Эволюция рыб - Википедия - Evolution of fish

В эволюция рыб началось около 530 миллионов лет назад во время Кембрийский взрыв. Именно в это время первые хордовые разработал череп и позвоночник, ведущий к первому краниаты и позвоночные. Первый рыбы родословные принадлежат к Агната, или рыба без челюсти. Ранние примеры включают Haikouichthys. Во время позднего Кембрийский, угревидная рыба без челюсти, называемая конодонты, и небольшая в основном бронированная рыба, известная как остракодермы, впервые появился. Большинство бесчелюстных рыб теперь вымерли; но сохранившиеся миноги может напоминать древнюю предварительно челюстную рыбу. Миноги относятся к Циклостомы, который включает дошедшие до нас миксина, и эта группа, возможно, рано отделилась от других бесчеловечных.
Раннее челюстные позвоночные вероятно, возник в конце Ордовик период. Впервые они представлены в Окаменелости от Силурийский двумя группами рыб: бронированной рыбой, известной как плакодермы, который произошел от остракодерм; и Acanthodii (или колючие акулы). Челюстные рыбы, которые сохранились до наших дней, также появились в конце силурия: Chondrichthyes (или хрящевую рыбу) и Osteichthyes (или костлявую рыбу). Костные рыбы разделились на две отдельные группы: Актиноптеригии (или рыба с луговыми плавниками) и Саркоптеригии (включая рыбу с лопастными плавниками).
Вовремя Девонский В этот период произошло значительное увеличение разнообразия рыб, особенно среди остракодерм и плакодерм, а также среди рыб с лопастными плавниками и ранних акул. Это привело к тому, что девон стал известен как возраст рыб. Именно от рыбы с лопастными плавниками четвероногие эволюционировали, четырехногие позвоночные, представленные сегодня амфибии, рептилии, млекопитающие, и птицы. Переходные четвероногие впервые появились в раннем девоне, а к концу девона появились первые четвероногие. Разнообразие челюстных позвоночных может указывать на эволюционное преимущество челюстных позвоночных. рот; но неясно, является ли преимущество шарнирной челюсти большей силой прикуса, улучшенным дыханием или комбинацией факторов. Рыбы не представляют собой монофилетический группа, но парафилетический один, поскольку они исключают четвероногие.[1]
Рыба, как и многие другие организмы, сильно пострадала от события вымирания на протяжении всей естественной истории. Самые ранние, Ордовикско-силурийские события вымирания, привело к потере многих видов. В вымирание в конце девона привело к исчезновению остракодерм и плакодерм к концу девона, а также других рыб. Колючие акулы вымерли в Пермско-триасовое вымирание; конодонты вымерли в Триасово-юрское вымирание. В Меловое – палеогеновое вымирание, и сегодняшний день Голоценовое вымирание, также повлияли на разнообразие рыб и рыбные запасы.
Обзор

Обычная классификация имеет живую позвоночные как подтип, сгруппированный в восемь классов на основе традиционных интерпретаций грубого анатомический и физиологический черты. В свою очередь, эти классы сгруппированы в позвоночных, имеющих четыре конечности ( четвероногие ) и те, которые этого не делают: Рыбы. В сохранившийся Классы позвоночных:[3]
- Рыбы:
- бесчелюстные рыбы (Агната)
- хрящевые рыбы (Хондрихтиес)
- рыбы с луговыми плавниками (Актиноптеригии)
- рыба с лопастными плавниками (Саркоптеригия)
- Тетраподы:
- амфибии (Амфибия)
- рептилии (Рептилии)
- птицы (Авес)
- млекопитающие (Млекопитающие)
Рыба, возможно, произошла от животного, похожего на коралловидное морской брызг (а оболочка ), личинки которых во многом напоминают ранних рыб. Первые предки рыб могли иметь сохранил личиночную форму в зрелом возрасте (как это делают сегодня некоторые морские брызги), хотя этот путь нельзя доказать.
Позвоночные, среди них первые Рыбы, возникла около 530 миллионов лет назад во время Кембрийский взрыв, что привело к увеличению разнообразия организмов.[4]



Первые предки рыб или животных, которые, вероятно, были тесно связаны с рыбами, были Пикайя, Haikouichthys и Myllokunmingia.[8][4] Эти три роды все появилось около 530 Ма. Пикайя имел примитивный нотохорда, структура, которая могла бы развиться в позвоночник потом. В отличие от другой фауны, доминировавшей в кембрии, эти группы имели основные позвоночные план тела: а нотохорда, рудиментарные позвонки и хорошо выраженные голова и хвост.[9] Всем этим ранним позвоночным не хватало челюсти в обычном смысле и полагались на фильтрующую подачу близко к морскому дну.[10]
За ними последовали бесспорные ископаемые позвоночные в виде рыб с тяжелыми панцирями, обнаруженные в породах с Ордовик Период 500–430 гг. Ма.
Первый челюстные позвоночные появился в конце Ордовик и стал обычным явлением в Девонский, часто известный как «Эпоха рыб».[11] Две группы костлявые рыбы, то актиноптеригии и саркоптеригия, развились и стали обычным явлением.[12] Девонский период также стал свидетелем гибели практически всех бесчелюстных рыб, за исключением миног и миксин, а также Плакодерми, группа бронированных рыб, доминировавшая в последнее время Силурийский. В девоне также наблюдался рост первых лабиринтодонты, который был переходным звеном между рыбами и амфибии.
Колонизация новых ниши привело к разнообразию строения тела, а иногда и к увеличению в размерах. В Девонский Период (395–345 млн лет) принес с собой таких гигантов, как плакодерма Дунклеостей, которые могли вырасти до семи метров в длину, и ранние дышащие воздухом рыбы, которые могли оставаться на суше в течение длительного времени. Среди этой последней группы были предковые амфибии.
В рептилии появились из лабиринтодонтов в последующие Каменноугольный период. В анапсид и синапсид рептилии были обычным явлением в конце Палеозой, в то время как диапсиды стал доминирующим во время Мезозойский. В море костлявые рыбы стал доминирующим.
Более поздние излучения, такие как излучение рыб в силурийский и девонский периоды, затрагивали меньшее количество таксонов, в основном с очень похожими строениями тела. Первыми животными, которые отважились на сушу, были членистоногие. У некоторых рыб были легкие и крепкие костлявые плавники, и они также могли заползать на сушу.
Рыба без челюсти


Бесчелюстные рыбы относятся к суперкласс Агната в филюм Хордовые, подтип Позвоночные. Агната происходит из Греческий, и означает «без челюстей».[13] Это исключает всех позвоночных с челюстями, известных как гнатостомы. Хотя это второстепенный элемент современного морского фауна, рыба без челюсти была известна среди первых рыб в раннем Палеозой. Два типа Ранний кембрий животное, имеющее плавники, позвоночное животное мускулатура и жабры известны с раннего кембрия. Маотяньшаньские сланцы из Китай: Haikouichthys и Myllokunmingia. Их предварительно назначил Агнате Жанвье. Третий возможный агнатид из того же региона - Haikouella. О возможном агнатиде, который не был официально описан, сообщил Симонетти из среднего кембрия. Burgess Shale из британская Колумбия.[нужна цитата ]
Многие ордовикские, силурийские и девонские агнаты были бронированы тяжелыми костлявыми пластинами сложной формы, полученными из минерализованной чешуи. Первые бронированные агнатаны - Остракодермы, предшественники костлявая рыба и, следовательно, к четвероногие (включая людей) - известны с середины Ордовик, а к позднему Силурийский агнатаны достигли высшей точки своей эволюции. Большинство остракодерм, таких как телодонты, остеостраки, и галеаспиды, были более тесно связаны с гнатостомами, чем с выжившими бесцветными, известными как циклостомы. Циклостомы, по-видимому, отделились от других бесчеловечных насекомых до эволюции дентина и кости, которые присутствуют у многих ископаемых бесчелюстных, в том числе конодонты.[14] Агнатанс отказался от Девонский и никогда не поправлялся.
Агнатаны в целом парафилетический,[15] потому что большинство вымерших бесчеловечных насекомых принадлежат к стволовая группа гнатостомов.[16][17] Недавние молекулярные данные, как из рРНК[18] и из мтДНК[19] решительно поддерживает теорию о том, что живые агнатаны, известные как циклостомы, находятся монофилетический.[20] В филогенетическая таксономия, отношения между животными обычно не делятся на ранги, а изображаются в виде вложенного «семейного дерева», известного как кладограмма. Филогенетическим группам даются определения, основанные на их отношениях друг с другом, а не чисто на физических признаках, таких как наличие позвоночника. Этот образец гнездования часто сочетается с традиционной таксономией в практике, известной как эволюционная таксономия.

В кладограмма ниже для бесчелюстных рыб основано на исследованиях, проведенных Филипп Жанвье и другие для Веб-проект "Древо жизни".[22] († = группа вымерла)
| Рыба без челюсти |
| |||||||||||||||||||||||||||||||||||||||
†Конодонты

Конодонты напоминали примитивных бесчелюстных угрей. Они появились 520 млн лет назад и были уничтожены 200 млн лет назад.[24] Первоначально они были известны только по зубчатым микрофоссилиям, названным конодонтовые элементы. Эти «зубы» по-разному интерпретировались как устройства для подачи фильтра или как «захватывающий и дробящий набор».[25] Конодонты имели длину от сантиметра до 40 см. Промиссум.[25] Их большие глаза были расположены сбоку, что делает маловероятным их хищническую роль. Сохранившаяся мускулатура намекает на то, что некоторые конодонты (Промиссум по крайней мере) были эффективными крейсерами, но неспособными развивать скорость.[25] В 2012 г. исследователи классифицировали конодонтов в филюм Хордовые на основе их плавников с плавниковыми лучами, шеврон -образные мышцы и нотохорда.[26] Некоторые исследователи считают их позвоночными, внешне похожими на современных. миксина и миноги,[27] хотя филогенетический анализ показывает, что они больше полученный чем любая из этих групп.[28]
†Остракодермы

Остракодермы (без кожуры) находятся бронированный бесчелюстные рыбы из Палеозой. Сегодня этот термин не часто встречается в классификациях, потому что он парафилетический или же полифилетический, и не имеет филогенетического значения.[29] Тем не менее, этот термин до сих пор неофициально используется для группирования бронированных бесчелюстных рыб.
Доспех остракодермы состоял из 3–5-миллиметровых многоугольных пластин, которые защищали голову и жабры, а затем перекрывались ниже по телу, как чешуя. Глаза были особенно защищены. Ранее хордовые использовали свои жабры как для дыхания, так и для питания, тогда как остракодермы использовали свои жабры для дыхание Только. У них было до восьми отдельных глоточных жаберных мешочков по бокам головы, которые были постоянно открыты без какой-либо защиты. крышка. В отличие от беспозвоночные это использование реснитчатый движение для перемещения пищи, остракодермы использовали свой мускулистый глотку для создания всасывания, которое тянуло маленькие и медленные добыча в их рты.
Первыми ископаемыми рыбами, которые были обнаружены, были остракодермы. В Швейцарский анатом Луи Агассис получил несколько окаменелостей костлявых бронированных рыб из Шотландия в 1830-х гг. Ему было трудно классифицировать их, поскольку они не походили ни на одно живое существо. Сначала он сравнил их с существующими бронированными рыбами, такими как сом и осетровые но позже, осознав, что у них нет подвижных челюстей, в 1844 году классифицировал их в новую группу «остракодермы».[30]
Остракодермы существовали в двух основных группах, более примитивные гетеростраки и цефаласпиды. Позже, около 420 миллионов лет назад, челюстная рыба произошла от одной из остракодерм. После появления челюстных рыб, большинство видов остракодерм пришло в упадок, и последние остракодермы вымерли в конце XIX века. Девонский период.[31]
Челюстная рыба
Эта статья включает в себя список общих Рекомендации, но он остается в основном непроверенным, потому что ему не хватает соответствующих встроенные цитаты. (Январь 2014) (Узнайте, как и когда удалить этот шаблон сообщения) |
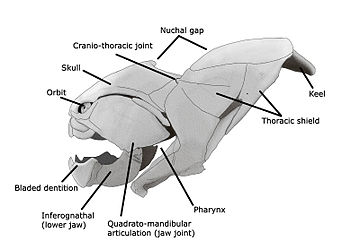

| Внешнее видео | |
|---|---|
Челюсть позвоночных, вероятно, первоначально возникла в Силурийский период и появился в Плакодерма рыбы, который далее диверсифицировался в Девонский. Два самых передних глоточные дуги считаются собственно челюстью и подъязычной дугой соответственно. Подъязычная система удерживает челюсть от мозговой оболочки черепа, обеспечивая большую подвижность челюстей. Уже давно предполагалось, что это парафилетический комплекс, ведущий к более производным гнатостомам, открытие Энтелогнатус предполагает, что плакодермы являются прямым предком современных костистых рыб.
Как и в большинстве позвоночные, рыбьи челюсти костлявый или же хрящевой и противостоят вертикально, составляя верхняя челюсть и нижняя челюсть. Челюсть происходит от двух самых передних. глоточные дуги поддерживает жабры и обычно несет многочисленные зубы. Предполагается, что череп последнего общего предка сегодняшних челюстных позвоночных напоминал акул.[32]
Считается, что изначальные селективные преимущества челюстей были связаны не с кормлением, а с увеличением эффективности дыхания. Челюсти использовались в буккальный насос (наблюдается у современных рыб и амфибии ), который перекачивает воду через жабры рыб или воздух в легкие у земноводных. В течение эволюционного периода было выбрано более привычное для людей использование челюстей для кормления, и это стало очень важной функцией у позвоночных. Много костистость рыбы существенно изменили свои челюсти для всасывающее питание и выступ челюсти, в результате получаются очень сложные челюсти с множеством задействованных костей.
Челюстные позвоночные и челюстные рыбы произошли от более ранних бесчелюстных рыб, а кладограмма внизу для челюстных позвоночных - продолжение кладограммы из раздела выше. († = группа вымерла)
| Челюстные позвоночные |
| ||||||||||||||||||||||||||||||||||||||||||||||||
†Плакодермы


Плакодермы, учебный класс Плакодерми (пластина без кожи), находятся вымерший бронированная доисторическая рыба, появившаяся около 430 млн лет назад в раннем-среднем силуре. В основном они были уничтожены во время Позднее девонское вымирание событие, 378 млн лет, хотя некоторые из них выжили и немного восстановили свое разнообразие во время Фаменский эпоха перед полным вымиранием в конце девона, 360 млн лет назад; в конечном итоге они являются предками современных позвоночных-гнатогомов.[33][34] Их голова и грудь были покрыты массивными и часто украшенными бронированными пластинами. Остальная часть тела была масштабированный или голые, в зависимости от вида. Броневой щит был шарнирным, при этом головная броня шарнирно соединялась с грудной. Это позволило плакодермам поднимать голову, в отличие от остракодерм. Плакодермы были первыми челюстными рыбами; их челюсти, вероятно, произошли от первого из их жабры арки. График справа показывает рост и исчезновение отдельных линий передачи плакодермы: Скребня, Ренанида, Антиархи, Petalichthyidae, Птиктодонтида и Артродира.
†Колючие акулы

Колючие акулы, класс Acanthodii, - это вымершие рыбы, которые имеют общие черты как с костными, так и с хрящевыми рыбами, хотя в конечном итоге более тесно связаны с последними и являются их предками. Несмотря на то, что их называли «колючими акулами», акантоды были предшественниками акул, хотя и породили их. Они появились в море в начале силурийского периода, примерно за 50 миллионов лет до появления первых акул. В конце концов конкуренция со стороны костистых рыб оказалась слишком сильной, и колючие акулы вымерли в пермские времена около 250 млн лет назад. По форме они напоминали акул, но их эпидермис был покрыт крошечными ромбовидными пластинками, похожими на чешуйки голостеи (Гарс, Bowfins ).
Хрящевые рыбы

Хрящевые рыбы, класс Chondrichthyes, состоящий из акулы, лучи и химеры, появившийся около 395 миллионов лет назад, в середине Девонский, произошедшие от акантодий. Класс содержит подклассы Голоцефали (химера ) и Elasmobranchii (акулы и лучи ). Излучение эласторазветвлений на диаграмме справа разделено на таксоны: Cladoselache, Eugeneodontiformes, Symmoriida, Xenacanthiformes, Ктенакантообразные, Hybodontiformes, Galeomorphi, Плоскообразные и Batoidea.
Костные рыбы
| Внешнее видео | |
|---|---|
Костные рыбы, класс Osteichthyes, характеризуются скорее костным скелетом, чем хрящ. Они появились в конце Силурийский, около 419 миллионов лет назад. Недавнее открытие Энтелогнатус настоятельно предполагает, что костистые рыбы (и, возможно, хрящевые рыбы, через акантодий) произошли от ранних плакодерм.[36] Подкласс Osteichthyes, лучеплавниковых рыб (Актиноптеригии ), стали доминирующая группа рыб в постпалеозойском и современном мире, насчитывающем около 30 000 ныне живущих видов.
Группы костных (и хрящевых) рыб, возникшие после девона, характеризовались постоянным улучшением кормодобывания и передвижения.[37]
Рыбы с лопастными плавниками

Рыбы с лопастными плавниками, рыба, принадлежащая к классу Sarcopterygii, в основном вымершие костистые рыбы, в основном характеризующиеся крепкими и короткодолевыми плавниками с прочным внутренним скелетом, космоидные весы и внутренние ноздри. Их плавники мясистые, дольчатый, парные плавники, соединенные с телом одной костью.[40] Плавники рыб с лопастными плавниками отличаются от плавников всех остальных рыб тем, что каждый имеет мясистый, лопастевидный чешуйчатый стебель, отходящий от тела. В грудной и тазовые плавники шарнирно сочленены, напоминая конечности четвероногих, предшественниками которых они были. Плавники превратились в ноги первых четвероногих наземных позвоночных, амфибии. У них также есть два спинных плавника с отдельными основаниями, в отличие от одного спинного плавника. рыба с плавниками. Головной мозг рыб с лопастными плавниками имеет примитивную линию петли, но у четвероногих и двоякодышащих рыб она отсутствует. У многих ранних рыб с лопастными плавниками симметричный хвост. У всех рыб с лопастными плавниками зубы покрыты истинным эмаль.
Рыбы с лопастными плавниками, такие как латимерия и двоякодышащая рыба, были самой разнообразной группой костистых рыб в девоне. Таксономисты, которые придерживаются кладистского подхода, включают группировку Тетрапода внутри Sarcopterygii, а четвероногие, в свою очередь, включают все виды четырехногих позвоночных.[41] Плавники рыб с лопастными плавниками, таких как латимерия, очень похожи на конечности конечностей четвероногих, ожидаемых от предков. Рыбы с лопастными плавниками, по-видимому, следовали двум различным линиям развития и, соответственно, разделены на два подкласса: Рипидистия (включая двоякодышащих и Тетраподоморфа, к которым относятся тетраподы) и Актинистия (латимерия). Первые рыбы с лопастными плавниками, обнаруженные в самом верхнем Силурийский (около 418 млн лет), очень похож на колючие акулы, который вымер в конце палеозоя. В начале – середине Девонский (416 - 385 млн лет назад), а хищный плакодермы доминировали над морями, некоторые рыбы с лопастными плавниками заходили в пресная вода среды обитания.

В раннем девоне (416-397 млн лет назад) лопастоперые рыбы разделились на две основные линии: латимерия и Rhipidistians. Первые никогда не покидали океаны, и их расцвет пришелся на Поздний девон и Каменноугольный от 385 до 299 млн лет назад, поскольку в эти периоды они встречались чаще, чем в любой другой период фанерозоя; латимерии по-прежнему живут в океаны (род Латимерия ). Рипидисты, предки которых, вероятно, жили в эстуарии, мигрировали в пресноводные местообитания. Они, в свою очередь, разделились на две основные группы: двоякодышащая рыба и тетраподоморфы. Наибольшее разнообразие двоякодышащих рыб было в Триасовый период; сегодня осталось менее десятка родов. Двоякодышащие рыбы развили первые протолёгкие и прото-конечности, развив способность жить вне водной среды в среднем девоне (397–385 млн лет назад). Первые тетраподоморфы, которые включали гигантских ризодонтов, имели ту же общую анатомию, что и двоякодышащие, которые были их ближайшими родственниками, но, похоже, они не покидали свою водную среду обитания до поздней девонской эпохи (385-359 млн лет назад), с появлением из четвероногие (четвероногие позвоночные). Тетраподы - единственные тетраподоморфы, которые выжили после девона. Лепестковые рыбы продолжались до конца палеозойской эры, неся большие потери во время Пермско-триасовое вымирание (251 млн лет).
Рыбы со скатными плавниками

Рыбы со скатными плавниками, класс Actinopterygii, отличаются от рыб с лопастными плавниками тем, что их плавники состоят из перепонок кожи, поддерживаемых шипами («лучами»), сделанными из кости или рога. Есть и другие различия в структурах дыхания и кровообращения. У рыб с лучевыми плавниками скелеты обычно состоят из настоящих костей, хотя это не относится к осетровые и веслонос.[44]
Рыбы с лучевыми плавниками - доминирующая группа позвоночных, в состав которой входит половина всех известных видов позвоночных. Они населяют морские глубины, прибрежные заливы, пресноводные реки и озера и являются основным источником пищи для людей.[44]
График
Преддевонский: Происхождение рыбы
| Кембрийский | Кембрийский (541–485 Ма ): Начало кембрия ознаменовалось Кембрийский взрыв, внезапное появление почти всех типов беспозвоночных животных (моллюски, медузы, черви и членистоногие, такие как ракообразные) в большом количестве. Первые позвоночные появились в виде примитивных рыб, которые впоследствии сильно разнообразились в силурии и девоне. | |||||
|---|---|---|---|---|---|---|
 | Пикайя |
Пикайя, вместе с Myllokunmingia и Haikouichthys ercaicunensis непосредственно ниже все кандидаты в летописи окаменелостей на звания «первое позвоночное животное» и «первая рыба». Пикайя это род появившихся около 530 Ма вовремя Кембрийский взрыв многоклеточной жизни. Pikaia gracilens (на фото) это переходное ископаемое между беспозвоночными и позвоночными,[45] и может быть самым ранним известным хордовый.[46][47] В этом смысле он мог быть изначальным предком рыб. Это было примитивное существо без видимых глаз, без четко очерченной головы и длиной менее 2 дюймов (5 сантиметров). Пикайя Это уплощенное вбок животное в форме листа, которое плавает, бросая свое тело в серию S-образных зигзагообразных изгибов, похожих на движения змей. Рыбы унаследовали то же движение плавания, но, как правило, у них более жесткий позвоночник. У него была пара больших головных щупалец и ряд коротких придатков, которые могут быть связаны с жаберными щелями по обе стороны от его головы. Пикайя показывает основные предпосылки для позвоночные. Уплощенное тело делится на пары сегментированные мышечные блоки, видимые как слабые вертикальные линии. Мышцы лежат по обе стороны гибкой структуры, напоминающей стержень, идущий от кончика головы к кончику хвоста.[48] | ||||
 | Haikouichthys | Haikouichthys (рыба из Хайкоу) - еще один род, который также упоминается в летописи окаменелостей около 530 млн лет назад и также отмечает переход от беспозвоночных к позвоночным.[7] Хайкоуичтис краниаты (животные с позвоночником и отчетливой головой). В отличие от ПикайяУ них были глаза. У них также был определенный череп и другие характеристики, которые привели к палеонтологи чтобы назвать ее настоящей черепной коробкой, и даже чтобы она была широко охарактеризована как одна из самых ранних рыб. Кладистический анализ указывает на то, что животное, вероятно, является базальным хордовый или базальный черепной;[49] но он не обладает достаточными функциями, которые будут включен неоспоримым даже в любом стволовая группа.[50][7] | ||||
| Myllokunmingia | Myllokunmingia - род, появившийся около 530 млн лет назад. Это хордовые, и было высказано мнение, что это позвоночное животное,[8] Он имеет длину 28 мм и высоту 6 мм и является одним из старейших черепных черепов. | |||||
| Конодонт | Конодонты (конические зубы) напоминали примитивных угрей. Они появились 495 млн лет и были уничтожены 200 млн лет назад.[24] Первоначально они были известны только по зубчатым микрофоссилиям, названным конодонтовые элементы. Эти «зубы» по-разному интерпретировались как устройства для подачи фильтра или как «захватывающий и дробящий набор».[25] Конодонты имели длину от сантиметра до 40 см. Промиссум.[25] Их большие глаза расположены сбоку, что делает маловероятным их хищническую роль. Сохранившаяся мускулатура намекает на то, что некоторые конодонты (Промиссум по крайней мере) были эффективными крейсерами, но неспособными развивать скорость.[25] В 2012 г. исследователи классифицировали конодонты в филюм Хордовые на основе их плавников с плавниковыми лучами, шеврон -образные мышцы и нотохорда.[26] Некоторые исследователи считают их позвоночными, внешне похожими на современных. миксина и миноги,[27] хотя филогенетический анализ показывает, что они больше полученный чем любая из этих групп.[28] | ||||
 | Остракодермы | Остракодермы (без кожуры) представляют собой одну из нескольких групп вымерших примитивных бесчелюстных рыб, которые были покрыты доспехом из костных пластин. Они появились в Кембрийский, около 510 миллионов лет назад, и стал вымерший ближе к концу Девонский около 377 миллионов лет назад. Первоначально у остракодермы плавники были плохо сформированы, и плавники, или конечности, впервые появились в этой группе. Они были покрыты костяной броней или чешуей и часто были менее 30 см (12 дюймов) в длину. | ||||
| Ордов- ician | Ордовик (485–443 млн лет назад): Рыбы, первые настоящие позвоночные в мире, продолжали развиваться, а те, у кого есть челюсти (Гнатостомы ), возможно, впервые появился в конце этого периода. Жизнь на суше еще не изменилась. | |||||
 | Арандаспис | Арандаспис - бесчелюстные рыбы, жившие в ранний ордовик, около 480–470 млн лет. Он был около 15 см (6 дюймов) в длину, с обтекаемым корпусом, покрытым рядами шишковидной брони. щитки. Передняя часть тела и голова были защищены твердыми пластинами с отверстиями для глаз, ноздрей и жабры. Хотя это было без челюсти, Арандаспис Возможно, у него во рту были подвижные тарелки, служившие губами, всасывающие частицы пищи. Низкое положение его пасти предполагает, что он добывал пищу на дне океана. У него не было плавников, и его единственным средством передвижения был горизонтально сплющенный хвост. В результате он, вероятно, плавал так же, как современный головастик.[51] | ||||
 | Астраспис | Астраспис (звездный щит) это вымерший род примитивных бесчелюстных рыб, родственных другим ордовикским рыбам, таким как Сакабамбаспис и Арандаспис. Окаменелости ясно показывают сенсорную структуру (система боковой линии). Расположение этих органов в виде правильных линий позволяет рыбе определять направление и расстояние, с которого происходит волнение в воде. Арандаспис Считается, что у них был подвижный хвост, покрытый небольшими защитными пластинами, и область головы, покрытая пластинами большего размера. Образец, описанный Сансом и другие. имели относительно большие боковые глаза и серию из восьми жабры проемы по бокам.[52] | ||||
| Птераспидоморфы | Птераспидоморфы вымерший класс ранних бесчелюстных рыб. Окаменелости показывают обширную защиту головы. У многих были гипоцеркальные хвосты, которые создавали подъемную силу для облегчения передвижения по воде их бронированных тел, которые были покрыты дермальной костью. У них также были сосущие части рта, а некоторые виды могли жить в пресной воде. | |||||
| Телодонты | Телодонты (зубы соска) представляют собой класс маленьких вымерших бесчелюстных рыб с характерной чешуей вместо больших пластин брони. Ведутся споры о том, представляют ли они монофилетическая группировка, или несопоставимые стволовые группы к основным линиям беззубый и челюстная рыба.[53] Телодонтов объединяет характерная «телодонтовая чешуя». Этот определяющий символ не обязательно является результатом общего происхождения, как это могло быть развивались независимо разными группами. Таким образом, обычно считается, что телодонты представляют полифилетическую группу.[54] Если они монофилетичны, нет никаких убедительных доказательств того, каково было их наследственное состояние.[55] Эти чешуйки легко рассеялись после смерти; их небольшой размер и устойчивость делают их наиболее распространенными окаменелостями позвоночных своего времени.[56][57] Рыба жила как в пресноводной, так и в морской среде, впервые появившись во время Ордовик, и гибну во время Франско-фаменское вымирание позднего Девонский. Они были преимущественно донными обитателями, питавшимися отложениями, хотя некоторые виды могли быть пелагическими. | |||||
| Ордовик закончился Ордовикско-силурийское вымирание (450–440 млн лет). Произошли два события, в результате которых погибло 27% всех семейств, 57% всех родов и от 60% до 70% всех видов.[58] Вместе они оцениваются многими учеными как вторые по величине из пяти крупных вымираний в истории Земли с точки зрения процента от роды которые вымерли. | ||||||
| Силурийский | Силурийский (443–419 млн лет. | |||||
 | Хотя традиционно девон называют веком рыб, недавние открытия показали, что силурийский период также был периодом значительной диверсификации. Челюстная рыба развита подвижные челюсти, адаптированный из опор передних двух-трех жаберные дуги | |||||
 | Анаспида | Анаспида (без щита) вымерший класс примитивных бесчелюстных позвоночных, живших во времена Силурийский и Девонский периоды.[59] Они классически считаются предками миноги.[60] Анаспиды были небольшими, преимущественно морскими агнатаны без тяжелого костяного щита и парных плавников, но сильно преувеличенные гипоцеркальный хвосты. Впервые они появились в Ранний силурийский период, и процветал до Позднее девонское вымирание,[61] где большинство видов, кроме миноги, вымерли. Необычно для Агнатан у анаспидов не было ни костяного щита, ни доспехов. Голова вместо этого покрыта множеством более мелких, слабо минерализованных чешуек.[62] | ||||
 | Остеостраки | Остеостраки («костлявые щиты») были классом безчелюстных рыб с костяными панцирями, которые жили с Средний силур к Поздний девон. Анатомически говоря, остеостраки, особенно девонские виды, были одними из самых продвинутых из всех известных бесчеловечных насекомых. Это связано с развитием парных плавников и их сложной анатомией черепа. Остеостраки были больше похожи на миноги чем у челюстных позвоночных, поскольку они имеют две пары полукружных каналов во внутреннем ухе, в отличие от трех пар, обнаруженных во внутреннем ухе челюстных позвоночных.[63] У большинства остеостраков был массивный головогрудный щит, но все виды среднего и позднего девона, по-видимому, имели уменьшенный, более тонкий и часто микромерный кожный скелет.[64] Вероятно, они были относительно хорошими пловцами, обладая спинные плавники, парный грудные плавники, и сильный хвост.[51] | ||||
 | Колючие акулы | Колючие акулы, более формально называемые «Акантоды» (с шипами), составляют класс Acanthodii. Впервые они появились в конце силурия ~ 420 млн лет назад и были одними из первых рыб, у которых появились челюсти. Они имеют общие черты с обоими хрящевые рыбы и костлявая рыба, но они не настоящие акулы, хотя и ведут к ним. Они вымерли до конца перми ~ 250 млн лет назад. Однако чешуя и зубы, относящиеся к этой группе, а также более производные челюстные рыбы, такие как хрящевые и костные рыбы, датируются Ордовик ~ 460 млн лет. Акантоды, как правило, были небольшими акулоподобными рыбами, от беззубых фильтраторов до зубатых хищников. Когда-то их часто относили к классу Плакодерми, но недавние авторитеты склонны относить скребней к парафилетическим сообществам, ведущим к современным хрящевым рыбам. Их различают в двух отношениях: они были самыми ранними из известных челюстных позвоночных, и у них были крепкие шипы поддерживая все их плавники, закрепленный на месте и неподвижный (как акула с спинной плавник ), важное защитное приспособление. Их окаменелости крайне редки. | ||||
 | Плакодермы | Плакодермы, (пластинчатая кожа), представляют собой группу панцирно-челюстных рыб класса Placodermi. Самые старые окаменелости появились в конце силурия и вымерли в конце девона. Недавние исследования показывают, что плакодермы, возможно, являются парафилетический группа базальный челюстные рыбы и ближайшие родственники всех ныне живущих челюстных позвоночных. Некоторые плакодермы были маленькими плоскими обитателями дна, например антиархи. Однако многие из них, особенно артродиры, были активными хищниками в разгар воды. Дунклеостей, который появился позже в девоне ниже, был самым большим и самым известным из них. Верхняя челюсть плотно срасталась с черепом, но между черепом и костной пластиной в области туловища имелся шарнирный сустав. Это позволяло запрокидывать верхнюю часть головы и, в артродиры, позволил им откусить больше. | ||||
 | Мегамастакс | Мегамастакс, (большой рот), это род из рыба с лопастными плавниками который жил в конце Силурийский период, ок. 423 миллион лет назад, в Китай. До открытия Мегамастакссчиталось, что челюстные позвоночные (гнатостомы ) были ограничены по размеру и вариациям до Девонский период. Мегамастакс известен только по костям челюсти и, по оценкам, достигал около 1 метра (3 фута 3 дюйма) в длину.[65] | ||||
 | Гуйю онейрос | Гуйю онейрос, самая ранняя из известных костистых рыб. Он сочетает в себе оба ребристый и ребристый особенности, хотя анализ совокупности его характеристик приближает его к рыбам с лопастными плавниками.[66][67][68] | ||||
| Андреолепис | Вымерший род Андреолепис включает самую раннюю из известных рыб с плавниками Андреолепис хедей, возникшие в позднем силуре около 420 млн лет назад.[69][70] | |||||
Девон: эра рыб
Масштаб оси: миллионы лет назад.
Девонский период делится на ранний, средний и поздний девон. К началу раннего девона 419 млн лет назад челюстные рыбы разделились на четыре отдельные клады: плакодермы и колючие акулы, оба из которых сейчас вымерли, а хрящевой и костлявые рыбы, оба из которых все еще существуют. Современная костистая рыба, класс Osteichthyes, появилось в конце Силурийский или ранний девон, около 416 миллионов лет назад. И хрящевые, и костные рыбы могли возникнуть либо от плакодерм, либо от колючих акул. Подкласс костистых рыб, рыбы с луговыми плавниками (Actinopterygii), стали доминирующая группа в постпалеозойском и современном мире - около 30 000 живых видов.
Уровень моря в девоне в целом был высоким. В морской фауне преобладали мшанки, разнообразный и богатый брахиоподы, загадочный гедереллоиды, микроконхид и кораллы. Лилиеподобный морские лилии были в изобилии, и трилобиты все еще были довольно распространены. Среди позвоночных бесчелюстные панцирные рыбы (остракодермы ) сократилось в разнообразии, в то время как челюстные рыбы (гнатостомы) одновременно увеличились как в море, так и в пресная вода. Бронированный плакодермы были многочисленны на более низких стадиях девонского периода, но вымерли в позднем девоне, возможно, из-за конкуренции за пищу с другими видами рыб. Ранний хрящевой (Chondrichthyes ) и костистая рыба (Osteichthyes ) также стали разнообразными и сыграли большую роль в девонских морях. Первый массовый род акул, Cladoselache, появился в океанах в девонский период. Большое разнообразие рыб в то время привело к тому, что девон получил в народной культуре название «Эпоха рыб».
Первый ребристый и ребристый костлявая рыба появились в девоне, а плакодермы начал доминировать почти во всех известных водных средах. Однако другой подкласс Osteichthyes, Саркоптеригии, включая рыбу с лопастными плавниками, в том числе латимерия и двоякодышащая рыба ) и четвероногие, была самой разнообразной группой костистых рыб в девоне. Саркоптериги в основном характеризуются внутренними ноздрями, лепестковыми плавниками, содержащими прочный внутренний скелет, и космоидные весы.
В среднем девоне 393–383 млн лет назад броненосный беззубый остракодерма разнообразие рыбы сокращалось; челюстные рыбы процветали и увеличивались в разнообразии как в океанах, так и в пресной воде. Мелкие, теплые, обедненные кислородом воды внутренних озер Девона, окруженные примитивными растениями, обеспечивали среду, необходимую некоторым ранним рыбам для развития таких важных характеристик, как хорошо развитые легкие и способность выползать из воды на сушу на долгое время. короткие периоды времени. Хрящевые рыбы, класс Chondrichthyes, состоящий из акулы, лучи и химеры, появившийся около 395 миллионов лет назад, в середине Девонский
В позднем девоне первые леса формировались на суше. Первые четвероногие появляются в летописи окаменелостей за период, начало и конец которого отмечены событиями вымирания. Так продолжалось до конца девона 359 млн лет назад. Предки всех четвероногие начали приспосабливаться к ходьбе по суше, их сильные грудные и тазовые плавники постепенно превратились в ноги (см. Тиктаалик ).[73] В океанах примитивный акулы стал более многочисленным, чем в Силурийский и поздний ордовик. Первый аммонит моллюски появившийся. Трилобиты, похожий на моллюска брахиоподы и великий коралловые рифы, все еще были обычным явлением.
В Позднее девонское вымирание произошел в начале последней фазы девона, фаменской фауны (граница фран-фамен), около 372,2 млн лет. Многие ископаемые Агнатан рыба, за исключением псаммостеид гетеростраки, последний раз появлялись незадолго до этого события. Кризис позднего девонского вымирания в первую очередь затронул морское сообщество и выборочно затронул мелководные теплые водные организмы, а не холодноводные. Наиболее важной группой, пострадавшей от этого вымирания, были строители рифов великих девонских рифовых систем.
Второй импульс гашения, Мероприятие Hangenberg закрыл девонский период и оказал драматическое влияние на фауну позвоночных. Плакодермы в основном вымерли во время этого события, как и большинство представителей других групп, включая рыб с лопастными плавниками, скребней и ранних четвероногих животных как в морских, так и в наземных средах обитания, оставив лишь несколько выживших. Это событие было связано с оледенением в умеренной и полярной зонах, а также с эвксинией и аноксией в морях.
| Девонский (419–359 млн лет назад): в начале девона впервые появились рыба с лопастными плавниками, предшественники четвероногие (животные с четырьмя конечностями). В этот период сформировались основные группы рыб, которых часто называют возраст рыбы.[74] Видеть Категория: Девонские рыбы. | ||||||
| D е v о п я а п | Рано Девонский | Ранний девон (419–393 млн лет назад): | ||||
|---|---|---|---|---|---|---|
 | Псаролепис | Псаролепис (пятнистая шкала) это род вымерших рыба с лопастными плавниками которые жили примерно от 397 до 418 млн лет. Окаменелости Псаролепис были найдены в основном на юге Китай и описан палеонтолог Сяобо Ю в 1998 году. Доподлинно неизвестно, какая группа Псаролепис принадлежит, но палеонтологи соглашаются, что это, вероятно, базальный род и, по-видимому, близок к общему предку лопастниковых и рыбы с луговыми плавниками.[75] | ||||
 | Холоптихий | Холоптихий вымерший род из порядок из поролеповидный рыба с лопастными плавниками, возрастом от 416 до 359 млн лет. Это был обтекаемый хищник длиной около 50 сантиметров (20 дюймов) (хотя он мог вырасти до 2,5 м), который питался другими костлявая рыба. Его округлая чешуя и форма тела указывают на то, что он мог быстро плыть по воде, чтобы поймать добычу.[76][77] Подобно другим Rhipidistians, у него был клык зубы на его вкус помимо более мелких зубов на челюстях. Его асимметричный хвост щеголял хвостовой плавник на его нижнем конце. Чтобы компенсировать нисходящий толчок, вызванный этим размещением плавников, Холоптихияs грудные плавники были помещены высоко на теле. | ||||
 | Птиктодонтида | В птиктодонтиды (клюв) вымершие монотипный порядок небронированного плакодермы, содержащий только одну семью. Они существовали от начала до конца девона. Птиктодонтиды с их большими головами, большими глазами и длинными телами очень напоминали современных химер (Голоцефали ). Их броня превратилась в узор из небольших пластинок вокруг головы и шеи. Как вымершие и родственные акантофоракиды, а также живых и неродственных голоцефалов, большинство птиктодонтид, как полагают, жили около морского дна и охотились на моллюски. | ||||
 | Petalichthyida | В Petalichthyida был порядок маленьких, сплющенных плакодермы которые существовали с начала девона до позднего девона. Их типом были расширенные плавники и многочисленные бугорки, украшавшие все пластины и чешуйки их доспехов. Они достигли пика разнообразия в раннем девоне и были обнаружены по всему миру. Поскольку у них были сжатые формы тела, предполагается, что они были обитателями дна, которые преследовали или устраивали засаду на более мелкую рыбу. Их диета не ясна, поскольку ни один из найденных окаменелостей не сохранил части рта. | ||||
 | Laccognathus | Laccognathus (ямка на челюсти) был родом амфибия рыба с лопастными плавниками, существовавшая 398–360 млн лет.[78] Они характеризовались тремя большими ямками (ямками) на внешней поверхности нижней челюсти, которые могли выполнять сенсорные функции.[79] Laccognathus вырос до 1–2 метров (3–7 футов) в длину. У них были очень короткие дорсовентрально сплюснутые головы, менее одной пятой длины тела.[80] Скелет имел такую структуру, что большие участки кожи натягивались на твердые костные пластинки. Эта кость состояла из особо плотных волокон - настолько плотных, что обмен кислорода через кожу был маловероятен. Скорее плотный окостенения служил для удержания воды внутри тела как Laccognathus путешествовал по суше между водоемами.[81] | ||||
| Середина Девонский | Средний девон (393–383 млн лет): Хрящевые рыбы, состоящий из акулы, лучи и химеры, появился около 395 млн лет назад. | |||||
| Диптерус | Диптерус (два крыла) вымерший род двоякодышащая рыба от 376–361 млн лет. Он был около 35 сантиметров (14 дюймов) в длину, в основном питался беспозвоночными, и имел легкие, а не воздушный пузырь. Как и его предок Dipnorhynchus на нёбе вместо настоящих зубов были зубчатые пластины. Однако в отличие от своих современных родственников, у которых спинной, каудальный, и анальный плавник слиты в одно, плавники у него еще разделились. Иначе Диптерус очень напоминал современных двоякодышащих рыб.[82] | |||||
 | Хейролепис | Хейролепис (ручной плавник) был родом рыбы с луговыми плавниками. Это был один из самых базальный из девонских рыб с лучевыми плавниками и считается первым, кто обладает «стандартными» кожными черепными костями, которые можно увидеть у более поздних рыб с лучевыми плавниками. Это была хищная пресноводная рыба длиной около 55 сантиметров (22 дюйма), и, судя по размеру ее глаз, она охотилась на зрение.[51] | ||||
 | Cladoselache | Cladoselache был первым массовым родом примитивных акул, появившимся около 370 млн лет назад.[83] Он вырос до 6 футов (1,8 м) в длину, с анатомическими особенностями, похожими на современные. макрель акула. У него было обтекаемое тело, почти полностью лишенное напольные весы, с пяти до семи жаберные щели и короткая закругленная морда с открытым ртом на передней части черепа.[83] У него был очень слабый челюстной сустав по сравнению с современными акулами, но он компенсировал это очень сильными мышцами, закрывающими челюсти. Его зубы были с множеством бугорков и гладкими краями, что делало их пригодными для хватания, но не для разрыва или жевания. Cladoselache поэтому, вероятно, схватил добычу за хвост и проглотил ее целиком.[83] У него были мощные кили, выходящие на боковую часть хвостового стебля, и полулунный хвостовой плавник, причем верхняя лопасть примерно того же размера, что и нижняя. Эта комбинация способствовала его скорости и ловкости, что было полезно при попытке перехитрить его вероятного хищника, тяжелобронированную 10-метровую рыбу-плакодерму Дунклеостей.[83] | ||||
| Коккостеус | Коккостеус (семенная кость) вымерший род артродир плакодерма. Большинство окаменелостей было найдено в пресноводных отложениях, хотя они могли попасть в соленую воду. Они выросли до 40 сантиметров (16 дюймов) в длину. Как и все другие артродиры, Коккостеус имел стык между панцирем тела и черепа. У него также был внутренний сустав между шеей. позвонки и задняя часть черепа, что позволяет ему открывать пасть еще шире. Наряду с более длинными челюстями это позволяло Коккостеус питаться довольно крупной добычей. Как и все другие артродиры, Коккостеус в его челюсти были встроены костные зубные пластины, образующие клюв. Клюв оставался острым за счет того, что края зубных пластин шлифовали друг о друга.[84] | |||||
 | Ботриолепис |
Ботриолепис (шкала без язв) был самым успешным родом антиарх плакодермы, если не самый успешный род любой плакодермы, с более чем 100 видами, распространенными в отложениях от среднего до позднего девона на всех континентах. | ||||
 | Питуриаспида | Питуриаспида (галлюциногенный щит) это класс, содержащий два причудливых вида бронированных беспощадных рыб с огромным носом трибуны. Они жили в устьях около 390 млн лет назад. В палеонтолог Гэвин Янг, назвал класс в честь галлюциногенного препарата Питури, так как он подумал, что у него могут быть галлюцинации при просмотре причудливых форм.[85] Более изученный вид выглядел как метательный дротик, с удлиненным щитом и копьевидным ростром. Другой вид выглядел как медиатор с хвостом, с меньшей и более короткой трибуной и более треугольным головным щитом. | ||||
| Позднее девонское вымирание: 375–360 млн лет. Продолжительная серия исчезновений уничтожила около 19% всех семейств, 50% всех родов.[58] и 70% всех видов. Это событие вымирания длилось, возможно, 20 млн лет, и есть свидетельства серии импульсов вымирания в этот период. | ||||||
| Поздно Девонский | Поздний девон (383–359 млн лет назад): | |||||
 | Дунклеостей |
Дунклеостей это род из артродир плакодермы существовавшие от 380 до 360 млн лет. Он вырос до 10 метров (33 фута) в длину.[86][87] и весил до 3,6 тонны.[88] Это было гиперплотоядный хищник вершины. Помимо современного Titanichthys (ниже) никакая другая плакодерма не могла сравниться с ней по размеру. Вместо зубов, Дунклеостей имел две пары острых костных пластинок, образующих клювовидную структуру. Помимо мегалодон, у него был самый сильный укус из всех рыб,[89] генерирует укусы в той же лиге, что и тиранозавр Рекс и современный крокодил.[90] | ||||
 | Titanichthys | Titanichthys это род гигантских аномальных морских плакодерма которые жили в мелководных морях. Многие виды подошли Дунклеостей по размеру и комплектации. Однако, в отличие от своего родственника, различные виды Титаничи у них были маленькие, малоэффективные на вид заглушки, у которых отсутствовала острая режущая кромка. Предполагается, что Titanichthys был питатель фильтра который использовал свой большой рот, чтобы глотать или вдыхать стайки маленьких, анчоусы -подобные рыбы, или возможно криль -подобно зоопланктон, и что ротовые пластины удерживали добычу, позволяя воде уйти, когда она закрывала свой рот. | ||||
 | Матерписцис |
Матерписцис (мать-рыба) это род из птиктодонтид плакодерма примерно от 380 млн лет. Известный только по одному экземпляру, он уникален тем, что еще не родился эмбрион присутствуют внутри и с замечательной сохранностью минерализованной структуры питания плаценты (пуповина ). Это делает Матерписцис первое известное позвоночное животное, показавшее живородство, или рожать, чтобы жить молодым.[91] Образец получил название Materpiscis attboroughi в честь Дэвид Аттенборо.[92] | ||||
 | Гинерия | Гинерия это род хищных рыб с лопастными плавниками, длиной около 2,5 м (8,2 фута), которые жили 360 миллионов лет назад.[93] | ||||
 | Ризодонты | Ризодонты были отрядом рыб с лопастными плавниками, дожившими до конца карбона, 377–310 млн лет. Они достигли огромных размеров. Самый крупный из известных видов, Rhizodus hibberti вырос до 7 метров в длину, что сделало его самой крупной из известных пресноводных рыб. | ||||
От рыбы до четвероногих

| От плавников до конечностей |
|---|
 Иллюстрация, показывающая, насколько необходимо изменить прикрепления задних конечностей у рыб с лопастными плавниками при переходе от рыб с лопастными плавниками (A) к ранним четвероногим (B), если рыба должна была стать наземным животным.  Сравнение плавников лопастоперых рыб и ног ранних четвероногих: 1. Тиктаалик 2. Пандерихтис 3. Евстеноптерон 4. Акантостега 5. Ихтиостега (задняя нога) |
Первый четвероногие - четвероногие, дышащие воздухом наземные животные, от которых произошли наземные позвоночные, включая человека. Они произошли от лопастоперых рыб клады. Саркоптеригии, появившись в прибрежных водах в среднем девоне и дав начало первым амфибии.[94]
Группа рыб с лопастными плавниками, которые были предками четвероногих, сгруппированы вместе как Рипидистия,[95] и первые четвероногие произошли от этих рыб за относительно короткий промежуток времени - 385–360 млн лет. Сами ранние группы четвероногих группируются как Лабиринтодонтия. Они сохранили водные, мальковидные головастики, система все еще встречается в современные амфибии. С 1950-х до начала 1980-х годов считалось, что четвероногие произошли от рыб, которые уже приобрели способность ползать по суше, возможно, чтобы они могли перейти из высыхающего бассейна в более глубокий. Однако в 1987 г. почти полные окаменелости Акантостега из примерно 363 Ма показал, что этот поздний девон переходный у животного были ноги, легкие и жабры, но оно никогда не могло выжить на суше: его конечности, запястья и голеностопные суставы были слишком слабыми, чтобы выдерживать его вес; его ребра были слишком короткими, чтобы его легкие не сжимались под его весом; его рыбоподобный хвостовой плавник мог быть поврежден при волочении по земле. Текущая гипотеза состоит в том, что Акантостега, который был около 1 метра (3,3 фута) в длину, был полностью водным хищником, который охотился на мелководье. Его скелет отличался от скелета большинства рыб тем, что позволяло ему поднимать голову, чтобы дышать воздухом, в то время как его тело оставалось под водой, в том числе: его челюсти показывают модификации, которые позволили бы ему глотать воздух; кости в задней части черепа соединены вместе, обеспечивая сильные точки прикрепления для мышц, поднимавших его голову; голова не соединяется с плечевой пояс и у него отчетливая шея.[96]

| Внешнее видео | |
|---|---|
1 2 3 4 5 | |
Распространение девонских наземных растений может помочь объяснить, почему дыхание воздухом было бы преимуществом: листья, падающие в ручьи и реки, способствовали бы росту водной растительности; это привлекло бы пасущихся беспозвоночных и мелкую рыбу, которая охотилась на них; они были бы привлекательной добычей, но окружающая среда была неподходящей для крупных морских хищных рыб; дыхание воздухом было бы необходимо, потому что в этих водах не хватало бы кислорода, так как теплая вода содержит меньше растворенного кислорода, чем более холодная морская вода, и поскольку разложение растительности потребовало бы часть кислорода.[96]
Существуют три основные гипотезы относительно того, как четвероногие развили свои короткие плавники (прото-конечности). Традиционное объяснение - это «гипотеза сужающегося водоема» или «гипотеза пустыни», выдвинутая американским палеонтологом. Альфред Ромер. Он полагал, что конечности и легкие могли развиться из-за необходимости находить новые водоемы по мере того, как высыхали старые водоемы.[98]
Вторая гипотеза - это «гипотеза приливов и отливов», выдвинутая в 2010 году группой польских палеонтологов во главе с Гжегожем Недзведским. Они утверждали, что саркоптериги, возможно, впервые появились на суше из приливные зоны а не внутренние водоемы. Их гипотеза основана на обнаружении следов Захелмие возрастом 395 миллионов лет в Захелмие, Польша, старейшие из когда-либо обнаруженных ископаемых останков четвероногих.[94][99]
Третья гипотеза, «гипотеза лесных угодий», была предложена американским палеонтологом Грегори Дж. Реталлаком в 2011 году. Он утверждает, что конечности могли развиться на мелководье в лесных массивах как средство навигации в среде, заполненной корнями и растительностью. Он основывал свои выводы на доказательствах того, что окаменелости переходных четвероногих постоянно обнаруживаются в местах обитания, которые ранее были влажными и лесными. поймы.[100]
Исследования Дженнифер А. Клак и ее коллеги показали, что самые ранние четвероногие животные, похожие на Акантостега, были полностью водными и совершенно непригодными для жизни на суше. Это контрастирует с более ранним представлением о том, что рыба сначала вторглась на сушу - либо в поисках добычи (как современные прыгуны ) или найти воду, когда пруд, в котором они жили, высох - а позже развились ноги, легкие и т. д.
Две идеи о гомология руки, руки и цифры существовали в течение последних 130 лет. Во-первых, цифры уникальны для четвероногих.[101][102] и во-вторых, что предшественники присутствовали в плавниках ранних саркоптерийский рыбы.[103] До недавнего времени считалось, что «генетические данные и данные окаменелостей подтверждают гипотезу о том, что цифры - это эволюционная новинка».[104]п. 640. Однако новое исследование, в котором была создана трехмерная реконструкция Пандерихтис, прибрежная рыба из Девонский Период 385 миллионов лет назад показывает, что у этих животных уже было много гомологичных костей, присутствующих в передних конечностях позвоночных с конечностями.[105] Например, у них было радиальный кости похожи на рудиментарные пальцы, но расположены в основании их плавников, напоминающих руки.[105] Таким образом, в эволюции четвероногие сдвиг так, что крайняя часть плавников была потеряна и в конечном итоге заменена ранними пальцами. Это изменение согласуется с дополнительными данными исследования актиноптериги, акулы и двоякодышащая рыба что пальцы четвероногих произошли от ранее существовавших дистальных радиусов, имеющихся у более примитивных рыб.[105][106] Споры по-прежнему существуют с Тиктаалик, позвоночное животное часто считается пропущенная ссылка между рыбами и наземными животными имел короткие, похожие на ноги конечности, без пальцевидных лучевых костей, найденных у Panderichthys. Исследователи статьи отметили, что «трудно сказать, означает ли это распределение символов, что Тиктаалик является аутапоморфный, что Panderichthys и четвероногие сходятся друг с другом, или что Panderichthys ближе к четвероногим, чем Tiktaalik. Во всяком случае, это демонстрирует, что переход рыба – четвероногие сопровождался значительным несовпадением признаков в функционально важных структурах ».[105]п. 638.
От конца девона до середины Каменноугольный в летописи окаменелостей наблюдается разрыв в 30 миллионов лет. Этот разрыв, называемый Разрыв ромера, отличается отсутствием ископаемых останков предков четвероногих и других позвоночных, которые выглядят хорошо приспособленными для жизни на суше.[107]
| Переход от рыб с лопастными плавниками к четвероногим | ||||||||
|---|---|---|---|---|---|---|---|---|
 ~385 Ма ~385 Ма | Евстеноптерон |
Род вымерший рыба с лопастными плавниками который приобрел культовый статус благодаря своим тесным отношениям с четвероногие. Ранние изображения этого животного показывают, что оно появилось на суше, однако теперь палеонтологи согласны с тем, что это было исключительно водное животное.[97] Род Евстеноптерон известен по нескольким видам, жившим в поздний девонский период, около 385 млн лет. Он был объектом интенсивных исследований с 1940-х по 1990-е годы. палеоихтиолог Эрик Ярвик.[108] | ||||||
 | Гогонас | Гогонас (морда от Гого) была рыба с лопастными плавниками, известная по сохранившимся в трехмерном виде окаменелостям возрастом 380 миллионов лет, найденным в Формация Гого. Это была небольшая рыба, достигающая 30-40 см (12-16 дюймов) в длину.[109] Его скелет имеет несколько черт, похожих на четвероногих. Они включали структуру своего среднее ухо, а его плавники показывают предшественников костей предплечья, радиус и локтевая кость. Исследователи полагают, что он использовал плавники, похожие на предплечья, чтобы выскочить из рифа и поймать добычу. Гогонас был впервые описан в 1985 г. Джон А. Лонг. Почти 100 лет Евстеноптерон был образцом для подражания для демонстрации этапов эволюции рыб с лопастными плавниками до четвероногих. Гогонас теперь заменяет Евстеноптерон как лучше сохранившийся представитель без какой-либо двусмысленности в интерпретации его анатомии. | ||||||
~ 385 млн лет | Пандерихтис | Адаптирован к илистому мелководью и способен к перемещению по мелководью или наземным движениям сгибания тела. Имел способность поддерживать себя.[110] У них были большие головы, похожие на четвероногих, и они, как полагают, являются наиболее обращенными к макушке стволовыми четвероногими рыбами с парными плавниками. | ||||||
~ 375 млн лет | Тиктаалик | Рыба с плавниками, похожими на конечности, которые могут вывести ее на сушу.[111] Это пример из нескольких линий древних саркоптерийских рыб, которые адаптировались к бедным кислородом мелководным местам обитания того времени, что привело к эволюции четвероногих.[96] Палеонтологи предполагают, что это представитель переход между позвоночными животными, не являющимися четвероногими (рыбами), такими как Пандерихтис, известные по окаменелостям возрастом 380 миллионов лет, и ранние четвероногие, такие как Акантостега и Ихтиостега, известный по окаменелостям возрастом около 365 миллионов лет. Его смесь примитивных рыб и унаследованных характеристик четвероногих привела одного из его первооткрывателей, Нил Шубин, чтобы охарактеризовать Тиктаалик как "рыбопод ".[112][113] | ||||||
 365 млн лет | Акантостега | Рано как рыба лабиринтодонт занявшие болота и изменившие представления о ранней эволюции четвероногих.[96] В нем было восемь цифр на каждой руке (количество пальцев на ступнях неясно) связаны перепонками, у него не было запястий, и в целом он был плохо приспособлен для выхода на сушу.[114] Последующие открытия выявили более ранние переходные формы между Акантостега и полностью рыбоподобные животные.[115] | ||||||
 374–359 млн лет | Ихтиостега |
До находок других ранних четвероногих и близкородственных рыб в конце 20 века, Ихтиостега стоял особняком как переходное ископаемое между рыбой и четвероногими, сочетая рыбий хвост и жабры с черепом и конечностями земноводного. У него были легкие и семь пальцев на конечностях, которые помогали ему перемещаться по мелководью в болотах. | ||||||
359–345 млн лет | Педерпес | Педерпес является самым ранним из известных полностью наземных четвероногих. Он включен сюда, чтобы завершить превращение рыб с лопастными плавниками в четвероногих, хотя Педерпес больше не рыба. | ||||||
К концу девона наземные растения стабилизировали пресноводные среды обитания, что позволило первым водно-болотное угодье экосистемы развиваться со все более сложными пищевые полотна это открыло новые возможности. Пресноводные среды обитания были не единственными местами, где можно было найти воду, наполненную органическими веществами и забитую растениями с густой растительностью у кромки воды. В то время также существовали болотистые места обитания, такие как мелкие водно-болотные угодья, прибрежные лагуны и большие солоноватоводные речные дельты, и есть много оснований предполагать, что именно в такой среде развивались четвероногие. Ранние ископаемые четвероногие были обнаружены в морских отложениях, и поскольку окаменелости примитивных четвероногих обычно разбросаны по всему миру, они, должно быть, распространились, следуя прибрежным линиям - они не могли жить только в пресной воде.
- Ископаемое освещает эволюцию конечностей от плавников Scientific American, 2 2 апреля 2004 г.
Пост девонский
- В Мезозойская эра началось около 250 миллионов лет назад в результате Пермско-триасовое событие, самый большой массовое вымирание в истории Земли и закончилась около 66 миллионов лет назад Меловое – палеогеновое вымирание, еще одно массовое вымирание, уничтожившее нептичьих динозавры, а также другие виды растений и животных. Его часто называют Возраст рептилий потому что рептилии были доминирующими позвоночными животными того времени. Мезозой стал свидетелем постепенного рифтинга суперконтинента. Пангея на отдельные участки суши. Климат чередовался между периодами потепления и похолодания; в целом Земля была горячее, чем сегодня.
- Мезозой увидел диверсификацию неоптериский рыб, клады, состоящие из голостейский и костистость Рыбы. Разнообразие форм тела в триасовом, юрском и раннемеловом периодах. неоптериский рыбы были зарегистрированы,[116] показывает, что накопление новых форм тела у костистых рыб было преимущественно постепенным в течение этого периода в 150 миллионов лет (250–100 млн лет назад). Кажется, что голостовые рыбы накапливают разнообразие форм тела (так называемое несоответствие) между ранний триас и Тоарский, после чего количество различий в их формах тела оставалось стабильным до конца Раннемеловой период.[116]
| Углерод бессмысленный | Каменноугольный (359–299 млн лет): Акулы прошел серьезный эволюционное излучение вовремя Каменноугольный.[117] Считается, что это эволюционное излучение произошло из-за упадка плакодермы в конце девонского периода вызвало много экологические ниши стать незанятым и позволить новым организмам развиваться и заполнять эти ниши.[117] | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Прибрежные моря в каменноугольном периоде c. 300 млн лет | Первые 15 миллионов лет в каменноугольном периоде осталось очень мало земных окаменелостей. Этот пробел в летописи окаменелостей называется Разрыв ромера после американского палеентолога Альфред Ромер. Хотя давно ведутся споры о том, является ли разрыв результатом окаменелости или связан с реальным событием, недавние исследования указывают на то, что в этот период наблюдалось падение уровня кислорода в атмосфере, что указывает на своего рода экологический коллапс.[118] Разрыв увидел кончину Девонский рыбоподобный ихтиостегальский лабиринтодонты и появление более продвинутых темноспондил и рептилиоморфан амфибии, которые так характерны для фауны наземных позвоночных каменноугольного периода. В Каменноугольный моря населяли многие рыбы, в основном Elasmobranchs (акулы и их родственники). К ним относятся некоторые, например Псаммод с дробящимися зубцами, похожими на мостовую, приспособленными для измельчения панцирей брахиопод, ракообразных и других морских организмов. У других акул были пронзительные зубы, такие как Symmoriida; некоторые, петалодонты, имел своеобразные циклоидные режущие зубы. Большинство акул были морскими, но Ксенакантида вторглись в пресные воды угольных болот. Среди костлявая рыба, то Palaeonisciformes обнаруженные в прибрежных водах, также, по-видимому, мигрировали в реки. Саркоптерийский рыба также была видна, и одна группа, Ризодонты, достиг очень большого размера. Большинство видов морских рыб каменноугольного периода были описаны в основном по зубам, шипам плавников и кожным косточкам, а пресноводные рыбы меньшего размера были сохранены целиком. Пресноводные рыбы были многочисленны и включают роды Ctenodus, Уронемус, Акантоды, Cheirodus, и Гиракант. | |||||||||
 | Stethacanthidae |  В результате эволюционного излучения каменноугольные акулы принимали самые разные причудливые формы, в том числе акулы этого семейства. Stethacanthidae, который обладал плоским щеткообразным спинным плавником с пятном зубчики на его вершине.[117] Стетакантус ' необычный плавник, возможно, использовался в брачных ритуалах.[117] Помимо плавников, Stethacanthidae напоминали Falcatus (ниже). | ||||||||
 | Falcatus | Falcatus это род малых кладодонт зубастые акулы, жившие 335–318 млн лет. Они были около 25–30 см (10–12 дюймов) в длину.[119] Для них характерны выступающие шипы плавников, загнутые вперед над головой. | ||||||||
 | Ородус | Ородус еще одна акула каменноугольного периода, род из семейства Orodontidae жившие в ранней перми с 303 по 295 млн лет. Он вырос до 2 м (6,5 футов) в длину. | ||||||||
| Пермский период | Пермский период (298–252 млн лет назад): | |||||||||
 | Акантоды | Акантоды вымерший род колючая акула.[120] Это было жабры но нет зубы,[121] и предположительно был питатель фильтра.[51] Акантоды имел только две кости черепа и был покрыт чешуей кубической формы. Каждая пара грудной и тазовые плавники был один позвоночник, как и одиночный анальный и спинные плавники Всего у него шесть шипов, что вдвое меньше, чем у многих других колючих акул.[51] Акантоды имеют общие качества как костных рыб (остеихти), так и хрящевых рыб (хондрихтисы), и было высказано предположение, что они могли быть стеблевыми хондрихтианами и стеблевыми гнатостомами.[122][123] | ||||||||
| Пермь завершилась самым обширным событие вымирания записано в палеонтология: the Пермско-триасовое вымирание. От 90% до 95% морских видов стали вымерший, а также 70% всех наземных организмов. Это также единственное известное массовое исчезновение насекомых.[124][125] Восстановление после пермо-триасового вымирания было длительным; для восстановления наземных экосистем потребовалось 30 миллионов лет,[126] а морские экосистемы заняли еще больше времени.[58] | ||||||||||
| Триасовый | Триасовый (252–201 млн лет назад): ихтиофауна триаса была удивительно однородной, что отражает тот факт, что очень немногие семьи пережили пермское вымирание. Значительное излучение рыбы с луговыми плавниками произошел в триасовый период, положив начало появлению многих современных рыб.[127] Видеть Категория: Триасовые рыбы. | |||||||||
 | Perleidus | Perleidus был рыба с плавниками из раннего триаса. Примерно 15 сантиметров (6 дюймов) в длину, это была пресноводная хищная рыба с челюстями, свисающими вертикально под черепной коробкой, что позволяло им широко открываться. Perleidus имел очень гибкий спинной и анальные плавники, с уменьшенным количеством плавниковых лучей, что сделало бы рыбу более подвижной в воде.[51] | ||||||||
 | Толстокожие |  Толстокожие - вымерший отряд лучевых плавниковых рыб, существовавший от среднего триаса до K-Pg вымирание (ниже). Для них характерны зазубренные грудные плавники, уменьшенные брюшные плавники и костный рострум. Их отношения с другими рыбами неясны. | ||||||||
 | Pholidophorus | Pholidophorus был вымерший род костистость, около 40 сантиметров (16 дюймов) в длину, примерно 240–140 млн лет назад. Хотя и не очень близок к современному сельдь, это было чем-то похоже на них. Был сингл спинной плавник, симметричный хвост и анальный плавник размещен ближе к задней части тела. У него были большие глаза, и он, вероятно, был быстро плавающим хищником, охотился планктонный ракообразные и более мелкую рыбу.[128] Очень рано костистость, Pholidophoris имел много примитивных характеристик, таких как ганоидная чешуя и позвоночник который частично состоял из хрящ, скорее, чем кость.[128] | ||||||||
| Триас закончился Триасово-юрское вымирание. Вымерло около 23% всех семейств, 48% всех родов (20% морских семейств и 55% морских родов) и от 70% до 75% всех видов.[129] Архозавры нединозавров продолжали доминировать в водной среде, в то время как диапсиды неархозавров продолжал доминировать в морской среде.[129] | ||||||||||
| Юрский | Юрский (201–145 млн лет назад): во время Юрский период, основными позвоночными животными, обитавшими в морях, были рыбы и морской рептилии. К последним относятся ихтиозавры которые были на пике своего разнообразия, плезиозавры, плиозавры, и морской крокодилы семей Телеозавры и Metriorhynchidae.[130] Многочисленные черепахи можно найти в озерах и реках.[131][132] Видеть Категория: Юрские рыбы. | |||||||||
 | Лидсихтис | Наряду со своими близкими родственниками пахикормид Боннерихтис и Ринконихтис, Лидсихтис является частью крупной фильтраторы которые плавали в мезозойских морях более 100 миллионов лет, от средней юры до конца Меловой период. Пахикормиды могут представлять раннюю ветвь Teleostei, группа самых современных костистых рыб; в таком случае Лидсихтис - самая крупная из известных костистых рыб.[133] В 2003 году был обнаружен ископаемый образец длиной 22 метра (72 фута).[134] | ||||||||
 | Ichthyodectidae |  Эта ископаемая рыба Ichthyodectidae из нижней юры - одна из наиболее хорошо сохранившихся ископаемых рыб в мире. Семья Ichthyodectidae (буквально «кусающие рыбу») были семьей морских актиноптеригий рыбы. Впервые они появились 156 млн лет назад. Поздняя юра и исчез во время вымирания K-Pg 66 млн лет назад. Они были самыми разнообразными на протяжении мелового периода. Иногда классифицируется как примитив костлявая рыба порядок Толстокожие, сегодня они обычно считаются членами отряда «рыб-бульдогов» Ichthyodectiformes в гораздо более продвинутых Остеоглоссоморфный. Большинство ихтиодектид имели длину от 1 до 5 метров (от 3,5 до 16,5 футов). Все известные таксоны мы хищники, питаясь мелкой рыбой; в некоторых случаях более крупные Ichthyodectidae охотились на более мелких членов семейства. У некоторых видов были удивительно большие зубы, а у других, например, Gillicus arcuatus, имели маленьких и засасывали свою добычу. Самый большой Ксифактин был 20 футов в длину и появился в позднем меловом периоде (внизу). | ||||||||
| Cret- крутой | Меловой (145–66 млн лет): Видеть Категория: Меловая рыба. | |||||||||
| Осетр | Истинный осетровые появляются в летописи окаменелостей в верхнем меловом периоде. С тех пор осетровые испытали на удивление мало морфологический изменения, указывающие на то, что их эволюция была исключительно медленной, и зарабатывающие на них неформальный статус как живые окаменелости.[135][136] Отчасти это объясняется большим интервалом генерации, допуском к широкому диапазону температура и соленость, отсутствие хищники из-за размеров и обилия предметов добычи в бентосный среда. | |||||||||
 | Кретоксирина | Cretoxyrhina mantelli был большим акула которые жили от 100 до 82 миллионов лет назад, в середине мелового периода. Она широко известна как акула Джинсу. Впервые эту акулу идентифицировал известный швейцарский натуралист, Луи Агассис в 1843 г., как Cretoxyhrina mantelli. Однако наиболее полный образец этой акулы был обнаружен в 1890 году охотником за окаменелостями Чарльзом Х. Штернбергом, который опубликовал свои результаты в 1907 году. Образец состоял из почти полного связанного позвоночного столба и более 250 связанных зубов. Такая исключительная сохранность ископаемых акул встречается редко, потому что скелет акулы состоит из хрящ, не склонный к окаменению. Чарльз окрестил образец Oxyrhina mantelli. Этот экземпляр представлял собой акулу длиной 20 футов (6,1 м). | ||||||||
 | Энход | Энход вымерший род костистых рыб. Он процветал в верхнем меловом периоде и был размером от маленького до среднего. Одним из наиболее примечательных атрибутов этого рода являются большие «клыки» на передней части верхней и нижней челюстей и на небных костях, что привело к его ошибочному прозвищу среди охотников за окаменелостями и палеоихтиологов «саблезубая сельдь». Эти клыки, наряду с длинным гладким телом и большими глазами, предполагают Энход был хищным видом. | ||||||||
 | Ксифактин |
Ксифактин - вымерший род крупных хищных морских костистых рыб позднего мела. Они выросли более чем на 4,5 метра (15 футов) в длину.[137] | ||||||||
 | Птиход | Птиход это род вымерших гибодонтиформный акула, жившая с позднего мела до палеогена.[138][139] Ptychodus mortoni (на фото) был около 32 футов (9,8 метра) в длину и был обнаружен в Канзас, Соединенные Штаты.[140] | ||||||||
| Конец мелового периода был отмечен Меловое – палеогеновое вымирание (Вымирание K-Pg). Есть существенные ископаемые летописи челюсть Рыбы через границу K – T, что является хорошим свидетельством закономерностей вымирания этих классов морских позвоночных. В хрящевые рыбы, примерно 80% акулы, лучи, и коньки семьи пережили вымирание,[141] и более 90% костистая рыба (костлявые рыбы) семьи выжили.[142] Имеются свидетельства массовой гибели костистых рыб на ископаемом участке непосредственно над пограничным слоем K – T на Остров Сеймур вблизи Антарктиды, по-видимому, вызванный K – Pg вымиранием.[143] Однако морская и пресноводная среда обитания рыб смягчила экологические последствия вымирания,[144] и данные показывают, что сразу после исчезновения резко увеличились размеры и численность костистых особей, по-видимому, из-за исчезновения их аммонит конкурентов (аналогичных изменений в популяциях акул по другую сторону границы не было).[145] | ||||||||||
| Кайнозойский Эра | Кайнозойская эра (66 млн лет по настоящее время): В нынешнюю эпоху наблюдается большое разнообразие костных рыб. Более половины всех ныне живущих видов позвоночных (около 32000 видов) - это рыбы (не четвероногие черепные), разнообразный набор линий, населяющих все водные экосистемы мира, от снежных гольянов (Cypriniformes) в Гималайских озерах на высоте более 4600 метров (15 100 футов) к камбалам (отряд Pleuronectiformes) в Глубине Челленджера, самой глубокой океанской траншеи на высоте около 11000 метров (36000 футов). Множество видов рыб являются основными хищниками в большинстве водоемов мира, как пресноводных, так и морских. | |||||||||
 | Амфистиум | Амфистиум Это ископаемая рыба возрастом 50 миллионов лет, которая была определена как ранний родственник камбалы и переходное ископаемое.[146] У типичной современной камбалы голова асимметрична, и оба глаза находятся на одной стороне головы. В Амфистиум, переход от типичной симметричной головы позвоночного животного является неполным, с одним глазом, расположенным ближе к макушке.[147] | ||||||||
 | Мегалодон |
Мегалодон вымерший вид акул, живший примерно от 28 до 1,5 млн лет. Это было похоже на коренастую версию Большая белая акула, но был намного больше, длина окаменелостей достигала 20,3 метра (67 футов).[148] Найдено во всех океанах[149] это был один из самых крупных и могущественных хищников в истории позвоночных,[148] и, вероятно, оказали глубокое влияние на морская жизнь.[150] | ||||||||
Доисторическая рыба
| Внешнее видео | |
|---|---|
Доисторические рыбы - это ранние рыбы, которые известны только из ископаемое записи. Они самые ранние из известных позвоночные, и включают первую и вымершую рыбу, пережившую Кембрийский к Третичный. Изучение доисторических рыб называется палеоихтиология. Несколько живых форм, таких как латимерия также называются доисторическими рыбами или даже живые окаменелости из-за их нынешней редкости и сходства с вымершими формами. Рыбы, ставшие недавно вымерший обычно не относятся к доисторическим рыбам.
Живые окаменелости

Бесчелюстные рыбы
Костные рыбы
- Арована и Арапайма
- Bowfin
- Латимерия
- Гар
- Квинслендская двоякодышащая рыба
- Protanguilla palau (угорь)
- Осетровые и веслонос
- Бичир
Акулы
В латимерия считалось, что вымерли 66 миллион лет назад, пока не появится живой экземпляр, принадлежащий порядок был обнаружен в 1938 году у побережья Южной Африки.[152][153]
Ископаемые участки

Некоторые окаменелости, давшие примечательные окаменелости рыб
- Эбби Вуд УОНИ
- Кровати Bracklesham
- Известняк Медвежьего Ущелья
- Burgess Shale
- Canowindra
- Крато Формирование
- Dura Den
- Формация Фелтвилл
- Национальный памятник Фоссил Бьютт
- Формирование меха
- Формация Гого
- Green's Creek
- Формация Грин Ривер
- Провинциальный парк Каква
- Land Grove Quarry
- Маотяншаньские сланцы
- Матанусская свита
- Ископаемые кровати McAbee
- Национальный парк Мигуаша
- MoClay
- Монте Болка
- Mount Ritchie
- Оркадский бассейн
- От пирса Портисхед до УОНИ Блэк Нор
- Формация Сантана
- Саутерхэм Грей Пит
- Формация Танет
- Формация Товако
- Weydale
- Чжоукоудянь
Коллекции ископаемых
| Часть серии по |
| Палеонтология |
|---|
 |
История палеонтологии |
Отрасли палеонтологии |
| Палеонтологический портал Категория |
Некоторые известные коллекции ископаемых рыб.
- Коллекция ископаемых рыб Музей естественной истории, Великобритания.
- Сбор и экспертиза Museum für Naturkunde, Германия.
- Ископаемые рыбы Полевой музей, Соединенные Штаты.
Палеоихтиологи
Палеоихтиология - это научное исследование доисторической жизни рыб. Ниже перечислены некоторые исследователи, внесшие заметный вклад в палеоихтиологию.
- Луи Агассис
- Мэри Эннинг
- Майкл Бентон
- Дерек Бриггс
- Ханс К. Бьерринг
- Джон Сэмюэл Бюджетт
- Фредерик Чепмен
- Дженни Клак
- Тед Даешлер
- Башфорд Дин
- Роберт Дик
- Филип Грей Эгертон
- Эдвин Шербон Хиллз
- Джеффри А. Хатчингс
- Томас Генри Хаксли
- Йохан Ашехуг Кюр
- Филипп Жанвье
- Эрик Ярвик
- Джордж В. Лаудер
- Джон А. Лонг
- Хью Миллер
- Чарльз Мур
- Пол Э. Олсен
- Хайнц Кристиан Пандер
- Элизабет Филпот
- Жан Пивето
- Колин Паттерсон
- Альфред Ромер
- Ира Рубинофф
- Нил Шубин
- Франц Штайндахнер
- Эрик Стенсио
- Рамзи Хитли Трэкэр
- Томас Стэнли Уэстолл
- Тиберий Корнелис Винклер
- Артур Смит Вудворд
Смотрите также
- Сравнительная анатомия
- Конвергентная эволюция у рыб
- Эволюция парных плавников
- Ихтиолит
- Список лет в палеонтологии
- Старый красный песчаник
- Пародии на символ ихтиса
- Доисторическая жизнь
- Прогулочная рыба - рыба с чертами четвероногих
- Палеонтология позвоночных
Рекомендации
Цитаты
- ^ Лекуантр и Ле Гюядер 2007
- ^ Бентон, М. Дж. (2005) Палеонтология позвоночных В архиве 2020-06-09 в Wayback Machine Джон Вили, 3-е издание, стр. 14. ISBN 9781405144490.
- ^ Ромер 1970.
- ^ а б Докинз 2004, п. 357.
- ^ Гевин, В. (2005). «Функциональная геномика сгущает биологический сюжет». PLOS Биология. 3 (6): e219. Дои:10.1371 / journal.pbio.0030219. ЧВК 1149496. PMID 15941356.
- ^ Геном ланцетника (амфиоксуса) и происхождение позвоночных В архиве 2016-03-04 в Wayback Machine Ars Technica, 19 июня 2008 г.
- ^ а б c Shu, D.-G .; Conway Morris, S .; Han, J .; и другие. (Январь 2003 г.). "Голова и позвоночник раннего кембрийского позвоночного Haikouichthys". Природа. 421 (6922): 526–529. Bibcode:2003Натура.421..526С. Дои:10.1038 / природа01264. PMID 12556891. S2CID 4401274.
- ^ а б Шу, Д-Г .; и другие. (4 ноября 1999 г.). «Нижнекембрийские позвоночные из южного Китая». Природа. 402 (6757): 42–46. Bibcode:1999Натура 402 ... 42S. Дои:10.1038/46965. S2CID 4402854.
- ^ Ваггонер, Бен. «Позвоночные животные: летопись окаменелостей». UCMP. В архиве из оригинала 29 июня 2011 г.. Получено 15 июля 2011.
- ^ Haines & Chambers 2005.
- ^ Британская энциклопедия 1954 г., п. 107.
- ^ Берг 2004, п. 599.
- ^ "агнатан". Оксфордский словарь английского языка (Интернет-ред.). Издательство Оксфордского университета. (Подписка или членство участвующего учреждения требуется.)
- ^ Бейкер, Клэр В.Х. (2008). «Эволюция и разработка клеток нервного гребня позвоночных». Текущее мнение в области генетики и развития. 18 (6): 536–543. Дои:10.1016 / j.gde.2008.11.006. PMID 19121930.
- ^ Пурнелл, М.А. (2001). Дерек Э. Г. Бриггс и Питер Р. Кроутер (ред.). Палеобиология II. Оксфорд: Blackwell Publishing. п. 401. ISBN 978-0-632-05149-6.
- ^ Чжао Вэнь-Цзинь; Чжу Минь (2007). «Диверсификация и фаунистический сдвиг силуро-девонских позвоночных Китая». Геологический журнал. 42 (3–4): 351–369. Дои:10.1002 / gj.1072. Архивировано из оригинал на 2013-01-05.
- ^ Сансом, Роберт С. (2009). «Филогения, классификация и полярность характера остеостраков (позвоночных)». Журнал систематической палеонтологии. 7: 95–115. Дои:10.1017 / S1477201908002551. S2CID 85924210. В архиве из оригинала от 19.10.2012. Получено 2013-01-16.
- ^ Маллатт Дж. И Дж. Салливан. 1998. (1998). «Последовательности 28S и 18S рибосомной ДНК поддерживают монофилию миног и миксин». Молекулярная биология и эволюция. 15 (12): 1706–1718. Дои:10.1093 / oxfordjournals.molbev.a025897. PMID 9866205.CS1 maint: несколько имен: список авторов (связь)
- ^ ДеЛарбре Кристиан; Галлут Кирилл; Барриэль Вероник; Жанвье Филипп; Гачелин Габриэль (2002). «Полная митохондриальная ДНК миксины, Eptatretus burgeri: сравнительный анализ последовательностей митохондриальной ДНК убедительно подтверждает монофилию циклостома». Молекулярная филогенетика и эволюция. 22 (2): 184–192. Дои:10.1006 / mpev.2001.1045. PMID 11820840.
- ^ Жанвье, П. 2010. «МикроРНК возрождают старые представления о дивергенции и эволюции бесчелюстных позвоночных». Труды Национальной академии наук (США) 107: 19137-19138. [1] В архиве 2015-09-24 на Wayback Machine "Хотя я был одним из первых сторонников парафилии позвоночных, я впечатлен доказательствами, предоставленными Heimberg et al. и готовы признать, что циклостомы на самом деле монофилетичны. Следствием этого является то, что они могут сказать нам немного, если вообще что-нибудь, о заре эволюции позвоночных, за исключением того, что интуиция зоологов 19-го века была правильной, предполагая, что эти странные позвоночные (в частности, миксины) сильно выродились и потеряли многие признаки из-за время."
- ^ а б Бентон, М. Дж. (2005) Vertebrate Palaeontology, Blackwell, 3-е издание, рис. 3.25 на странице 73.
- ^ Жанвье, Филипп (1997) Позвоночные. Животные с позвоночником В архиве 2013-03-12 в Wayback Machine. Версия от 01 января 1997 г. в Веб-проект "Древо жизни" В архиве 2011-05-15 на Wayback Machine
- ^ Паттерсон, Колин (1987). Молекулы и морфология в эволюции: конфликт или компромисс?. Кембридж, Великобритания: Издательство Кембриджского университета. п. 142. ISBN 978-0-521-32271-3. В архиве из оригинала от 24.06.2016.
- ^ а б Де Ренци М, Будоров К, Сударь М (1996). «Вымирание конодонтов в терминах дискретных элементов на границе триаса и юры». Cuadernos de Geología Ibérica. 20: 347–364. В архиве из оригинала на 2016-08-06. Получено 2013-01-20.
- ^ а б c d е ж Gabbott, S.E .; Р. Дж. Олдридж; Дж. Н. Терон (1995). «Гигантский конодонт с сохранившейся мышечной тканью из верхнего ордовика Южной Африки». Природа. 374 (6525): 800–803. Bibcode:1995Натура 374..800Г. Дои:10.1038 / 374800a0. S2CID 4342260.
- ^ а б Бриггс, Д. (май 1992 г.). «Конодонты: основная вымершая группа, добавленная к позвоночным». Наука. 256 (5061): 1285–1286. Bibcode:1992Научный ... 256.1285Б. Дои:10.1126 / science.1598571. PMID 1598571.
- ^ а б Милсом, Клэр; Ригби, Сью (2004). «Позвоночные». Окаменелости с первого взгляда. Виктория, Австралия: Blackwell Publishing. п. 88. ISBN 978-0-632-06047-4.
- ^ а б Donoghue, P.C.J .; Forey, P.L .; Олдридж, Р.Дж. (2000). «Конодонтовое родство и хордовые филогении». Биологические обзоры. 75 (2): 191–251. Дои:10.1111 / j.1469-185X.1999.tb00045.x. PMID 10881388. S2CID 22803015. В архиве из оригинала 13.04.2020. Получено 2008-04-07.
- ^ Бентон 1999, п. 44.
- ^ Мэйси, Джон Г. (1996). Обнаружение ископаемых рыб (иллюстрированный ред.). Нью-Йорк: Генри Холт и компания. п.37.
- ^ Конструкция челюсти позвоночного заблокирована рано В архиве 2012-09-08 в Wayback Machine
- ^ Davis, S .; Finarelli, J .; Коутс, М. (2012). «Акантоды и акулоподобные состояния у последнего общего предка современных гнатостомов». Природа. 486 (7402): 247–250. Bibcode:2012Натура.486..247D. Дои:10.1038 / природа11080. PMID 22699617. S2CID 4304310. В архиве с оригинала 25 марта 2019 г.. Получено 25 марта 2019.
- ^ «Морда древней рыбы показывает корни современной челюсти». Природа. 25 сентября 2013 г. В архиве с оригинала 31 октября 2013 г.. Получено 26 сентября, 2013.
- ^ Мередит Смит, Моя; Кларк, Бретт; Гуже, Даниэль; Йохансон, Зерина (2017-08-17). «Эволюционное происхождение зубов у челюстных позвоночных: противоречивые данные по пластинам аканторацида ('Placodermi')». Палеонтология. 60 (6): 829–836. Дои:10.1111 / pala.12318. ISSN 0031-0239.
- ^ Бентон, М. Дж. (2005) Vertebrate Palaeontology, Blackwell, 3-е издание, рис. 7.13 на странице 185.
- ^ Чжу, Мин; Сяобо Ю; Пер Эрик Альберг; Брайан Чу; Цзин Лу; Туо Цяо; Цинмин Цюй; Венцзинь Чжао; Ляньтао Цзя; Хеннинг Блом; Юань Чжу (2013). «Силурийская плакодерма с остеихтиановыми краевыми костями челюсти». Природа. 502 (7470): 188–193. Bibcode:2013Натура.502..188Z. Дои:10.1038 / природа12617. PMID 24067611. S2CID 4462506.
- ^ Хельфман и другие 2009, п. 198.
- ^ Аллен, Г. Р., С. Х. Мидгли, М. Аллен. Полевой справочник пресноводных рыб Австралии. Ред. Ян Найт / Венди Булгин. Перт, Вашингтон: Музей Западной Австралии, 2002. С. 54–55.
- ^ Амемия, К. Т.; Альфельди, Дж; Ли, А. П.; Поклонники; Филипп, H; MacCallum, I; и другие. (2013). «Геном африканских латимерии дает представление об эволюции четвероногих». Природа. 496 (7445): 311–316. Bibcode:2013Натура.496..311A. Дои:10.1038 / природа12027. ЧВК 3633110. PMID 23598338.
- ^ Клак, Дженнифер А. (2012). Укрепление позиций: происхождение и эволюция четвероногих. Издательство Индианского университета. ISBN 9780253356758.
- ^ Нельсон 2006.
- ^ Йохансон Зерина; Лонг Джон А .; Талант Джона А .; Жанвье Филипп; Уоррен Джеймс У. (2006). «Древнейшая целакант из раннего девона Австралии». Письма о биологии. 2 (3): 443–46. Дои:10.1098 / rsbl.2006.0470. ЧВК 1686207. PMID 17148426. Архивировано из оригинал 19 февраля 2013 г.
- ^ Форей 1998.
- ^ а б Введение в актиноптеригию В архиве 2013-02-17 в Wayback Machine Музей палеонтологии, Калифорнийский университет.
- ^ Докинз 2004, п. 289: «Очевидно, у позвоночных животных должны были быть предки, жившие в кембрии, но предполагалось, что они были беспозвоночными предшественниками настоящих позвоночных - протохордовых. Пикайя широко рекламировалась как старейшее ископаемое протохордовое животное».
- ^ Моррис SC (1979). «Фауна сланцев Берджесса (средний кембрий)». Ежегодный обзор экологии и систематики. 10: 327–349. Дои:10.1146 / annurev.es.10.110179.001551. JSTOR 2096795.
- ^ Моррис СК, Кэрон Дж. Б. (2012). "Pikaia gracilens Walcott, хордовые стволовые группы из среднего кембрия Британской Колумбии " (PDF). Биологические обзоры. 87 (2): 480–512. Дои:10.1111 / j.1469-185X.2012.00220.x. PMID 22385518. S2CID 27671780. Архивировано из оригинал (PDF) на 2012-05-27. Получено 2013-01-19.
- ^ Палмер 2000, п. 66-67.
- ^ Кембрий: 2 В архиве 2013-02-17 в Wayback Machine Палеос. Изменено 26 октября 2002 г. Проверено 20 января 2013 г.
- ^ Donoghue, P.C.J .; Пурнелл, М.А. (2005). «Дублирование генома, вымирание и эволюция позвоночных» (PDF). Тенденции в экологии и эволюции. 20 (6): 312–319. Дои:10.1016 / j.tree.2005.04.008. PMID 16701387. Архивировано из оригинал (PDF) на 2008-12-17. Получено 2013-01-08.
- ^ а б c d е ж Палмер, Д., изд. (1999). Иллюстрированная энциклопедия динозавров и доисторических животных Маршалла. Лондон: Marshall Editions. п. 23. ISBN 978-1-84028-152-1.
- ^ Сансом И.Дж., Смит М.П., Смит М.М., Тернер П. (1997). "Астраспис: Анатомия и гистология ордовикских рыб » (PDF). Палеонтология. 40 (3): 625–642. Архивировано из оригинал (PDF) на 2013-10-29.
- ^ Тернер, С .; Тарлинг, Д. Х. (1982). «Распространение телодонта и других бесчеловечных галактик как тесты континентальных реконструкций нижнего палеозоя». Палеогеография, палеоклиматология, палеоэкология. 39 (3–4): 295–311. Bibcode:1982ППП .... 39..295Т. Дои:10.1016 / 0031-0182 (82) 90027-X.
- ^ Сарджант и Холстед.
- ^ Донохью 2000, п. 206.
- ^ Тернер1999, п. 42–78.
- ^ Ранний и средний силурийский период. Видеть Казлев, М.А .; Белый, Т (6 марта 2001 г.). "Палеос: Телодонти". Palaeos.com. Архивировано из оригинал на 2007-10-28. Получено 30 октября, 2007.
- ^ а б c Баэз. Джон (2006) Вымирание В архиве 2013-01-30 в Wayback Machine Калифорнийский университет. Проверено 20 января 2013 года.
- ^ Альберг 2001, п. 188.
- ^ Паттерсон 1987, п. 142.
- ^ Холл и Ханкен 1993, п. 131.
- ^ Жанвье 2003.
- ^ Сансом, Р. С. (2009). «Филогения, классификация и полярность характера остеостраков (позвоночных)». Журнал систематической палеонтологии. 7: 95–115. Дои:10.1017 / S1477201908002551. S2CID 85924210.
- ^ Отто, М .; Лаурин, М. (2001). «Микроанатомия дермального скелета Balticaspis latvica (Остеостраки, средний девон) ". Журнал палеонтологии позвоночных. 21 (1): 186–189. Дои:10.1671 / 0272-4634 (2001) 021 [0186: motdso] 2.0.co; 2. В архиве из оригинала 2020-09-03. Получено 2014-05-04.
- ^ Чу, Брайан; Чжу, Мин; Чжао, Вэньцзинь; Цзя, Ляотао; Чжу, Юань (2014). «Крупнейшее силурийское позвоночное животное и его палеоэкологические последствия». Научные отчеты. 4: 5242. Bibcode:2014НатСР ... 4E5242C. Дои:10.1038 / srep05242. ЧВК 4054400. PMID 24921626.
- ^ Чжу М., Чжао В., Цзя Л., Лу Дж., Цяо Т., Цюй Q (2009). «Самый древний сочлененный остейхтиан обнаруживает мозаичные гнатостомы». Природа. 458 (7237): 469–474. Bibcode:2009Натура.458..469Z. Дои:10.1038 / природа07855. PMID 19325627. S2CID 669711.
- ^ Коутс М.И. (2009). «Палеонтология: за пределами эпохи рыб». Природа. 458 (7237): 413–414. Bibcode:2009 Натур.458..413C. Дои:10.1038 / 458413a. PMID 19325614. S2CID 4384525.
- ^ Фарингула В архиве 2012-03-09 в Wayback MachineНаучные блоги, 1 апреля 2009 г.
- ^ Мин. Z; Шульце, Ганс-Петер (1997). «Самая старая саркоптеригийская рыба» (PDF). Lethaia. 30 (4): 293–304. Дои:10.1111 / j.1502-3931.1997.tb00472.x. В архиве из оригинала 2020-09-03. Получено 2017-12-12.
- ^ Мерсс Т. (2001). "Андреолепис (Actinopterygii) в верхнем силуре северной Евразии ». Известия Эстонской академии наук. 50 (3): 174–189. В архиве из оригинала на 2020-08-01. Получено 2020-06-07.
- ^ Parry, S.F .; Благородный С.Р .; Crowley Q.G .; Wellman C.H. (2011). «Ограничение возраста U – Pb с высокой точностью на Rhynie Chert Konservat-Lagerstätte: шкала времени и другие последствия». Журнал геологического общества. Лондон: Геологическое общество. 168 (4): 863–872. Дои:10.1144/0016-76492010-043.
- ^ Kaufmann, B .; Trapp, E .; Мезгер, К. (2004). «Численный возраст горизонтов Келлвассера верхнего франа (верхнего девона): новая датировка циркона U-Pb из Штайнбруха Шмидта (Келлервальд, Германия)». Журнал геологии. 112 (4): 495–501. Bibcode:2004JG .... 112..495K. Дои:10.1086/421077.
- ^ Амос, Джонатан (6 января 2010 г.). "Ископаемые следы" старейших путешественников по земле'". Новости BBC. В архиве с оригинала 24 августа 2018 г.. Получено 4 декабря 2012.
- ^ «Музей Века Рыб - Дом». www.ageoffishes.org.au. В архиве из оригинала 14 марта 2019 г.. Получено 25 марта 2019.
- ^ Бентон 2005, п. 65.
- ^ Иллюстрированная энциклопедия динозавров и доисторических животных Маршалла, 1999 г., п. 43.
- ^ Холланд Т (2010). «Остеихтиановые останки верхнего девона из реки Генуя, Виктория, Австралия» (PDF). Воспоминания музея Виктории. 67: 35–44. Дои:10.24199 / j.mmv.2010.67.04.[постоянная мертвая ссылка ]
- ^ Делл'Амор, К. (12 сентября 2011 г.). «Древняя зубастая рыба, найденная в Арктике - гигантские реки». Национальная география Ежедневные новости. В архиве из оригинала 24 сентября 2011 г.. Получено 13 сентября, 2011.
- ^ Том Аврил (12 сентября 2011 г.). «Ископаемые остатки рыб проливают свет на фазу« Еврамерика »». Спрашивающий. В архиве из оригинала 15 сентября 2012 г.. Получено 15 сентября, 2011.
- ^ Воробьева, Е. (2006). "Новый вид Laccognathus (Porolepiform Crossopterygii) из девона Латвии ». Палеонт. Дж. Physorg.com. 40 (3): 312–322. Дои:10.1134 / S0031030106030129. S2CID 129696592.
- ^ Вицман, Ф. (2011). "Новый вид Laccognathus (Sarcopterygii, Porolepiformes) из позднего девона острова Элсмир, Нунавут, Канада ". Журнал палеонтологии позвоночных. 31 (5): 981–996. Дои:10.1080/02724634.2011.599462. S2CID 140604758.
- ^ Иллюстрированная энциклопедия динозавров и доисторических животных Маршалла, 1999 г., п. 45.
- ^ а б c d Иллюстрированная энциклопедия динозавров и доисторических животных Маршалла, 1999 г., п. 26.
- ^ Иллюстрированная энциклопедия динозавров и доисторических животных Маршалла, 1999 г., п. 32.
- ^ Длинный 1996.
- ^ Древняя рыба с укусом убийцы В архиве 2012-09-29 в Wayback Machine. Новости науки. 19 мая 2009 года.
- ^ Иллюстрированная энциклопедия динозавров и доисторических животных Маршалла, 1999 г., п. 33.
- ^ Рыба-монстр сокрушила противника самым сильным укусом В архиве 2013-04-02 в Wayback Machine. Сидней Морнинг Геральд. 30 ноября 2006 г.
- ^ «Пресс-релиз: Древний хищник обладал сильнейшей укусой из всех рыб, соперничая с укусами крупных аллигаторов и T. rex». Полевой музей через EurekAlert !. 28 ноября 2006 г. В архиве с оригинала 10 августа 2017 г.. Получено 9 августа, 2017.
- ^ Рой Бритт, Роберт (28 ноября 2006 г.). "У доисторических рыб были самые мощные челюсти". LiveScience. В архиве из оригинала 2020-09-03. Получено 2009-04-26.
- ^ Long, J. A .; Trinajstic, K .; Young, G.C .; Зенден, Т. (2008). «Живорождение в девонский период». Природа. 453 (7195): 650–652. Bibcode:2008Натура.453..650л. Дои:10.1038 / природа06966. PMID 18509443. S2CID 205213348.
- ^ Ископаемое обнаруживает древнейшее живорождение В архиве 2009-02-28 в Wayback Machine Новости BBC, 28 мая 2008 г.
- ^ Томсон, К. С. (1968). «Новая девонская рыба (Crossopterygii: Rhipidistia), рассматриваемая в связи с происхождением Амфибии». Postilla. 124.
- ^ а б Гжегож Недзведски; Петр Шрек; Катажина Наркевич; Марек Наркевич; Пер Э. Альберг (2010). «Пути тетрапод раннего среднего девона в Польше». Природа. Издательская группа "Природа". 463 (7277): 43–48. Bibcode:2010Натура.463 ... 43N. Дои:10.1038 / природа08623. PMID 20054388. S2CID 4428903.
- ^ Гордон, М. С .; Graham, J. B .; Ван, Т. (2004). «Возвращаясь к вторжению позвоночных на землю». Физиологическая и биохимическая зоология. 77 (5): 697–699. Дои:10.1086/425182. S2CID 83750933.
- ^ а б c d Клак, Дж. А. (Ноябрь 2005 г.). "Попасть на землю". Scientific American. Архивировано из оригинал on 2020-07-28. Получено 2008-09-06.
- ^ а б Лаурин М .; Meunier F.J .; Germain D .; Лемуан М. (2007). "Микроанатомическое и гистологическое исследование парного скелета плавников девонского саркоптерига Eusthenopteron Foordi". Журнал палеонтологии. 81: 143–153. Дои:10.1666 / 0022-3360 (2007) 81 [143: AMAHSO] 2.0.CO; 2. ISSN 0022-3360.
- ^ «В 2011 году у перехода рыба-четвероногая появилась новая гипотеза». Наука 2.0. 27 декабря 2011 г. В архиве с оригинала 14 января 2012 г.. Получено 2 января, 2012.
- ^ Shanta Barley (6 января 2010 г.). «Обнаружены древнейшие следы четвероногого позвоночного». Новый ученый. В архиве из оригинала 17 марта 2010 г.. Получено 3 января, 2010.
- ^ Реталлак, Грегори (2011). "Гипотеза Вудленда для эволюции девонских тетрапод". Журнал геологии. Издательство Чикагского университета. 119 (3): 235–258. Bibcode:2011JG .... 119..235R. Дои:10.1086/659144. S2CID 128827936.
- ^ Холмгрен, Н. (1933). «О происхождении конечности четвероногих». Acta Zoologica. 14 (2–3): 185–295. Дои:10.1111 / j.1463-6395.1933.tb00009.x.
- ^ Воробьева Е.И. (1992). «Роль развития и функции в формировании четвероногих грудных плавников». Ж. Общ. Биол. 53: 149–158.
- ^ Уотсон DMS (1913). "На конечности примитивного четвероногого". Анат. Anzeiger. 44: 24–27.
- ^ Шубин Н., Табин С., Кэрролл С. (1997). «Ископаемые, гены и эволюция конечностей животных». Природа. 388 (6643): 639–48. Bibcode:1997Натура.388..639S. Дои:10.1038/41710. PMID 9262397. S2CID 2913898.
- ^ а б c d Boisvert CA, Марк-Курик Э., Альберг PE (2008). «Грудной плавник Пандерихтиса и происхождение цифр». Природа. 456 (7222): 636–8. Bibcode:2008Натура.456..636Б. Дои:10.1038 / природа07339. PMID 18806778. S2CID 2588617. В архиве из оригинала от 04.01.2014. Получено 2013-02-15.
- ^ Тан, Кер (24 сентября 2008 г.). «У древних рыб были примитивные пальцы рук и ног». National Geographic News. В архиве с оригинала от 27 сентября 2008 г.
- ^ Ahlberg, P.E .; Мильнер, А. Р. (апрель 1994 г.). «Происхождение и ранняя диверсификация четвероногих». Природа. 368 (6471): 507–514. Bibcode:1994Натура.368..507А. Дои:10.1038 / 368507a0. S2CID 4369342.
- ^ Геологическая служба Канады (2008-02-07). «Прошлые жизни: Хроники канадской палеонтологии: Евстеноптерон - принц Мигуаша». Архивировано из оригинал на 2004-12-11. Получено 2009-02-10.
- ^ «Архивная копия». Архивировано из оригинал на 2012-04-20. Получено 2013-01-24.CS1 maint: заархивированная копия как заголовок (связь)
- ^ Природа: Тазовый плавник и пояс Пандерихтис и происхождение передвижения четвероногих В архиве 2012-08-12 в Wayback Machine
- ^ Эдвард Б. Даешлер; Нил Х. Шубин; Фариш А. Дженкинс-младший (6 апреля 2006 г.). «Девонская рыба, похожая на четвероногих, и эволюция строения тела четвероногих». Природа. 440 (7085): 757–763. Bibcode:2006Натура.440..757D. Дои:10.1038 / природа04639. PMID 16598249.
- ^ Джон Нобл Уилфорд, Нью-Йорк Таймс, Ученые называют ископаемые рыбки недостающим звеном В архиве 2017-11-15 в Wayback Machine, 5 апреля 2006 г.
- ^ Шубин, Нил (2008). Ваша внутренняя рыба. Пантеон. ISBN 978-0-375-42447-2.
- ^ "Acanthostega gunneri В архиве 2004-12-24 на Wayback Machine," Девонские времена.
- ^ Даешлер, Э. Б., Шубин, Н. Х. и Дженкинс, Ф. А. (апрель 2006 г.). «Девонская рыба, похожая на четвероногих, и эволюция строения тела четвероногих» (PDF). Природа. 440 (7085): 757–763. Bibcode:2006Натура.440..757D. Дои:10.1038 / природа04639. PMID 16598249. S2CID 4413217. Архивировано из оригинал (PDF) на 2012-03-01.CS1 maint: несколько имен: список авторов (связь)
- ^ а б Кларк, Джон Т .; Фридман, Мэтт (август 2018). «Разнообразие форм тела у рыб-неоптерисов триаса-раннего мела: устойчивое неравенство голостовых и преимущественно постепенное увеличение фенотипического разнообразия костистых рыб». Палеобиология. 44 (3): 402–433. Дои:10.1017 / pab.2018.8. ISSN 0094-8373. S2CID 90207334. В архиве из оригинала 2020-09-03. Получено 2019-12-12.
- ^ а б c d Р. Эйдан Мартин. «Золотой век акул». Биология акул и скатов. В архиве из оригинала от 22 мая 2008 г.. Получено 2008-06-23.
- ^ Ward P .; Labandeira, C .; Лаурин, М .; Berner, R.A .; и другие. (2006). «Подтверждением разрыва Ромера является низкий кислородный интервал, ограничивающий время первоначальной наземной трансформации членистоногих и позвоночных». Труды Национальной академии наук. 103 (45): 16818–16822. Bibcode:2006PNAS..10316818W. Дои:10.1073 / pnas.0607824103. ЧВК 1636538. PMID 17065318.
- ^ Лунд, Ричард; Гроган, Эйлин (2006). «Ископаемые рыбы Медвежьего ущелья: Falcatus falcatus". Архивировано из оригинал на 21.08.2008.
- ^ Эдинбург, Королевское физическое общество (1880 г.). "Труды Королевского физического общества Эдинбурга". V: 115. В архиве из оригинала на 2020-08-01. Получено 2020-06-07. Цитировать журнал требует
| журнал =(помощь) - ^ Николсон, Генри Аллейн; Ричард Лидеккер (1889 г.). Руководство по палеонтологии. п. 966. В архиве из оригинала на 2020-08-01. Получено 2020-06-07.
- ^ "Исследование проливает новый свет на общего предка всех челюстных позвоночных". Sci-News.com. 15 июня 2012 г. В архиве из оригинала 21 октября 2012 г.
- ^ Коутс, Майкл I .; Finarelli, John A .; Дэвис, Сэмюэл П. (1 июня 2012 г.). «Акантоды и акулоподобные состояния у последнего общего предка современных гнатостомов». Природа. 486 (7402): 247–250. Bibcode:2012Натура.486..247D. Дои:10.1038 / природа11080. PMID 22699617. S2CID 4304310.
- ^ Олден, Эндрю (17 марта 2017 г.). «Пермско-триасовое вымирание». ThoughtCo. В архиве с оригинала 25 марта 2019 г.
- ^ «Архивная копия». Архивировано из оригинал на 2012-09-20. Получено 2012-12-04.CS1 maint: заархивированная копия как заголовок (связь)
- ^ Sahney, S .; Бентон, М.Дж. (2008). «Восстановление после самого глубокого массового вымирания всех времен». Труды Королевского общества B: биологические науки. 275 (1636): 759–65. Дои:10.1098 / rspb.2007.1370. ЧВК 2596898. PMID 18198148.
- ^ Костные рыбы В архиве 2013-06-06 на Wayback Machine Морской мир. Проверено 2 февраля 2013 года.
- ^ а б Иллюстрированная энциклопедия динозавров и доисторических животных Маршалла, 1999 г., п. 38–39.
- ^ а б «вымирание». Math.ucr.edu. Архивировано из оригинал на 2012-08-11. Получено 2008-11-09.
- ^ Мотани, Р.(2000), Rulers of the Jurassic Seas, Scientific American, том 283, вып. 6
- ^ Крылья, Оливер; Раби, Мартон; Schneider, Jörg W .; Шверманн, Леони; солнце, Ge; Чжоу, Чан-Фу; Джойс, Уолтер Г. (2012). «Огромное ложе юрской черепахи из бассейна Турфан в Синьцзяне, Китай». Naturwissenschaften. 114 (11): 925–935. Bibcode:2012NW ..... 99..925W. Дои:10.1007 / s00114-012-0974-5. PMID 23086389. S2CID 17423081.
- ^ Гэннон, Меган (31 октября 2012 г.). «В Китае найдено кладбище юрских черепах». Livescience.com. В архиве из оригинала 5 ноября 2012 г.. Получено 4 декабря, 2012.
- ^ Листон, Джефф, М. Ньюбри, Т. Чалландс и К. Адамс (2013) "Рост, возраст и размеры юрских пахикормид. Leedsichthys problematicus (Osteichthyes: Actinopterygii) В архиве 2016-03-04 в Wayback Machine. "в Дж. Арратиа, Х. П. Шульце и М. В. Х. Уилсон (ред.) Мезозойские рыбы 5 - Глобальное разнообразие и эволюция. Страницы 145-175, F. Pfeil. ISBN 9783899370539.
- ^ "Самая большая рыба из когда-либо найденных" обнаружена в Великобритании. В архиве 2016-08-12 в Wayback Machine National Geographic News, 1 октября 2003 г.
- ^ Б.Г. Гардинер (1984) Осетровые как живые ископаемые. Стр. 148–152 в N. Eldredge and S.M. Стэнли, ред. Живые окаменелости. Спрингер-Верлаг, Нью-Йорк.
- ^ Krieger J .; Фюрст П.А. (2002). «Доказательства замедленной скорости молекулярной эволюции в отряде Acipenseriformes». Молекулярная биология и эволюция. 19 (6): 891–897. Дои:10.1093 / oxfordjournals.molbev.a004146. PMID 12032245.
- ^ Рафферти, Джон П. (2010) Мезозойская эра: эпоха динозавров В архиве 2020-08-01 в Wayback Machine Стр. 219, Издательская группа Розен. ISBN 9781615301935.
- ^ Ископаемые (Смитсоновские справочники) Дэвид Уорд (Стр.200)
- ^ База данных палеобиолой Ptychodus entry В архиве 2013-07-26 в Wayback Machine доступ 23.08.09
- ^ "BBC - Earth News - В Канзасе обнаружено окаменелость гигантской хищной акулы"
- ^ Маклауд, Н., Роусон, П.Ф., Фори, П.Л., Баннер, Ф.Т., Будагер-Фадель, М.К., Баун, П.Р., Бернетт, Дж. А., Чемберс, П., Калвер, С., Эванс, С. Е., Джеффри, К., Камински, Массачусетс, Лорд, А.Р., Милнер, А.С., Милнер, А.Р., Моррис, Н., Оуэн, Э., Розен, Б.Р., Смит, А.Б., Тейлор, П.Д., Уркхарт, И. и Янг, мл. «Меловой – третичный биотический переход». Журнал геологического общества. 154 (2): 265–292. Bibcode:1997JGSoc.154..265M. Дои:10.1144 / gsjgs.154.2.0265. S2CID 129654916.CS1 maint: несколько имен: список авторов (связь)
- ^ Паттерсон, С. (1993). Osteichthyes: Teleostei. В: The Fossil Record 2 (Бентон, MJ, редактор). Springer. стр.621–656. ISBN 978-0-412-39380-8.
- ^ Zinsmeister WJ (1 мая 1998 г.). «Открытие горизонта смертности рыб на границе К – Т на острове Сеймур: переоценка событий в конце мелового периода». Журнал палеонтологии. 72 (3): 556–571. Дои:10.1017 / S0022336000024331. В архиве из оригинала 30.10.2007. Получено 2007-08-27.
- ^ Робертсон Д.С., Маккенна М.С., Мультяшный OB, Надежда S, Лиллегрейвен Дж. А. (2004). «Выживание в первые часы кайнозоя» (PDF). Бюллетень GSA. 116 (5–6): 760–768. Bibcode:2004GSAB..116..760R. Дои:10.1130 / B25402.1. Архивировано из оригинал (PDF) на 2015-10-22. Получено 2016-01-06.
- ^ Sibert, E.C .; Норрис, Р. Д. (2015-06-29). «Новая эра рыб, инициированная массовым вымиранием мелового и палеогенового периода». PNAS. 112 (28): 8537–8542. Bibcode:2015ПНАС..112.8537С. Дои:10.1073 / pnas.1504985112. ЧВК 4507219. PMID 26124114.
- ^ «Находка странной рыбы противоречит аргументу разумного замысла». Национальная география. 9 июля 2008 г. В архиве из оригинала на 2018-05-28. Получено 2008-07-17.
- ^ Мэтт Фридман (10 июля 2008 г.). «Эволюционное происхождение асимметрии камбалы». Природа. 454 (7201): 209–212. Bibcode:2008Натура.454..209F. Дои:10.1038 / природа07108. PMID 18615083. S2CID 4311712.
- ^ а б Wroe, S .; Huber, D. R .; Lowry, M .; McHenry, C .; Морено, К .; Clausen, P .; Ferrara, T. L .; Cunningham, E .; Dean, M. N .; Саммерс, А. П. (2008). «Трехмерный компьютерный анализ механики челюсти белой акулы: насколько сильно может укусить большая белая акула?» (PDF). Журнал зоологии. 276 (4): 336–342. Дои:10.1111 / j.1469-7998.2008.00494.x. В архиве (PDF) из оригинала на 2018-11-27. Получено 2013-01-12.
- ^ Пимиенто, Каталина; Дана Дж. Эрет; Брюс Дж. Макфадден; Гордон Хаббелл (10 мая 2010 г.). Степанова, Анна (ред.). «Древний питомник вымершей гигантской акулы мегалодона из миоцена Панамы». PLOS ONE. Панама: PLoS.org. 5 (5): e10552. Bibcode:2010PLoSO ... 510552P. Дои:10.1371 / journal.pone.0010552. ЧВК 2866656. PMID 20479893.
- ^ Ламбер, Оливье; Джованни Биануччи; Клаас Пост; Кристиан де Мюисон; Родольфо Салас-Гисмонди; Марио Урбина; Джелле Реумер (1 июля 2010 г.). «Гигантский укус нового кашалота-хищника из миоценовой эпохи Перу». Природа. Перу. 466 (7302): 105–108. Bibcode:2010Натура.466..105л. Дои:10.1038 / природа09067. PMID 20596020. S2CID 4369352.
- ^ Myxini В архиве 2017-12-15 в Wayback Machine - Музей палеонтологии Калифорнийского университета
- ^ Смит, Дж. Л. Б. (1956). Старые Четвероногие: История Целаканта. Лонгманс Грин.
- ^ Lavett Smith, C .; Rand, Charles S .; Шеффер, Бобб; Ац, Джеймс В. (1975). «Латимерия, живая латимерия, яйцеживородящая». Наука. 190 (4219): 1105–6. Bibcode:1975Научный ... 190.1105Л. Дои:10.1126 / science.190.4219.1105. S2CID 83943031.
Библиография
- Альберг, Пер Эрик (2001). Основные события в ранней эволюции позвоночных: палеонтология, филогения, генетика и развитие. Вашингтон, округ Колумбия: Тейлор и Фрэнсис. ISBN 978-0-415-23370-5. В архиве из оригинала от 16.06.2016.
- Бентон, Майкл Дж. (2005): Палеонтология позвоночных
- Бентон, Майкл (2009) Палеонтология позвоночных В архиве 2014-07-05 в Wayback Machine Выпуск 3, John Wiley & Sons. ISBN 9781405144490.
- Берг, Линда Р .; Эльдра Перл Соломон; Дайана В. Мартин (2004). Биология. Cengage Learning. ISBN 978-0-534-49276-2.
- Хейнс, Тим; Чемберс, Пол (2005). «Полное руководство по доисторической жизни». Книги Светлячка. Цитировать журнал требует
| журнал =(помощь) - Клаудсли-Томпсон, Дж. Л. (2005). Экология и поведение мезозойских рептилий. 9783540224211: Springer.CS1 maint: location (связь)
- Докинз, Ричард (2004). Сказка предков: паломничество к заре жизни. Бостон: Компания Houghton Mifflin. ISBN 978-0-618-00583-3.
- Донохью, П. К., П. Л. Фори и Р. Дж. Олдридж (2000). «Конодонтовое родство и хордовая филогения». Биологические обзоры Кембриджского философского общества. 75 (2): 191–251. Дои:10.1111 / j.1469-185X.1999.tb00045.x. PMID 10881388. S2CID 22803015.CS1 maint: несколько имен: список авторов (связь)
- Британская энциклопедия (1954). Британская энциклопедия: новый обзор универсальных знаний. 17.
- Фори, Питер Л. (1998) История латимерийских рыб. Лондон: Чепмен и Холл.
- Холл, Брайан Кейт; Ханкен, Джеймс (1993). Череп. Чикаго: Издательство Чикагского университета. ISBN 978-0-226-31568-3. В архиве из оригинала на 20.05.2016. Получено 2016-01-27.
- Хелфман Джи, Коллетт БиБи, Фэйси Д.Х. и Боуэн Б.В. (2009) Разнообразие рыб: биология, эволюция и экология В архиве 2014-11-14 на Wayback Machine Вили-Блэквелл. ISBN 978-1-4051-2494-2
- Жанвье, Филипп (2003). Ранние позвоночные. Издательство Оксфордского университета. ISBN 978-0-19-852646-9.
- Lecointre, G; Ле Гуядер, H (2007). «Древо жизни: филогенетическая классификация». Справочная библиотека издательства Гарвардского университета. Цитировать журнал требует
| журнал =(помощь) - Лонг, Джон А. Восстание рыб: 500 миллионов лет эволюции. Балтимор: Издательство Университета Джона Хопкинса, 1996. ISBN 0-8018-5438-5
- Нельсон, Джозеф С. (2006). Рыбы мира. Джон Уайли и сыновья, Inc. ISBN 978-0-471-25031-9.
- Палмер, Д., изд. (1999). Иллюстрированная энциклопедия динозавров и доисторических животных Маршалла. Лондон: Marshall Editions. п. 26. ISBN 978-1-84028-152-1.
- Палмер, Д. (2000). «Атлас доисторического мира». Филадельфия: Marshall Publishing Ltd. Цитировать журнал требует
| журнал =(помощь) - Паттерсон, Колин (1987). Молекулы и морфология в эволюции: конфликт или компромисс?. Кембридж, Великобритания: Издательство Кембриджского университета. ISBN 978-0-521-32271-3. В архиве из оригинала от 24.06.2016. Получено 2016-01-27.
- Sarjeant, William Antony S .; Л. Б. Холстед (1995). Окаменелости позвоночных и эволюция научных концепций: труды, посвященные Беверли Холстед. ISBN 978-2-88124-996-9. В архиве из оригинала на 16.02.2017. Получено 2020-06-07.
- Ромер, А.С. (1970). «Тело позвоночных» (4-е изд.). Лондон: W.B. Сондерс. Цитировать журнал требует
| журнал =(помощь) - Тернер, С. (1999). «Раннесилурийско-раннедевонские комплексы телодонтов и их возможное экологическое значение». В А. Ж. Буко; Дж. Лоусон (ред.). Палеосообщества, Международная программа геологической корреляции 53, Проект Экостратиграфия, Заключительный отчет. Издательство Кембриджского университета.
дальнейшее чтение
| Внешнее видео | |
|---|---|
- Бентон MJ (1998) «Качество летописи окаменелостей позвоночных» Страницы 269–303 в Donovan, SK и Paul CRC (ред.), Адекватность летописи окаменелостей. Вайли. ISBN 9780471969884.
- Клотье Р. (2010). «Летопись окаменелостей онтогенеза рыб: понимание закономерностей и процессов развития» (PDF). Семинары по клеточной биологии и биологии развития. 21 (4): 400–413. Дои:10.1016 / j.semcdb.2009.11.004. PMID 19914384.[постоянная мертвая ссылка ]
- Жанвье, Филипп (1998) Ранние позвоночные, Оксфорд, Нью-Йорк: Издательство Оксфордского университета. ISBN 0-19-854047-7
- Лонг, Джон А. (1996) Восстание рыб: 500 миллионов лет эволюции Издательство Университета Джона Хопкинса. ISBN 0-8018-5438-5
- McKenzie DJ, Фаррелл AP и Браунер CJ (2011) Физиология рыб: примитивные рыбы Академическая пресса. ISBN 9780080549521.
- Мэйси Дж. Г. (1996) Обнаружение ископаемых рыб Холт. ISBN 9780805043662.
- Рядом, T.J .; Дорнбург, А .; Eytan, R.I .; Keck, B.P .; Smith, W.L .; Kuhn, K.L .; Moore, J.A .; Цена, S.A .; Burbrink, F.T .; Фридман, М. (2013). «Филогения и темп диверсификации в сверхизлучении колючих рыб». Труды Национальной академии наук. 110 (31): 12738–12743. Bibcode:2013ПНАС..11012738Н. Дои:10.1073 / pnas.1304661110. ЧВК 3732986. PMID 23858462.
- Шубин, Нил (2009) Ваша внутренняя рыба: путешествие в 3,5-миллиардную историю человеческого тела Винтажные книги. ISBN 9780307277459.
- Знакомство с позвоночными Музей палеонтологии, Калифорнийский университет.
внешняя ссылка
- Ископаемая рыба
- Происхождение рыбы
- Обзор эволюции – Карл Саган
- Происхождение позвоночных Марк В. Киршнер, iBioSeminars.
- 150 миллионов лет эволюции рыб в одной удобной фигурке НаучныйАмериканский, 29 августа 2013 г.
- Музей возраста рыб, Canowindra - постоянная выставка одних из лучших из тысяч окаменелостей девонского периода, найденных поблизости.
| О рыбы |  | ||||
|---|---|---|---|---|---|
| Анатомия и физиология | |||||
| Сенсорный системы | |||||
| Размножение | |||||
| Передвижение | |||||
| Другой поведение | |||||
| К среда обитания | |||||
| Другие типы | |||||
| Коммерческий |
| ||||
| Основные группы | |||||
| Списки | |||||
| |||||
| Предтечи |  | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Рыба без челюсти |
| ||||||||||||
| Челюстная рыба |
| ||||||||||||
| Списки | |||||||||||||
| Связанный | |||||||||||||
† вымерший | |||||||||||||

| Плавники |  | |
|---|---|---|
| Конечности | ||
| Крылья | ||
| Эволюция | ||
| Связанный | ||