая позволяет им легче держать голову над водой во время плавания, и большие перепончатые лапы с толстыми матами, которые играют роль лопастей при плавании. Он также развил маленькие сосочки и вакуумные присоски на подошвах, чтобы уменьшить вероятность скольжения по льду, наряду с меньшими ушами для уменьшения потери тепла, веками, которые действуют как солнцезащитные очки, приспособлениями для их мясной диеты, большой объем желудка, позволяющий кормить по условно-патологическому принципу, и способность голодать до девяти месяцев, перерабатывая мочевину.[278][279] Этот пример представляет собой макроэволюционное изменение, включающее слияние нескольких областей эволюционной биологии, например адаптация посредством естественного отбора, географической изоляции, видообразования и гибридизации.
Доказательства окраски

Окраска животных предоставили важные ранние доказательства эволюции путем естественного отбора в то время, когда было мало прямых доказательств. Во второй половине XIX века были обнаружены три основные функции окраски, которые впоследствии использовались в качестве доказательства отбора: камуфляж (защитная окраска); мимикрия, и то и другое Батезиан и Мюллериан; и апосематизм. После косвенных доказательств, представленных Дарвином в О происхождении видов, и учитывая отсутствие механизмов генетической изменчивости или наследственности в то время, натуралисты, включая современников Дарвина, Генри Уолтер Бейтс и Фриц Мюллер искали доказательства из того, что они могли наблюдать в поле.[281]
Мимикрия и апосематизм
Бейтс и Мюллер описали формы мимикрии, которые теперь носят их имена, основываясь на своих наблюдениях за тропическими бабочками. Эти весьма специфические образцы окраски легко объясняются естественным отбором, поскольку хищники, такие как птицы, которые охотятся на зрение, чаще ловят и убивают насекомых, которые менее хорошо имитируют неприятные модели, чем те, которые лучше имитируют; но закономерности иначе объяснить трудно.[282] Дарвинисты, такие как Альфред Рассел Уоллес и Эдвард Бэгнолл Поултон, а в 20 веке Хью Котт и Бернард Кеттлуэлл, искали доказательства того, что имеет место естественный отбор.[283][284]
Камуфляж
В 1889 году Уоллес отметил, что снежный камуфляж, особенно оперение и мех, которые менялись в зависимости от времени года, предложили очевидное объяснение как приспособление к укрыванию.[285][280] Книга Поултона 1890 года, Цвета животных, написано во время упадка дарвинизма, использовала все формы окраски, чтобы отстаивать естественный отбор.[286] Котт описал множество видов камуфляжа, мимикрии и предупреждающей окраски в своей книге 1940 года. Адаптивная окраска животных, и в частности его рисунки совпадающая деструктивная окраска на лягушках убедили других биологов, что эти обманчивые отметины были результатом естественного отбора.[283] Кеттлвелл экспериментировал эволюция пяденицы, показывая, что вид имел адаптированы, поскольку загрязнение изменило окружающую среду; это предоставило неопровержимые доказательства дарвиновской эволюции.[284]
Данные математического моделирования и симуляции
Информатика позволяет итерация самоизменяющихся сложные системы быть изученным, позволяющим математически понять природу процессов, лежащих в основе эволюции; предоставление доказательств скрытых причин известных эволюционных событий. Эволюция специфических клеточных механизмов, таких как сплайсосомы которые могут превратить геном клетки в огромную мастерскую из миллиардов взаимозаменяемых частей, которые могут создавать инструменты, которые создают нас, могут быть впервые изучены точным образом.
«Потребовалось более пяти десятилетий, но теперь электронный компьютер достаточно мощный, чтобы моделировать эволюцию»,[287] помощь биоинформатика в попытке решить биологические проблемы.
Вычислительная эволюционная биология позволила исследователям проследить эволюцию большого числа организмов путем измерения изменений в их ДНК, а не только с помощью физической систематики или физиологических наблюдений. Он сравнил целые геномы, что позволило изучить более сложные эволюционные события, такие как дупликация гена, горизонтальный перенос генов и предсказание факторов, важных для видообразования. Это также помогло построить сложные вычислительные модели популяций для прогнозирования результатов работы системы с течением времени, а также для отслеживания и обмена информацией о все большем количестве видов и организмов.
В будущем планируется восстановить более сложное древо жизни.
Кристоф Адами, профессор Институт Кека сделал это в Эволюция биологической сложности:
- Чтобы обосновать или противостоять тенденции эволюции сложности в биологической эволюции, сложность должна быть как строго определена, так и измерима. Недавнее теоретико-информационное (но интуитивно очевидное) определение идентифицирует сложность генома с объемом информации, которую последовательность хранит о своем окружении. Мы исследуем эволюцию сложности генома в популяциях цифровых организмов и подробно отслеживаем эволюционные переходы, которые увеличивают сложность. Мы показываем это, потому что естественный отбор заставляет геномы вести себя как естественные "Максвелл Демон "в фиксированной среде геномная сложность вынуждена возрастать.[288]
Дэвид Дж. Эрл и Майкл В. Дим - профессора в Университет Райса указал на это в Эволюционируемость - это выбираемая черта:
- Не только жизнь эволюционировала, но и жизнь эволюционировала, чтобы развиваться. То есть корреляции внутри белковой структуры эволюционировали, и механизмы манипулирования этими корреляциями развивались одновременно. Темпы, с которыми происходят различные события в иерархии эволюционных движений, не случайны или произвольны, а выбираются дарвиновской эволюцией. Разумно, быстрые или экстремальные изменения окружающей среды приводят к отбору для большей эволюционируемости. Этот отбор не запрещен причинно-следственной связью и сильнее всего проявляется на самых крупных шагах внутри мутационной иерархии. Многие наблюдения в рамках эволюционной биологии, которые до сих пор считались эволюционными случайностями или случайностями, объясняются отбором по эволюционируемости. Например, иммунная система позвоночных показывает, что изменчивое окружение антигенов оказывает селективное давление на использование адаптируемых кодонов и полимераз низкой точности во время соматическая гипермутация. Аналогичная движущая сила для предвзятого использования кодонов в результате продуктивно высокой частоты мутаций наблюдается в белке гемагглютинина грипп А.[289]
Компьютерное моделирование эволюции линейных последовательностей продемонстрировало важность рекомбинации блоков последовательности, а не только точечного мутагенеза. Повторяющиеся циклы точечного мутагенеза, рекомбинации и отбора должны позволить in vitro молекулярная эволюция сложных последовательностей, таких как белки ».[290] Эволюционная молекулярная инженерия, также называемая направленной эволюцией или in vitro Молекулярная эволюция включает в себя повторяющийся цикл мутаций, размножения с рекомбинацией и выбора наиболее подходящих индивидуальных молекул (белков, ДНК и РНК). Можно заново пережить естественную эволюцию, показывая нам возможные пути от каталитических циклов, основанных на белках, к основанным на РНК и основанным на ДНК.[290][291][292][293]
В Вида компьютерное моделирование использовалось для проверки доказательств общего происхождения и естественного отбора.[294][295] В одном примере это было использовано, чтобы продемонстрировать, что естественный отбор может способствовать альтруизму, что было предсказано, но трудно проверить эмпирически. При более высоких скоростях репликации, допускаемых моделированием, это становится наблюдаемым.[296]
Смотрите также
использованная литература
- ^ Маунт, Д. (2004). Биоинформатика: анализ последовательности и генома (2-е изд.). Пресса лаборатории Колд-Спринг-Харбор: Колд-Спринг-Харбор, Нью-Йорк. ISBN 978-0-87969-608-5.
- ^ Пенни, Дэвид; Foulds, L.R .; Хенди, М. Д. (1982). «Проверка теории эволюции путем сравнения филогенетических деревьев, построенных из пяти различных белковых последовательностей». Природа. 297 (5863): 197–200. Bibcode:1982Натура.297..197P. Дои:10.1038 / 297197a0. PMID 7078635. S2CID 4270111.
- ^ «Эукариоты». www.tolweb.org. Получено 23 июн 2018.
- ^ Макс, Эдвард (5 мая 2003 г.). «Плагиат ошибок и молекулярная генетика». Архив Talk Origins. Получено 4 мая 2018.
- ^ Футуйма, Дуглас Дж. (1998). Эволюционная биология (3-е изд.). Sinauer Associates. С. 108–110. ISBN 978-0-87893-189-7.
- ^ Haszprunar (1995). «Mollusca: Целоматные турбеллярии или мезенхиматозные кольчатые червяки?». У Тейлора (ред.). Происхождение и эволюционное излучение Mollusca: столетний симпозиум Малакологического общества Лондона. Оксфорд: Издательство Оксфордского университета. ISBN 978-0-19-854980-2.
- ^ Козмик, З .; Daube, M .; Frei, E .; Norman, B .; Kos, L .; Dishaw, L.J .; Noll, M .; Пятигорский, Дж. (2003). «Роль генов Pax в эволюции глаза: ген PaxB книдарии, объединяющий функции Pax2 и Pax6» (PDF). Клетка развития. 5 (5): 773–785. Дои:10.1016 / S1534-5807 (03) 00325-3. PMID 14602077.
- ^ Лэнд, М.Ф. и Нильссон, Д.-Э., Глаза животных, Oxford University Press, Oxford (2002). ISBN 0-19-850968-5.
- ^ Chen, F.C .; Ли, W.H. (2001). «Геномные расхождения между людьми и другими гоминоидами и эффективный размер популяции общего предка людей и шимпанзе». Американский журнал генетики человека. 68 (2): 444–56. Дои:10.1086/318206. ЧВК 1235277. PMID 11170892.
- ^ Cooper, G.M .; Брудно, М .; Грин, E.D .; Batzoglou, S .; Сидов, А. (2003). «Количественные оценки расхождения последовательностей для сравнительного анализа геномов млекопитающих». Genome Res. 13 (5): 813–20. Дои:10.1101 / гр.1064503. ЧВК 430923. PMID 12727901.
- ^ Картинка с надписью «Хромосома 2 человека и ее аналоги у обезьян» в статье Сравнение хромосом человека и человекообразных обезьян как свидетельство общего предка В архиве 2011-08-20 в WebCite Это буквально изображение связи у людей, которая связывает две отдельные хромосомы у нечеловеческих обезьян, создавая одну хромосому у людей. Кроме того, хотя этот термин первоначально относился к свидетельствам окаменелостей, это тоже след из прошлого, соответствующий некоторым живым существам, которые при жизни физически воплощали эту связь.
- ^ Нью-Йорк Таймс отчет Все еще развиваются, человеческие гены рассказывают новую историю, на основе Карта недавнего положительного отбора в геноме человека, заявляет Международный проект HapMap "предоставляет самые убедительные доказательства того, что люди все еще развиваются" и подробно описывает некоторые из этих свидетельств.
- ^ Альбертс, Брюс; Джонсон, Александр; Льюис, Джулиан; Рафф, Мартин; Робертс, Кейт; Уолтер, Питер (Март 2002 г.). Молекулярная биология клетки (4-е изд.). Рутледж. ISBN 978-0-8153-3218-3.
- ^ «Конвергентные доказательства эволюции». В архиве 01.12.2010 в Wayback Machine Философский интеллект: эволюция для всех. 26 ноября 2010 г.
- ^ Петров, Д.А .; Хартл, Д.Л. (2000). «Эволюция псевдогена и естественный отбор для компактного генома». Журнал наследственности. 91 (3): 221–7. Дои:10.1093 / jhered / 91.3.221. PMID 10833048.
- ^ а б Сяо-Цзе, Лу; Ай-Мэй, Гао; Ли-Хуан, Цзи; Цзян, Сюй (1 января 2015 г.). «Псевдоген при раке: реальные функции и многообещающая сигнатура». Журнал медицинской генетики. 52 (1): 17–24. Дои:10.1136 / jmedgenet-2014-102785. ISSN 0022-2593. PMID 25391452.
- ^ Ванин, Э Ф (1985). «Обработанные псевдогены: характеристики и эволюция». Ежегодный обзор генетики. 19 (1): 253–272. Дои:10.1146 / annurev.ge.19.120185.001345. ISSN 0066-4197. PMID 3909943.
- ^ Герштейн, Марк (2006). «Псевдогены в областях ENCODE: согласованная аннотация, анализ транскрипции и эволюции» (PDF). Геномные исследования. 17 (6): 839–51. Дои:10.1101 / гр.5586307. ЧВК 1891343. PMID 17568002. Получено 23 июн 2018.
- ^ "Что такое мусорная ДНК?". Новости-Medical.net. 7 мая 2010. Получено 23 июн 2018.
- ^ Okamoto, N .; Иноуэ, И. (2005). «Вторичный симбиоз в процессе». Наука. 310 (5746): 287. Дои:10.1126 / science.1116125. PMID 16224014. S2CID 22081618.
- ^ Okamoto, N .; Иноуэ, И. (2006). «Hatena arenicola gen. Et sp. Nov., Катаблефарид, который, вероятно, приобретает пластид». Протист. 157 (4): 401–19. Дои:10.1016 / j.protis.2006.05.011. PMID 16891155.
- ^ МакЭндрю, Алек. Хромосома 2 человека - это слияние двух наследственных хромосом.. Доступ 18 мая 2006 г.
- ^ Доказательства общего предка: хромосома человека 2 (видео) 2007
- ^ Yunis, J.J .; Пракаш О. (1982). «Происхождение человека: хромосомное графическое наследие». Наука. 215 (4539): 1525–1530. Bibcode:1982Научный ... 215.1525Y. Дои:10.1126 / science.7063861. PMID 7063861.
- ^ Хромосомы человека и обезьяны В архиве 2011-08-20 в WebCite; по состоянию на 8 сентября 2007 г.
- ^ Аварелло, Розамария; Pedicini, A; Кайуло, А; Zuffardi, O; Фраккаро, М. (1992). «Доказательства наличия предкового альфоидного домена на длинном плече хромосомы 2 человека». Генетика человека. 89 (2): 247–9. Дои:10.1007 / BF00217134. PMID 1587535. S2CID 1441285.
- ^ а б Ijdo, J. W .; Бальдини, А; Уорд, округ Колумбия; Ридерс, СТ; Уэллс, Р.А. (1991). "Происхождение хромосомы 2 человека: исконное слияние теломер и теломер". Труды Национальной академии наук. 88 (20): 9051–5. Bibcode:1991PNAS ... 88.9051I. Дои:10.1073 / пнас.88.20.9051. ЧВК 52649. PMID 1924367.
- ^ а б Аминокислотные последовательности в белках цитохрома с разных видов, адаптировано из Strahler, Arthur; Наука и история Земли, 1997. стр. 348.
- ^ Lurquin, P.F .; Стоун, Л. (2006). Гены, культура и эволюция человека: синтез. Blackwell Publishing, Incorporated. п. 79. ISBN 978-1-4051-5089-7.
- ^ а б Теобальд, Дуглас (2004). «29+ доказательств макроэволюции; функциональная избыточность белков]». Архив Talk Origins.
- ^ Кастресана, Дж. (2001). «Цитохром б Филогения и систематика человекообразных обезьян и млекопитающих ». Молекулярная биология и эволюция. 18 (4): 465–471. Дои:10.1093 / oxfordjournals.molbev.a003825. PMID 11264397.
- ^ Van Der Kuyl, A.C .; Dekker, J.T .; Гоудсмит, Дж. (1999). «Открытие нового эндогенного ретровируса типа C (FcEV) у кошек: доказательства того, что RD-114 является рекомбинантным рекомбинантным вирусом BaEVEnv FcEVGag-Pol / павиана». Журнал вирусологии. 73 (10): 7994–8002. Дои:10.1128 / JVI.73.10.7994-8002.1999. ЧВК 112814. PMID 10482547.
- ^ Свердлов, Э. (Февраль 2000 г.). «Ретровирусы и эволюция приматов». BioEssays. 22 (2): 161–71. Дои:10.1002 / (SICI) 1521-1878 (200002) 22: 2 <161 :: AID-BIES7> 3.0.CO; 2-X. PMID 10655035.
- ^ Belshaw, R .; Pereira, V .; Katzourakis, A .; и другие. (Апрель 2004 г.). «Длительная реинфекция генома человека эндогенными ретровирусами». Труды Национальной академии наук Соединенных Штатов Америки. 101 (14): 4894–9. Bibcode:2004ПНАС..101.4894Б. Дои:10.1073 / pnas.0307800101. ЧВК 387345. PMID 15044706.
- ^ Bonner, T.I .; О'Коннелл, С .; Коэн, М. (август 1982 г.). «Клонированные эндогенные ретровирусные последовательности из ДНК человека». Труды Национальной академии наук Соединенных Штатов Америки. 79 (15): 4709–13. Bibcode:1982PNAS ... 79.4709B. Дои:10.1073 / пнас.79.15.4709. ЧВК 346746. PMID 6181510.
- ^ Джонсон, Велкин Э .; Гроб, Джон М. (31 августа 1999 г.). «Построение филогении приматов из последовательностей древних ретровирусов». Труды Национальной академии наук. 96 (18): 10254–10260. Bibcode:1999PNAS ... 9610254J. Дои:10.1073 / пнас.96.18.10254. ISSN 0027-8424. ЧВК 17875. PMID 10468595.
- ^ Паллен, Марк (2009). Грубый путеводитель по эволюции. Грубые направляющие. С. 200–206. ISBN 978-1-85828-946-5.
- ^ Tanaka, G .; Hou, X .; Максимум.; Edgecombe, G.D .; Штраусфельд, штат Нью-Джерси (октябрь 2013 г.). «Хелицератный образец нейронного основания у кембрийского большого членистоногого придатка». Природа. 502 (7471): 364–367. Bibcode:2013Натура.502..364Т. Дои:10.1038 / природа12520. PMID 24132294. S2CID 4456458.
- ^ Эндрюс, Рой Чепмен (3 июня 1921 г.). "Замечательный случай внешних задних конечностей у горбатого кита" (PDF). Американский музей Novitates.
- ^ а б Холл, Брайан К. (1995), «Атавизмы и атавистические мутации», Природа Генетика, 10 (2): 126–127, Дои:10.1038 / ng0695-126, PMID 7663504, S2CID 27868367
- ^ а б c d е ж г час я Холл, Брайан К. (1984), «Механизмы развития, лежащие в основе атавизмов», Биологические обзоры, 59 (1): 89–124, Дои:10.1111 / j.1469-185x.1984.tb00402.x, PMID 6367843, S2CID 29258934
- ^ Ламберт, Кэти. (2007-10-29) HowStuffWorks "Как работают атавизмы". Animals.howstuffworks.com. Проверено 6 декабря 2011.
- ^ а б Томич, Ненад; и другие. (2011), «Атавизмы: медицинские, генетические и эволюционные последствия», Перспективы биологии и медицины, 54 (3): 332–353, Дои:10.1353 / pbm.2011.0034, PMID 21857125, S2CID 40851098
- ^ Рейно, А. (1977), Сомиты и ранний морфогенез конечностей рептилий. В морфогенезе конечностей и сомитов позвоночных, Cambridge University Press, Лондон, стр. 373–386.
- ^ Табучи, Хироко (2006), У дельфинов могут быть `` остатки '' ног, Livescience.com
- ^ Тайсон, Р .; Graham, J.P .; Colahan, P.T .; Берри, C.R. (2004). «Скелетный атавизм у миниатюрной лошади». Ветеринарная радиология и ультразвук. 45 (4): 315–7. Дои:10.1111 / j.1740-8261.2004.04060.x. PMID 15373256.
- ^ Симпсон, Г. Г. (1951), Лошади: история семейства лошадей в современном мире и шестьдесят миллионов лет эволюции., Oxford University Press
- ^ Дао, Ань Х .; Нетски, Мартин Г. (1984), "Человеческие хвосты и псевдохвосты", Патология человека, 15 (5): 449–453, Дои:10.1016 / S0046-8177 (84) 80079-9, PMID 6373560
- ^ Катя Доумс; и другие. (2007), «Реэволюция сексуальности нарушает закон Долло», Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ., 104 (17): 7139–7144, Bibcode:2007PNAS..104.7139D, Дои:10.1073 / pnas.0700034104, ЧВК 1855408, PMID 17438282
- ^ Харрис, Мэтью П .; и другие. (2006), "Развитие зубов первого поколения архозавров у куриного мутанта", Текущая биология, 16 (4): 371–377, Дои:10.1016 / j.cub.2005.12.047, PMID 16488870
- ^ Майкл Ф. Уайтинг; и другие. (2003), «Утрата и восстановление крыльев у палочников», Природа, 421 (6920): 264–267, Bibcode:2003Натурал.421..264Вт, Дои:10.1038 / природа01313, PMID 12529642, S2CID 962571
- ^ а б Роберт Дж. Райков; и другие. (1979), «Эволюционное восстановление утраченной предковой мышцы в сообществе шалашников», Кондор, 81 (2): 203–206, Дои:10.2307/1367290, JSTOR 1367290
- ^ Роберт Дж. Райкоу (1975), «Эволюционное возрождение мускулов предков как аномалии развития у двух видов птиц», Кондор, 77 (4): 514–517, Дои:10.2307/1366113, JSTOR 1366113
- ^ Э. Эванш (1959), «Аномалии подъязычных мышц у собак (Canis familis)», Анатомическая запись, 133 (2): 145–162, Дои:10.1002 / ар.1091330204, PMID 13670435, S2CID 33397424
- ^ Замок, Уильям Э. (1906), Происхождение полидактильной расы морских свинок (49-е изд.), Вашингтонский институт Карнеги
- ^ Хелд, Льюис И. (2010). "Загадка Evo-Devo моделирования человеческого волоса". Эволюционная биология. 37 (2–3): 113–122. Дои:10.1007 / s11692-010-9085-4. S2CID 12112693.
- ^ а б Футуйма, Дуглас Дж. (1998). Эволюционная биология (3-е изд.). Sinauer Associates Inc. стр. 122. ISBN 978-0-87893-189-7.
- ^ а б 29+ свидетельств макроэволюции: Часть 1. Talkorigins.org. Проверено 6 декабря 2011.
- ^ Койн, Джерри А. (2009). Почему эволюция верна. Викинг. стр.8–11. ISBN 978-0-670-02053-9.
- ^ Дарвин, Чарльз (1859). О происхождении видов. Джон Мюррей. п.420.
- ^ Туоми, Дж. (1981). «Структура и динамика дарвиновской эволюционной теории» (PDF). Syst. Zool. 30 (1): 22–31. Дои:10.2307/2992299. JSTOR 2992299.
- ^ Aravind, L .; Iyer, L.M .; Анантараман, В. (2003). «Два лица Альбы: эволюционная связь между белками, участвующими в структуре хроматина и метаболизмом РНК». Геномная биология. 4 (10): R64. Дои:10.1186 / gb-2003-4-10-r64. ЧВК 328453. PMID 14519199.
- ^ Brochu, C.A .; Wagner, J. R .; Jouve, S .; Sumrall, C.D .; Денсмор, Л. Д. (2009). «Исправление исправлено: консенсус по поводу значения Crocodylia и того, почему это важно» (PDF). Syst. Биол. 58 (5): 537–543. Дои:10.1093 / sysbio / syp053. PMID 20525607. Архивировано из оригинал (PDF) 27 сентября 2013 г.
- ^ Бок, В. Дж. (2007). «Объяснения в эволюционной теории» (PDF). J Zool Syst Evol Res. 45 (2): 89–103. Дои:10.1111 / j.1439-0469.2007.00412.x. Архивировано из оригинал (PDF) 12 мая 2012 г.
- ^ Клюге 1999
- ^ Лаурин 2000
- ^ Фитцхью 2006, п. 31 год
- ^ Клюге 1999, п. 432
- ^ Слифкин, Натан (2006). Вызов творения…. Зоопарк Тора. С. 258–9. ISBN 978-1-933143-15-6.
- ^ Сентер, Фил; и другие. (2015), «Рудиментарные биологические структуры: проверка креационистских гипотез, применимая в классе», Американский учитель биологии, 77 (2): 99–106, Дои:10.1525 / abt.2015.77.2.4, S2CID 85573136
- ^ Аттила Регоэс; и другие. (2005), «Импорт белка, репликация и наследование остатков митохондрий», Журнал биологической химии, 280 (34): 30557–30563, Дои:10.1074 / jbc.M500787200, PMID 15985435
- ^ Секигучи, Хироши; и другие. (2002), "Остаточные хлоропласты в гетеротрофных страменопилах". Pteridomonas danica и Настой цилиофриса (Dictyochophyceae) ", Протист, 153 (2): 157–167, Дои:10.1078/1434-4610-00094, PMID 12125757
- ^ Паула Дж. Рудалл; и другие. (2002), «Цветочная анатомия и систематика луковых растений с особым упором на Gilliesia, Предполагаемый мимик насекомого с сильно зигоморфными цветками ", Американский журнал ботаники, 89 (12): 1867–1883, Дои:10.3732 / ajb.89.12.1867, PMID 21665616
- ^ Strittmatter, Lara I .; и другие. (2002), "Субдиоэзия в Consolea Spinosissima (Cactaceae): Система разведения и эмбриологические исследования », Американский журнал ботаники, 89 (9): 1373–1387, Дои:10.3732 / ajb.89.9.1373, PMID 21665739, S2CID 17169760
- ^ Эшман, Тиа-Линн (2003), «Ограничения на эволюцию самцов и половой диморфизм: полевые оценки генетической архитектуры репродуктивных признаков в трех популяциях гинодобных» Fragaria virginiana", Эволюция, 57 (9): 2012–2025, Дои:10.1554/02-493, PMID 14575323, S2CID 198153247
- ^ Голонка, Аннетт М .; и другие. (2005), "Ветровое опыление, половой диморфизм и изменения цветочных признаков Schiedea (Caryophyllaceae) ", Американский журнал ботаники, 92 (9): 1492–1502, Дои:10.3732 / ajb.92.9.1492, PMID 21646167
- ^ Walker-Larsen, J .; Хардер, Л. Д. (2001), «Рудиментарные органы как возможности для функциональных инноваций: пример стаминодии Penstemon», Эволюция, 55 (3): 477–487, Дои:10.1111 / j.0014-3820.2001.tb00782.x, PMID 11327156, S2CID 19533785
- ^ Gomez, Nadilla N .; Шоу, Рут Г. (2006), «Влияние инбридинга на мужскую и женскую фертильность и наследование мужского бесплодия в Немофила Menziesii (Hydrophyllaceae) ", Американский журнал ботаники, 93 (5): 739–746, Дои:10.3732 / ajb.93.5.739, PMID 21642137
- ^ Бейдер, Ларс; Холл, Брайан К. (2002), "Конечности у китов и безграничность у других позвоночных: механизмы эволюционной трансформации и трансформации развития и потери", Эволюция и развитие, 4 (6): 445–458, Дои:10.1046 / j.1525-142x.2002.02033.x, PMID 12492145, S2CID 8448387
- ^ а б Койн, Джерри А. (2009). Почему эволюция верна. Викинг. стр.60. ISBN 978-0-670-02053-9.
- ^ Вест-Эберхард, Мэри Джейн (2003). Пластичность развития и эволюция. Издательство Оксфордского университета. п. 232. ISBN 978-0-19-512234-3.
- ^ П. К. Симойнс-Лопес; C. S. Gutstein (2004), "Заметки об анатомии, расположении и гомологии костей таза у мелких китообразных (Cetacea, Delphinidae, Pontoporiidae)", Латиноамериканский журнал водных млекопитающих, 3 (2): 157–162, Дои:10.5597 / lajam00060
- ^ «Пример 1. Живые киты и дельфины с задними конечностями». Дуглас Теобальд. Получено 20 марта 2011.
- ^ Гарнер, Стефан; и другие. (2006), «Гибридизация, стабильность развития и функциональность морфологических признаков у жужелицы Carabus solieri (Coleoptera, Carabidae)», Биологический журнал Линнеевского общества, 89: 151–158, Дои:10.1111 / j.1095-8312.2006.00668.x
- ^ Джеффри, Уильям Р. (2008), «Новые модельные системы в evo-DevO: пещерные рыбы и микроэволюция развития», Эволюция и развитие, 10 (3): 256–272, Дои:10.1111 / j.1525-142X.2008.00235.x, ЧВК 3577347, PMID 18460088
- ^ Койн, Джерри А. (2009). Почему эволюция верна. Викинг. стр.59–60. ISBN 978-0-670-02053-9.
- ^ Абед Э. Зубидат; и другие. (2010), "Фотоэнтренмент у слепых и зрячих видов грызунов: реакция на фотофазный свет с разными длинами волн", Журнал экспериментальной биологии, 213 (Пт 24): 4213–4222, Дои:10.1242 / jeb.048629, PMID 21113002
- ^ Кирни, Морин; Майсано, Джессика Андерсон; Роу, Тимоти (2005), "Черепная анатомия вымерших амфисбайцев" Rhineura hatcherii (Squamata, Amphisbaenia) на основе рентгеновской компьютерной томографии высокого разрешения », Журнал морфологии, 264 (1): 1–33, Дои:10.1002 / jmor.10210, PMID 15549718, S2CID 7504517
- ^ Дэвид С. Калвер (1982), Пещерная жизнь: эволюция и экология, Издательство Гарвардского университета, ISBN 9780674330191
- ^ Койн, Джерри А. (2009). Почему эволюция верна. Викинг. стр.57–59. ISBN 978-0-670-02053-9.
- ^ Эрин Э. Максвелл; Hans C.E. Larsson (2007), «Остеология и миология крыла эму (Dromaius novaehollandiae) и ее влияние на эволюцию рудиментарных структур», Журнал морфологии, 268 (5): 423–441, Дои:10.1002 / jmor.10527, PMID 17390336, S2CID 12494187
- ^ Майкл Г. Гласспул (1982), Атлас офтальмологии (1-е изд.), MTP Press Unlimited, p. 9
- ^ Rehoreka, Susan J .; Смит, Тимоти Д. (2006), «Железа Хардера приматов: существует ли она на самом деле?», Анн Анат, 188 (4): 319–327, Дои:10.1016 / j.aanat.2006.01.018, PMID 16856596
- ^ Андраде, Джулия Б .; Lewis, Ryshonda P .; Сентер, Фил (2016), «Аппендикулярные скелеты пяти азиатских видов сцинков из родов. Брахимелы и Офиомор, в том числе виды с рудиментарными отростками », Амфибия-Рептилии, 37 (4): 337–344, Дои:10.1163/15685381-00003062
- ^ Морин Кирни (2002), «Аппендикулярный скелет у амфисбенов (Reptilia: Squamata)», Копея, 2002 (3): 719–738, Дои:10.1643 / 0045-8511 (2002) 002 [0719: asiars] 2.0.co; 2
- ^ Цуйиджи, Таканобу; Кирни, Морин; Оливье Риппель (2006), "Первый отчет о мышце грудного пояса у змей, с комментариями о шейно-спинной границе змеи", Копея, 2006 (2): 206–215, Дои:10.1643 / 0045-8511 (2006) 6 [206: froapg] 2.0.co; 2
- ^ Макгоуэн, Майкл Р .; Кларк, Клей; Гейтси, Джон (2008), "Подгеном рудиментарных обонятельных рецепторов китов-одонтоцетов: филогенетическое соответствие между методами согласования генов и деревьев и методами суперматрицы", Систематическая биология, 57 (4): 574–590, Дои:10.1080/10635150802304787, PMID 18686195
- ^ Nweeia, Martin T .; и другие. (2012), «Рудиментарная анатомия зубов и номенклатура клыков монодона-моноцероса», Анатомический рекорд, 295 (6): 1006–1016, Дои:10.1002 / ar.22449, PMID 22467529, S2CID 22907605
- ^ Таг, Роберт Г. (2002), "Изменчивость метаподиалов у приматов с рудиментарными цифрами: Ателес Жоффройи, Колобус гереза, и Perodicticus potto", Американский журнал физической антропологии, 117 (3): 195–208, Дои:10.1002 / ajpa.10028, PMID 11842399
- ^ Peterková, R .; и другие. (2002), "Развитие рудиментарных зачатков зубов как часть одонтогенеза мышей", Соединительная ткань исследования, 43 (2–3): 120–128, Дои:10.1080/03008200290000745, PMID 12489147, S2CID 33141711
- ^ Лиман, Эмили Р .; Иннан, Хидеки (2003), «Расслабленное избирательное давление на важный компонент трансдукции феромона в эволюции приматов», PNAS, 100 (6): 3328–3332, Bibcode:2003ПНАС..100.3328Л, Дои:10.1073 / pnas.0636123100, ЧВК 152292, PMID 12631698
- ^ Чжан, Цзяньчжи; Уэбб, Дэвид М. (2003), «Эволюционное ухудшение пути вомероназальной трансдукции феромона у катарринных приматов», PNAS, 100 (14): 8337–8341, Bibcode:2003PNAS..100.8337Z, Дои:10.1073 / пнас.1331721100, ЧВК 166230, PMID 12826614
- ^ Тамацу; и другие. (2007), «Остатки вибриссальных капсульных мышц существуют в верхней губе человека», Клиническая анатомия, 20 (6): 628–631, Дои:10.1002 / ок. 20497, PMID 17458869, S2CID 21055062
- ^ Мбака, Годвин О .; Эдживунми, Адедайо Б. (2009), «Распространенность отсутствия ладонной мышцы - исследование в популяции йоруба», Ольстерский медицинский журнал, 78 (2): 90–93, ЧВК 2699194, PMID 19568443
- ^ Друин, Гай; и другие. (2011), «Генетика потери витамина С у позвоночных», Текущая геномика, 12 (5): 371–378, Дои:10.2174/138920211796429736, ЧВК 3145266, PMID 22294879
- ^ Жибер, Жан-Мишель; и другие. (2000), "Barnacle Duplicate" Гравированный гены: Дивергентные паттерны экспрессии и свидетельства наличия остатков брюшной полости ", Эволюция и развитие, 2 (4): 194–202, Дои:10.1046 / j.1525-142x.2000.00059.x, PMID 11252562, S2CID 41665533
- ^ Фрейер, Клаудиа; Ренфри, Мэрилин Б. (2009), "Плацента желточного мешка млекопитающих", Журнал экспериментальной зоологии, 312B (6): 545–554, Дои:10.1002 / jez.b.21239, PMID 18985616
- ^ Браванд, Дэвид; и другие. (2008), «Потеря генов яичного желтка у млекопитающих и происхождение лактации и плацентации», PLOS Биология, 6 (3): e63, Дои:10.1371 / journal.pbio.0060063, ЧВК 2267819, PMID 18351802
- ^ Майерс, П. З. (2008), «Репродуктивная история записана в геноме», Фарингула, ScienceBlogs
- ^ Дэвид Седер; и другие. (1997), «О развитии конечностей китообразных: I. Рудиментация задних конечностей у пятнистого дельфина (Стенелла аттенуата)", Европейский журнал морфологии, 35 (1): 25–30, Дои:10.1076 / ejom.35.1.25.13058, PMID 9143876
- ^ Boke, Norman H .; Андерсон, Эдвард Ф. (1970), "Структура, развитие и таксономия в роде Лофофора", Американский журнал ботаники, 57 (5): 569–578, Дои:10.2307/2441055, JSTOR 2441055
- ^ Мальте Эльбрехтер; Эберхарт Шнепф (1996) "Gymnodinium chlorophorum, новая зеленая цветущая диноагеллята (Gymnodiniales, Dinophyceae) с рудиментарным эндосимбионтом празинофита », Phycologia, 35 (5): 381–393, Дои:10.2216 / i0031-8884-35-5-381.1
- ^ Джеффри С. Принс; Пол Мика Джонсон (2006), «Ультраструктурное сравнение Аплизия и Долабрифера Чернильные железы предполагают клеточные участки производства белка против хищников и обработки водорослевого пигмента », Журнал исследований моллюсков, 72 (4): 349–357, Дои:10.1093 / mollus / eyl017
- ^ Ридли, Марк (2004). Эволюция (3-е изд.). Блэквелл Паблишинг. п. 282. ISBN 978-1-4051-0345-9.
- ^ а б Докинз, Ричард (2009). Величайшее шоу на Земле: свидетельства эволюции. Bantam Press. стр.364–365. ISBN 978-1-4165-9478-9.
- ^ Уильямс, Г. (1992). Естественный отбор: области, уровни и проблемы. Oxford Press. ISBN 978-0-19-506932-7.
- ^ «Признания дарвиниста». Найлз Элдридж. Получено 22 июн 2010.
- ^ Койн, Джерри А. (2009). Почему эволюция верна. Викинг. стр.26–28. ISBN 978-0-670-02053-9.
- ^ Дональд Р. Протеро (2013), Оживление окаменелостей: введение в палеобиологию (3-е изд.), Columbia University Press, стр. 21 год
- ^ "Часто задаваемые вопросы". Музей естественной истории Фонда округа Лос-Анджелес. Архивировано из оригинал 11 марта 2011 г.. Получено 21 февраля 2011.
- ^ Дин, Уильям Ричард Джон; Милтон, Сюзанна Джейн (1999). Кару: экологические закономерности и процессы. Издательство Кембриджского университета. п. 31. ISBN 978-0-521-55450-3.
- ^ Шадевальд, Роберт Дж. (1982). «Шесть аргументов о потопе, на которые креационисты не могут ответить». Журнал Creation Evolution. 3: 12–17.
- ^ а б Дональд Р. Протеро (2008), «Какое недостающее звено?», Новый ученый, 197 (2645): 35–41, Дои:10.1016 / s0262-4079 (08) 60548-5
- ^ «Очевидно, что у позвоночных должны были быть предки, жившие в кембрии, но считалось, что они были беспозвоночными предшественниками настоящих позвоночных - протохордовых. Pikaia широко рекламировалась как старейшее ископаемое протохордовое животное». Ричард Докинз 2004 Рассказ предков Стр. 289, ISBN 0-618-00583-8
- ^ Chen, J. Y .; Huang, D. Y .; Ли, К. В. (1999). «Раннекембрийский череповидный хордовый». Природа. 402 (6761): 518–522. Bibcode:1999Натура 402..518C. Дои:10.1038/990080. S2CID 24895681.
- ^ Shu, D. G .; Morris, S.C .; Han, J .; Zhang, Z. F .; Yasui, K .; Janvier, P .; Chen, L .; Zhang, X. L .; Liu, J. N .; Li, Y .; Лю, H. -Q. (2003), "Голова и позвоночник раннего кембрийского позвоночного. Haikouichthys", Природа, 421 (6922): 526–529, Bibcode:2003Натура.421..526С, Дои:10.1038 / природа01264, PMID 12556891, S2CID 4401274
- ^ Лежандр, Серж (1989). Les communautés de mammifères du Paléogène (Eocène supérieur et Oligocène) Западной Европы: структуры, среда и эволюция. München: F. Pfeil. п. 110. ISBN 978-3-923871-35-3.
- ^ Академия естественных наук - Джозеф Лейди - Американские лошади В архиве 2012-03-05 в Wayback Machine
- ^ Шубин, Нил (2008). Ваша внутренняя рыба. Пантеон. ISBN 978-0-375-42447-2.
- ^ Niedzwiedzki, G .; Szrek, P .; Narkiewicz, K .; Narkiewicz, M .; Альберг, П. (2010). «Пути тетрапод раннего среднего девона в Польше». Природа. 463 (7227): 43–48. Bibcode:2010Натура.463 ... 43N. Дои:10.1038 / природа08623. PMID 20054388. S2CID 4428903.
- ^ Кота-Санчес, Х. Уго; Бомфим-Патрисио, Марсия К. (2010). «Морфология семян, полиплоидия и история эволюции эпифитного кактуса. Rhipsalis baccifera (Cactaceae) " (PDF). Polibotanica. 29: 107–129. Получено 28 февраля 2013.
- ^ Менкхорст, Питер; Найт, Фрэнк (2001). Полевой справочник по млекопитающим Австралии. Издательство Оксфордского университета. п. 14. ISBN 978-0-19-550870-3.
- ^ Оджи, Майкл; Гуден, Бретт; Мюссер, Энн (2006). Ехидна: необычное млекопитающее, откладывающее яйца.. CSIRO Publishing. ISBN 9780643092044.
- ^ "Белые медведи / среда обитания и распространение". Парки и развлечения SeaWorld. Архивировано из оригинал 25 октября 2013 г.. Получено 21 февраля 2011.
- ^ "Сирены мира". Спасите ламантинский клуб. Получено 21 февраля 2011.
- ^ Континентальный дрейф и эволюция В архиве 2006-02-11 в Wayback Machine. Biology.clc.uc.edu (25 марта 2001 г.). Проверено 6 декабря 2011.
- ^ а б c d Койн, Джерри А. (2009). Почему эволюция верна. Викинг. стр.99–110. ISBN 978-0-670-02053-9.
- ^ Мерфи, Джеймс Б.; Чиофи, Клаудио; де ла Пануз, Коломба; Уолш, Trooper, ред. (2002). Комодские драконы: биология и сохранение (серия по биологии и сохранению зоопарков и аквариумов). Вашингтон.: Смитсоновские книги. ISBN 978-1-58834-073-3.
- ^ Бурдик, Алан (25 марта 2007 г.). «Чудо-земля Сокотры, Йемен». АЛАН БЕРДИК. Получено 8 июля 2010.
- ^ «Туатара». Экология Новой Зеландии: живые ископаемые. TerraNature Trust. 2004 г.. Получено 10 ноября 2006.
- ^ «Факты о туатаре». Сохранение: местные виды. Подразделение по угрожаемым видам, Департамент охраны природы, правительство Новой Зеландии. Архивировано из оригинал 30 сентября 2007 г.. Получено 10 февраля 2007.
- ^ «Самый разыскиваемый в Новой Каледонии». Получено 8 июля 2010.
- ^ «Гигантская пушистохвостая облачная крыса (Crateromys schadenbergi)». Архивировано из оригинал 30 июня 2010 г.. Получено 8 июля 2010.
- ^ Рабор, Д.С. (1986). Путеводитель по филиппинской флоре и фауне. Центр управления природными ресурсами, Министерство природных ресурсов и Университет Филиппин.
- ^ Хамфри, Роберт Р. (1974). Буджум и его дом. Университет Аризоны Press. ISBN 978-0816504367.
- ^ Шофилд, Джеймс (27 июля 2001 г.). "Исчезающий тюлень-нерпа на Байкале". The Moscow Times. Получено 27 сентября 2007.
- ^ Болдуин, Б.Г .; Робишо, Р. Х. (1995). «Историческая биогеография и экология Гавайского альянса серебряных мечей (Asteraceae). Новые молекулярные филогенетические перспективы». В Wagner, W. L .; Функ, В. А. (ред.). Гавайская биогеография: эволюция архипелага с горячей точкой. Вашингтон: Пресса Смитсоновского института. С. 259–287.
- ^ «Адаптивная радиация и гибридизация в Гавайском альянсе серебряного меча». Отделение ботаники Гавайского университета.
- ^ Паллен, Марк (2009). Грубый путеводитель по эволюции. Грубые направляющие. п. 87. ISBN 978-1-85828-946-5.
- ^ Irwin, D.E .; Irwin, J.H .; Прайс, Т.Д. (2001). «Кольцевые виды как мосты между микроэволюцией и видообразованием» (PDF). Genetica. 112–113: 223–43. Дои:10.1023 / А: 1013319217703. PMID 11838767. S2CID 7108000.
- ^ Докинз, Ричард (2004). Рассказ предков. Хоутон Миффлин. п. 303. ISBN 978-0-618-00583-3.
- ^ Дэвис, Пол и Кенрик, Пол. 2004. Ископаемые растения. Smithsonian Books (совместно с Лондонским музеем естественной истории), Вашингтон, округ Колумбия. ISBN 1-58834-156-9
- ^ Койн, Джерри А. (2009). Почему эволюция верна. Викинг. п.103. ISBN 978-0-670-02053-9.
- ^ Pioneer Productions (19 января 2010 г.). "Руководство по эпизодам". Как была создана Земля. Сезон 2. Эпизод 8. Исторический канал.
- ^ Ло, Чжэ-Си; Цзи, Цян; Wible, John R .; Юань, Чун-Си (12 декабря 2003 г.). «Раннемеловое трибосфеновое млекопитающее и эволюция метатерии». Наука. 302 (5652): 1934–1940. Bibcode:2003Наука ... 302.1934Л. Дои:10.1126 / science.1090718. PMID 14671295. S2CID 18032860.
- ^ Nilsson, M. A .; Чураков, Г .; Sommer, M .; Van Tran, N .; Земанн, А .; Brosius, J .; Шмитц, Дж. (27 июля 2010 г.). Пенни, Дэвид (ред.). «Отслеживание эволюции сумчатых животных с использованием вставок архаичных геномных ретропозонов». PLOS Биология. 8 (7): e1000436. Дои:10.1371 / journal.pbio.1000436. ЧВК 2910653. PMID 20668664.
- ^ Вудберн, Майкл О .; Зинсмейстер, Уильям Дж. (Октябрь 1982 г.). «Ископаемые млекопитающие из Антарктиды». Наука. 218 (4569): 284–286. Bibcode:1982Наука ... 218..284Вт. Дои:10.1126 / science.218.4569.284. PMID 17838631. S2CID 32304737.
- ^ Goin, Francisco J .; и другие. (Декабрь 1999 г.). «Новые открытия« супподобных »сумчатых из Антарктиды (остров Сеймур, средний эоцен)». Журнал эволюции млекопитающих. 6 (4): 335–365. Дои:10.1023 / А: 1027357927460. S2CID 2319361.
- ^ Регуэро, Марсело А .; Marenssi, Sergio A .; Сантильяна, Серджио Н. (май 2002 г.). «Антарктический полуостров и Южная Америка (Патагония) Палеогеновые наземные фауны и окружающая среда: биогеографические связи». Палеогеография, палеоклиматология, палеоэкология. 179 (3–4): 189–210. Bibcode:2002ППП ... 179..189р. Дои:10.1016 / S0031-0182 (01) 00417-5.
- ^ Миллс, Уильям Джеймс. Изучение полярных границ: историческая энциклопедия, ABC-CLIO, 2003. ISBN 1-57607-422-6, ISBN 978-1-57607-422-0
- ^ Goin, F.J .; Reguero, M.A .; Pascual, R .; von Koenigswald, W .; Woodburne, M.O .; Case, J.A .; Marenssi, S.A .; Vieytes, C .; Бискайно, С.Ф. (2006). «Первое гондванатерийское млекопитающее из Антарктиды». Геологическое общество, Лондон, Специальные публикации. 258 (1): 135–144. Bibcode:2006ГСЛСП.258..135Г. Дои:10.1144 / GSL.SP.2006.258.01.10. S2CID 129493664.
- ^ Протеро, Дональд Р .; Шох, Роберт М. (2002). Рога, бивни и ласты: эволюция копытных млекопитающих. JHU пресс. п. 45. ISBN 978-0-8018-7135-1.
- ^ а б Джеймс М. Собел; и другие. (2009), «Биология видообразования», Эволюция, 64 (2): 295–315, Дои:10.1111 / j.1558-5646.2009.00877.x, PMID 19891628, S2CID 10168162
- ^ а б Джерри А. Койн; Х. Аллен Орр (2004), Видообразование, Sinauer Associates, стр. 5, ISBN 978-0-87893-089-0
- ^ Рэйвен, Питер Х. (2005). Биология растений (7-е изд.). Нью-Йорк: W.H. Фримен. ISBN 978-0-7167-6284-3. OCLC 183148564.
- ^ Хаас, Дж. У. мл. (Июнь 2000 г.). "Преподобный доктор Уильям Х. Даллинджер F.R.S .: ранний защитник теистической эволюции. и Враг спонтанного зарождения ". Перспективы науки и христианской веры. 52: 107–117. Получено 15 июн 2010.
- ^ а б Ле Паж, Майкл (16 апреля 2008 г.). «Н.С.: бактерии совершают главный эволюционный сдвиг в лаборатории». Новый ученый. Получено 9 июля 2012.
- ^ Блаунт З.Д .; Borland C.Z .; Ленский, Р. (Июнь 2008 г.). «Историческая случайность и эволюция ключевого нововведения в экспериментальной популяции Escherichia coli». Труды Национальной академии наук Соединенных Штатов Америки. 105 (23): 7899–906. Bibcode:2008PNAS..105.7899B. Дои:10.1073 / pnas.0803151105. ЧВК 2430337. PMID 18524956.
- ^ Ленски, Ричард Э .; Травизано, Майкл (1994). «Динамика адаптации и диверсификации: эксперимент 10 000 поколений с бактериальными популяциями». PNAS. 91 (15): 6808–6814. Bibcode:1994PNAS ... 91.6808L. Дои:10.1073 / пнас.91.15.6808. ЧВК 44287. PMID 8041701.
- ^ Турко, Патрик; и другие. (2016), «Быстрая эволюционная потеря сопротивления металлов, обнаруженная в результате вылупления яиц, вылупившихся десятилетиями», Эволюция, 70 (2): 398–407, Дои:10.1111 / evo.12859, PMID 26768308, S2CID 205124469
- ^ Танака, Т .; Хашимото, Х. (1989). «Лекарственная устойчивость и возможность передачи штаммов Shigella, выделенных в 1986 году в Японии». Кансенсогаку Засши. 63 (1): 15–26. Дои:10.11150 / kansenshogakuzasshi1970.63.15. PMID 2501419.
- ^ Thwaites, W.M. (Лето 1985 г.). «Новые белки без Божьей помощи». Журнал Creation Evolution. 5 (2): 1–3.
- ^ Эволюция и информация: нейлоновый жук. Nmsr.org. Проверено 6 декабря 2011.
- ^ Тан, Кер (23 сентября 2005 г.). «Почему ученые отвергают разумный замысел». Новости NBC.
- ^ Миллер, Кеннет Р. Только теория: эволюция и битва за душу Америки (2008) стр. 80–82
- ^ Шон Станковски; Мэтью А. Стрейсфельд (2015), «Интрогрессивная гибридизация способствует адаптивной дивергенции в недавнем излучении цветков обезьян», Труды Королевского общества B: биологические науки, 282 (1814): 20151666, Дои:10.1098 / rspb.2015.1666, ЧВК 4571715, PMID 26311673
- ^ а б Новости науки, Темная сила: пигмент, похоже, находит хорошее применение радиации, Неделя от 26 мая 2007 г .; Vol. 171, № 21, с. 325 от Давиде Кастельвекки
- ^ Дадачева, Е .; Bryan, R.A .; Хуанг, X .; Moadel, T .; Schweitzer, A.D .; Aisen, P .; Носанчук, Ю.Д .; Касадеваль, А. (2007). Резерфорд, Джулиан (ред.). «Ионизирующее излучение изменяет электронные свойства меланина и усиливает рост меланизированных грибов». PLOS ONE. 2 (5): e457. Bibcode:2007PLoSO ... 2..457D. Дои:10.1371 / journal.pone.0000457. ЧВК 1866175. PMID 17520016.
- ^ а б Джон А. Эндлер (1980). «Естественный отбор по цветным узорам у Poecilia reticulata». Эволюция. 34 (1): 76–91. Дои:10.2307/2408316. JSTOR 2408316. PMID 28563214.
- ^ Дэвид Н. Резник; Фрэнк Х. Шоу; Ф. Хелен Родд; Рут Г. Шоу (1997). «Оценка скорости эволюции природных популяций гуппи (Poecilia reticulata)". Наука. 275 (5308): 1934–1937. Дои:10.1126 / science.275.5308.1934. PMID 9072971. S2CID 18480502.
- ^ Дэвид Резник; Джон А. Эндлер (1982). «Влияние хищничества на эволюцию жизненного цикла тринидадских гуппи (Poecilia reticulata)». Эволюция. 36 (1): 160–177. Дои:10.2307/2407978. JSTOR 2407978. PMID 28581096.
- ^ Совет медицинских исследований (Великобритания) (21 ноября 2009 г.). «Ген устойчивости к заболеванию головного мозга развивается в сообществе Папуа-Новой Гвинеи; может помочь в понимании CJD». Science Daily (онлайн). Новости науки. Получено 22 ноября 2009.
- ^ Mead, S .; Whitfield, J .; Poulter, M .; Shah, P .; Uphill, J .; Кэмпбелл, Т .; Аль-Дуджайли, H .; Hummerich, H .; Beck, J .; Mein, C.A .; Verzilli, C .; Whittaker, J .; Alpers, M. P .; Коллиндж, Дж. (2009). «Новый вариант защитного прионного протеина, который колокализируется при воздействии Куру» (PDF). Медицинский журнал Новой Англии. 361 (21): 2056–2065. Дои:10.1056 / NEJMoa0809716. PMID 19923577.
- ^ Byars, S. G .; Ewbank, D .; Govindaraju, D. R .; Стернс, С. К. (2009). «Естественный отбор в современной человеческой популяции». Труды Национальной академии наук. 107 (suppl_1): 1787–1792. Bibcode:2010PNAS..107.1787B. Дои:10.1073 / pnas.0906199106. ЧВК 2868295. PMID 19858476.
- ^ Непереносимость сои и лактозы Обратный путь: соевое питание
- ^ Enattah, N.S .; Sahi, T .; Savilahti, E .; Terwilliger, J.D .; Peltonen, L .; Ярвеля, И. (2002). «Идентификация варианта, связанного с гиполактазией взрослого типа». Природа Генетика. 30 (2): 233–7. Дои:10,1038 / ng826. PMID 11788828. S2CID 21430931.
- ^ Карри, Эндрю (31 июля 2013 г.). «Археология: молочная революция». Природа. 500 (7460): 20–2. Bibcode:2013Натура 500 ... 20С. Дои:10.1038 / 500020a. PMID 23903732.
- ^ Ласточка DM (2003). «Генетика персистенции лактазы и непереносимости лактозы». Ежегодный обзор генетики. 37: 197–219. Дои:10.1146 / annurev.genet.37.110801.143820. PMID 14616060.
- ^ Харриет, Коулз (20 января 2007 г.). «Ген лактазы в Африке: вы пьете молоко?». Геном человека, Wellcome Trust. Архивировано из оригинал 29 сентября 2008 г.. Получено 18 июля 2008.
- ^ Тишкофф, С.А .; Reed, F.A .; Ranciaro, A .; и другие. (Январь 2007 г.). «Конвергентная адаптация персистенции лактазы человека в Африке и Европе». Природа Генетика. 39 (1): 31–40. Дои:10,1038 / ng1946. ЧВК 2672153. PMID 17159977.
- ^ а б c d е ж г час я Herrel, A .; Huyghe, K .; Vanhooydonck, B .; и другие. (Март 2008 г.). «Быстрое крупномасштабное эволюционное расхождение в морфологии и производительности, связанное с использованием другого пищевого ресурса». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 105 (12): 4792–5. Bibcode:2008PNAS..105.4792H. Дои:10.1073 / pnas.0711998105. ЧВК 2290806. PMID 18344323.
- ^ а б c d Барт Вервуст; Ирена Грбак; Рауль Ван Дамм (август 2007 г.). "Различия в морфологии, производительности и поведении между недавно разошедшимися популяциями Podarcis sicula зеркальные различия в давлении хищников ". Ойкос. 116 (8): 1343–1352. Дои:10.1111 / j.0030-1299.2007.15989.x. S2CID 55765174.
- ^ «Ящерицы быстро развиваются после знакомства с островом». Национальная география. 21 апреля 2008 г.
- ^ Майерс, П.З. (23 апреля 2008 г.). "Все еще просто ящерица". ScienceBlogs.
- ^ Кларк, Брайан В .; и другие. (2013). «Различия в устойчивости к полициклическим ароматическим углеводородам и ПХБ-126 среди субпопуляций Fundulus heteroclitus по всему устью реки Элизабет, специфичные для соединений и смесей». Экологические науки и технологии. 47 (18): 10556–10566. Дои:10.1021 / es401604b. ЧВК 4079253. PMID 24003986.
- ^ Уэлш, Дженнифер (17 февраля 2011 г.). «Рыба эволюционировала, чтобы выживать в токсинах ГМ в реке Гудзон». LiveScience. Получено 19 февраля 2011.
- ^ а б c Исаак Вирджин; и другие. (2011). «Механические основы устойчивости к ПХБ в Атлантическом Томкоде из реки Гудзон». Наука. 331 (6022): 1322–1325. Bibcode:2011Научный ... 331.1322W. Дои:10.1126 / science.1197296. ЧВК 3246799. PMID 21330491.
- ^ Cheptou, P .; Carrue, O .; Rouifed, S .; Кантарел, А. (2008). «Быстрая эволюция распространения семян в городской среде среди сорняков Crepis sancta». Труды Национальной академии наук. 105 (10): 3796–9. Bibcode:2008PNAS..105.3796C. Дои:10.1073 / pnas.0708446105. ЧВК 2268839. PMID 18316722.
- ^ «Эволюция в городских джунглях». Архивировано из оригинал 8 ноября 2012 г.. Получено 8 июля 2010.
- ^ «Человеческая деятельность увеличивает размер мозга у животных». Йельский научный. 21 декабря 2013 г.. Получено 16 марта 2015.
- ^ Snell-Rood, Emilie C .; Вик, Наоми (2013). «Антропогенная среда оказывает переменный отбор на емкость черепа у млекопитающих». Труды Королевского общества B: биологические науки. 280 (1769): 20131384. Дои:10.1098 / rspb.2013.1384. ЧВК 3768300. PMID 23966638.
- ^ а б c Розенблюм, Эрика Бри (2006), «Конвергентная эволюция и дивергентный отбор: ящерицы в экотоне Белых песков», Американский натуралист, 167 (1): 1–15, Дои:10.1086/498397, PMID 16475095, S2CID 10765531
- ^ Розенблюм, Эрика Бри (2007), «Мультилокусная перспектива колонизации, сопровождаемая отбором и потоком генов», Эволюция, 61 (12): 2971–2985, Дои:10.1111 / j.1558-5646.2007.00251.x, PMID 17976180, S2CID 2322067
- ^ а б c Де Роше, Симона (2014), «Помимо черного и белого: расходящееся поведение и производительность у трех быстро развивающихся видов ящериц в Белых Песках», Биологический журнал Линнеевского общества, 111: 169–182, Дои:10.1111 / bij.12165
- ^ а б c Робертсон, Жанна Мари (2009), «Быстрое расхождение окраски социальных сигналов в экотоне Белых песков для трех видов ящериц в условиях сильного естественного отбора», Биологический журнал Линнеевского общества, 98 (2): 243–255, Дои:10.1111 / j.1095-8312.2009.01291.x
- ^ Шемске, Дуглас В. (2000), «Понимание происхождения видов», Эволюция, 54 (3): 1069–1073, Дои:10.1554 / 0014-3820 (2000) 054 [1069: utoos] 2.3.co; 2
- ^ Маллет, Джеймс (2001), «Революция видообразования», Журнал эволюционной биологии, 14 (6): 887–888, Дои:10.1046 / j.1420-9101.2001.00342.x, S2CID 36627140
- ^ Койн, Джерри А .; Орр, Х. Аллен (2004), Видообразование, Sinauer Associates, стр. 37, ISBN 978-0-87893-089-0
- ^ Флорин, А.-Б .; А. Один (2002), «Лабораторная среда не способствует аллопатрическому видообразованию», Журнал эволюционной биологии, 15: 10–19, Дои:10.1046 / j.1420-9101.2002.00356.x, S2CID 85410953
- ^ Койн, Джерри А .; Орр, Х. Аллен (2004), Видообразование, Sinauer Associates, стр. 88–89, ISBN 978-0-87893-089-0
- ^ Либхерр, Джеймс К .; МакХью, Джозеф В. в Реш, В. Х .; Карде, Р. Т. (редакторы) 2003. Энциклопедия насекомых. Академическая пресса.
- ^ а б c d е ж Бентон, Майкл Дж .; Пирсон, Пол Н. (2001), "Видообразование в летописи окаменелостей", Тенденции в экологии и эволюции, 16 (7): 405–411, Дои:10.1016 / S0169-5347 (01) 02149-8, PMID 11403874
- ^ Эрвин, Д. Х. и Ансти, Р. Л. (1995), Новые подходы к видообразованию в летописи окаменелостей, Columbia University Press, стр. 22
- ^ Лазарь, Давид; и другие. (1995), "Симпатрическое видообразование и филетические изменения в Globorotalia truncatulinoides", Палеобиология, 21 (1): 28–51, Дои:10.1017 / s0094837300013063
- ^ Kellogg, Davida E .; Хейс, Джеймс Д. (1975), "Микроэволюционные закономерности в позднекайнозойских радиоляриях", Палеобиология, 1 (2): 150–160, Дои:10.1017 / s0094837300002347
- ^ Хейс, Джеймс Д. (1970), "Стратиграфия и эволюционные тенденции радиолярий в глубоководных отложениях северной части Тихого океана", Мемуары Геологического общества Америки, 126: 185–218, Дои:10.1130 / MEM126-p185, ISBN 978-0-8137-1126-3
- ^ Ульф Сёрханнус; и другие. (1998), «Кладогенетические и анагенетические изменения морфологии Rhizosolenia praebergonii Мухина ", Историческая биология, 1 (3): 185–205, Дои:10.1080/08912968809386474
- ^ Сёрханнус, Ульф; и другие. (1991), «Итеративная эволюция в роде диатомовых. Ризосоления Эренберг », Lethaia, 24 (1): 39–44, Дои:10.1111 / j.1502-3931.1991.tb01178.x
- ^ Пирсон, Пол Н .; Эзард, Томас Х. Г. (2014). «Эволюция и видообразование в эоценовых планктонных фораминиферах Turborotalia» (PDF). Палеобиология. 40: 130–143. Дои:10.1666/13004. S2CID 5625312.
- ^ Гингерич, П. Д. (1985), «Виды в летописи окаменелостей: концепции, тенденции и переходы», Палеобиология, 11: 27–41, Дои:10.1017 / s0094837300011374
- ^ Гингерич П. Д. (1976), "Палеонтология и филогения: закономерности эволюции на уровне видов у ранних третичных млекопитающих", Американский журнал науки, 276 (1): 1–28, Bibcode:1976AmJS..276 .... 1G, Дои:10.2475 / ajs.276.1.1
- ^ Chaline, J .; и другие. (1993), «Морфологические тенденции и скорость эволюции арвиколид (Arvicolidae, Rodentia): к модели прерывистого равновесия / неравновесия», Четвертичный международный, 19: 27–39, Bibcode:1993QuInt..19 ... 27C, Дои:10.1016 / 1040-6182 (93) 90019-в
- ^ Райс, Уильям Р .; Соль, Джордж У. (1990). «Эволюция репродуктивной изоляции как коррелированного характера в условиях симпатрии: экспериментальные данные». Эволюция, Общество изучения эволюции. 44 (5): 1140–1152. Дои:10.1111 / j.1558-5646.1990.tb05221.x. PMID 28563894. S2CID 9145271.
- ^ «Эволюция репродуктивной изоляции как взаимосвязанного характера в условиях симпатрии: экспериментальные данные» (PDF). Уильям Р. Райс, Джордж У. Солт. Архивировано из оригинал (PDF) 13 мая 2012 г.. Получено 23 мая 2010.
- ^ «Наблюдаемые экземпляры видообразования, 5.3.5 Симпатрическое видообразование у Drosophila melanogaster». Джозеф Боксхорн. Получено 23 мая 2010.
- ^ Скотт П. Иган; и другие. (2012), «Дивергентное использование растений-хозяев способствует репродуктивной изоляции среди популяций цинипидных галлов», Письма о биологии, 8 (4): 605–608, Дои:10.1098 / рсбл.2011.1205, ЧВК 3391443, PMID 22337505
- ^ Egan, Scott P .; и другие. (2013), "Параллельные паттерны морфологических и поведенческих изменений среди ассоциированных с хозяином популяций двух видов галловых ос", PLOS ONE, 8 (1): e54690, Bibcode:2013PLoSO ... 854690E, Дои:10.1371 / journal.pone.0054690, ЧВК 3549985, PMID 23349952
- ^ Feder, J.L .; Roethele, J.B .; Фильчак, К .; Niedbalski, J .; Ромеро-Северсон, Дж. (1 марта 2003 г.). «Доказательства инверсионного полиморфизма, связанного с формированием симпатрической расы хозяев у яблонной личинки мухи, Rhagoletis pomonella». Генетика. 163 (3): 939–53. ЧВК 1462491. PMID 12663534.
- ^ Berlocher, S.H .; Буш, Г.Л. (1982). «Электрофоретический анализ филогении Rhagoletis (Diptera: Tephritidae)». Систематическая зоология. 31 (2): 136–55. Дои:10.2307/2413033. JSTOR 2413033.
- ^ Berlocher, S.H .; Федер, Дж. Л. (2002). «Симпатрическое видообразование у насекомых-фитофагов: выход за пределы споров?». Анну Рев Энтомол. 47: 773–815. Дои:10.1146 / annurev.ento.47.091201.145312. PMID 11729091. S2CID 9677456.
- ^ Буш, Г.Л. (1969). «Формирование симпатрической расы хозяев и видообразование у плодоядных мух рода Rhagoletis (Diptera: Tephritidae)». Эволюция. 23 (2): 237–51. Дои:10.2307/2406788. JSTOR 2406788. PMID 28562891.
- ^ Prokopy, R.J .; Diehl, S.R .; Кули, С.С. (1988). «Поведенческие данные для рас-хозяев мух Rhagoletis pomonella» (PDF). Oecologia. 76 (1): 138–47. Bibcode:1988Oecol..76..138P. Дои:10.1007 / BF00379612. HDL:2027.42/47773. JSTOR 4218647. PMID 28312391. S2CID 19076040.
- ^ Feder, J.L .; Roethele, J.B .; Wlazlo, B .; Берлоше, С. (1997). «Селективное поддержание аллозимных различий между симпатрическими хозяевами яблонной мухи». Proc Natl Acad Sci USA. 94 (21): 11417–21. Bibcode:1997PNAS ... 9411417F. Дои:10.1073 / пнас.94.21.11417. ЧВК 23485. PMID 11038585.
- ^ а б c «Лондонский подземный источник новых форм насекомых». Времена. 26 августа 1998 г.
- ^ а б Fonseca, D.M .; Keyghobadi, N; Малькольм, Калифорния; Мехмет, К; Шаффнер, Ф; Моги, М; Fleischer, RC; Вилкерсон, Р. К. (2004). «Новые векторы в комплексе Culex pipiens» (PDF). Наука. 303 (5663): 1535–8. Bibcode:2004Наука ... 303.1535F. Дои:10.1126 / science.1094247. PMID 15001783. S2CID 40158868. Архивировано из оригинал (PDF) 23 июля 2011 г.
- ^ а б Бурдик, Алан (2001). «Насекомое из метро - Лондон, Англия. Подземный дом для различных видов комаров». Естественная история.
- ^ Byrne, K .; Николс, Р.А. (1999). «Culex pipiens в лондонских подземных туннелях: различие между наземными и подземными популяциями». Наследственность. 82 (1): 7–15. Дои:10.1038 / sj.hdy.6884120. PMID 10200079.
- ^ Виноградова, Е.Б .; Шайкевич, Э. (2007). «Морфометрические, физиологические и молекулярные характеристики подземных популяций городского комара Culex pipiens Linnaeus f. Molestus Forskål (Diptera: Culicidae) из нескольких регионов России» (PDF). Европейский вестник комаров. 22: 17–24.
- ^ а б c Больно, Карла; и другие. (2008), «Мультилокусный тест одновременной дивергенции через Панамский перешеек с использованием щелкающих креветок из рода Alpheus», Эволюция, 63 (2): 514–530, Дои:10.1111 / j.1558-5646.2008.00566.x, PMID 19154357, S2CID 11820649
- ^ К. Монтес; и другие. (2015), «Среднемиоценовое закрытие Центральноамериканского морского пути», Наука, 348 (6231): 226–229, Bibcode:2015Научный ... 348..226М, Дои:10.1126 / science.aaa2815, PMID 25859042
- ^ Ноултон, Нэнси (1993), "Дивергенция в белках, митохондриальной ДНК и репродуктивной совместимости через Панамский перешеек", Наука, 260 (5114): 1629–1632, Bibcode:1993Наука ... 260.1629K, Дои:10.1126 / science.8503007, PMID 8503007, S2CID 31875676
- ^ а б Ноултон, Нэнси; Вейгт, Ли А. (1998), «Новые даты и новые показатели расхождения через Панамский перешеек», Proc. R. Soc. Лондон. B, 265 (1412): 2257–2263, Дои:10.1098 / rspb.1998.0568, ЧВК 1689526
- ^ а б c Rieseberg, Loren H .; Вендел, Джонатан (2004), «Вид растений - Восстание бедных кузенов», Новый Фитолог, 161: 1–21, Дои:10.1111 / j.1469-8137.2004.00957.x
- ^ а б Лафон-Пласетт, Клеман; и другие. (2016), «Текущие исследования видообразования растений: раскрытие процессов и механизмов, лежащих в основе эволюции барьеров репродуктивной изоляции», Новый Фитолог, 209 (1): 29–33, Дои:10.1111 / nph.13756, PMID 26625345
- ^ Анакер, Брайан Л .; Штраус, Шарон Ю. (2014), «География и экология видов растений: перекрытие ареалов и дивергенция ниш у родственных видов», Proc. R. Soc. B, 281 (1778): 20132980, Дои:10.1098 / rspb.2013.2980, ЧВК 3906944, PMID 24452025
- ^ Хегарти, Мэтью Дж .; Хискок, Саймон Дж. (2004), "Гибридное видообразование у растений: новые идеи молекулярных исследований", Новый Фитолог, 165 (2): 411–423, Дои:10.1111 / j.1469-8137.2004.01253.x, PMID 15720652, S2CID 29320650
- ^ Вендел, Джонатан Ф. (январь 2000 г.). «Эволюция генома полиплоидов». Молекулярная биология растений. 42 (1): 225–249. Дои:10.1023 / А: 1006392424384. ISSN 0167-4412. PMID 10688139. S2CID 14856314.
- ^ Семон, Мари; Вулф, Кеннет Х. (декабрь 2007 г.). «Последствия дупликации генома». Текущее мнение в области генетики и развития. 17 (6): 505–512. Дои:10.1016 / j.gde.2007.09.007. ISSN 0959-437X. PMID 18006297.
- ^ а б Комай, Лука (ноябрь 2005 г.). «Преимущества и недостатки полиплоидности». Природа Обзоры Генетика. 6 (11): 836–846. Дои:10.1038 / nrg1711. ISSN 1471-0056. PMID 16304599. S2CID 3329282.
- ^ Солтис, Памела С.; Солтис, Дуглас Э. (20 июня 2000 г.). «Роль генетических и геномных атрибутов в успехе полиплоидов». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 97 (13): 7051–7057. Bibcode:2000PNAS ... 97.7051S. Дои:10.1073 / pnas.97.13.7051. ISSN 0027-8424. ЧВК 34383. PMID 10860970.
- ^ Маварес, Хесус; Салазар, Камило А .; Бермингем, Элдридж; и другие. (15 июня 2006 г.). «Видообразование путем гибридизации в Геликоний бабочки ». Природа. 441 (7095): 868–871. Bibcode:2006Натура.441..868М. Дои:10.1038 / природа04738. ISSN 0028-0836. PMID 16778888. S2CID 2457445.
- ^ Рэмси, Джастин; Шемске, Дуглас В. (ноябрь 1998 г.). «Пути, механизмы и скорость образования полиплоидов у цветковых растений». Ежегодный обзор экологии и систематики. 29: 467–501. Дои:10.1146 / annurev.ecolsys.29.1.467. ISSN 1545-2069. S2CID 31637733.
- ^ Отто, Сара П.; Уиттон, Жаннетт (декабрь 2000 г.). «Заболеваемость и эволюция полиплоидов». Ежегодный обзор генетики. 34: 401–437. CiteSeerX 10.1.1.323.1059. Дои:10.1146 / annurev.genet.34.1.401. ISSN 0066-4197. PMID 11092833.
- ^ а б Эрик Баак; и другие. (2015), «Истоки репродуктивной изоляции у растений», Новый Фитолог, 207 (4): 968–984, Дои:10.1111 / nph.13424, PMID 25944305, S2CID 30147815
- ^ Вальехо-Марин, Марио (2012). «Mimulus peregrinus (Phrymaceae): новый британский аллополиплоидный вид». Фитоключи. 14 (14): 1–14. Дои:10.3897 / phytokeys.14.3305. ЧВК 3492922. PMID 23170069.
- ^ Вальехо-Марин, Марио; Баггс, Ричард Дж .; Кули, Ариэль М .; Пузи, Джошуа Р. (2015). «Видообразование путем дупликации генома: повторяющееся происхождение и геномный состав недавно сформировавшегося аллополиплоидного вида Mimulus peregrinus». Эволюция. 69 (6): 1487–1500. Дои:10.1111 / evo.12678. ЧВК 5033005. PMID 25929999.
- ^ Карпеченко, Г. (1927). «Полиплоидные гибриды Raphanus sativus Икс Brassica oleracea L ". Бык. Appl. Бот. 17: 305–408.
- ^ Терасава, Y (1933). "Переход между Брассико-рафанус и B. chinensis и Raphanus sativus". Японский журнал генетики. 8 (4): 229–230. Дои:10.1266 / jjg.8.229.
- ^ Лоу, Эндрю Дж .; Эбботт, Ричард Дж. (1996). "Происхождение новых видов аллополиплоидов Senecio camrensis (сложноцветный) и его связь с эндемиками Канарских островов Senecio tenerifae". Американский журнал ботаники. 83 (10): 1365–1372. Дои:10.2307/2446125. JSTOR 2446125.
- ^ Койн, Джерри А. (2009). Почему эволюция верна. Группа пингвинов. стр.187–189. ISBN 978-0-670-02053-9.
- ^ Ботанический сад Миссури. "Веб-дисплей TROPICOS Senecio vulgaris L ". База номенклатурных и образцовых данных. Государственная библиотека Миссури. Архивировано из оригинал 2 августа 2013 г.. Получено 1 февраля 2008.
- ^ а б Бомблис, Кирстен; Лемпе, Янне; Эппле, Петра; Вартманн, Норман; Ланц, Криста; Дангл, Джеффри Л .; Вайгель, Детлеф (2007). «Аутоиммунный ответ как механизм синдрома несовместимости типа Добжанского-Мюллера у растений». ПЛОС Биол. 5 (9): e236. Дои:10.1371 / journal.pbio.0050236. ЧВК 1964774. PMID 17803357.
- ^ «Новые виды растений возникают из-за конфликтов между генами иммунной системы». Эд Йонг. Архивировано из оригинал 3 февраля 2010 г.. Получено 22 мая 2010.
- ^ Первес, Уильям Кирквуд; Садава, Дэвид Э .; Orians, Gordon H .; Хеллер, Х. Крейг (2006). Жизнь, наука биология (7-е изд.). Sinaur Associates, Inc. стр. 487. ISBN 978-0-7167-9856-9.
- ^ Солтис, Пэм (17 марта 2011 г.). "Исследователь УФ: исследование цветущих растений" фиксирует эволюцию в действии'". EurekAlert, Американская ассоциация развития науки. Получено 28 марта 2011.
- ^ Buggs, Ричард Дж. А .; Чжан, Линьцзин; Майлз, Николас; Тейт, Дженнифер А .; Гао, Лу; Вэй, Ву; Schnable, Patrick S .; Барбазук, В. Брэд; Солтис, Памела С. (2011). «Транскриптомный шок порождает эволюционную новизну в недавно созданном природном аллополиплоидном заводе». Текущая биология. 21 (7): 551–6. Дои:10.1016 / j.cub.2011.02.016. PMID 21419627. S2CID 18417406.
- ^ Bearhop, S .; Fiedler, W .; Фернесс, R.W .; Votier, S.C .; Waldron, S .; Newton, J .; Bowen, G.J .; Berthold, P .; Фарнсворт, К. (2005). «Ассортативное спаривание как механизм быстрой эволюции миграционной пропасти». Наука. 310 (5747): 502–504. Bibcode:2005Наука ... 310..502B. Дои:10.1126 / science.1115661. PMID 16239479. S2CID 6415789. Вспомогательные онлайн-материалы
- ^ Йонг, Эд (3 декабря 2009 г.). «Британские кормильцы разделили черных шапок на две генетически разные группы». НаукаБлоги. Получено 21 мая 2010.
- ^ Тоблер, Майкл (2009). «Способствует ли хищное насекомое расхождению между популяциями рыб, приспособленных к пещерам и к поверхности?». Письма о биологии. 5 (4): 506–509. Дои:10.1098 / rsbl.2009.0272. ЧВК 2781934. PMID 19443506.
- ^ «Гигантское насекомое разделяет пещерных рыб на отдельные популяции». Эд Йонг. Архивировано из оригинал 1 февраля 2010 г.. Получено 22 мая 2010.
- ^ Ж. П. Дупе; Дж. Х. Инглэнд; М. Фурзе; Д. Паэткау (2007 г.), «Наблюдение медведя гризли (Ursus arctos) в Канаде на самом севере: фотографические данные и данные ДНК с острова Мелвилл, Северо-Западные территории», Арктический, 60: 271–276
- ^ Грей, А. П. (1972), Гибриды млекопитающих. Контрольный список с библиографией (2-е изд.), Farnham Royal, Slough SL2 3BN, England: Сельскохозяйственные бюро Содружества
- ^ Кучера, Верена Э; Бидон, Тобиас; Хейлер, Фрэнк; Роди, Юлия Л .; Файн, Стивен Р .; Янке, Аксель (2014). «Медведи в лесу генных деревьев: филогенетический вывод усложнен неполной сортировкой по происхождению и потоком генов». Молекулярная биология и эволюция. 31 (8): 2004–2017. Дои:10.1093 / molbev / msu186. ЧВК 4104321. PMID 24903145.
- ^ а б Lindqvist, C .; Schuster, S.C .; Sun, Y .; Talbot, S.L .; Qi, J .; Ratan, A .; Tomsho, L.P .; Kasson, L .; Zeyl, E .; Aars, J .; Miller, W .; Ingolfsson, O .; Бахманн, Л .; Уиг, О. (2010). «Полный митохондриальный геном челюстной кости плейстоцена раскрывает происхождение белого медведя». Труды Национальной академии наук. 107 (11): 5053–7. Bibcode:2010PNAS..107.5053L. Дои:10.1073 / pnas.0914266107. ЧВК 2841953. PMID 20194737.
- ^ Hailer, F .; Кучера, В. Э .; Hallstrom, B.M .; Klassert, D .; Fain, S. R .; Леонард, Дж. А .; Arnason, U .; Янке, А. (2012). «Ядерные геномные последовательности показывают, что белые медведи - древняя и отчетливая медвежья линия» (PDF). Наука. 336 (6079): 344–7. Bibcode:2012Наука ... 336..344H. Дои:10.1126 / наука.1216424. HDL:10261/58578. PMID 22517859. S2CID 12671275.
- ^ Liu S .; и другие. (2014), «Популяционная геномика выявляет недавнее видообразование и быструю эволюционную адаптацию белых медведей», Ячейка, 157 (4): 785–794, Дои:10.1016 / j.cell.2014.03.054, ЧВК 4089990, PMID 24813606
- ^ Миллер, Вт; Schuster, SC; Уэлч, AJ; Ратан, А; Бедоя-Рейна, ОС; Чжао, Ф; Kim, HL; Burhans, RC; Драуц, Д.И.; Виттекиндт, NE; Томшо, LP; Ибарра-Лаклетт, Э; Эррера-Эстрелла, L; Павлин, E; Фарли, S; Sage, GK; Роде, К; Оббард, М; Montiel, R; Бахманн, Л; Ingólfsson, O; Aars, J; Майлунд, Т; Wiig, O; Talbot, SL; Линдквист, C (2012). «Геномы белых и бурых медведей выявляют древние примеси и демографические следы прошлых изменений климата». Труды Национальной академии наук Соединенных Штатов Америки. 109 (36): E2382 – E2390. Bibcode:2012PNAS..109E2382M. Дои:10.1073 / pnas.1210506109. ЧВК 3437856. PMID 22826254.
- ^ «Эволюция». Международные полярные медведи. Получено 30 января 2014.
- ^ Хансман, Джаред (6 августа 2008 г.). «Адаптивные черты белого медведя (Ursus maritimus)». Helium, Inc. Получено 30 января 2014.
- ^ а б Тикелл, У. Л. Н. (март 2003 г.). «Белое оперение». Водно-болотные птицы: Международный журнал биологии водоплавающих птиц. 26 (1): 1–12. Дои:10.1675 / 1524-4695 (2003) 026 [0001: wp] 2.0.co; 2. JSTOR 1522461.
- ^ Ларсон, Эдвард Дж. (2004). Эволюция: замечательная история научной теории. Нью-Йорк: Современная библиотека. стр.121–123, 152–157. ISBN 978-0-679-64288-6.
- ^ Боулер, Питер Дж. (1983). Затмение дарвинизма: антидарвиновские эволюционные теории в период около 1900 года. Издательство Университета Джона Хопкинса. С. 29, 250. ISBN 978-0-8018-4391-4.
- ^ а б Котт, Хью Б. (1940). Адаптивная окраска животных. Метуэн. стр.68 –72.
- ^ а б Cook, L.M .; Grant, B.S .; Saccheri, I.J .; Маллет, Джеймс (2012). «Избирательное истребление пернатой мотылька птицами: последний эксперимент Майкла Майеруса». Письма о биологии. 8 (4): 609–612. Дои:10.1098 / рсбл.2011.1136. ЧВК 3391436. PMID 22319093.
- ^ Уоллес, Альфред Рассел (2015) [1889]. Дарвинизм - изложение теории естественного отбора - с некоторыми ее приложениями. Читать книги. п. 180. ISBN 978-1-4733-7510-9.
- ^ Поултон, Эдвард Бэгнолл (1890). Цвета животных. п. Разверните после страницы 339 и повсюду.
- ^ Моделируемая эволюция становится сложной. Trnmag.com (2008-05-08). Проверено 6 декабря 2011.
- ^ Adami, C .; Ofria, C .; Кольер, Т. (2000). «Эволюция биологической сложности». Proc Natl Acad Sci USA. 97 (9): 4463–8. arXiv:физика / 0005074. Bibcode:2000PNAS ... 97.4463A. Дои:10.1073 / pnas.97.9.4463. ЧВК 18257. PMID 10781045.
- ^ Earl, D.J .; Дим, М.В. (2004). «Эволюционируемость - это выбираемая черта». Proc Natl Acad Sci USA. 101 (32): 11531–6. arXiv:q-bio / 0407012. Bibcode:2004PNAS..10111531E. Дои:10.1073 / pnas.0404656101. ЧВК 511006. PMID 15289608.
- ^ а б Стеммер, В. (1994). «Перетасовка ДНК путем случайной фрагментации и повторной сборки: рекомбинация in vitro для молекулярной эволюции». Proc Natl Acad Sci USA. 91 (22): 10747–51. Bibcode:1994PNAS ... 9110747S. Дои:10.1073 / пнас.91.22.10747. ЧВК 45099. PMID 7938023.
- ^ Заутер, Э. (27 марта 2006 г.). """Ускоренная эволюция" превращает фермент РНК в фермент ДНК In vitro". TSRI - Новости и обзоры. 6 (11).
- ^ Молекулярная эволюция. kaist.ac.kr
- ^ Молекулярная эволюция in vitro. Isgec.org (1975-08-04). Проверено 6 декабря 2011.
- ^ Освальд, Том (18 июля 2001 г.). «Цифровые организмы, используемые для подтверждения эволюционного процесса». EurekAlert! Новости науки. Американская ассоциация развития науки.
- ^ Дибас, Шерил (7 мая 2003 г.). «Эксперименты с искусственной жизнью показывают, как могут развиваться сложные функции». EurekAlert! Новости науки. Американская ассоциация развития науки.
- ^ Clune, J; Голдсби, HJ; Ofria, C; Пеннок, RT (2011). «Выборочное давление для точного нацеливания на альтруизм: данные цифровой эволюции для трудных для проверки аспектов теории инклюзивной приспособленности» (PDF). Труды Королевского общества. Королевское общество. 278. Архивировано из оригинал (PDF) 7 декабря 2011 г.
Источники
- Биологическая наука. Оксфорд. 2002 г.
- Клегг, С.Дж. (1998). Генетика и эволюция. Лондон: Дж. Мюррей. ISBN 978-0-7195-7552-5.
- Койн, Джерри А. (2009). Почему эволюция верна. Нью-Йорк: Издательство Оксфордского университета. ISBN 978-0-19-923084-6.
- Дарвин, Чарльз 24 ноября 1859 г. На Происхождение видов посредством естественного отбора или сохранения благоприятных рас в борьбе за жизнь. Лондон: Джон Мюррей, Albemarle Street. 502 страницы. Перепечатано: Gramercy (22 мая 1995 г.). ISBN 0-517-12320-7
- Докинз, Ричард (2009). Величайшее шоу на Земле: свидетельства эволюции. Bantam Press. ISBN 978-1-4165-9478-9.
- Эндлер, Джон А. (1986). Естественный отбор в дикой природе. Нью-Джерси: Издательство Принстонского университета. ISBN 978-0-691-08387-2.
- Фитцхью, Кирк (2006). «-1-« Свидетельства »эволюции против« свидетельств »разумного замысла: параллельные заблуждения». Музей естественной истории округа Лос-Анджелес.
- Грант, Питер Р .; Грант, Б. Розмари (2014). 40 лет эволюции: зяблики Дарвина на острове Дафна Мейджор. Издательство Принстонского университета. п. 432. ISBN 978-0691160467.
- Hill, A .; Беренсмейер, А. К. (1980). Окаменелости в процессе становления: тафономия позвоночных и палеоэкология. Чикаго: Издательство Чикагского университета. ISBN 978-0-226-04169-8.
- Клюге, Арнольд Г. (1999). «Наука филогенетической систематики: объяснение, предсказание и проверка». Кладистика. 15 (4): 429–436. Дои:10.1111 / j.1096-0031.1999.tb00279.x. HDL:2027.42/73926. S2CID 6034651.
- Лорен, Мишель (2000). «Ранняя эволюция четвероногих» (PDF). Дерево. 15 (3): 118–123. Дои:10.1016 / s0169-5347 (99) 01780-2. PMID 10675932. Архивировано из оригинал (PDF) 22 июля 2012 г.. Получено 10 ноября 2017.
- Майр, Эрнст (2001). Что такое эволюция. Нью-Йорк: Основные книги. ISBN 978-0-465-04426-9.
- Paul, C.R.C .; Донован, С. (1998). Адекватность летописи окаменелостей. Нью-Йорк: Джон Вили. ISBN 978-0-471-96988-4.
- Протеро, Дональд (2007). Эволюция: что говорят окаменелости и почему это важно. Издательство Колумбийского университета. ISBN 978-0231139625.
- Шубин, Нил (2008). Ваша внутренняя рыба: путешествие в 3,5 миллиарда лет истории человеческого тела. Случайный дом. ISBN 978-0-375-42447-2.
- Собер, Эллиотт (2008). Доказательства и эволюция: логика науки. Издательство Кембриджского университета. ISBN 978-0-521-87188-4.
внешние ссылки
- Ресурсы по развитию национальных академий
- Архив TalkOrigins – 29+ свидетельств макроэволюции: научное обоснование общего происхождения
- Архив TalkOrigins – Часто задаваемые вопросы об окаменелостях переходных позвоночных
- Понимание эволюции: ваш универсальный источник информации об эволюции
- Национальная академия прессы: учение об эволюции и природе науки
- Эволюция - Предоставлено PBS.
- Новости эволюции от сети новостей генома (GNN)
- Эволюция путем естественного отбора - Введение в логику теории эволюции путем естественного отбора
- Howstuffworks.com - Как работает эволюция
- 15 самоцветов эволюции
