Плезиозаврия - Plesiosauria
| Плезиозавры | |
|---|---|
 | |
| Восстановленный скелет Плезиозавр | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Класс: | Рептилии |
| Суперзаказ: | †Зауроптеригия |
| Clade: | †Пистозаврия |
| Порядок: | †Плезиозаврия Blainville, 1835 |
| Подгруппы | |
| |
В Плезиозаврия (/ˌпляsяəˈsɔːrяə,-zя-/;[1][2] Греческий: πλησίος, плезиос, что означает "рядом" и sauros, что означает "ящерица") или плезиозавры являются порядок или клады вымерших Мезозойский морские рептилии (морской Sauropsida ), принадлежащих Зауроптеригия.
Плезиозавры впервые появились в последнем Триасовый Период, возможно, в Ретиан стадии около 203 миллионов лет назад.[3] Они стали особенно распространены в период Юрский Период, процветающий до их исчезновения из-за Меловое – палеогеновое вымирание в конце Меловой Период, около 65 миллионов лет назад. У них было всемирное океаническое распространение.
Плезиозавры были одними из первых обнаруженных ископаемых рептилий. В начале девятнадцатого века ученые осознали, насколько отличительным было их телосложение, и в 1835 году их назвали отдельным отрядом. Первый род плезиозавров, одноименный Плезиозавр, был назван в 1821 году. С тех пор было описано более сотни достоверных видов. В начале двадцать первого века количество открытий увеличилось, что привело к лучшему пониманию их анатомии, взаимоотношений и образа жизни.
Плезиозавры имели широкое плоское тело и короткий хвост. Их конечности превратились в четыре длинных ласта, приводимые в движение сильными мышцами, прикрепленными к широким костным пластинам, образованным плечевым поясом и тазом. Ласты летели по воде. Плезиозавры дышали воздухом и рожали живых детенышей; есть признаки того, что они были теплокровными.
Плезиозавры показали два основных морфологический типы. Некоторые виды с телосложением «плезиозавроморф» имели (иногда чрезвычайно) длинные шеи и маленькие головы; они были относительно медленными и ловили мелких морских животных. У других видов, некоторые из которых достигают длины до семнадцати метров, «плиозавроморф» имел короткую шею и большую голову; это были высшие хищники, быстрые охотники на крупную добычу. Эти два типа связаны с традиционным строгим разделением плезиозавров на два подотряда: длинношеие. Плезиозавроидеа и короткая шея Плиозаавроидная. Современные исследования, однако, показывают, что некоторые группы с «длинной шеей» могли иметь некоторых членов с короткой шеей или наоборот. Поэтому были введены чисто описательные термины «плезиозавроморф» и «плиозавроморф», которые не подразумевают прямой связи. «Плезиозавроидеа» и «плиозавроидеа» сегодня имеют более ограниченное значение. Термин «плезиозавр» правильно используется для обозначения Plesiosauria в целом, но неофициально он иногда означает только длинношеие формы, старые Plesiosauroida.
История открытия
Ранние находки
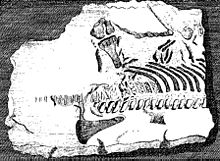
Элементы скелета плезиозавров - одни из первых окаменелостей вымерших рептилий, признанных таковыми.[4] В 1605 г. Ричард Верстеген из Антверпен проиллюстрировано в его Восстановление истлевшего интеллекта позвонки плезиозавра, которые он назвал рыбами и видел как доказательство того, что Великобритания когда-то был связан с европейским континентом.[5] Валлийский Эдвард Лхойд в его Lithophylacii Brittannici Ichnographia с 1699 г. также включены изображения позвонков плезиозавра, которые снова считались позвонками рыб или Ихтиоспондили.[6] Другие натуралисты в семнадцатом веке добавили останки плезиозавра в свои коллекции, такие как Джон Вудворд; только намного позже они были признаны плезиозаврическими и сегодня частично сохранились в Sedgwick Museum.[4]
В 1719 г. Уильям Стьукли описал частичный скелет плезиозавра, на который его внимание обратил прадед Чарльз Дарвин, Роберт Дарвин из Элстона. Каменная плита добыта в карьере на Фульбек в Линкольншир и использовался вместе с окаменелостью на нижней стороне, чтобы укрепить наклон водопоя в Элстон в Ноттингемшир. После того, как в нем были обнаружены странные кости, он был выставлен в местном доме священника, как останки грешника, утонувшего в монастыре. Великий потоп. Стукели подтвердил, что "делювиальный «природа, но понимала, что она представляет собой какое-то морское существо, возможно, крокодила или дельфина.[7] Экземпляр сегодня хранится в Музей естественной истории инвентарный номер БМНХ Р.1330. Это самый ранний обнаруженный более или менее полный скелет ископаемой рептилии в музейной коллекции. Возможно, это относится к Плезиозавр долиходеирус.[4]
В течение восемнадцатого века количество открытий английских плезиозавров быстро увеличивалось, хотя все они носили более или менее фрагментарный характер. Важными коллекционерами были преподобные Уильям Маунси и Баптист Ноэль Тернер, активный в Долина Бельвуар, коллекции которого в 1795 г. описаны Джон Николлс в первой части его История и древности графства Лестершир.[8] Один из частичных скелетов плезиозавра Тернера до сих пор сохраняется как образец BMNH R.45 в Британском музее естественной истории; это сегодня упоминается Талассиодракон.[4]
Название Плезиозавр

В начале девятнадцатого века плезиозавры были еще малоизвестны, и их особенное строение не было изучено. Никаких систематических различий между ихтиозавры, поэтому окаменелости одной группы иногда объединяли с окаменелостями другой, чтобы получить более полный образец. В 1821 году частичный скелет обнаружен в коллекции полковника Томас Джеймс Берч,[9] был описан Уильям Конибер и Генри Томас Де ла Беш, и признаны представляющими особую группу. Был назван новый род, Плезиозавр. Родовое название произошло от греческого πλήσιος, Plèsios, "ближе к" и латинизированный савр, в значении «ящерица», чтобы выразить Плезиозавр был в Цепь бытия более близко расположен к Sauria, особенно крокодил, чем Ихтиозавр, который имел форму более низшей рыбы.[10] Таким образом, название следует читать скорее как «приближающийся к саурии» или «почти рептилия», чем как «близкая ящерица».[11] Части образца все еще присутствуют в Музей естественной истории Оксфордского университета.[4]
Вскоре после этого морфология стал намного более известным. В 1823 году Томас Кларк сообщил о почти полном черепе, вероятно, принадлежащем Талассиодракон, который сейчас сохраняется Британская геологическая служба как образец BGS GSM 26035.[4] В том же году коммерческий коллекционер окаменелостей Мэри Эннинг и ее семья обнаружили почти полный скелет в Лайм Реджис в Дорсет, Англия, на том, что сегодня называется Юрское побережье. Он был приобретен Герцог Бекингемский, который предоставил его геологу Уильям Бакленд. Он, в свою очередь, позволил Конибиру описать это 24 февраля 1824 года в лекции для Геологическое общество Лондона,[12] во время того же собрания, на котором впервые было названо имя динозавра, Мегалозавр. Две находки показали уникальное и причудливое строение животных, которое в 1832 году профессор Бакленд сравнил с «морским змеем, пробегающим через черепаху». В 1824 году Конибер также предоставил конкретное имя к Плезиозавр: долиходеирус, что означает «длинный шею». В 1848 году скелет был куплен Британским музеем естественной истории и внесен в каталог как образец BMNH 22656.[4] Когда лекция была опубликована, Конибер также назвал второй вид: Плезиозавр гигантский. Это была форма с короткой шеей, позже присвоенная Плиозаавроидная.[13]

Широкой публике плезиозавры стали известны благодаря двум богато иллюстрированным публикациям коллекционера. Томас Хокинс: Воспоминания об ихтиосаури и плезиозавре 1834 г.[14] и Книга Великих морских драконов 1840 года. Хокинс придерживался весьма своеобразного взгляда на животных,[15] видя в них чудовищные творения дьявола, во время доадамитский этап истории.[16] В конце концов Хокинс продал свои ценные и красиво отреставрированные экземпляры Британскому музею естественной истории.[17]
В течение первой половины девятнадцатого века количество находок плезиозавров неуклонно росло, особенно благодаря открытиям в морских скалах Лайм-Реджиса. Сэр Ричард Оуэн в одиночку названо около сотни новых видов. Однако большинство их описаний было основано на отдельных костях без достаточного диагноза, чтобы можно было отличить их от других видов, которые были описаны ранее. Многие из новых видов, описанных в это время, впоследствии были признанный недействительным. Род Плезиозавр особенно проблематичен, так как большинство новых видов были помещены в него так, что он стал таксон для мусорной корзины. Постепенно были названы другие роды. Хокинс уже создал новые роды, хотя они больше не считаются действительными. В 1841 году Оуэн назвал Плиозавр брахидейрус. это этимология упоминалось ранее Плезиозавр долиходейрус так как это происходит от πλεῖος, плея, «более полно», отражая, что, по словам Оуэна, он был ближе к Саурии, чем Плезиозавр. Его видовое название означает «с короткой шеей».[18] Позже Pliosauridae были признаны имеющими морфологию, фундаментально отличную от плезиозаврид. Семья Плезиозавры уже был придуман Джон Эдвард Грей в 1825 г.[19] В 1835 г. Анри Мари Дюкроте де Бленвиль назвал отряд самим Plesiosauria.[20]
Американские открытия
Во второй половине девятнадцатого века важные находки были сделаны за пределами Англии. Хотя сюда входили и некоторые немецкие открытия, в основном это были плезиозавры, обнаруженные в отложениях американского мелового периода. Западный внутренний морской путь, то Мел Ниобрара. Одна окаменелость, в частности, знаменует начало Bone Wars между конкурирующими палеонтологами Эдвард Дринкер Коуп и Отниэль Чарльз Марш.
В 1867 году врач Теофил Тернер около Форт Уоллес в Канзас обнаружил скелет плезиозавра, который он подарил Коупу.[21] Коуп попытался реконструировать животное, предположив, что чем длиннее конечность позвоночника, тем короче шея. Вскоре он заметил, что скелет, обретающий форму под его руками, обладал некоторыми очень особыми качествами: шейные позвонки имели шевроны, а суставные поверхности хвостовых позвонков были ориентированы задом наперед.[22] Взволнованный, Коуп пришел к выводу, что открыл совершенно новую группу рептилий: Стрептозаврия или «обращенные заурианы», которые будут отличаться перевернутыми позвонками и отсутствием задних конечностей, причем основной движущей силой является хвост.[23] Опубликовав описание этого животного,[24] сопровождаемая иллюстрацией в учебнике о рептилиях и амфибиях,[25] Коп пригласил Марша и Джозеф Лейди восхищаться его новым Эласмозавр Platyurus. Некоторое время послушав интерпретацию Коупа, Марш предположил, что более простым объяснением странного строения было бы то, что Коуп перевернул позвоночный столб по отношению к телу в целом. Когда Коуп с негодованием отреагировал на это предложение, Лейди молча взял череп и приложил его к предполагаемому последнему хвостовому позвонку, которому он идеально подошел: на самом деле это был первый шейный позвонок, к которому все еще был прикреплен кусок заднего черепа.[26] Утомленный, Коуп попытался уничтожить все издание учебника и, когда это не удалось, немедленно опубликовал улучшенное издание с правильной иллюстрацией, но с идентичной датой публикации.[27] Он оправдал свою ошибку, заявив, что был введен в заблуждение самим Лейди, который, описывая образец Чимолиазавр, также перевернул позвоночник.[28] Позже Марш утверждал, что этот роман был причиной его соперничества с Коупом: «с тех пор он был моим заклятым врагом». И Коуп, и Марш в своем соперничестве назвали множество родов и видов плезиозавров, большинство из которых сегодня считаются недействительными.[29]
На рубеже веков большая часть исследований плезиозавров проводилась бывшим учеником Марша, профессором Сэмюэл Венделл Уиллистон. В 1914 году Уиллистон опубликовал свой Водные рептилии прошлого и настоящего.[30] Несмотря на лечение морских рептилий в целом, в течение многих лет он оставался самым обширным общим текстом о плезиозаврах.[31] В 2013 году первый современный учебник готовил Оливье Риппель. В середине двадцатого века США оставались важным центром исследований, главным образом благодаря открытиям Сэмюэл Пол Уэллс.
Недавние открытия
В то время как в течение девятнадцатого и большей части двадцатого века новые плезиозавры описывались в количестве трех или четырех родов каждое десятилетие, темп внезапно ускорился в 1990-х годах, когда в этот период было обнаружено семнадцать плезиозавров. Темпы открытий ускорились в начале двадцать первого века, и каждый год было названо около трех или четырех плезиозавров.[32] Это означает, что около половины известных плезиозавров относительно новы в науке, что является результатом гораздо более интенсивных полевых исследований. Некоторые из них происходят вдали от традиционных областей, например в новых сайтах, разработанных в Новая Зеландия, Аргентина, Чили,[33] Норвегия, Япония, Китай и Марокко, но места, где были сделаны наиболее оригинальные открытия, оказались по-прежнему продуктивными, и важные новые находки были найдены в Англии и Германии. Некоторые из новых родов представляют собой переименование уже известных видов, которые считались достаточно разными, чтобы гарантировать отдельное название рода.
В 2002 году прессе было объявлено о «Монстре Арамберри». Обнаружен в 1982 г. в селе Арамберри, в северном мексиканском штате Нуэво-Леон, изначально он был классифицирован как динозавр. Экземпляр на самом деле является очень большим плезиозавром, возможно, достигающим в длину 15 м (49 футов). В СМИ были опубликованы преувеличенные сообщения, в которых утверждалось, что он был длиной 25 метров (82 фута) и весил до 150 000 килограммов (330 000 фунтов), что сделало бы его одним из крупнейших хищников всех времен. Эта ошибка была резко увековечена в BBC с документальный серии Прогулка с динозаврами, который также преждевременно классифицировал его как Лиоплевродон ферокс.[34][35]
В 2004 году местный рыбак обнаружил совершенно неповрежденного молодняка плезиозавра. Бриджуотер-Бэй Национальный заповедник в Сомерсете, Великобритания. Окаменелость, датируемая 180 миллионов лет назад, на что указывает аммониты связанный с ним, измеряется 1,5 метра (4 фута 11 дюймов) в длину и может быть связан с Ромалеозавр. Вероятно, это наиболее хорошо сохранившийся из всех обнаруженных плезиозавров.[36][37][38]
В 2005 году останки трех плезиозавров (Dolichorhynchops herschelensis ) обнаружен в 1990-х гг. Гершель, Саскачеван были обнаружены как новый вид доктором Тамаки Сато, японским палеонтологом позвоночных.[39]
В 2006 году объединенная группа американских и аргентинских следователей (последний из Аргентинский антарктический институт и Музей Ла-Платы ) обнаружил скелет молодого плезиозавра длиной 1,5 метра (4 фута 11 дюймов) на Остров Вега в Антарктиде.[40] Окаменелость в настоящее время выставлена в геологическом музее Школа горного дела и технологий Южной Дакоты.[41]
В 2008 году ископаемые останки неописанного плезиозавра, которого назвали Хищник X, теперь известный как Pliosaurus funkei, были обнаружены в Свальбард.[нужна цитата ] Он имел длину 12 м (39 футов) и силу укуса 149 килоньютон (33000 фунтов).ж) является одним из самых мощных из известных.[42]
Увеличилось не только количество полевых открытий, но и с 1950-х годов плезиозавры стали предметом более обширной теоретической работы. Новый метод кладистика впервые позволил точно рассчитать их эволюционные отношения. Было опубликовано несколько гипотез о том, как они охотились и плавали, включая общие современные представления о биомеханика и экология. Многие недавние открытия проверили эти гипотезы и породили новые.
В декабре 2017 года на континенте Антарктида был найден большой скелет плезиозавра, самого старого существа на континенте и первого представителя его вида в Антарктиде.[43]
Эволюция

Плезиозаврия берет свое начало в Зауроптеригия, группа возможно архозавроморф рептилии, вернувшиеся в море. Продвинутая подгруппа завроптеригий, хищные Эусауроптеригия с маленькими головами и длинными шеями, разделенными на две ветви во время Верхний триас. Один из них, Nothosauroida сохранены в рабочем состоянии локтевые и коленные суставы; но другой, Пистозаврия, стал более полно адаптироваться к образу жизни морских обитателей. Их позвоночный столб стал жестче, и основная движущая сила во время плавания исходила уже не от хвоста, а от конечностей, которые превратились в ласты.[44] Пистозаврии стали теплокровными и живородящий, рожая жить молодым.[45] Рано, базальный, участники группы, традиционно называемой "Пистозавриды ", все еще были в основном прибрежными животными. Их плечевые пояса оставались слабыми, таз не могли выдержать силу сильного плавательного гребка, и их ласты были тупыми. Позже откололась более продвинутая группа пистозавров: плезиозавры. У них были усиленные плечевые пояса, более плоский таз и более заостренные ласты. Другие приспособления, позволяющие им колонизировать открытое море, включали жесткие суставы конечностей; увеличение количества фаланг кисти и стопы; более плотное латеральное соединение фаланги пальцев стопы и пальцев ног и укороченный хвост.[46][47]

С самого начала Юрский, то Геттангский стадия известна богатая радиация плезиозавров, подразумевая, что группа, должно быть, уже диверсифицировалась в Поздний триас; Однако из этого разнообразия было обнаружено лишь несколько очень базальных форм. Дальнейшая эволюция плезиозавров очень спорна. Различные кладистические анализы не привели к консенсусу относительно взаимоотношений между основными подгруппами плезиозавров. Традиционно плезиозавров делят на длинношеих. Плезиозавроидеа и короткошерстный Плиозаавроидная. Однако современные исследования показывают, что некоторые группы с длинной шеей могли иметь членов с короткой шеей. Чтобы избежать путаницы между филогения, эволюционные отношения и морфология Из-за того, как построено животное, формы с длинной шеей называются «плезиозавроморфами», а формы с короткой шеей - «плиозавроморфами», при этом виды «плезиозавроморфов» обязательно более тесно связаны друг с другом, чем с формами «плиозавроморф». .[48]

В последний общий предок Плезиозаврия, вероятно, была довольно маленькой формой с короткой шеей. В течение ранней юры подгруппой с наибольшим количеством видов была подгруппа Rhomaleosauridae, возможно, очень базальное отщепление видов, которые также были короткошеими. Плезиозавры в этот период были не более пяти метров (шестнадцати футов) в длину. Посредством Тоарский около 180 миллионов лет назад другие группы, в том числе Плезиозавры, стали более многочисленными, и у некоторых видов появилась более длинная шея, в результате чего общая длина тела достигала десяти метров (33 футов).[49]
В середине юры очень большой Pliosauridae эволюционировал. Они характеризовались большой головой и короткой шеей, например Лиоплевродон и Simolestes. У этих форм были черепа до трех метров (десять футов) в длину, до семнадцати метров (56 футов) и веса в десять тонн. У плиозавридов были большие конические зубы, и они были доминирующими морскими хищниками своего времени. В то же время, примерно 160 миллионов лет назад, Cryptoclididae Присутствовали более короткие виды с длинной шеей и маленькой головой.[50]
В Leptocleididae излучался во время Раннемеловой период. Это были довольно маленькие формы, которые, несмотря на свою короткую шею, могли быть более близки к Plesiosauridae, чем к Pliosauridae. Позднее в раннем меловом периоде Elasmosauridae появившийся; это были одни из самых длинных плезиозавров, достигавшие пятнадцати метров (пятьдесят футов) в длину из-за очень длинной шеи, содержащей до 76 позвонков, больше, чем у любого другого известного позвоночного животного. Pliosauridae все еще присутствовали, как показывают крупные хищники, такие как Кронозавр.[50]
В начале Поздний мел, то Ихтиозаврия вымерли; возможно, группа плезиозавров эволюционировала, чтобы заполнить свои ниши: Поликотилиды с короткой шеей и необычно удлиненной головой с узкой мордой. В позднем меловом периоде у эласмозавридов было еще много видов.[50]
Все плезиозавры стали вымерший в результате K-T событие в конце мелового периода, примерно 66 миллионов лет назад.[51]
Отношения
В современном филогения, клады определены группы, которые содержат все виды, принадлежащие определенной ветви эволюционного древа. Один из способов определить кладу - позволить ей состоять из последний общий предок двух таких видов и всех его потомков. Такая клада называется "клады узлов ". В 2008, Патрик Дракенмиллер и Энтони Рассел таким образом определили плезиозаврию как группу, состоящую из последнего общего предка Плезиозавр долихохейрус и Peloneustes филарх и все его потомки.[52] Плезиозавр и Peloneustes представляли основные подгруппы Plesiosauroida и Pliosauroida и были выбраны по историческим причинам; любой другой вид из этих групп был бы достаточен.
Другой способ определить кладу - позволить ей состоять из всех видов, более тесно связанных с определенным видом, который в любом случае желательно включить в кладу, чем с другим видом, который, напротив, желает исключить. Такая клада называется "стволовая клада ". Такое определение имеет то преимущество, что легче включить все виды с определенным морфология. Плезиозаврия была в 2010 г. Хиллари Кетчум и Роджер Бенсон определяется как таковой таксон на основе стебля: "все таксоны более тесно связаны с Плезиозавр долиходейрус и Pliosaurus brachydeirus чем Августазавр хагдорни ". Кетчум и Бенсон (2010) также создали новую кладу Neoplesiosauria, a таксон на основе узлов это было определено как "Плезиозавр долиходейрус, Pliosaurus brachydeirus, их последний общий предок и все его потомки ".[50] Клада Neoplesiosauria очень вероятно материально идентична Plesiosauria. Sensu Дракенмиллер и Рассел, таким образом, обозначали один и тот же вид, и этот термин должен был заменить это понятие.
Бенсон и другие. (2012) обнаружили, что традиционная плиозаавроидная парафилетический по отношению к Plesiosauroida. Было обнаружено, что Rhomaleosauridae находятся за пределами Neoplesiosauria, но все еще в пределах Plesiosauria. Рано Карнийский пистозавр Бобозавр оказался на шаг впереди, чем Августазавр по отношению к плезиозаврам и, следовательно, он по определению представляет собой базового известного плезиозавра. Этот анализ был сосредоточен на базальных плезиозаврах и, следовательно, только на одном производном плиозавриде и одном криптоклидиан были включены, в то время как эласмозавриды не были включены вообще. Более подробный анализ, опубликованный Бенсоном и Дракенмиллером в 2014 году, не смог разрешить отношения между линиями в основании Плезиозаврии.[53]

Следующее кладограмма следует анализу Бенсона и Дракенмиллера (2014).[53]

| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Описание
Размер

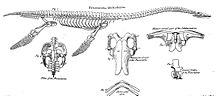
В целом, длина взрослых плезиозавров варьировала от 1,5 метров (4,9 футов) до примерно 15 метров (49 футов). Таким образом, в группу вошли одни из крупнейших морских высшие хищники в Окаменелости, примерно равный самому длинному ихтиозавры, мозазавриды, акулы и зубатые киты по размеру. Некоторые останки плезиозавров, такие как набор тщательно реконструированных фрагментов нижней челюсти длиной 2,875 метра (9,43 фута), сохранившийся в Музей Оксфордского университета и относиться к Pliosaurus rossicus (ранее упоминалось Стретозавр[54] и Лиоплевродон ), обозначенная длина 17 метров (56 футов). Однако недавно утверждалось, что его размер в настоящее время не может быть определен из-за того, что они плохо реконструированы, а длина 12,7 метра (42 фута) более вероятна.[55] MCZ 1285, образец, в настоящее время относящийся к Кронозавр королевский, от Раннемеловой период из Австралия, по оценкам, длина черепа составляла 2,21–2,85 м (7,3–9,4 фута).[55][56]
Скелет
Типичный плезиозавр имел широкое, плоское тело и короткое хвостик. Плезиозавры сохранили у своих предков две пары конечностей, которые превратились в большие ласты.[57] Плезиозавры были связаны с более ранними Nothosauridae,[58] у которого было более похожее на крокодила тело. Расположение ласт необычно для водных животных, поскольку, вероятно, все четыре конечности использовались для перемещения животного по воде вверх и вниз. Хвост, скорее всего, использовался только для помощи в управлении направлением. Это контрастирует с ихтиозавры а позже мозазавры, в котором хвост обеспечивал главную движущую силу.[59]
Чтобы привести в действие ласты, плечевой пояс и таз были сильно изменены, превратившись в широкие костные пластинки на нижней стороне тела, которые служили поверхностью прикрепления для больших групп мышц, способных тянуть конечности вниз. В плече коракоидный стал самым большим элементом, покрывающим большую часть груди. В лопатка был намного меньше, образуя внешнюю переднюю кромку ствола. К середине он продолжался в ключица и, наконец, небольшой межключичная кость. Как и большинство четвероногие плечевой сустав образован лопаткой и коракоидом. В тазу костная пластинка образована седалищная кость сзади и больше лобковая кость перед ней. В подвздошная кость, который у наземных позвоночных несет вес задней конечности, стал маленьким элементом сзади, больше не прикрепленным ни к лобковой, ни к бедренной кости. Тазобедренный сустав образован седалищной и лобковой костью. Грудные и тазовые пластины соединялись между собой пластрон костная клетка, образованная парными ребра живота что у каждого была средняя и внешняя секции. Такое расположение обездвиживало весь ствол.[59]
Чтобы стать ластами, конечности значительно изменились. Конечности были очень большими, каждая примерно равной длине туловища. Передние и задние конечности сильно напоминали друг друга. В плечевая кость в плече, а бедренная кость в верхней части ноги превратились в большие плоские кости, расширенные на их внешних концах. Локтевые и коленные суставы перестали функционировать: нижняя рука и голень не могли сгибаться по отношению к элементам верхней конечности, а образовывали их плоское продолжение. Все внешние кости стали плоскими опорными элементами ласт, плотно соединенными друг с другом и с трудом могли вращаться, сгибаться, растягиваться или раскладываться. Это было верно в отношении локтевая кость, радиус, пястные кости и пальцы, а также большеберцовая кость, малоберцовая кость, плюсневые кости и пальцы ног. Кроме того, чтобы удлинить ласты, число фаланг увеличилось до восемнадцати подряд, что называется гиперфалангия. Ласты не были идеально плоскими, но имели слегка изогнутый верхний профиль, как у профиль, чтобы уметь «летать» по воде.[59]

Хотя плезиозавры мало различались по строению туловища и могут быть названы «консервативными» в этом отношении, между подгруппами были существенные различия в форме шеи и черепа. Плезиозавров можно разделить на два основных морфологических типа, которые различаются головой и шея размер. «Плезиозавроморфы», такие как Cryptoclididae, Elasmosauridae, и Плезиозавры, имел длинные шеи и маленькие головы. «Плиозавроморфы», такие как Pliosauridae и Rhomaleosauridae, имел более короткие шеи с большой удлиненной головой. Изменения длины шеи были вызваны не удлинением отдельных шейных позвонков, а увеличением количества этих шейных позвонков. Эласмозавр имеет семьдесят два шейных позвонка; известный рекорд принадлежит elasmosaurid Альбертонектес, с семьюдесятью шестью шейками матки.[60] Большое количество подразумеваемых суставов подсказывало ранним исследователям, что шея должна была быть очень гибкой; действительно, лебединое изгибание шеи считалось возможным - в исландский плезиозаврами даже называют Svanelur, "лебединые ящерицы". Однако современные исследования подтвердили более раннюю гипотезу Уиллистона о том, что длинные пластинчатые шипы на вершине позвонков, отросток Spinosi, сильно ограничивает вертикальное движение. Хотя горизонтальное изгибание было менее ограниченным, в целом шея должна была быть довольно жесткой и, конечно, не могла изгибаться в змеевидные спирали. Это еще более верно для короткошеих «плиозавромофов», у которых было всего одиннадцать шейных позвонков. В ранних формах амфицеллярный шейные позвонки амфиплат имели двуглавые шейные ребра; более поздние формы имели одноголовые ребра. В оставшейся части позвоночник, номер спинные позвонки варьировала от девятнадцати до тридцати двух, крестцовых позвонков от двух до шести и хвостовых позвонков от двадцати одного до тридцати двух. Эти позвонки все еще обладали первоначальными отростками, унаследованными от наземных предков зауроптеригий, и не превратились в простые диски, подобные рыбам, как это произошло с позвонками ихтиозавров. Хвостовые позвонки имели шевронные кости. Спинные позвонки плезиозавров легко узнать по двум крупным foramina subcentralia, парные сосудистые отверстия на нижней стороне.[59]
Череп плезиозавров показал "эвяпсид "состояние без нижнего височные отверстия, отверстия в нижней части задней части. Верхние височные отверстия образовывали большие отверстия по бокам задней крыши черепа, прикрепляя мышцы к нижней челюсти. Как правило, теменные кости были очень большими, с гребнем средней линии, но чешуйчатые кости обычно образуют арку, исключая теменные кости из затылок. Глазницы большие, как правило, наискось вверх; у плиозаврид глаза были больше направлены вбок. Глаза поддерживали склеральные кольца, форма которого показывает, что они были относительно плоскими, приспособлением для дайвинга. Расположенные спереди внутренние ноздри, хоаны, имеют небные канавки для отвода воды, поток которой будет поддерживаться за счет гидродинамического давления на расположенные сзади, перед глазницами, наружные ноздри во время плавания. Согласно одной из гипотез, во время прохождения через носовые протоки вода «пахла» обонятельным эпителием.[61][62] Однако ближе к задней части во нёбе присутствует вторая пара отверстий; более поздняя гипотеза утверждает, что это настоящие хоаны а передняя пара реально представляла собой парную солевые железы.[63] Расстояние между глазницами и ноздрями было так ограничено, потому что носовые кости были сильно редуцированы, многие виды даже отсутствовали. В предчелюстные кости непосредственно коснулся лобные кости, причем эласмозавриды даже обращаются назад к теменные кости. Часто слезные кости тоже не хватало.[47]

Форма и количество зубов были очень разными. У некоторых форм были сотни игольчатых зубов. У большинства видов были более крупные конические зубы с круглым или овальным сечением. Таких зубов было от четырех до шести в верхней челюсти и от четырнадцати до двадцати пяти в верхней. верхняя челюсть; количество в нижних челюстях примерно равнялось количеству черепа. Зубы помещались в лунки, имели вертикальную морщинистую эмаль и не имели настоящей режущей кромки или карина. У некоторых видов передние зубы были значительно длиннее, чтобы хватать добычу.
Мягкие ткани
Остатки мягких тканей плезиозавров встречаются редко, но иногда, особенно в сланец месторождений, они частично сохранились, например показывая очертания тела. Ранним открытием в этом отношении был голотип Плезиозавр conybeari (на данный момент Аттенборозавр ). Из таких находок известно, что кожа была гладкой, без видимых чешуек, но с небольшими морщинками, что задний край ласт значительно заходил за кости конечностей;[64] и что на хвосте был вертикальный плавник, как сообщает Вильгельм Дамес в его описании Плезиозавр guilelmiimperatoris (на данный момент Селейозавр ).[65] Возможность хвостовой двуустки была подтверждена недавними исследованиями формы хвостового отдела позвоночника. Пантозавр, Криптоклидус и Ромалеозавр zetlandicus.[66][67][68] Исследование 2020 года утверждает, что хвостовой плавник имел горизонтальную конфигурацию.[69]
Палеобиология


Еда
Вероятный источник пищи плезиозавров варьировался в зависимости от того, принадлежали ли они к длинношеей "плезиозавроморфной" форме или короткошейным "плиозавроморфам".
Чрезвычайно длинные шеи "плезиозавроморфов" вызвали спекуляции относительно их функции с того самого момента, как их особенное строение стало очевидным. Конибер предложил три возможных объяснения. Шея могла служить для перехвата быстро движущейся рыбы в погоне. В качестве альтернативы плезиозавры могли отдыхать на морском дне, а голова была отправлена на поиски добычи, что, казалось, подтверждалось тем фактом, что глаза были направлены относительно вверх. Наконец, Конибер предположил возможность того, что плезиозавры плавали по поверхности, позволяя своей шее опускаться вниз в поисках пищи на более низких уровнях. Все эти интерпретации предполагали, что шея очень гибкая. Современное представление о том, что шея на самом деле была довольно жесткой с ограниченным вертикальным движением, потребовало новых объяснений. Одна из гипотез состоит в том, что длина шеи позволяла удивить косяки рыб, причем голова появлялась раньше, чем вид или волна давления хобота могли их предупредить. «Плезиозавроморфы» охотились визуально, о чем свидетельствуют их большие глаза, и, возможно, использовали направленное чувство обоняния. Твердые и мягкотелые головоногие моллюски, вероятно, составляли часть их рациона. Их челюсти были, вероятно, достаточно сильны, чтобы прокусить твердые панцири этого типа добычи. Были найдены ископаемые образцы с раковинами головоногих моллюсков в желудке.[70] В костлявая рыба (Osteichthyes ), которые в юрском периоде стали более разнообразными, вероятно, тоже были добычей. Совершенно другая гипотеза утверждает, что «плезиозавроморфы» питались снизу. Жесткие шеи использовали бы, чтобы бороздить морское дно, поедая бентос. Это было бы доказано длинными бороздами, присутствующими на дне древних морей.[71][72] Такой образ жизни был предложен в 2017 году для Morturneria.[73] «Плезиозавроморфы» не были хорошо приспособлены к ловле крупной, быстро движущейся добычи, поскольку их длинные шеи, хотя и выглядели обтекаемыми, вызывали огромные трение кожи. Санкар Чаттерджи предположил в 1989 г., что некоторые Cryptocleididae были подвесные питатели, фильтрующие планктон. Aristonectes например имел сотни зубов, что позволяло ему просеивать мелкие Ракообразные из воды.[74]
Короткошеие «плиозавроморфы» были лучшими плотоядными животными, или высшие хищники, в своих соответствующих пищевые сети.[75] Они были хищниками-преследователями[76] или устраивать засаду на хищников с добычей различного размера и приспособленных кормушек; их зубы можно было использовать, чтобы проткнуть мягкую добычу, особенно рыбу.[77] Их головы и зубы были очень большими, чтобы хватать и разрывать крупных животных. Их морфология позволяла быстро плавать. Они тоже охотились визуально.
Плезиозавры сами были добычей других хищников, о чем свидетельствуют следы укусов, оставленные акулой, которые были обнаружены на окаменелом плавнике плезиозавра.[78] и окаменелые останки содержимого желудка мозазавра, которые считаются останками плезиозавра.[79]
Скелеты также были обнаружены с гастролиты, камни в их желудках, но помогают ли они расщеплять пищу, особенно головоногих, в мышечном желудке или плавучесть, или и то, и другое не установлено.[80][81] Однако общий вес гастролитов, обнаруженных у различных особей, оказывается недостаточным для изменения плавучести этих крупных рептилий.[82] Первые гастролиты плезиозавра, обнаруженные Мауизавр гарднери, сообщили Гарри Говьер Сили в 1877 г.[83] Количество этих камней на одну особь часто очень велико. В 1949 году окаменелость Альзадазавр (образец SDSM 451, позже переименованный в Стиксозавр ) показал 253 из них.[84] Кроме того, размер отдельных камней часто бывает значительным. В 1991 г. был исследован образец эласмозаврида KUVP 129744, содержащий гастролит диаметром 17 сантиметров и весом 1,3 кг; и камень несколько короче - 1490 грамм. Всего присутствовало сорок семь гастролитов с общим весом тринадцать килограммов. Размер камней был замечен как указание на то, что они были проглочены не случайно, а намеренно, возможно, животное преодолело большие расстояния в поисках подходящего типа камня.[85]
Передвижение
Флиппер движение

Характерная форма тела с четырьмя шлепками вызвала немало спекуляций о том, какие удары использовали плезиозавры. Единственная современная группа с четырьмя ластами - это морские черепахи, которые используют только переднюю пару для движения. Конибер и Бакленд уже сравнивали ласты с птичьими крыльями. Однако такое сравнение было малоинформативным, так как механика полета птиц в этот период была плохо изучена. К середине девятнадцатого века обычно считалось, что плезиозавры использовали гребные движения. Ласты должны были быть перемещены вперед в горизонтальном положении, чтобы минимизировать трение, а затем повернуты в осевом направлении в вертикальное положение, чтобы их оттянуть назад, что привело бы к максимально возможному реактивная сила. Фактически, такой метод был бы очень неэффективным: ход возврата в этом случае не генерирует тяги, а задний ход создает огромную турбулентность. В начале двадцатого века недавно открытые принципы полета птиц подсказали нескольким исследователям, что плезиозавры, такие как черепахи и пингвины, могут летать во время плавания. Это было, например, предложено Эберхард Фраас в 1905 г.,[86] а в 1908 г. Отенио Лотар Франц Антон Луи Абель.[87] В полете движение ласт более вертикальное, его острие описывает овал или восьмерку. В идеале плавник сначала перемещается под наклоном вперед и вниз, а затем, после небольшого втягивания и поворота, пересекает этот путь снизу, чтобы подтянуть его вперед и вверх. Во время обоих гребков вниз и вверх по Принцип Бернулли, прямая и восходящая тяга создается выпуклым изогнутым верхним профилем ласта, передняя кромка слегка наклонена относительно потока воды, а турбулентность минимальна. Однако, несмотря на очевидные преимущества такого способа плавания, в 1924 г. было проведено первое систематическое исследование мускулатуры плезиозавров. Дэвид Мередит Сирс Уотсон пришел к выводу, что они все же выполнили гребное движение.[88]
В середине двадцатого века «модель гребли» Уотсона оставалась доминирующей гипотезой относительно плавательного гребка плезиозавра. В 1957 г. Ламберт Беверли Холстед В то время, используя фамилию Тарло, предложил вариант: задние конечности гребли бы в горизонтальной плоскости, но передние конечности гребли бы, двигаясь вниз и назад.[89][90] В 1975 г. традиционной модели бросили вызов Джейн Энн Робинсон, который возродил «летающую» гипотезу. Она утверждала, что основные группы мышц были оптимально размещены для вертикального движения ласт, а не для горизонтального вытягивания конечностей, и что форма плечевых и тазобедренных суставов препятствовала бы вертикальному вращению, необходимому для гребли.[91] В следующей статье Робинсон предположил, что кинетическая энергия создаваемые силами, действующими на туловище при ударах, сохранялись и высвобождались как упругая энергия в грудной клетке, что обеспечивает особенно эффективную и динамичную двигательную установку.[92]
В модели Робинсона и ход вниз, и ход вверх были бы сильными. В 1982 году ее раскритиковал Самуэль Тарситано, Эберхард Фрей и Юрген Рисс, который утверждал, что, хотя мышцы на нижней стороне плечевых и тазовых пластин были явно достаточно мощными, чтобы тянуть конечности вниз, сопоставимые группы мышц наверху этих пластин для подъема конечностей просто отсутствовали, и если бы они присутствовали , не могли быть применены с применением силы, их выпуклость несла опасность повреждения внутренних органов. Они предложили более ограниченную модель полета, в которой мощный удар вниз сочетался с в значительной степени отсутствием мощности, при этом ласт возвращался в исходное положение за счет импульса движущегося вперед и временно опускающегося тела.[93][94] Эта модифицированная летающая модель стала популярной интерпретацией. Меньше внимания альтернативной гипотезе уделили Стивен Годфри в 1984 году, в котором было предложено, чтобы и передние, и задние конечности выполняли глубокое гребущее движение назад в сочетании с мощным восстановительным движением вперед, напоминая движения передних конечностей морских львов.[95]
В 2010, Фрэнк Сандерс и Кеннет Карпентер опубликовал исследование, в котором пришел к выводу, что модель Робинсона верна. Frey & Riess ошиблись бы в своем утверждении, что плечевые и тазовые пластины не имеют мышц, прикрепленных к их верхним сторонам. Хотя эти группы мышц, вероятно, были не очень мощными, это можно было легко компенсировать за счет больших мышц спины, особенно Musculus latissimus dorsi, который был бы хорошо развит с учетом высоких шипов на позвоночнике. Более того, плоское строение плечевых и тазобедренных суставов ясно указывало на то, что основное движение было вертикальным, а не горизонтальным.[96]
Походка

Как все четвероногие конечностями плезиозавры должны были походка, скоординированный рисунок движения ласт, в данном случае. Из бесконечного числа возможностей на практике внимание в основном было направлено на вопрос о том, двигались ли передняя пара и задняя пара одновременно, так что все четыре ласта задействованы одновременно, или поочередно, причем задействована каждая пара. в очереди. Frey & Riess в 1991 году предложили альтернативную модель, которая имела бы преимущество более непрерывной силовой установки.[97] В 2000 г. Theagarten Lingham-Soliar уклонился от ответа, заключив, что, как и морские черепахи, плезиозавры использовали только переднюю пару для мощного удара. Задняя пара использовалась бы только для управления. Лингхам-Солиар вывел это из формы тазобедренного сустава, которая допускала только ограниченное вертикальное движение. Кроме того, разделение движущей силы и функции рулевого управления облегчило бы общую координацию тела и предотвратило бы слишком экстремальные подача. Он отверг гипотезу Робинсона о том, что упругая энергия хранится в грудной клетке, считая ребра слишком жесткими для этого.[98]
Интерпретация Frey & Riess стала доминирующей, но была оспорена в 2004 году Сандерсом, который экспериментально показал, что, в то время как альтернативное движение могло вызвать чрезмерную качку, одновременное движение вызвало бы только небольшую высоту звука, что могло быть легко контролируется задними ластами. Из других осевых перемещений прокатка можно было контролировать, попеременно задействуя ласты с правой или левой стороны, и рыскание длинной шеей или вертикальным хвостовым плавником. Сандерс не верил, что задняя пара не использовалась для движения, заключая, что ограничения, налагаемые тазобедренным суставом, были очень относительными.[99] В 2010 году Сандерс и Карпентер пришли к выводу, что при чередовании походки турбулентность, вызванная передней парой, препятствовала бы эффективному действию задней пары. Кроме того, длительная фаза планирования после одновременного столкновения была бы очень энергоэффективной.[96] Также возможно, что походка была необязательной и адаптирована к обстоятельствам. Во время быстрого устойчивого преследования было бы полезно другое движение; в засаде одновременный удар сделал бы возможной максимальную скорость. При поиске добычи на большом расстоянии сочетание одновременного движения с скольжением стоило бы меньше всего энергии.[100] В 2017 году исследование Люка Маскатта с использованием модели робота пришло к выводу, что задние ласты активно использовались, что позволило увеличить движущую силу на 60% и повысить эффективность на 40%. Удар был бы наиболее мощным при слегка чередующейся походке, когда задние ласты включались сразу после передних ласт, чтобы извлечь выгоду из их следа. Однако не было бы единой оптимальной фазы для всех условий, походка, вероятно, была изменена, как того требовала ситуация.[101]
Скорость

В общем, определить максимальную скорость вымерших морских существ сложно. Для плезиозавров это усложняется из-за отсутствия единого мнения об их гребке ласт и походке. Нет точных расчетов их Число Рейнольдса. Отпечатки окаменелостей показывают, что кожа была относительно гладкой, без чешуек, и это могло уменьшить форма перетащить.[96] На коже присутствуют мелкие морщинки, которые могли помешать отделению ламинарный поток в пограничный слой и тем самым уменьшил трение кожи.
Устойчивая скорость можно оценить, рассчитав тянуть упрощенной модели тела, к которой можно подойти вытянутый сфероид, и устойчивый уровень выработки энергии мышцы. Первое исследование этой проблемы было опубликовано Джуди Массар в 1988 г.[102] Даже при допущении низкого гидродинамический КПД 0,65 модель Массара, казалось, показывала, что плезиозавры, если бы они были теплокровными, двигались бы со скоростью четыре метра в секунду, или около четырнадцати километров в час, что значительно превышало известные скорости современных дельфинов и китов.[103] Однако в 2002 г. Рёске Мотани показал, что формулы, которые использовал Массар, были ошибочными. Пересчет с использованием исправленных формул дал скорость полметра в секунду (1,8 км / ч) для хладнокровного плезиозавра и полтора метра в секунду (5,4 км / ч) для эндотермический плезиозавр. Даже самая высокая оценка примерно на треть ниже скорости дошедших до нас. Китообразные.[104]
Массар также попытался сравнить скорости плезиозавров со скоростью двух других основных групп морских рептилий, Ихтиозаврия и Mosasauridae. Она пришла к выводу, что плезиозавры были примерно на двадцать процентов медленнее, чем развитые ихтиозавры, которые использовали очень эффективное туннельное движение, колеблющее только хвост, но на пять процентов быстрее, чем мозазавриды, которые, как предполагалось, плавали с неэффективным угрюмым движением ног. тело.[103]
Многие виды плезиозавров могли значительно различаться по скорости плавания, что отражает различные формы тела, присутствующие в группе. В то время как короткошеие «плиозавроморфы» (например, Лиоплевродон ), возможно, были быстрыми пловцами, «плезиозавроморфы» с длинной шеей были созданы больше для маневренности, чем для скорости, замедляемые сильным трением кожи, но способные к быстрому перекатывающемуся движению. Некоторые формы с длинной шеей, такие как Elasmosauridae, также имеют относительно короткие короткие ласты с низким соотношение сторон, дополнительно снижая скорость, но улучшая крен.[105]
Дайвинг
Доступно немного данных, которые точно показывают, насколько глубоко ныряли плезиозавры. То, что они нырнули на значительную глубину, подтверждается следами декомпрессионная болезнь. Руководители плеча и бедра со многими окаменелостями некроз костной ткани из-за слишком быстрого всплытия после глубоких погружений. Однако это не позволяет определить точную глубину, поскольку ущерб мог быть вызван несколькими очень глубокими погружениями или, альтернативно, большим количеством относительно неглубоких спусков. Позвонки не имеют такого повреждения: они, вероятно, были защищены превосходным кровоснабжением, что стало возможным благодаря артериям, входящим в кость через две foramina subcentralia, большие отверстия в их нижней части.[106]
Спуску помог бы отрицательный Сила Архимеда, т. е. плотнее воды. Конечно, это помешало бы появиться снова. Шоу молодых плезиозавров пахиостоз, чрезмерная плотность костной ткани, которая могла иметь увеличенный относительный вес. У взрослых особей более губчатая кость. Гастролиты были предложены как метод увеличения веса[107] или даже как средство достижения нейтрального плавучесть, проглотив или выплюнув их снова по мере необходимости.[108] Их также можно было использовать для повышения стабильности.[109]
Относительно большие глаза Cryptocleididae рассматривались как адаптация к глубокому дайвингу.[нужна цитата ]
Роль хвоста
Исследование 2020 года показало, что зауроптеригианы полагаются на вертикальные удары хвостом во многом как китообразные. У плезиозавров туловище было жестким, поэтому это действие было более ограниченным и было связано с ластами.[69]
Метаболизм
Традиционно предполагалось, что вымершие группы рептилий были хладнокровными, как современные рептилии. Новое исследование последних десятилетий привело к выводу, что некоторые группы, такие как теропод динозавры и птерозавры, скорее всего, были теплокровными. Трудно определить, были ли плезиозавры также теплокровными. Один из признаков высокого метаболизм наличие быстрорастущих фиброламеллярная кость. Однако пахиостоз у молодых особей затрудняет установление наличия такой кости у плезиозавров. Тем не менее, было возможно проверить его появление на более базальных членах более обширной группы, к которой принадлежали плезиозавры, Зауроптеригия. Исследование 2010 года показало, что фиброламеллярная кость изначально присутствовала у зауроптеригов.[110] Последующая публикация в 2013 году показала, что Nothosauridae отсутствовал этот тип костного матрикса, но этот базальный Пистозаврия обладал им, что было признаком более высокого метаболизма.[111] Таким образом, больше скупой предположить, что более производные пистозавры, плезиозавры, также имели более быстрый метаболизм. В статье, опубликованной в 2018 году, на основе количественного остеогистологического моделирования утверждалось, что плезиозавры имеют показатели метаболизма в покое (RMR) у ряда птиц.[112] Однако эти результаты проблематичны с точки зрения общих принципов физиологии позвоночных (см. Закон Клейбера ) и данные изотопных исследований зубной эмали плезиозавра действительно предполагают эндотермию при более низких RMR, с предполагаемой температурой тела прибл. 26 ° С.[113]
Размножение

Как вообще рептилии яйцекладущий, до конца двадцатого века считалось возможным, что более мелкие плезиозавры, возможно, заползли на пляж, чтобы отложить яйца, как современные черепахи. Их сильные конечности и плоская нижняя сторона, казалось, сделали это возможным. Этот метод, например, защищал Холстед. Однако, поскольку у этих конечностей больше не было функциональных локтевых или коленных суставов, а нижняя часть из-за своей плоской поверхности вызывала бы сильное трение, уже в девятнадцатом веке было выдвинуто предположение, что плезиозавры были живородящий. Кроме того, было трудно представить, как самые крупные виды, размером с китов, могли выжить при высадке на берег. Находки окаменелостей эмбрионов ихтиозавров показали, что по крайней мере одна группа морских рептилий родила живых детенышей. Первым, кто заявил, что подобные эмбрионы были обнаружены у плезиозавров, был Гарри Говьер Сили, который сообщил в 1887 году, что приобрел узелок с четырьмя-восемью крошечными скелетами.[114] В 1896 году он более подробно описал это открытие.[115] Если бы они были подлинными, эмбрионы плезиозавров были бы очень маленькими, как у ихтиозавров. Однако в 1982 г. Ричард Энтони Талборн показал, что Сили был обманут "подделанной" окаменелостью гнезда раков.[116]
Настоящий образец плезиозавра, найденный в 1987 году, в конечном итоге доказал, что плезиозавры родили живых детенышей:[117] Эта окаменелость беременной Поликотил латиппинус показывает, что эти животные родили одного крупного детеныша и, вероятно, вложили в свое потомство родительскую заботу, как и современные киты.Птенец был 1,5 метра (пять футов) в длину и, таким образом, крупнее по сравнению с его матерью длиной в пять метров (шестнадцать футов), что указывает на K-стратегия в репродукции.[118] Мало что известно о темпах роста или возможных половой диморфизм.
Социальное поведение и интеллект
Исходя из родительской заботы, на которую указывает большой размер молодых людей, можно сделать вывод, что социальное поведение в целом было относительно сложным.[117] Неизвестно, охотились ли плезиозавры стаями. Их относительный размер мозга кажется типичным для рептилий. Из чувств важны зрение и обоняние, а не слух; эласмозавриды утратили стремени полностью. Было высказано предположение, что у некоторых групп в черепе находились чувствительные к электричеству органы.[119][120]
Палеопатология
Некоторые окаменелости плезиозавра показывают патологии, результат болезни или старости. В 2012 г. нижняя челюсть из Плиозавр был описан с челюстным суставом, явно пораженным артрит, типичный признак старение.[121]
Распределение
Плезиозавры были найдены на всех континентах, в том числе Антарктида.[122]
Стратиграфическое распространение
Ниже приводится список геологических образований, которые дали окаменелости плезиозавра.
В современной культуре
Было высказано предположение, что легенды из морские змеи и современные наблюдения предполагаемых монстры в озерах или море можно объяснить выживанием плезиозавров в наше время. Это криптозоологический предложение было отклонено научное сообщество в целом, который считает, что это основано на фантазиях и лженаука. Предполагаемые туши плезиозавров представляют собой частично разложившиеся трупы гигантские акулы вместо.[128][129][130]
В то время Лохнесское чудовище часто сообщается, что он похож на плезиозавра, но также часто описывают, что он выглядит совершенно иначе. Было представлено несколько причин, по которым это вряд ли может быть плезиозавром. Они включают предположение, что вода в озере слишком холодная для предполагаемого хладнокровный рептилии, чтобы иметь возможность легко выжить, предположение, что дышащих воздухом животных будет легко увидеть всякий раз, когда они появляются на поверхности, чтобы дышать,[131] тот факт, что озеро слишком маленькое и содержит недостаточно пищи, чтобы поддерживать размножающуюся колонию крупных животных, и, наконец, тот факт, что озеро образовалось всего 10 000 лет назад в конце последнего Ледниковый период, а последнее появление в окаменелостях плезиозавров датируется более 66 миллионами лет назад.[132] Частые объяснения наблюдений включают: волны, плавание неодушевленных предметов, фокусы света, плавание известных животных и розыгрыши.[133] Тем не менее, в народном воображении плезиозавры стали отождествляться с монстром Лох-Несса. Это имело преимущество, сделав группу более известной для широкой публики, но недостаток в том, что люди не могут серьезно относиться к этой теме, заставляя палеонтологов снова и снова объяснять, что плезиозавры действительно существуют, а не являются просто созданиями из мифов или фантазий.[134]
Смотрите также
использованная литература
- ^ «Плезиозавр». Словарь Merriam-Webster.
- ^ «Плезиозавр». Dictionary.com Несокращенный. Случайный дом.
- ^ "Справочник плезиозавров". Архивировано из оригинал 4 марта 2016 г.. Получено 20 апреля 2013.
- ^ а б c d е ж г Эванс, М. (2010). «Роли музеев, коллекций и коллекционеров в ранней истории палеонтологии рептилий». В Муди, Ричард; MoodyBuffetaut, E .; MoodyNaish, D .; MoodyMartill, D. M. (ред.). Динозавры и другие вымершие ящеры: историческая перспектива. Геологическое общество Лондона. С. 5–31. ISBN 978-1-86239-311-0.
- ^ Ричард Верстеган, 1605 г., Восстановление истлевшего интеллекта или Nationum Origo, Р. Бруней, Антверпен
- ^ Lhuyd, E., 1699, Lithophylacii Brittannici Ichnographia, sive Lapidum aliorumque Fossilium Brittanicorum singulari figurà insignium, Лондон
- ^ Стьюкли, Вт (1719 г.). «Отчет о слепке почти всего скелета большого животного из очень твердого камня, недавно представленный Королевским обществом из Ноттингемшира». Философские труды. 30 (360): 963–968. Дои:10.1098 / рстл.1717.0053.
- ^ Николлс, Дж., 1795 г., История и древности графства Лестершир. Том I, Джон Николлс, Лондон
- ^ Конибер, У.Д. (1822). "Дополнительные примечания по ископаемым родам Ихтиозавр и Плезиозавр". Труды Лондонского геологического общества. 2: 103–123. Дои:10.1144 / transgslb.1.1.103. S2CID 129545314.
- ^ De la Beche, H.T .; Конибер, W.D. (1821). «Уведомление об открытии нового животного, образующего связь между ихтиозавром и крокодилом, вместе с общими замечаниями по остеологии ихтиозавра». Труды Лондонского геологического общества. 5: 559–594.
- ^ "Plesiosaur_Names". oceansofkansas.com.
- ^ Конибер, У.Д. (1824). "Об открытии почти идеального скелета Плезиозавр". Труды Лондонского геологического общества. 2: 382–389.
- ^ Benson, R.B.J .; Evans, M .; Smith, A.S .; Sassoon, J .; Moore-Faye, S .; Ketchum, H.F .; Форрест, Р. (2013). "Гигантский череп плиозаврида из поздней юры Англии". PLOS ONE. 8 (5): e65989. Bibcode:2013PLoSO ... 865989B. Дои:10.1371 / journal.pone.0065989. ЧВК 3669260. PMID 23741520.
- ^ Хокинс, Т. Х. (1834). «Воспоминания об Ихтиосаури и Плезиозавре; вымерших монстрах древней Земли» (PDF). Релфе и Флетчер. Архивировано из оригинал (PDF) на 2005-08-30.
- ^ Петерсон, А. (2012). «Ужасные ящерицы и гнев Бога: как христианство и романтизм 19 века повлияли на визуальные представления динозавров и наше восприятие древнего мира» (PDF). Стэнфордский исследовательский журнал бакалавриата.
- ^ Хокинс, Т. Х. (1840). Книга Великих морских драконов, Ихтиосаури и Плезиозавров, Гедолим Танинум Моисея. Вымершие монстры Древней Земли. У. Пикеринг, Лондон. С. 1–27.
- ^ Кристофер Макгоуэн, 2001 год, Искатели драконов, Кембридж, Массачусетс, издательство Perseus Publishing
- ^ Оуэн, Р. (1841). "Описание некоторых останков гигантского крокодилового ящера, вероятно морского, из Нижних Зеленых песков в Хите и зубов из той же формации в Мейдстоне, относящихся к роду Полиптиходон". Труды ассоциации геологов. 3: 449–452.
- ^ Эдвард Грей, Джон (1825). «Краткий обзор родов рептилий и амфибий с описанием некоторых новых видов». Анналы философии (Британский музей). 10: 193–217.
- ^ де Бленвиль, Х. М. Д. (1835). "Description de quelques espèces de la Californie, precédée de l'analyse d'une système générale d'Erpetologie et d'Amphibiologie". Nouvelles Archives du Muséum d'Histoire Naturelle (На французском). 4: 233–296.
- ^ Коуп, Э. (1868). "[Резолюция с благодарностью доктора Теофилуса Тернера за пожертвование скелета Эласмозавр Platyurus]". Proc. Акад. Nat. Sci. Phila. 20: 314.
- ^ Коуп, Э. (1868). "Замечания о новом эналиозавре, Эласмозавр Platyurus". Труды Академии естественных наук Филадельфии. 20: 92–93.
- ^ Коуп, Э. (1869). «О рептильных отрядах Pythonomorpha и Streptosauria». Труды Бостонского общества естествознания. XII: 250–266.
- ^ Коуп, Э. (1868). «О новом большом эналиозавре». Американский журнал науки серии. 46 (137): 263–264.
- ^ Коуп, Э. Д. (1869). «Зауроптеригия». Сводка вымерших батрахий и рептилий Северной Америки, часть I. Новая серия. 14. Труды Американского философского общества. С. 1–235.
- ^ Лейди, Дж (1870). «Об Elasmosaurus platyurus Копа». Американский журнал науки серии. 49 (147): 392.
- ^ Коуп, Э. (1870). «Сводка вымерших батрахий и рептилий Северной Америки». Труды Американского философского общества. Новая серия. 14 (1): 1–252. Дои:10.2307/1005355. JSTOR 1005355.
- ^ Коуп, Э. (1870). "Об Elasmosaurus platyurus Cope". Американский журнал науки серии. 50 (148): 140–141.
- ^ Эллис (2003), стр. 129
- ^ Уиллистон, С.В., 1914 г., Водные рептилии прошлого и настоящего. Издательство Чикагского университета. Чикаго, Иллинойс. 251 стр.
- ^ Дэвидсон, Дж. П. (2015). «Непонятые морские рептилии: Художественные реконструкции доисторической морской жизни в конце девятнадцатого века». Труды Канзасской академии наук. 118 (1–2): 53–67. Дои:10.1660/062.118.0107. S2CID 83904449.
- ^ Смит, A.S., 2003, Кладистический анализ плезиозаврии (Reptilia: Sauropterygia). Магистерская диссертация по палеобиологии, Бристольский университет, 91 стр.
- ^ Отеро, Родриго А .; Суарес, Марио; Ле Ру, Якобус П. (2009). «Первое упоминание о плезиозаврах Elasmosaurid (Sauropterygia: Plesiosauria) в верхних слоях формации Dorotea, поздний мел (маастрихт), Пуэрто-Наталес, Чилийская Патагония». Андская геология. 36 (2): 342–350. Дои:10.4067 / s0718-71062009000200008.
- ^ Форрест, Ричард. «Лиоплевродон». Место обитания Плезиозавра. Архивировано из оригинал 15 июля 2011 г.. Получено 18 сентября 2017.
- ^ Форрест, Ричард. "Монстр Арамберри"'". Место обитания Плезиозавра. Архивировано из оригинал 3 сентября 2011 г.. Получено 18 сентября 2017.
- ^ Ларкин, Найджел; О'Коннор, Соня; Парсонс, Деннис (2010). «Виртуальная и физическая подготовка плезиозавра Колларда из Бриджуотер-Бей, Сомерсет, Великобритания». Геологический куратор. 9 (3): 107.
- ^ Форрест, Ричард. "Плезиозавр-воротник". Архивировано из оригинал на 2013-01-17. Получено 31 октября 2012.
- ^ Ларкин, Найджел. «Подготовка и сохранение важного шестифутового скелета плезиозавра для Сомерсетского музея». Получено 31 октября 2012.
- ^ Сато, Тамаки (205). «Новый поликотилидный плезиозавр (Reptilia: Sauropterygia) из формации Bearpaw в верхнемеловом периоде в Саскачеване, Канада». Журнал палеонтологии. 79: 969-980.
- ^ «Hallazgo de un ejemplar completeto de plesiosaurio joven». Архивировано из оригинал на 2013-07-18. Получено 2013-04-22. (На испанском)
- ^ Ледфорд, Х. (2006). «Редкая окаменелость рептилий найдена в Антарктиде». Новости природы. Дои:10.1038 / news061211-4. S2CID 85361720.
- ^ «ХИЩНИК Икс - музей естествознания». 21 марта 2009 г. Архивировано с оригинал 21 марта 2009 г.
- ^ Хигнетт, Кэтрин (22.12.2017). «Плезиозавр: древнее морское чудовище, обнаруженное в Антарктиде». Newsweek. Получено 2017-12-23.
- ^ Риппель, О. (2000). Зауроптеригия I. Handbuch der Paläoherpetologie (на немецком языке). 12А. Verlag Dr. Friedrich Pfeil. С. 1–134.
- ^ Cheng, Y-N .; Wu, X-C .; Джи, К. (2004). «Китайские морские рептилии родили детенышей живыми». Природа. 432 (7015): 383–386. Bibcode:2004Натура 432..383С. Дои:10.1038 / природа03050. PMID 15549103. S2CID 4391810.
- ^ Сторрс, Г. (1993). «Функция и филогения в эволюции зауроптеригов (Diapsida)». Американский журнал науки. 293A: 63–90. Bibcode:1993AmJS..293 ... 63S. Дои:10.2475 / ajs.293.A.63.
- ^ а б Риппель, О., 1997, «Введение в зауроптеригию», В: Callaway, J. M. & Nicholls, E. L. (ред.), Древние морские рептилии С. 107–119. Academic Press, Сан-Диего, Калифорния
- ^ О'Киф, Ф. (2002). «Эволюция морфотипов плезиозавров и плиозавров в Plesiosauria (Reptilia: Sauropterygia)». Палеобиология. 28: 101–112. Дои:10.1666 / 0094-8373 (2002) 028 <0101: teopap> 2.0.co; 2.
- ^ Роджер Б. Дж. Бенсон; Марк Эванс; Патрик С. Дракенмиллер (2012). Лалуэза-Фокс, Карлес (ред.). «Высокое разнообразие, низкая диспропорция и малый размер тела у плезиозавров (рептилий, зауроптеригией) на границе триаса и юры». PLOS ONE. 7 (3): e31838. Bibcode:2012PLoSO ... 731838B. Дои:10.1371 / journal.pone.0031838. ЧВК 3306369. PMID 22438869.
- ^ а б c d Ketchum, H.F .; Бенсон, Р. Б. Дж. (2010). «Глобальные взаимосвязи Plesiosauria (Reptilia, Sauropterygia) и решающая роль выборки таксонов в определении результатов филогенетических анализов». Биологические обзоры Кембриджского философского общества. 85 (2): 361–392. Дои:10.1111 / j.1469-185X.2009.00107.x. PMID 20002391. S2CID 12193439.
- ^ Баккер, Р. (1993). «Циклы вымирания плезиозавров - события, знаменующие начало, середину и конец мелового периода». В Caldwell, W.G.E .; Кауфман, Э. (ред.). Эволюция Западного внутреннего бассейна. Геологическая ассоциация Канады. С. 641–664.
- ^ Druckenmiller, P. S .; Рассел, А. П. (2008). «Филогения Plesiosauria (Sauropterygia) и ее влияние на систематический статус Leptocleidus Andrews, 1922». Zootaxa. 1863: 1–120. Дои:10.11646 / zootaxa.1863.1.1.
- ^ а б Benson, R. B.J .; Дракенмиллер, П. С. (2013). «Фаунистический круговорот морских четвероногих во время юрско-мелового перехода». Биологические обзоры. 89 (1): 1–23. Дои:10.1111 / brv.12038. PMID 23581455. S2CID 19710180.
- ^ Tarlo, L.B.H. (1959). "Стретозавр gen nov., гигантский плиозавр из Киммериджской глины ». Палеонтология. 2 (2): 39–55.
- ^ а б МакГенри, Колин Ричард (2009). "Пожиратель богов: палеоэкология мелового плиозавра" Кронозавр королевский"(PDF): 1–460
- ^ Benson, R. B.J .; Evans, M .; Smith, A. S .; Sassoon, J .; Moore-Faye, S .; Ketchum, H.F .; Форрест, Р. (2013). Батлер, Ричард Дж (ред.). "Гигантский череп плиозаврида из поздней юры Англии". PLOS ONE. 8 (5): e65989. Bibcode:2013PLoSO ... 865989B. Дои:10.1371 / journal.pone.0065989. ЧВК 3669260. PMID 23741520.
- ^ Колдуэлл, Майкл В; 1997b. Модифицированное окостенение перихондра и развитие лопаточных конечностей у ихтиозавров и плезиозавров; Журнал палеонтологии позвоночных 17 (3); 534-547
- ^ Сторрс, Гленн У .; 1990. Филогенетические отношения пахиплеврозавров и нотозавровидных рептилий (Diapsida: Sauropterygia); Журнал палеонтологии позвоночных; 10 (Дополнение к № 3)
- ^ а б c d Смит, Адам Стюарт (2008). «Объяснение окаменелостей 54: Плезиозавры». Геология сегодня. 24 (2): 71–75. Дои:10.1111 / j.1365-2451.2008.00659.x.
- ^ Кубо, Тай; Митчелл, Марк Т .; Хендерсон, Дональд М. (2012). "Альбертонектес вандервельдей, новый эласмозавр (Reptilia, Sauropterygia) из верхнего мела Альберты ». Журнал палеонтологии позвоночных. 32 (3): 557–572. Дои:10.1080/02724634.2012.658124. S2CID 129500470.
- ^ Cruickshank, A.R.I .; Small, P.G .; Тейлор, М.А. (1991). «Дорсальные ноздри и гидродинамическое обоняние под водой у плезиозавров». Природа. 352 (6330): 62–64. Bibcode:1991 Натур. 352 ... 62C. Дои:10.1038 / 352062a0. S2CID 4353612.
- ^ Браун, Д. С .; Круикшэнк А. Р. И. (1994). "Череп келловейского плезиозавра Cryptoclidus eurymerus и зауроптеригийскую щеку ". Палеонтология. 37 (4): 941–953.
- ^ Buchy, M C .; Frey, E .; Солсбери, С. (2006). «Внутренняя анатомия черепа плезиозаврии (Reptilia, Sauropterygia): свидетельство функционального вторичного неба». Lethaia. 39 (4): 289–303. Дои:10.1080/00241160600847488.
- ^ Huene, Ф. фон (1923). "Ein neuer Plesiosaurier aus dem oberen Lias Württembergs". Jahreshefte des Vereins für Vaterländische Naturkunde в Вюртемберге. 79: 1–21.
- ^ Дамы, W (1895). "Die Plesiosaurier der Süddeutschen Liasformation". Abhandlungen der Königlich Preussischen Akademie der Wissenschaften zu Berlin. 1895: 1–81.
- ^ Вильгельм, Британская Колумбия, 2010 г., Новая анатомия криптоклидных плезиозавров с комментариями по осевой локомоции. Докторская диссертация, Университет Маршалла, Хантингтон, Западная Вирджиния. Соединенные Штаты Америки
- ^ Wilhelm, B.C .; О'Киф, Ф. (2010). "Новый частичный скелет Пантозавр полосатый, криптоклеидоидный плезиозавр из верхнеюрской формации Сандэнс в Вайоминге ". Журнал палеонтологии позвоночных. 30 (6): 1736–1742. Дои:10.1080/02724634.2010.521217. S2CID 36408899.
- ^ Смит, Адам С. (2013). «Морфология хвостовых позвонков в Ромалеозавр zetlandicus и обзор свидетельств существования хвостового плавника у Плезиозаврии ". Палудикола. 9 (3): 144–158.
- ^ а б Сенников, А.Г. (2019). «Особенности строения и двигательной функции хвоста при зауроптеригии». Бюллетень биологии. 46 (7): 751–762. Дои:10.1134 / S1062359019070100. S2CID 211217453.
- ^ McHenry, C.R .; Cook, A.G .; Вро, С. (2005). «Плезиозавры донного питания». Наука. 310 (5745): 75. Дои:10.1126 / science.1117241. PMID 16210529. S2CID 28832109.
- ^ «Показано донное питание плезиозавра». Новости BBC. 17 октября 2005 г.. Получено 21 мая 2012.
- ^ Гейстер, Дж (1998). "Lebensspuren von Meersauriern und ihren Beutetieren im mittleren Jura (Callovien) von Liesberg, Schweiz". Фации. 39 (1): 105–124. Дои:10.1007 / bf02537013. S2CID 127249009.
- ^ О'Киф, Ф .; Otero, R .; Soto-Acuña, S .; О'Горман, Дж .; Годфри, S .; Чаттерджи, С. (2017). "Черепная анатомия Morturneria seymourensis из Антарктиды, а также эволюция фильтрующего питания плезиозавров австралийского позднего мела ». Журнал палеонтологии позвоночных. 37 (4): e1347570. Дои:10.1080/02724634.2017.1347570. S2CID 91144814.
- ^ Чаттерджи, С. и Смолл, Б.Дж., 1989, "Новые плезиозавры из верхнего мела Антарктиды", В: Crame, J. (ed) Происхождение и эволюция антарктической биоты, стр. 197-215, Издательство Геологического общества, Лондон.
- ^ "Справочник плезиозавров". Получено 20 апреля 2013.
- ^ Massare, J.A. (1992). «Древние мореплаватели». Естественная история. 101: 48–53.
- ^ Дж. Массаре (1987). «Морфология зубов и кормовые предпочтения мезозойских морских рептилий». J. Vert. Палеонтол. 7 (2): 121–137. Дои:10.1080/02724634.1987.10011647.
- ^ Эверхарт, М. Дж. (2005). "Следы укусов на весле эласмозавра (зауроптеригия; Плезиозаврия) из мела Ниобрара (верхний мел) как вероятное свидетельство кормления пластинчатой акулой, Cretoxyrhina mantelli". Палеонтология позвоночных. 2 (2): 14–24.
- ^ Эверхарт, М. Дж. (2004). «Плезиозавры как пища мозазавров; новые данные о содержимом желудка Tylosaurus proriger (Squamata; Mosasauridae) из формации Ниобрара в западном Канзасе ". Мозазавр. 7: 41–46.
- ^ Уиллистон, Сэмюэл Вендель; 1904. Камни желудка плезиозавров. Наука 20; 565
- ^ Эверхарт, М. Дж. (2000). «Гастролиты, связанные с плезиозавром, остаются в пачке Шарон-Спрингс из сланца Пьер (поздний мел), западный Канзас». Kansas Acad. Sci. Транс. 103 (1–2): 58–69. Дои:10.2307/3627940. JSTOR 3627940.
- ^ Cerda, A; Сальгадо, Л. (2008). "Gastrolitos en un plesiosaurio (Sauropterygia) de la Formación Allen (Campaniano-Maastrichtiano), provincia de Río Negro, Патагония, Аргентина". Амегиниана. 45: 529–536.
- ^ Сили, Х.Г. (1877). "На Mauisaurus gardneri Сили, эласмозавр с основания Голта Фолкстона ". Ежеквартальный журнал Лондонского геологического общества. 33 (1–4): 541–546. Дои:10.1144 / gsl.jgs.1877.033.01-04.32. S2CID 128691833.
- ^ Уэллс, С.П .; Бамп, Дж. Д. (1949). "Альзадазавр пембертони, новый эласмозавр из верхнего мела Южной Дакоты ». Журнал палеонтологии. 23 (5): 521–535.
- ^ Эверхарт, М.Дж. (2000). «Гастролиты, связанные с плезиозавром, остаются в пачке Шарон-Спрингс сланца Пьер (поздний мел), Западный Канзас». Сделки Канзасской академии наук. 103 (1–2): 58–69.
- ^ Фраас, Э. (1905). "Reptilien und Säugetiere in ihren Anpassungserscheinungen an das marine Leben". Jahresheften des Vereins für Vaterländische Naturkunde в Вюртемберге. 29: 347–386.
- ^ Абель, О. (1908). "Die Anpassungsformen der Wirbeltiere an das Meeresleben". Schriften des Vereines zur Verbreitung Naturwissenschaftlicher Kenntnisse в Вене. 48 (14): 395–422.
- ^ Уотсон, Д. (1924). «Плечевой пояс и передние конечности эласмозаврид». Труды Лондонского зоологического общества. 1924 (2): 885–917.
- ^ Тарло, Л. (1957). "Лопатка Pliosaurus macromerus Филлипс ". Палеонтология. 1: 193–199.
- ^ Холстед, Л. (1989). «Передвижение плезиозавра». Журнал геологического общества. 146 (1): 37–40. Bibcode:1989JGSoc.146 ... 37H. Дои:10.1144 / gsjgs.146.1.0037. S2CID 219541473.
- ^ Робинсон, Дж. (1975). «Передвижение плезиозавров». Neues Jahrbuch für Geologie und Paläontologie, Абхандлунген. 149 (3): 286–332.
- ^ Робинсон, Дж. (1977). «Межкорпоральная передача силы у плезиозавров». Neues Jahrbuch für Geologie und Paläontologie, Абхандлунген. 153 (1): 88–128.
- ^ Tarsitano, S .; Рис, Дж. (1982). «Передвижение плезиозавра - полет под водой против гребли». Neues Jahrbuch für Geologie und Paläontologie, Абхандлунген. 164 (1–2): 193–194. Дои:10.1127 / njgpa / 164/1982/188.
- ^ Frey, E .; Рейсс, Дж. (1982). «Соображения относительно передвижения плезиозавра». Neues Jahrbuch für Geologie und Paläontologie, Абхандлунген. 164 (1–2): 188–192. Дои:10.1127 / njgpa / 164/1982/193.
- ^ Годфри, Стивен Дж. (1984). «Субаквальная передвижение плезиозавра: переоценка». Neues Jahrbuch für Geologie und Paläontologie. 1984 (11): 661–672. Дои:10.1127 / njgpm / 1984/1984/661.
- ^ а б c Сандерс, Ф .; Карпентер, К .; Рид, В .; Рид, Дж. (2010). «Плавание плезиозавра восстановлено на основе анализа скелета и экспериментальных результатов». Труды Канзасской академии наук. 113 (1/2): 1–34. Дои:10.1660/062.113.0201. S2CID 86491931.
- ^ Рис, Дж. И Э. Фрей, 1991. "Эволюция подводного полета и передвижения плезиозавров", В: J.M.V. Райнер и Р.Дж. Вуттон (ред.) Биомеханика в эволюции, Кембридж, Англия: Издательство Кембриджского университета, стр. 131-144.
- ^ Лингхэм-Солиар, Т. (2000). «Передвижение плезиозавра: проблема с четырьмя крыльями реальна или просто атеоретическое упражнение?». Neues Jahrbuch für Geologie und Paläontologie, Абхандлунген. 217: 45–87. Дои:10.1127 / njgpa / 217/2000/45.
- ^ Сандерс, Ф .; Карпентер, К .; Рид, В .; Рид, Дж. (2004). «Плавание плезиозавра восстановлено на основе анализа скелета и экспериментальных результатов». Журнал палеонтологии позвоночных. 24: 108A – 109A. Дои:10.1080/02724634.2004.10010643. S2CID 220415208.
- ^ Long, J. H .; Schumaker, J .; Ливингстон, Н .; Кемп, М. (2006). «Четыре ласта или два? Тетраподальное плавание с водным роботом». Биоинспирация и биомиметика. 1 (1): 20–29. Bibcode:2006БиБи .... 1 ... 20л. Дои:10.1088/1748-3182/1/1/003. PMID 17671301.
- ^ Muscutt, Luke E .; Дайк, Гарет; Weymouth, Gabriel D .; Наиш, Даррен; Палмер, Колин; Ганапатисубрамани, Бхаратрам (2017). «Метод плавания плезиозавров с четырьмя ластами обеспечивает эффективное и эффективное передвижение». Труды Королевского общества B. 284 (1861): 20170951. Дои:10.1098 / rspb.2017.0951. ЧВК 5577481. PMID 28855360.
- ^ Massare, J.A. (1988). «Плавательные возможности мезозойских морских рептилий: значение для методов хищничества». Палеобиология. 14 (2): 187–205. Дои:10.1017 / s009483730001191x.
- ^ а б Massare, J. A., 1994, "Плавательные способности мезозойских морских рептилий: обзор", In: L. Maddock et al. (ред.) Механика и физиология плавания животных, Кембридж, Англия: Издательство Кембриджского университета, стр. 133-149.
- ^ Мотани, Р. (2002). «Оценка скорости плавания вымерших морских рептилий: новый взгляд на энергетический подход». Палеобиология. 28 (2): 251–262. Дои:10.1666 / 0094-8373 (2002) 028 <0251: sseoem> 2.0.co; 2.
- ^ О'Киф, Ф. (2001). «Экоморфология геометрии плавников плезиозавра» (PDF). Журнал эволюционной биологии. 14 (6): 987–991. CiteSeerX 10.1.1.579.4702. Дои:10.1046 / j.1420-9101.2001.00347.x. S2CID 53642687.
- ^ Rothschild, B.M .; Сторрс, Г. (2003). «Синдром декомпрессии у плезиозавров (Sauropterygia: Reptilia)». Журнал палеонтологии позвоночных. 23 (2): 324–328. Дои:10.1671 / 0272-4634 (2003) 023 [0324: dsipsr] 2.0.co; 2.
- ^ Тейлор, М.А. (1981). «Плезиозавры - оснастка и балластировка». Природа. 290 (5808): 628–629. Bibcode:1981Натура.290..628Т. Дои:10.1038 / 290628a0. S2CID 10700992.
- ^ Тейлор, M.A., 1993, "Желудочные камни для питания или плавучести? Возникновение и функция гастролитов у морских четвероногих", Philosophical Transactions of the Royal Society of London B 341: 163–175
- ^ Хендерсон, Д. (2006). «Плавающая точка: вычислительное исследование плавучести, равновесия и гастролитов у плезиозавров». Lethaia. 39 (3): 227–244. Дои:10.1080/00241160600799846.
- ^ Кляйн, Н. (2010). «Гистология длинных костей зауроптеригии из нижних тростников Германской впадины дает неожиданные последствия для филогении». PLOS ONE. 5 (7): e11613. Bibcode:2010PLoSO ... 511613K. Дои:10.1371 / journal.pone.0011613. ЧВК 2908119. PMID 20657768.
- ^ Краль, Анна; Кляйн, Николь; Сандер, П. Мартин (2013). "Эволюционные последствия расходящихся гистологий длинных костей Нотозавр и Пистозавр (Зауроптеригия, триас) ». BMC Эволюционная биология. 13: 123. Дои:10.1186/1471-2148-13-123. ЧВК 3694513. PMID 23773234.
- ^ Fleischle, Corinna V .; Винтрич, Таня; Сандер, П. Мартин (2018-06-06). «Количественные гистологические модели предполагают эндотермию у плезиозавров». PeerJ. 6: e4955. Дои:10.7717 / peerj.4955. ISSN 2167-8359. ЧВК 5994164. PMID 29892509.
- ^ Бернар, Орелиен; Лекюер, Кристоф; Винсент, Пегги; Амиот, Ромен; Бардет, Натали; Баффето, Эрик; Куни, Жиль; Фурель, Франсуа; Мартино, Франсуа; Мазен, Жан-Мишель; Приёр, Абель (11.06.2010). «Регулирование температуры тела некоторыми морскими рептилиями мезозоя». Наука. 328 (5984): 1379–1382. Bibcode:2010Sci ... 328.1379B. Дои:10.1126 / science.1187443. ISSN 1095-9203. PMID 20538946. S2CID 206525584.
- ^ Сили, Х.Г. (1888). «О способах развития молодежи в Плезиозавр". Отчет Британской ассоциации содействия развитию науки; Сентябрь в Манчестере. 1887: 697–698.
- ^ Seeley, H.G .; 1896; "На колчеданной конкреции из Лиаса Уитби, которая, по-видимому, показывает внешнюю форму тела эмбрионов одного вида Плезиозавр", Годовой отчет Йоркширского философского общества стр. 20–29
- ^ Талборн, Р.А. (1982). «Эмбрионы лиасовых плезиозавров переосмыслены как норы креветок». Палеонтология. 25: 351–359.
- ^ а б O'Keefe, F.R .; Чиаппе, Л.М. (2011). «Живородство и история жизни K-Selected в мезозойском морском плезиозавре (Reptilia, Sauropterygia)». Наука. 333 (6044): 870–873. Bibcode:2011Sci ... 333..870O. Дои:10.1126 / science.1205689. PMID 21836013. S2CID 36165835.
- ^ Уэлш, Дженнифер (11 августа 2011 г.). «Беременные окаменелости предполагают, что древние« морские чудовища »родились живыми молодыми». LiveScience. Получено 21 мая 2012.
- ^ О'Горман, J.P .; Гаспарини, З. (2013). "Пересмотр Sulcusuchus erraini (Sauropterygia, Polycotylidae) из верхнего мела Патагонии, Аргентина ". Алчеринга. 37 (2): 161–174. Дои:10.1080/03115518.2013.736788. S2CID 131429825.
- ^ Foffa, D .; Sassoon, J .; Манжета, A.R .; Mavrogordato, M.N .; Бентон, М.Дж. (2014). «Сложная ростральная нервно-сосудистая система гигантского плиозавра». Naturwissenschaften. 101 (5): 453–456. Bibcode:2014NW .... 101..453F. Дои:10.1007 / s00114-014-1173-3. PMID 24756202. S2CID 7406418.
- ^ Sassoon, J .; Noe, L.F .; Бентон, М.Дж. (2012). «Анатомия черепа, таксономические значения и палеопатология верхнеюрского плиозавра (Reptilia: Sauropterygia) из Вестбери, Уилтшир, Великобритания». Палеонтология. 55 (4): 743–773. Дои:10.1111 / j.1475-4983.2012.01151.x.
- ^ Чаттерджи, Санкар; Смолл, Брайан Дж .; Никелл, М. В. (1984). «Позднемеловые морские рептилии из Антарктиды;». Антарктический журнал США. 19 (5): 7–8.
- ^ ماذا تعرفون عن الـ "بليزوصور"؟ شاهدوا ما تم اكتشافه في سوريا مؤخراً. CNN (на арабском). 30 августа 2017.
- ^ Таня Винтрич; Сёдзи Хаяси; Александра Хусай; Ясухиса Накадзима; П. Мартин Сандер (2017). «Триасовый плезиозавр скелет и гистология кости информируют об эволюции уникального строения тела». Достижения науки. 3 (12): e1701144. Bibcode:2017SciA .... 3E1144W. Дои:10.1126 / sciadv.1701144. ЧВК 5729018. PMID 29242826.
- ^ «Материал: YPM 1640» в «Возникновении эласмозавридов ...» Эверхарт (2006), стр. 173.
- ^ а б c d «Таблица 13.1: Плезиозавры» в Эверхарте (2005). Океаны Канзаса, стр. 245.
- ^ «Материал: YPM 1640», в «Возникновение эласмозавридов ...» Эверхарт (2006), стр. 172.
- ^ Анонимный (отчет AP). Японский ученый утверждает, что морское существо могло быть связано с видом акул. Нью-Йорк Таймс, 26 июля 1977 г.
- ^ "Морское чудовище или акула: туша предполагаемого плезиозавра". paleo.cc.
- ^ Кимура С., Фуджи К. и другие. Морфология и химический состав рогового волокна неопознанного существа, пойманного у берегов Новой Зеландии. В КПК 1978 г. С. 67–74.
- ^ Великий морской змей, Antoon Cornelis Oudemans, 2009, Cosimo Inc. ISBN 978-1-60520-332-4 п. 321.
- ^ "Жизнь". Новый ученый.
- ^ "Существа Кроули". www.crawley-creatures.com. Архивировано из оригинал на 2009-05-21. Получено 2013-04-22.
- ^ Эллис (2003), стр. 1–3.
дальнейшее чтение
- Callaway, J.M .; Николлс, Э. Л. (1997). «Зауроптеригия». Древние морские рептилии. Академическая пресса. ISBN 978-0-12-155210-7.
- Карпентер, К. (1996). «Обзор плезиозавров с короткой шеей из мелового периода в западной части Северной Америки». Neues Jahrbuch für Geologie und Paläontologie, Абхандлунген. 201 (2): 259–287. Дои:10.1127 / njgpa / 201/1996/259.
- Карпентер, К. 1997. Сравнительная анатомия черепа двух североамериканских меловых плезиозавров. Стр. 91–216, в Calloway J. M. и E. L. Nicholls, (ред.), Ancient Marine Reptiles, Academic Press, San Diego.
- Карпентер, К. (1999). «Ревизия североамериканских эласмозавров из мелового периода западных интерьеров». Палудикола. 2 (2): 148–173.
- Cicimurri, D .; Эверхарт, М. (2001). «Эласмозавр с содержимым желудка и гастролитами из сланца Пьер (поздний мел) Канзаса». Пер. Канс. Акад. Наука. 104 (3 & 4): 129–143. Дои:10.1660 / 0022-8443 (2001) 104 [0129: AEWSCA] 2.0.CO; 2.
- Коуп, Э. Д. (1868). "Замечания о новом эналиозавре, Эласмозавр Platyurus". Труды Академии естественных наук Филадельфии. 20: 92–93.
- Эллис, Р. 2003: Морские драконы (Kansas University Press )
- Эверхарт, М. Дж. (2002). «Где бродили эласмозавры». Доисторические времена. 53: 24–27.
- Эверхарт, М.Дж. 2005. «Где бродили эласмозавры», Глава 7 в Океаны Канзаса: естественная история внутренней западной части моря, Издательство Индианского университета, Блумингтон, 322 с.
- Эверхарт, М.Дж. 2005. Океаны Канзаса: естественная история внутренней западной части моря. Издательство Индианского университета, Блумингтон, 322 стр.
- Эверхарт, М.Дж. (2005). «Гастролиты, связанные с плезиозаврами, останки в пачке Шарон-Спрингс (поздний мел) сланца Пьер, Западный Канзас». Kansas Acad. Sci. Транс. 103 (1–2): 58–69.
- Эверхарт, Майкл Дж (2006). «Возникновение Elasmosaurids (Reptilia: Plesiosauria) в меле Ниобрара в Западном Канзасе». Палудичила. 5 (4): 170–183.
- Хампе, О., 1992: Курьер Forsch.-Inst. Зенкенберг 145: 1-32
- Лингхэм-Солиар, Т. (1995). "в". Фил. Пер. R. Soc. Лондон. 347: 155–180.
- О'Киф, Ф. Р. (2001). «Кладистический анализ и таксономическая ревизия Plesiosauria (Reptilia: Sauropterygia);». Acta Zool. Фенница. 213: 1–63.
- (), 1997: в Отчеты Национального центра научного образования, 17.3 (Май / июнь 1997 г.) стр. 16–28.
- Каддуми, Х. Ф., 2009. Ископаемые остатки фауны Харрана и прилегающих территорий. Публикации Музея естественной истории Вечной реки, Иордания. 324 с.
- Сторрс, Г. В., 1999. Исследование плезиозаврии (Diapsida: Sauropterygia) из мела Ниобрара (верхний мел) в центральной части Северной Америки. Палеонтологические материалы Канзасского университета, (Н.С.), No. 11, 15 с.
- Уэллс, С. П. 1943. Плезиозавры эласмозавров с описанием нового материала из Калифорнии и Колорадо. Мемуары Калифорнийского университета 13: 125-254. фиг.1-37., пл.12-29.
- Уэллс, С. П. 1952. Обзор североамериканских меловых эластозавров. Публикации Калифорнийского университета по геологии 29: 46-144, фиг. 1-25.
- Уэллс, С. П. 1962. Новый вид эластозавров из апта Колумбии и обзор плезиозавров мелового периода. Публикации Калифорнийского университета по геологии 46, 96 стр.
- Уайт, Т., 1935: в Случайные статьи Boston Soc. Nat. Hist. 8: 219-228
- Уиллистон, С. У. 1890. Новый плезиозавр из мелового периода Ниобрара в Канзасе. Труды Канзасской академии наук 12: 174-178, 2 рис.
- Уиллистон, С. У. (1902). "Восстановление Dolichorhynchops osborni, новый меловой плезиозавр ». Бюллетень науки Канзасского университета. 1 (9): 241–244, 1 пласт.
- Уиллистон, С. У. 1903. Плезиозавры Северной Америки. Полевой Колумбийский музей, публикация 73, серия геологии 2 (1): 1-79, 29 пл.
- Уиллистон, С. У. (1906). «Североамериканские плезиозавры: Эласмозавр, Чимолиазавр, и Поликотил". Американский журнал науки. 21 (123): 221–234, 4 пл. Bibcode:1906AmJS ... 21..221W. Дои:10.2475 / ajs.s4-21.123.221.
- Уиллистон, С. В. (1908). «Североамериканские плезиозавры: Тринакромерум". Журнал геологии. 16 (8): 715–735. Bibcode:1908JG ..... 16..715Вт. Дои:10.1086/621573.
- Бардет, Натали; Каппетта, Анри; Переда Субербиола, Ксабье (2000). «Фауна морских позвоночных из позднемеловых фосфатов Сирии». Геологический журнал. Кембриджский университет. 137 (3): 269–290. Дои:10.1017 / S0016756800003988.
внешние ссылки
- Сайт Плезиозавра. Ричард Форрест.
- Справочник плезиозавров. Адам Стюарт Смит.
- Плезиозаврия техническое определение в Справочнике плезиозавров
- Часто задаваемые вопросы о плезиозаврах. Раймонд Таддеус К. Анког.
- Океаны Канзаса Палеонтология. Майк Эверхарт.
- "Окаменелость плезиозавра найдена в Бриджуотер-Бей ". Служба округа музеев Сомерсерта. (самое известное ископаемое)
- "Охотники за окаменелостями обнаружили 50-тонное чудовище из доисторических глубин ". Аллан Холл и Марк Хендерсон. Times Online, 30 декабря 2002 г. (Монстр Арамберри)
- Рептилии триаса жили молодыми.
- Молодь плезиозавра Бриджуотер-Бэй
- Насколько хороша летопись окаменелостей плезиозавра? Блог Laelaps.
| ||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||






