E. coli эксперимент долгосрочной эволюции - E. coli long-term evolution experiment

В Кишечная палочка долгосрочный эволюционный эксперимент (LTEE) - это постоянное исследование в экспериментальная эволюция во главе с Ричард Ленски который отслеживал генетические изменения в 12 изначально идентичных популяциях бесполых кишечная палочка бактерии с 24 февраля 1988 г.[2] Популяции достигли рубежа в 50 000 поколений в феврале 2010 года.[Обновить].[3] Ленский исполнил 10-тысячный передача эксперимента 13 марта 2017 г.[4] Население достигло 73,500 поколений в начале 2020 года, незадолго до того, как оно было заморожено из-за пандемии COVID-19.[5][6]
В ходе эксперимента Ленски и его коллеги сообщили о широком спектре фенотипических и генотипических изменений в развивающихся популяциях. В их число вошли изменения, произошедшие во всех 12 популяциях, и другие, которые проявились только в одной или нескольких популяциях. Например, во всех 12 популяциях наблюдалась схожая картина быстрого улучшения приспособленности, которое со временем замедлялось, скорости роста и увеличения размера клеток. У половины популяций развились дефекты репарации ДНК, которые вызвали мутаторные фенотипы, отмеченные повышенной частотой мутаций. Самая поразительная адаптация, о которой сообщалось до сих пор, - это эволюция аэробного роста на цитрате, что необычно для Кишечная палочка, в одной популяции в какой-то момент между поколениями от 31 000 до 31 500.[7][8]
Ленский принял решение приостановить эксперимент 9 марта 2020 года, когда его лаборатория временно закрылась в качестве меры предосторожности против распространения вируса. COVID-19. Все линии длительного хранения были помещены в хранилище со сверхнизкими температурами до тех пор, пока их нельзя будет разморозить для продолжения эксперимента.[9]
4 мая 2020 года Ленский объявил о 5-летнем продлении гранта через Программа долгосрочных исследований в области экологической биологии Национального научного фонда (LTREB) который поддерживает LTEE. Он также объявил, что эксперимент будет передан под наблюдение доктора Джеффри Э. Баррика, доцента кафедры молекулярных биологических наук Техасского университета в Остине, в течение следующих 5 лет.[10] Доктор Баррик ранее был исследователем с докторской степенью у доктора Ленски и внес большой вклад в исследования, основанные на LTEE.[10]
Экспериментальный подход
Долгосрочный эволюционный эксперимент был разработан как открытое средство эмпирического исследования центральных характеристик эволюция. Эксперимент был начат с тремя основными целями:
- Изучить динамику эволюции, включая скорость эволюционных изменений.
- Исследовать повторяемость эволюции.
- Чтобы лучше понять взаимосвязь между изменениями на фенотипический и генотипический уровни.[11]
По мере продолжения эксперимента его масштабы росли по мере того, как возникали новые вопросы в эволюционной биологии, которые можно было использовать для решения, по мере того, как эволюция популяций представляла новые явления для изучения, а также по мере развития технологий и методологических методов.[12]
Использование Кишечная палочка поскольку экспериментальный организм позволил изучить многие поколения и большие популяции за относительно короткий период времени. К тому же из-за длительного использования Кишечная палочка в качестве основной модели организма в молекулярная биология был доступен широкий спектр инструментов, протоколов и процедур для изучения изменений на генетическом, фенотипическом и физиологическом уровнях.[13] Бактерии также можно заморозить и сохранить, оставаясь при этом жизнеспособными. Это позволило создать то, что Ленски называет «замороженной летописью окаменелостей» образцов эволюционирующих популяций, которые могут быть восстановлены в любое время. Эта замороженная летопись окаменелостей позволяет перезапускать популяции в случаях заражения или других нарушений в эксперименте, а также позволяет изолировать и сравнивать живые образцы предков и эволюционировавших клонов. Ленский выбрал Кишечная палочка штамм, который воспроизводит только бесполым путем, не хватает плазмиды это может позволить бактериальная конъюгация, и не имеет жизнеспособных профаг. Как следствие, эволюция в эксперименте происходит только за счет основных эволюционных процессов мутация, генетический дрейф, и естественный отбор. Эта строгая асексуальность также означает, что генетические маркеры сохраняться в родословных и кладах общее происхождение, но иначе не может распространиться среди населения.[11]
Ленски решил провести эксперимент с бактериями, выращенными в минимальной среде с ограничением глюкозы под названием DM25,[14] который изначально был разработан Бернард Дэвис для использования в изоляции ауксотрофный мутанты Кишечная палочка с помощью пенициллин как селективный агент.[15][16] DM25 содержит низкую концентрацию глюкозы.[14] Ленский выбрал эту концентрацию, чтобы упростить анализ эволюции популяций за счет сокращения клональное вмешательство, в котором несколько версий аллели конкурируют в эволюционирующей популяции, уменьшая при этом возможность эволюции экологических взаимодействий.[11] Эта концентрация используемой глюкозы поддерживает максимальную популяцию в 500 миллионов клеток предка в 10 мл культуры, хотя теперь максимум варьируется среди эволюционировавших популяций.[15] DM25 также содержит большое количество цитрата (примерно в 11 раз больше, чем концентрация глюкозы), который изначально был включен Дэвисом, поскольку он улучшал эффективность уничтожения пенициллин во время его экспериментов, хотя теперь известно, что он помогает в E. coli 's получение железа из среды.[15][17]
Методы
12 популяций содержатся в инкубаторе 37 ° C (99 ° F) в лаборатории Ленски в Университет штата Мичиган. Каждый день 1% каждой популяции переносят в колбу со свежей питательной средой DM25. Разбавление означает, что в каждой популяции ежедневно происходит 6,64 поколения или удвоения. Большие репрезентативные образцы каждой популяции замораживают глицерином в качестве криопротектор каждые 500 поколений (75 дней). Бактерии в этих образцах остаются жизнеспособными и могут быть восстановлены в любое время. Эта коллекция образцов называется «замороженной летописью окаменелостей» и обеспечивает историю эволюции каждой популяции на протяжении всего эксперимента. Население также регулярно проверяется на предмет изменений в средний фитнес, а также регулярно проводятся дополнительные эксперименты для изучения интересных изменений в популяциях.[18] По состоянию на апрель 2016 г.[Обновить], то Кишечная палочка популяции изучались на протяжении более 64 500 поколений и, как считается, претерпели достаточно спонтанные мутации что каждый возможный точечная мутация в Кишечная палочка геном произошел несколько раз.[7]
Основное напряжение
Напряжение Кишечная палочка Ленски выбрал для использования в эксперименте по долгосрочному развитию «штамм Bc251», как описано в статье Сеймура Ледерберга от 1966 года через Брюса Левина, который использовал его в эксперименте по бактериальной экологии в 1972 году. Определяющие генетические особенности этот штамм были: T6р, Ул.р, р−м−, Ара− (не может расти на арабиноза ).[2] Ленски обозначил оригинальный штамм-основатель как REL606. Перед началом эксперимента Ленский выделил ара+ вариант штамма, при котором точечная мутация в ара оперон восстановил рост на арабинозе, которую он обозначил как штамм REL607. Начав долгосрочный эволюционный эксперимент, Ленский основал шесть популяций с шестью отдельными ара− колонии REL606. Эти популяции обозначаются как Ara-1 - Ara-6. Ленский также основал еще шесть популяций из шести особей ара.+ колонии REL607. Их называют популяциями от Ara + 1 до Ara + 6. Маркерные различия позволяют дифференцировать штаммы на пластинах Tetrazolium Arabinose, на которых Ara− колонии выглядят красными, а Ара+ колонии от белого до розового. В ходе эксперимента каждая популяция накопила большое количество различных мутаций, которые позволяют использовать дополнительные средства идентификации штаммов по популяции их происхождения.
Полученные результаты
Изменения в фитнесе
Многое в ходе анализа эксперимента было посвящено тому, как изменилась приспособленность популяций по отношению к их предкам. Все популяции показали образец быстрого увеличения относительной приспособленности в течение ранних поколений, причем это увеличение замедлялось со временем. К 20 000 поколениям популяция росла примерно на 70% быстрее, чем у предков.[11] Это увеличение и замедление роста продолжалось в последующих поколениях. Исследование 2013 Wiser et al. сообщили о постоянном улучшении на 50 000 поколений по сравнению с образцами, выделенными на 40 000 поколений. Они обнаружили, что повышение физической формы соответствует сила закона модель намного лучше, чем гиперболические модели, которые использовались ранее. Поскольку модель степенного закона описывает постоянно замедляющееся увеличение, не имеющее верхнего предела, в то время как гиперболическая модель подразумевает жесткий предел, работа предполагает, что рост будет продолжаться без ограничений, поскольку в популяциях фиксировались все более низкие мутации полезности.[21] Дальнейшая работа, опубликованная в 2015 году, сообщила о результатах более 1100 новых тестов на пригодность, в которых изучались изменения приспособленности на протяжении 60 000 поколений. Данные снова соответствуют предложенной модели степенного закона и, действительно, соответствуют прогнозам модели на основе более ранних данных. Эти результаты предполагают, что, в отличие от предыдущего мышления, адаптация и адаптивное расхождение потенциально могут увеличиваться бесконечно, даже в постоянной среде.[22][23][24]
Эволюция генома
На сегодняшний день сообщается, что из 12 популяций у шести обнаружились дефекты в способности восстановить ДНК, что значительно увеличивает скорость мутаций в этих штаммах.[8][25][26] Хотя считается, что бактерии в каждой популяции произвели сотни миллионов мутаций в течение первых 20000 поколений, Ленски подсчитал, что в течение этого периода времени было получено только от 10 до 20 полезных мутаций. фиксация в каждой популяции менее 100 точечных мутаций (включая нейтральные мутации ) достижение фиксации в каждой популяции.[11] В 2009 году Barrick et al. сообщили о результатах определения последовательностей генома из нескольких временных точек в популяции Ara-1. Они обнаружили, что, в отличие от снижения скорости улучшения приспособленности, накопление мутаций было линейным и напоминало часы, хотя несколько линий свидетельств предполагали, что большая часть накопления была полезной, а не нейтральной.[27]
Эволюция увеличения размера клеток во всех двенадцати популяциях
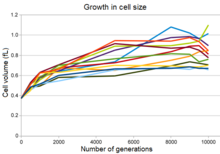
Все двенадцать экспериментальных популяций демонстрируют увеличение размера клеток одновременно с уменьшением максимальной плотности популяции, а во многих популяциях - более округлую форму клеток.[28] Это изменение было частично результатом мутации, изменившей выражение гена для пенициллин-связывающий белок, что позволило мутантным бактериям вытеснить бактерии-предки в условиях длительного эволюционного эксперимента. Однако, хотя эта мутация увеличилась фитнес в этих условиях это также увеличивало чувствительность бактерий к осмотический стресс и уменьшили их способность выживать в течение длительного периода в культурах со стационарной фазой.[28]
Экологическая специализация
В ходе эксперимента популяции эволюционировали и стали специализироваться на том ресурсе глюкозы, на котором они растут. Впервые это было описано в 2000 году, когда Купер и Ленски продемонстрировали, что все популяции испытали распад неиспользуемых метаболических функций после 20 000 поколений, что ограничило диапазон веществ, на которых могли расти бактерии. Их анализ показал, что этот распад был вызван антагонистическая плейотропия, в котором мутации, улучшающие способность расти на глюкозе, снижали или устраняли способность расти на других веществах.[29] Более позднее исследование Лейби и Маркса, в котором использовались более продвинутые методы, показало, что большая часть распада, идентифицированного Купером и Ленски, была экспериментальными артефактами, что потеря неиспользуемых функций была не такой обширной, как предполагалось на первый взгляд, и что некоторые неиспользуемые функции были улучшены. Более того, они пришли к выводу, что метаболические потери были вызваны не антагонистической плейотропией, а нейтральным накоплением мутаций в неиспользуемых частях генома, предполагая, что адаптация к простой среде не обязательно может привести к специализации.[30]
Эволюция сбалансированного полиморфизма и простых экосистем
Два различных варианта, S и L, были идентифицированы в популяции, обозначенной Ara-2, в 18000 поколениях на основании их образования малых и больших колоний соответственно.[31] Клоны типов S и L могли стабильно сосуществовать в совместном культивировании друг с другом, что указывает на то, что они занимали разные ниши в популяции. Это было подтверждено открытием, что L-тип имел преимущество во время роста на глюкозе, но что S имел преимущество во время стационарной фазы после того, как глюкоза закончилась. Было обнаружено, что эти два типа первоначально развились до 6000 поколений, а затем сосуществовали после этого.[31] Филогенетический анализ клонов двух типов, выделенных из разных поколений, показал, что типы S и L принадлежали к разным сосуществующим линиям в популяции и, возможно, подвергались зарождающемуся видообразованию.[32]
Эволюция использования аэробных цитратов в одной популяции
Фон

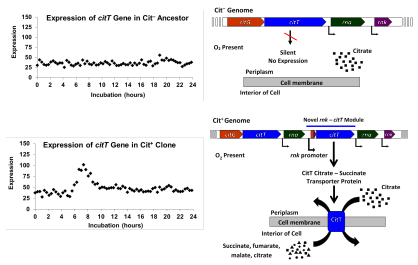
Кишечная палочка обычно не может расти аэробно на цитрат из-за неспособности экспрессировать переносчик цитрата в присутствии кислорода.[34] Тем не мение, Кишечная палочка имеет полный цикл лимонной кислоты, и поэтому метаболизирует цитрат в качестве промежуточного продукта во время аэробного роста других веществ, включая глюкозу. Наиболее Кишечная палочка может расти анаэробно на цитрате через ферментация, если вспомогательный субстрат, такой как глюкоза, доступен для обеспечения восстанавливающей способности.[7][34][35][36] В анаэробный рост возможен за счет экспрессии трансмембранного гена цитрат-сукцинатного антипортера, CITT, который был впервые идентифицирован в 1998 году. Этот ген совместно регулируется с другими генами, участвующими в цитратной ферментации, обнаруженными на cit оперон, который включается только при отсутствии кислорода.[34][37]
Неспособность к аэробному росту на цитрате, называемая Cit.− фенотип, считается определяющей характеристикой Кишечная палочка как вид, который был ценным средством дифференциации Кишечная палочка от патогенных Сальмонелла. Хотя Cit+ штаммы Кишечная палочка были выделены из экологических и сельскохозяйственных образцов, в каждом таком случае было обнаружено, что этот признак связан с присутствием плазмиды, несущей чужеродный переносчик цитрата.[38] Единый, спонтанный Cit+ мутант Кишечная палочка было сообщено Холлом в 1982 году.[39] Этот мутант был выделен во время продолжительной селекции на рост другого нового вещества в питательной среде, которая также содержала цитрат. Генетический анализ Холла показал, что основная мутация была сложной, но в конечном итоге он не смог идентифицировать точные изменения или вовлеченные гены, что привело его к гипотезе об активации загадочного гена-переносчика.[39] Области генома, до которых Холлу удалось сузить локализацию изменений, не соответствуют известному местоположению CITT ген, идентифицированный 16 лет спустя, а также физиологические характеристики в анализах переноса Cit Hall.+ мутанты соответствуют ожидаемым для аэробной экспрессии переносчика CitT.[38][40]
Cit+ развивается в LTEE
В 2008 году команда Ленского во главе с Захари Д. Блаунт, сообщили, что способность к аэробному росту на цитрате развивалась у одной популяции. Примерно в поколении 33 127 наблюдалось резкое увеличение мутности в популяции, обозначенной как Ara-3. Они обнаружили, что популяция содержала клоны, которые могли расти в аэробных условиях на цитрате (Cit+). Эта метаболическая способность позволила популяции вырасти в несколько раз больше, чем раньше, из-за большого количества цитрата, присутствующего в среде. Исследование замороженных образцов ископаемых популяций показало, что Cit+ клоны могли быть выделены уже в 31 500 поколениях. Cit+ было обнаружено, что варианты в популяции обладают рядом генетических маркеров, уникальных для популяции Ara-3; это наблюдение исключало возможность того, что клоны были контаминантами, а не спонтанными мутантами. В серии экспериментов, которые «переиграли» ленту эволюции Ara-3 от Cit− клонов, выделенных из образцов, замороженных в различные моменты истории популяции, они продемонстрировали, что способность к аэробному росту на цитрате с большей вероятностью будет повторно развиваться в подмножестве генетически чистых, эволюционировавших клонов. В этих экспериментах они наблюдали 19 новых независимых экземпляров Cit+ повторная эволюция, но только если исходить из клонов, выделенных после 20,000 поколения. Тесты флуктуации показали, что клоны этого поколения, а затем и более поздние, демонстрировали скорость мутации Cit+ признак, который был значительно выше, чем у предков. Даже в этих более поздних клонах скорость мутации Cit+ было порядка одного появления на триллион клеточных делений.[7]
Ленский и его коллеги пришли к выводу, что эволюция Cit+ функция в этой популяции возникла из-за одной или нескольких более ранних, возможно, неадаптивных, «потенцирующих» мутаций, которые увеличили скорость мутации до доступного уровня. Данные свидетельствуют о том, что использование цитрата связано как минимум с двумя мутациями после этих «усиливающих» мутаций. В более общем плане авторы предполагают, что эти результаты указывают, следуя аргументу Стивен Джей Гулд, «эта историческая случайность может иметь глубокое и длительное влияние» на ход эволюции.[7] Эти открытия стали рассматриваться как важный пример влияния исторической случайности на эволюцию.[15][41][42]
Геномный анализ Cit+ черта и значение для эволюционных инноваций
В 2012 году Ленски и его команда сообщили о результатах геномного анализа Cit+ признак, проливающий свет на генетическую основу и эволюционную историю признака. Исследователи секвенировали полные геномы двадцати девяти клонов, выделенных из различных временных точек в истории популяции Ara-3. Они использовали эти последовательности, чтобы реконструировать филогенетическую историю популяции; эта реконструкция показала, что население разделилось на три клады 20 000 поколений. Cit+ варианты эволюционировали в одном из них, который они назвали Clade 3. Клоны, потенцированные в более ранних исследованиях, были распределены среди всех трех клад, но были чрезмерно представлены в Clade 3. Это привело исследователей к выводу, что были были по крайней мере две потенцирующие мутации, вовлеченные в Cit+ эволюция.[8]
Исследователи также обнаружили, что все Cit+ клоны имели мутации, в которых дуплицировался или амплифицировался сегмент ДНК из 2933 пар оснований. Дублированный сегмент содержал ген CITT для белка-переносчика цитрата, используемого при анаэробном росте на цитрате. Дупликация является тандемной, и в результате копии расположены лицом к хвосту по отношению друг к другу. Эта новая конфигурация поместила копию ранее безмолвного, невыраженного CITT под контролем соседних rnk промотор гена, который управляет экспрессией при наличии кислорода. Этот новый rnk-citT модуль создал новую нормативную схему для CITT, активируя экспрессию переносчика цитрата в присутствии кислорода и, таким образом, обеспечивая аэробный рост цитрата.[8]
Движение этого rnk-citT модуля в геном потенцированного Cit− было показано, что клона достаточно для получения Cit+ фенотип. Однако первоначальный Cit+ фенотип, обусловленный дупликацией, был очень слабым и давал только ~ 1% -ное улучшение приспособленности. Исследователи обнаружили, что количество копий rnk-citT модуль пришлось увеличить для усиления Cit+ достаточно, чтобы бактерии могли хорошо расти на цитрате. Дальнейшие мутации после Cit+ бактерии стали доминировать в популяции, продолжали накапливать улучшенный рост на цитрате.
Исследователи пришли к выводу, что эволюция Cit+ проявился в трех различных фазах: (1) накапливались мутации, которые увеличивали скорость мутации Cit+, (2) сам признак проявился в слабой форме, и (3) признак был улучшен более поздними мутациями. Blount et al. предположил, что эта модель может быть типичной для того, как в целом развиваются новые черты, и предложил трехступенчатую модель эволюционных инноваций:
- Потенцирование: развивается генетический фон, при котором признак становится мутационно доступным, что делает возможной эволюцию признака.
- Актуализация: происходит мутация, которая приводит к появлению признака, делая его проявленным, хотя, вероятно, в слабой форме.
- Уточнение: Как только признак существует, если он обеспечивает селективное преимущество, накапливаются мутации, которые улучшают признак, делая его эффективным. Эта фаза является открытой и будет продолжаться до тех пор, пока возникают уточняющие мутации, а признак остается полезным.[8][15]
Эта модель получила признание в эволюционной биологии. В 2015 году палеонтолог Дуглас Эрвин предложили модификацию четырехступенчатой модели, чтобы лучше отразить возможное различие между эволюционной новизной и эволюционной инновацией, а также подчеркнуть важность условий окружающей среды: потенцирование, генерация новых фенотипов (актуализация), адаптивное совершенствование и эксплуатация (преобразование новизна новшества, поскольку она становится важной для экологического становления организмов-обладателей).[44]
Исследование потенцирования
В 2014 году исследовательская группа под руководством Эрика Квандта в лаборатории Джеффри Баррика в Техасский университет в Остине описал применение новой техники, называемой рекурсивной рекомбинацией и секвенированием в масштабе всего генома (REGRES), для выявления потенцирующих мутаций среди 70 присутствующих в линии Ara-3, которые привели к развитию Cit+.[45] В этом методе использовалось несколько раундов процесса, в котором F плазмида основан спряжение Между 33000 поколением Cit+ clone, CZB154, и Cit− основание клона LTEE для очистки мутаций, не требуемых ни для проявления слабой, ни для сильной формы Cit+ черта, которую они называют Cit++. Они обнаружили, что rnk-citT модуль, отвечающий за фенотипический переход на Cit+ было достаточно, чтобы произвести слабый Cit+ фенотип предка. Они также идентифицировали мутацию, которая произошла в линии, ведущей к CZB154, после начальной эволюции Cit+ что дало сильный, Cit++ фенотип предка без мутации, кроме rnk-citT модуль. Эта мутация, обнаруженная в регуляторной области гена, называемого dctA, вызвало резкое увеличение выраженности Транспортер DctA, который выполняет импорт C4-дикарбоксилаты в клетку. Они обнаружили, что это увеличенное выражение DctA позволяет Cit+ клетки для повторного захвата сукцинат, малат, и фумарат высвобождается в среду транспортером CitT во время импорта цитрата. Они выявили аналогичную мутацию в Cit++ клоны в популяции Ara-3, которые увеличивали экспрессию DctA за счет восстановления функции гена, который его регулирует, dcuS, который был деактивирован в предковом клоне. Quandt et al. пришел к выводу, что dctA мутация участвовала не в потенцировании, а в уточнении. Это заставило их предположить, что эволюция Cit+ в популяции Ara-3, возможно, зависели от генетического фона и популяционно-специфической экологии, которые позволили раннему, слабому Cit+ варианты, чтобы сохраняться в популяции достаточно долго, чтобы возникли уточняющие мутации и сделать рост на цитрате достаточно сильным, чтобы обеспечить значительное улучшение пригодности.
Позже Квандт и его коллеги опубликовали результаты, окончательно идентифицирующие мутацию, которая действительно усиливала Cit+ эволюция.[46] Эта мутация была в gltA ген, который кодирует цитрат-синтаза, фермент, участвующий в потоке углерода в цикл лимонной кислоты. Он имел эффект увеличения активности цитрат-синтазы, и они показали, что он позволяет улучшить рост ацетат. Более того, с gltA мутация, rnk-citT модуль, вызывающий Cit+ trait имеет нейтральный или слегка полезный эффект приспособленности, в то время как без него модуль был бы сильно вредным. В gltA мутация поэтому, по-видимому, позволила ранний, слабый Cit+ варианты сохраняться в популяции до тех пор, пока не могут произойти уточняющие мутации в соответствии с их более ранними выводами. После сильного Cit++ фенотип развился, повышенная активность цитратсинтазы стала пагубной. Исследователи обнаружили, что более поздние мутации в gltA противодействовал первой мутации, снижая активность цитрат-синтазы и улучшая рост цитрата. Они пришли к выводу, что серия мутаций в gltA сначала потенцированный, а затем уточненный рост на цитрате. Они также предположили, что родословная, в которой Cit+ возникла, возможно, заняла нишу в Ara-3, основанную на росте на ацетате, и что потенцирующие мутации, которые привели к эволюции Cit+ в Ara-3 были первоначально приспособлены к использованию ацетата.
Расследование пост-Cit+ экология и стойкое разнообразие
Небольшая субпопуляция Cit− клетки, неспособные расти на цитрате и принадлежащие к отдельной кладе, сохранялись в популяции после Cit+ клетки стали доминирующими. Ранние результаты показали, что это разнообразие частично связано с Cit− клетки лучше растут на глюкозе в среде.[7] Тернер и др. позже обнаружил, что еще одним фактором сосуществования было то, что Cit− клетки развили способность перекрестного питания на Cit+ большинство. Они показали, что Cit+ клетки высвобождают сукцинат, малат, и фумарат во время роста на цитрате, поскольку переносчик CitT выкачивает эти вещества из клетки, накачивая цитрат в клетку. Cit− клетки быстро развили способность расти на этих веществах из-за мутации, которая восстанавливала экспрессию соответствующего белка-переносчика, который молчал у предка.[47]
Cit− субпопуляция в конечном итоге ушла вымерший среди населения от 43 500 до 44 000 поколений. Было показано, что это вымирание произошло не из-за Cit+ большинство развивается, чтобы иметь возможность вторгаться ниша, занятая Cit− меньшинство. Действительно, Cit− клоны могли вторгнуться в Cit+ популяции после вымирания. Более того, в эксперименте, в котором они перезапустили двадцать копий популяции Ara-3 из образца, замороженного за 500 поколений до исчезновения, Turner et al. обнаружил, что Cit− субпопуляция не исчезла ни в одной из повторностей после 500 поколений эволюции. Одна из этих реплик продолжалась 2500 поколений, в течение которых Cit− продолжали сосуществовать. Исследователи пришли к выводу, что исчезновение Cit− произошло из-за какого-то неизвестного «редкого нарушения окружающей среды», аналогичного тому, которое может повлиять на естественные популяции.[48] Последняя реплика была интегрирована в основной эксперимент LTEE, став тринадцатой популяцией, Ara-7.[49]
Критика результатов использования цитрата
Другие исследователи экспериментировали с развитием аэробных цитрат, использующих Кишечная палочка. Дастин Ван Хофвеген и др., Работающие в лаборатории умный дизайн сторонник Скотт Миннич, смогли выделить 46 независимых цитрат-утилизирующих мутантов Кишечная палочка всего за от 12 до 100 поколений, используя очень длительный отбор в условиях голодания, во время которого бактерии будут быстрее обнаруживать больше мутаций.[50] В их исследованиях секвенирование геномной ДНК показало усиление CITT и dctA loci и перестройка ДНК были тем же классом мутаций, который был идентифицирован в эксперименте Ричарда Ленски и его команды. Они пришли к выводу, что редкость мутанта, использующего цитрат, в исследованиях Ленски, скорее всего, была результатом селективных экспериментальных условий, используемых его командой, а не уникальным событием эволюционного видообразования.[50]
Джон Рот и Софи Майснье-Патен рассмотрели подходы как к отложенным мутациям группы Ленски, так и к быстрым мутациям группы Ван Хофвегеса на Кишечная палочка. Они утверждают, что обе команды испытали одну и ту же последовательность потенцирования, актуализации и уточнения, приведшую к аналогичному Cit.+ варианты.[51] По их словам, период менее суток, в течение которого использование цитрата будет выбираться, с последующим 100-кратным разбавлением и периодом роста глюкозы, который не может быть выбран для использования цитрата, в конечном итоге снизил вероятность Кишечная палочка способность накапливать ранние адаптивные мутации от одного периода отбора к другому.[51] С другой стороны, команда Ван Хофвегена разрешила непрерывный период отбора продолжительностью 7 дней, что привело к более быстрому развитию применения цитратов. Кишечная палочка. Рот и Майнье-Патен предполагают, что серийное разведение Кишечная палочка и короткий период отбора для использования цитрата в условиях LTEE постоянно препятствовал каждому поколению Кишечная палочка от достижения следующих стадий использования аэробных цитратов.[51]
В ответ Блаунт и Ленски признают, что проблема не в экспериментах или данных, а в интерпретациях, сделанных Ван Хофвегеном и др. и Мезнье-Патен и Рот.[52] Ленский отмечает, что стремительная эволюция Cit+ не обязательно был неожиданностью, поскольку его команда также смогла создать несколько Cit+ мутанты через несколько недель во время экспериментов по воспроизведению, о которых они сообщили в статье 2008 года, в которой его команда впервые описала эволюцию аэробного использования цитрата в LTEE.[53] Кроме того, Ленски критикует описание Ван Хофвегена и др. Первоначальной эволюции Cit+ как «событие видообразования», указав, что LTEE не был разработан для выделения мутантов, использующих цитрат, или для борьбы с видообразованием, поскольку в своей статье 2008 года они сказали, что «став Citrate+ был лишь первым шагом на пути к возможному видообразованию ", и поэтому не предлагал, чтобы Cit+ мутанты принадлежали к другому виду, но это видообразование могло быть возможным следствием эволюции признака.[53] Ленски признает, что ученые, в том числе он и его команда, часто используют сокращенный язык и жаргон при обсуждении видообразования вместо того, чтобы писать более тщательно и точно по этому вопросу, и это может вызвать проблемы.[53] Однако он отмечает, что биологи-эволюционисты считают видообразование процессом, а не событием.[53] Он также критикует Ван Хофвегена и др. и Рот и Майснье-Патен за постулирование «ложных дихотомий» в отношении сложной концепции исторической случайности. Он утверждает, что историческая случайность означает, что история имеет значение, и что в их статье 2008 года представлены данные, показывающие, что эволюция Cit+ в LTEE был обусловлен мутациями, накопленными ранее. Он заключает, что «... историческая случайность была задействована и продемонстрирована в конкретном контексте, а именно в контексте появления Cit+ в LTEE - это не означает, что появление Cit+ исторически обусловлен в других экспериментальных контекстах, и в этом отношении другие изменения в LTEE исторически случайны - на самом деле, некоторые другие эволюционные изменения в LTEE были в высшей степени предсказуемыми и не (или, по крайней мере, не очевидно) зависели от предыдущих мутаций в населения ".[53]
Смотрите также
Рекомендации
- ^ Пенниси, Элизабет (14 ноября 2013 г.). «Человек, разливший в бутылки эволюцию». Наука. 342 (6160): 790–793. Bibcode:2013Наука ... 342..790П. Дои:10.1126 / science.342.6160.790. PMID 24233702.
- ^ а б Ленский, Ричард Э. (2000). «Источник основного напряжения». Домашняя страница Ричарда Э. Ленски. Университет штата Мичиган. Получено 2008-06-18.
- ^ "Ричард Э. Ленски (@RELenski) | Twitter". twitter.com. Получено 2016-07-02.
- ^ "Морщинки времени". Возвращение к Теллиамеду. 2017-03-13. Получено 2017-03-13.
- ^ bewilderbeast (24.02.2020). «Мы прерываем этот неприятный вирус хорошими новостями о бактериях». Возвращение к Теллиамеду. Получено 2020-08-06.
- ^ «Мы прерываем этот эксперимент». Возвращение к Теллиамеду. 2020-03-09. Получено 2020-08-06.
- ^ а б c d е ж Блаунт, Захари Д.; Borland, Christina Z .; Ленский, Ричард Э. (2008). "Историческая случайность и эволюция ключевой инновации в экспериментальной популяции кишечная палочка". Труды Национальной академии наук. 105 (23): 7899–906. Bibcode:2008PNAS..105.7899B. Дои:10.1073 / pnas.0803151105. JSTOR 25462703. ЧВК 2430337. PMID 18524956.
- ^ а б c d е Блаунт З.Д., Баррик Дж. Э., Дэвидсон С. Дж., Ленски Р. Э. (27.09.2012). "Геномный анализ ключевой инновации в экспериментальной кишечная палочка численность населения". Природа. 489 (7417): 513–518. Bibcode:2012Натура 489..513Б. Дои:10.1038 / природа11514. ЧВК 3461117. PMID 22992527.
- ^ «Мы прерываем этот эксперимент». Возвращение к Теллиамеду. 2020-03-09. Получено 2020-03-09.
- ^ а б «Еще пять лет». Возвращение к Теллиамеду. 2020-05-04. Получено 2020-05-09.
- ^ а б c d е Ленский, Ричард Э. (2010). "Фенотипическая и геномная эволюция во время эксперимента с бактериями поколения 20000 кишечная палочка". Отзывы о селекции растений. С. 225–265. Дои:10.1002 / 9780470650288.ch8. ISBN 978-0-471-46892-9.
- ^ Фокс, Джереми У .; Ленски, Ричард Э. (23 июня 2015 г.). "Отсюда и в вечность - теория и практика действительно длительного эксперимента". PLOS Биология. 13 (6): e1002185. Дои:10.1371 / journal.pbio.1002185. ЧВК 4477892. PMID 26102073.
- ^ Блаунт, Захари Д. (25 марта 2015 г.). «Неисчерпаемый потенциал кишечной палочки». eLife. 4. Дои:10.7554 / eLife.05826. ЧВК 4373459. PMID 25807083.
- ^ а б «Жидкая среда DM25». myxo.css.msu.edu. Получено 2016-05-24.
- ^ а б c d е Блаунт, Захари Д. (август 2016 г.). «Пример эволюционного непредвиденного обстоятельства». Исследования по истории и философии науки Часть C: Исследования по истории и философии биологических и биомедицинских наук. 58: 82–92. Дои:10.1016 / j.shpsc.2015.12.007. PMID 26787098.
- ^ Фитцджеральд, Джордж; Уильямс, Лютер С. (апрель 1975 г.). «Модифицированная процедура обогащения пенициллина для отбора бактериальных мутантов». Журнал бактериологии. 122 (1): 345–346. Дои:10.1128 / JB.122.1.345-346.1975. ЧВК 235679. PMID 1091629.
- ^ Wagegg, Вт; Braun, V (January 1981). "Ferric citrate transport in Escherichia coli requires outer membrane receptor protein fecA". Журнал бактериологии. 145 (1): 156–163. Дои:10.1128/JB.145.1.156-163.1981. ЧВК 217256. PMID 7007312.
- ^ Ленский, Ричард Э. (2000). "Overview of the Кишечная палочка long-term evolution experiment". Домашняя страница Ричарда Э. Ленски. Университет штата Мичиган. Получено 2008-06-18.
- ^ "Фото". Blount Lab. Получено 2016-05-28.
- ^ "Overview of the Кишечная палочка long-term evolution experiment". myxo.css.msu.edu. Получено 2016-05-28.
- ^ Wiser, M. J.; Ribeck, N.; Lenski, R. E. (14 November 2013). "Long-Term Dynamics of Adaptation in Asexual Populations". Наука. 342 (6164): 1364–1367. Bibcode:2013Sci...342.1364W. Дои:10.1126/science.1243357. PMID 24231808. S2CID 15341707.
- ^ Scharping, Nathaniel (2015-12-16). "Could Evolution Ever Yield a 'Perfect' Organism?". Откройте для себя журнал. Архивировано из оригинал 18 декабря 2015 г.. Получено 18 декабря 2015.
- ^ Lenski, Richard E; и другие. (2015). "Sustained fitness gains and variability in fitness trajectories in the long-term evolution experiment with кишечная палочка". Труды Королевского общества B: биологические науки. 282 (1821): 20152292. Дои:10.1098/rspb.2015.2292. ЧВК 4707762. PMID 26674951.
- ^ Kaznatcheev, Artem (May 2019). "Computational Complexity as an Ultimate Constraint on Evolution". Генетика. 212 (1): 245–265. Дои:10.1534/genetics.119.302000. ЧВК 6499524. PMID 30833289.
- ^ Sniegowski, Paul D.; Gerrish, Philip J.; Lenski, Richard E. (June 1997). "Evolution of high mutation rates in experimental populations of E. coli". Природа. 387 (6634): 703–705. Bibcode:1997Natur.387..703S. Дои:10.1038/42701. PMID 9192894. S2CID 4351382.
- ^ Barrick, J. E.; Lenski, R. E. (23 September 2009). "Genome-wide Mutational Diversity in an Evolving Population of Escherichia coli". Симпозиумы Колд-Спринг-Харбор по количественной биологии. 74: 119–129. Дои:10.1101/sqb.2009.74.018. ЧВК 2890043. PMID 19776167.
- ^ Баррик, Джеффри Э .; Yu, Dong Su; Yoon, Sung Ho; Jeong, Haeyoung; Oh, Tae Kwang; Schneider, Dominique; Ленски, Ричард Э .; Kim, Jihyun F. (18 October 2009). "Genome evolution and adaptation in a long-term experiment with Escherichia coli". Природа. 461 (7268): 1243–1247. Bibcode:2009Natur.461.1243B. Дои:10.1038/nature08480. PMID 19838166. S2CID 4330305.
- ^ а б Philippe, Nadège; Pelosi, Ludovic; Ленски, Ричард Э .; Schneider, Dominique (2008). "Evolution of Penicillin-Binding Protein 2 Concentration and Cell Shape during a Long-Term Experiment with кишечная палочка". Журнал бактериологии. 191 (3): 909–21. Дои:10.1128/JB.01419-08. ЧВК 2632098. PMID 19047356.
- ^ Cooper, Vaughn S.; Lenski, Richard E. (October 2000). "The population genetics of ecological specialization in evolving Escherichia coli populations". Природа. 407 (6805): 736–739. Bibcode:2000Natur.407..736C. Дои:10.1038/35037572. PMID 11048718. S2CID 205009743.
- ^ Leiby, Nicholas; Marx, Christopher J.; Moran, Nancy A. (18 February 2014). "Metabolic Erosion Primarily Through Mutation Accumulation, and Not Tradeoffs, Drives Limited Evolution of Substrate Specificity in Escherichia coli". PLOS Биология. 12 (2): e1001789. Дои:10.1371/journal.pbio.1001789. ЧВК 3928024. PMID 24558347.
- ^ а б Rozen, Daniel E.; Lenski, Richard E. (January 2000). "Long-Term Experimental Evolution in кишечная палочка. VIII. Dynamics of a Balanced Polymorphism". Американский натуралист. 155 (1): 24–35. Дои:10.1086/303299. PMID 10657174.
- ^ Rozen, Daniel E.; Schneider, Dominique; Lenski, Richard E. (27 June 2005). "Long-Term Experimental Evolution in Escherichia coli. XIII. Phylogenetic History of a Balanced Polymorphism". Журнал молекулярной эволюции. 61 (2): 171–180. Bibcode:2005JMolE..61..171R. Дои:10.1007/s00239-004-0322-2. PMID 15999245. S2CID 6970967.
- ^ "On the Evolution of Citrate Use". Возвращение к Теллиамеду. 2016-02-20. Получено 2016-05-26.
- ^ а б c "Cell Biology: The Use of Citrate". EVO-ED. Университет Мичигана.
- ^ Lara, F.J.S; Stokes, J.L. (1952). "Oxidation of citrate by кишечная палочка". Журнал бактериологии. 63 (3): 415–420. Дои:10.1128/JB.63.3.415-420.1952. ЧВК 169284. PMID 14927574.
- ^ Lütgens, M.; Gottschalk, G. (1 July 1980). "Why a Co-substrate is Required for Anaerobic Growth of Escherichia coli on Citrate". Микробиология. 119 (1): 63–70. Дои:10.1099/00221287-119-1-63. PMID 6997437.
- ^ Pos, Klaas Martinus; Димрот, Питер; Bott, Michael (August 1998). "The кишечная палочка Citrate Carrier CitT: a Member of a Novel Eubacterial Transporter Family Related to the 2-Oxoglutarate/Malate Translocator from Spinach Chloroplasts". Журнал бактериологии. 180 (16): 4160–4165. Дои:10.1128/JB.180.16.4160-4165.1998. ЧВК 107412. PMID 9696764.
- ^ а б Reynolds, C H; Silver, S (December 1983). "Citrate utilization by Escherichia coli: plasmid- and chromosome-encoded systems". Журнал бактериологии. 156 (3): 1019–1024. Дои:10.1128/JB.156.3.1019-1024.1983. ЧВК 217945. PMID 6358185.
- ^ а б Hall, B G (July 1982). "Chromosomal mutation for citrate utilization by Escherichia coli K-12". Журнал бактериологии. 151 (1): 269–273. Дои:10.1128/JB.151.1.269-273.1982. ЧВК 220237. PMID 7045076.
- ^ "кишечная палочка K-12 substr. MG1655 citT". ecocyc.org. Получено 2016-05-23.
- ^ Desjardins, Eric (2011-01-01). "Historicity and Experimental Evolution". Биология и философия. 26 (3): 339–364. Дои:10.1007/s10539-011-9256-4. S2CID 83908986.
- ^ Битти, Джон; Carrera, Isabel (2011-01-01). "When What Had to Happen Was Not Bound to Happen: History, Chance, Narrative, Evolution". Journal of the Philosophy of History. 5 (3): 471–495. Дои:10.1163/187226311x599916.
- ^ "Фото". Blount Lab. Получено 2017-10-06.
- ^ Erwin, Douglas H. (October 2015). "Novelty and Innovation in the History of Life". Текущая биология. 25 (19): R930–R940. Дои:10.1016/j.cub.2015.08.019. PMID 26439356.
- ^ Quandt, Erik M.; Deatherage, Daniel E .; Эллингтон, Эндрю Д.; Georgiou, George; Barrick, Jeffrey E. (11 February 2014). "Recursive genomewide recombination and sequencing reveals a key refinement step in the evolution of a metabolic innovation in кишечная палочка". Труды Национальной академии наук. 111 (6): 2217–2222. Bibcode:2014PNAS..111.2217Q. Дои:10.1073/pnas.1314561111. ЧВК 3926077. PMID 24379390.
- ^ Quandt, Erik M; Gollihar, Jimmy; Blount, Zachary D; Ellington, Andrew D; Georgiou, George; Barrick, Jeffrey E (14 October 2015). "Fine-tuning citrate synthase flux potentiates and refines metabolic innovation in the Lenski evolution experiment". eLife. 4. Дои:10.7554/eLife.09696. ЧВК 4718724. PMID 26465114.
- ^ Turner, Caroline B.; Блаунт, Захари Д.; Mitchell, Daniel H.; Lenski, Richard E. (2015-06-17). "Evolution and coexistence in response to a key innovation in a long-term evolution experiment with кишечная палочка". bioRxiv 10.1101/020958.
- ^ Saey, Tina Hesman (2015-09-09). "Extinction in lab bottle was a fluke, experiment finds". Новости науки. Получено 2016-06-04.
- ^ Turner, Caroline B.; Блаунт, Захари Д.; Ленски, Ричард Э .; Cohan, Frederick M. (18 November 2015). "Replaying Evolution to Test the Cause of Extinction of One Ecotype in an Experimentally Evolved Population". PLOS ONE. 10 (11): e0142050. Bibcode:2015PLoSO..1042050T. Дои:10.1371/journal.pone.0142050. ЧВК 4651540. PMID 26581098.
- ^ а б Van Hofwegen, Dustin J.; Hovde, Carolyn J.; Minnich, Scott A.; Silhavy, T. J. (1 April 2016). "Rapid Evolution of Citrate Utilization by кишечная палочка by Direct Selection Requires citT and dctA". Журнал бактериологии. 198 (7): 1022–1034. Дои:10.1128/JB.00831-15. ЧВК 4800869. PMID 26833416.
- ^ а б c Roth, John R.; Maisnier-Patin, Sophie; Silhavy, T. J. (1 April 2016). "Reinterpreting Long-Term Evolution Experiments: Is Delayed Adaptation an Example of Historical Contingency or a Consequence of Intermittent Selection?". Журнал бактериологии. 198 (7): 1009–1012. Дои:10.1128/JB.00110-16. ЧВК 4800865. PMID 26883821.
- ^ "Similar Data, Different Conclusions | The Scientist Magazine®". The Scientist. Получено 2016-05-21.
- ^ а б c d е Lenski, Richard (2016-02-20). "On the Evolution of Citrate Use". Возвращение к Теллиамеду.
дальнейшее чтение
- Докинз, Ричард (2009). "Forty-Five Thousand Generations of Evolution in the Lab". Величайшее шоу на Земле: свидетельства эволюции. Нью-Йорк: Свободная пресса. стр.116–33. ISBN 978-1-4165-9478-9.
- John Timmer (17 November 2013). "After 50,000 generations, bacteria are still evolving greater fitness". Ars Technica.
внешняя ссылка
- Кишечная палочка Long-term Experimental Evolution Project Site
- Bacteria make major evolutionary shift in the lab Боб Холмс Новый ученый 9 июня 2008 г.
- Evolution: Past, Present and Future Ричард Ленски
- List of publications on the experiment
- Online Publication of paper on Rapid evolution of citrate utilization