Анатомия птиц - Википедия - Bird anatomy

Анатомия птиц, или физиологическая структура из птицы 'тел, показывает множество уникальных приспособлений, в основном помогающих полет. У птиц есть свет система скелета и легкий, но мощный мускулатура который вместе с кровеносный и дыхательные системы способен на очень высокий метаболический ставки и кислород питания, позвольте птице летать. Развитие клюв привело к эволюция специально адаптированного пищеварительная система. Эти анатомический специализации заработали птиц свои собственные учебный класс в позвоночное животное филюм.
Система скелета
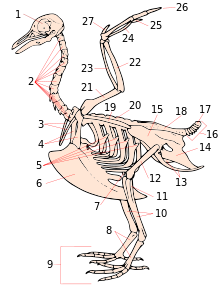
1. череп
2. шейные позвонки
3. фуркула
4. коракоидный
5. крючковидные отростки ребер
6. киль
7. надколенник
8. предплюсна
9. цифры
10. большеберцовая кость (большеберцовая кость )
11. малоберцовая кость (большеберцовая кость )
12. бедренная кость
13. седалищная кость (безымянный )
14. лобок (безымянный)
15. Illium (безымянный)
16. хвостовые позвонки
17. пигостиль
18. синсакрум
19. лопатка
20. спинные позвонки
21. плечевая кость
22. локтевая кость
23. радиус
24. запястье (запястный сустав )
25. пясть (запястный сустав )
26. цифры
27. Alula
У птиц много полых костей (пневматизированный ) с перекрещиванием стойки или же фермы для структурных сила. Количество полых костей варьируется у разных видов, хотя больше всего у крупных летающих и планирующих птиц. Респираторный воздушные мешочки часто образуют воздушные карманы в полуполых костях скелета птицы.[1] Кости ныряющие птицы часто менее полые, чем у не ныряющих видов. Пингвины, гагары,[2] и тупики полностью лишены пневматических костей.[3][4] Нелетающие птицы, Такие как страусы и эму, пневматизировали бедра[5] и, в случае эму, пневматический шейные позвонки.[6]
Осевой скелет
Скелет птицы высоко адаптированный за полет. Он очень легкий, но достаточно прочный, чтобы выдерживать подчеркивает взлета, полета и посадки. Одна из ключевых адаптаций - это слияние кости в одиночный окостенения, такой как пигостиль. Из-за этого у птиц обычно меньше костей, чем у других наземных позвоночных. У птиц тоже нет зубов или даже нет челюсть, а вместо этого клюв, который намного легче. Клювы многих птенцов имеют выступ, называемый яичный зуб, что облегчает их выход из амниотическое яйцо, который падает после проникновения в яйцо.
Позвоночник
Эта секция нужны дополнительные цитаты для проверка. (Январь 2020) (Узнайте, как и когда удалить этот шаблон сообщения) |
Позвоночный столб делится на пять частей: позвонки:
- Шейный (11–25): (шея)
- Туловище: (спинной или грудной) позвонки обычно срастаются в нотариум.
- Synsacrum: (сросшиеся позвонки спины также срослись с бедрами / тазом). Этот регион похож на крестец у млекопитающих и уникальна для голубей, потому что представляет собой слияние крестцового, поясничного и хвостового позвонков. Он прикреплен к тазу и поддерживает наземные движения ног голубя.
- Хвостовой (5-10): эта область похожа на копчик у млекопитающих и помогает контролировать движение перьев во время полета.
- Пигостиль (хвост): эта область состоит из 4-7 сросшихся позвонков и является местом прикрепления перьев.


Шея птицы состоит из 13–25 шейные позвонки позволяя птицам иметь повышенную гибкость.[7] Гибкая шея позволяет многим птицам с неподвижными глазами более продуктивно двигать головой и сосредотачивать взгляд на объектах, находящихся близко или далеко.[8] У большинства птиц шейных позвонков примерно в три раза больше, чем у людей, что обеспечивает повышенную стабильность во время быстрых движений, таких как полет, посадка и взлет.[9] Шея играет роль в покачивании головой, что присутствует как минимум у 8 из 27 отрядов птиц, включая Колумбообразные, Galliformes, и Gruiformes.[10] Качание головой - это оптокинетический ответ который стабилизирует окружение птиц, когда они чередуются между фазой толчка и фазой удержания.[11] Качание головой синхронно с ногами, так как голова движется в соответствии с остальным телом.[11] Данные различных исследований показывают, что основная причина покачивания головой у некоторых птиц заключается в стабилизации их окружения, хотя неясно, почему некоторые, но не все отряды птиц демонстрируют покачивание головой.[12]
Птицы единственные позвоночные иметь сросшиеся ключицы и килевидная грудина.[7] Киловая грудина служит местом прикрепления мышц, используемых во время полета или плавания.[7] Нелетающие птицы, такие как страусы, лишены килевой грудины и имеют более плотные и тяжелые кости по сравнению с летающими птицами.[13] Плавучие птицы имеют широкую грудину, ходячие птицы - длинную, а у летающих птиц грудина почти одинаковой ширины и высоты.[14]
Сундук состоит из фуркула (поперечный рычаг) и коракоидный (ключица), которая вместе с лопатка, сформировать грудной пояс. Сторона груди образована ребрами, которые встречаются у грудины (средняя линия груди).
Ребра
У птиц есть крючковые процессы на ребрах. Это крючковидные отростки кости, которые помогают укрепить грудную клетку, перекрывая ребро позади них. Эта особенность также встречается у туатара (Сфенодон ).
Череп
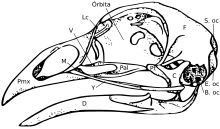
Череп состоит из пяти основных костей: лобной (верхняя часть головы), теменной (затылок), предчелюстной и носовой (верхняя часть головы). клюв ) и нижней челюсти (нижний клюв). Череп нормальной птицы обычно весит около 1% от общей массы тела птицы. Глаз занимает значительную часть черепа и окружен склеротическое кольцо для глаз, кольцо крошечных костей. Эта характеристика также наблюдается у их кузенов-рептилий.
Вообще говоря, птичьи черепа состоят из множества мелких неперекрывающихся костей. Педоморфоз, поддержание предкового состояния у взрослых, как полагают, способствовало эволюции птичьего черепа. По сути, черепа взрослых птиц будут напоминать ювенильную форму их черепа. теропод динозавр предки.[15] По мере развития птичьего происхождения и появления педоморфоза они утратили заглазничная кость за глазом, эктоптеригоид на задней стороне неба и зубов.[16][17] В нёбо структуры также сильно изменились с изменениями, в основном сокращениями, наблюдаемыми в птиергоиде, небной части и скуловой кости. Также произошло уменьшение аддукторных камер. [17] Все эти состояния наблюдаются у ювенильных форм их предков. В предчелюстной кость также гипертрофированный сформировать клюв, в то время как верхняя челюсть уменьшилась, как предполагают оба аспекта развития [15] и палеонтологические [18] исследования. Это расширение клюва произошло одновременно с потерей функциональной руки и развитием точки на передней части клюва, напоминающей «палец».[17] В предчелюстная кость также известно, что он играет большую роль в пищевом поведении рыб.[19][20]
Строение черепа птиц имеет важное значение для их пищевого поведения. Птицы демонстрируют независимое движение костей черепа, известное как черепной кинезис. Краниальный кинезис у птиц бывает нескольких форм, но все они возможны благодаря анатомии черепа. Животные с большими перекрывающимися костями (включая предков современных птиц)[21] имеют акинетические (не кинетические) черепа.[22][23] По этой причине утверждалось, что педоморфный птичий клюв можно рассматривать как эволюционное новшество.[17]
У птиц есть диапсид череп, как у рептилий, с предлакримальной ямкой (присутствует у некоторых рептилий). Череп имеет одиночный затылочный мыщелок.[24]
Аппендикулярного скелета
Эта секция нужны дополнительные цитаты для проверка. (Сентябрь 2018 г.) (Узнайте, как и когда удалить этот шаблон сообщения) |
Плечо состоит из лопатка (лопатка), коракоидный, и плечевая кость (плечо). Плечевая кость присоединяется к радиус и локтевая кость (предплечье), чтобы сформировать локоть. В запястье и пясть образуют «запястье» и «руку» птицы, а цифры сливаются вместе. Кости крыла очень легкие, поэтому птице легче летать.
Бедра состоят из таза, который включает в себя три основные кости: подвздошная кость (верхняя часть бедра), седалищная кость (стороны бедра) и лобок (перед бедра). Они слиты в один ( безымянная кость ). Безыменные кости имеют эволюционное значение, поскольку они позволяют птицам откладывать яйца. Они встречаются в вертлужная впадина (тазобедренный сустав) и сочленяется с бедренной костью, которая является первой костью задней конечности.
Верхняя часть ноги состоит из бедренной кости. В коленном суставе бедро соединяется с большеберцовая кость (голень) и малоберцовая кость (сторона голени). В предплюсна образует верхнюю часть стопы, пальцы составляют пальцы ног. Кости ног у птиц самые тяжелые, что способствует низкому центру тяжести, что облегчает полет. Скелет птицы составляет всего около 5% от общей массы тела.
У них сильно вытянутый четырехлучевой таз, похожий на некоторых рептилий. Задняя конечность имеет внутриплюсневой сустав, который встречается также у некоторых рептилий. Имеется обширное сращение позвонков туловища, а также сращение с грудной пояс.
Крылья
Ноги
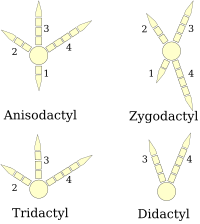
(схемы правой ноги)
Птичьи лапки классифицируются как анизодактиль, зигодактиль, гетеродактиль, синдактиль или же пампродактиль.[25] Анизодактиль - наиболее распространенное расположение пальцев у птиц, с тремя пальцами вперед и одним назад. Это распространено в певчие птицы и другие сидящие птицы, а также охотничьи птицы любят орлы, ястребы, и соколы.
Синдактилия, встречающаяся у птиц, похожа на анизодактилию, за исключением того, что второй и третий пальцы (внутренний и средний обращенные вперед пальцы) или три пальца слиты вместе, как в пояс зимородка Серил Альцион. Это характерно для Ракообразные (зимородки, пчеловоды, ролики, так далее.).
Зигодактиль (от греч. Ζυγον, а ярмо ) у ног два пальца обращены вперед (цифры два и три) и два назад (пальцы один и четыре). Такое расположение чаще всего встречается в древесный виды, особенно те, которые лазают дерево стволами или карабкаться сквозь листву. Зигодактилия встречается в попугаи, дятлы (включая мерцает ), кукушки (включая Roadrunners ), и немного совы. Найдены следы зигодактиля, датируемые 120–110 гг. Ма (рано Меловой ), За 50 миллионов лет до первых идентифицированных окаменелостей зигодактиля.[26]
Гетеродактилия похожа на зигодактилию, за исключением того, что третья и четвертая цифры указывают вперед, а первая и вторая - назад. Это встречается только в трогоны, тогда как пампродактиль - это устройство, при котором все четыре пальца могут быть направлены вперед, или птицы могут вращать два внешних пальца назад. Это характеристика стрижи (Apodidae ).
Эволюция
Изменение задних конечностей
Значительное сходство в строении задних конечностей птиц и динозавры связано с их способностью ходить на двух ногах, или двуногие.[27] В ХХ веке преобладало мнение, что переход к двуногому движению произошел за счет превращения передних конечностей в крылья. Современные ученые считают, что, наоборот, это было необходимым условием возникновения полета.[28]
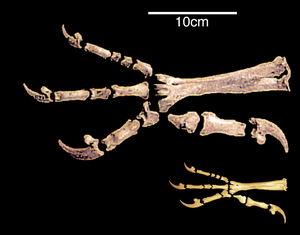
Переход к использованию для движения только задних конечностей сопровождался увеличением жесткости поясничной и крестцовой областей. Лобковые кости птиц и некоторых двуногих динозавров повернуты назад. Ученые связывают это со смещением центра тяжести тела назад. Причиной такого сдвига называют переход к двуногию или развитие мощных передних конечностей, как у археоптерикса.[29][30] Большой и тяжелый хвост двуногих динозавров мог быть дополнительной опорой. Частичное сокращение хвоста и последующее формирование пигостиля происходило из-за отклонения назад первого пальца задней конечности; у динозавров с длинным жестким хвостом развитие стопы происходило иначе. Этот процесс, по-видимому, шел параллельно у птиц и динозавров. В общем, анизодактильная лапа, которая также лучше схватывает и позволяет уверенно двигаться как по земле, так и по веткам, является наследственной для птиц. На этом фоне выделяются птерозавры, которые в процессе неудачных эволюционных изменений не могли полноценно передвигаться на двух ногах, а вместо этого разработали летательный аппарат, принципиально отличный от птиц.[31]
Изменения передних конечностей
Изменения задних конечностей не повлияли на расположение передних конечностей, которые у птиц остались сбоку, а у динозавров перешли на парасагиттальную ориентацию.[32] В то же время, как передние, освобожденные от опорной функции, имели широкие возможности для эволюционных изменений. Сторонники гипотезы бега считают, что полет сформировался за счет быстрого бега, подпрыгивания и затем скольжения. Передние конечности могли использоваться для захвата после прыжка или как «сети для ловли насекомых», животные могли ими махать, помогая себе во время прыжка. Согласно древесной гипотезе, предки птиц лазили по деревьям с помощью передних конечностей, а оттуда планировали, после чего переходили в бегство.[33]
Мышечная система

У большинства птиц около 175 различных мышц, в основном управляющих крыльями, кожей и ногами. В целом мышечная масса птиц сконцентрирована вентрально. Самыми крупными мышцами у птицы являются грудные или большая грудная мышца, которые управляют крыльями и составляют около 15–25% массы тела летающей птицы. Они обеспечивают мощный ход крыла, необходимый для полета. Глубокая мышца (под) грудных мышц - это supracoracoideus, или малая грудная мышца. Он поднимает крыло между взмахами крыльев. Обе группы мышц прикрепляются к килю грудины. Это примечательно, потому что у других позвоночных есть мышцы, поднимающие верхние конечности, как правило, прикрепленные к участкам задней части позвоночника. Супракоракоидус и грудные мышцы вместе составляют около 25–40% от общей массы тела птицы.[34] Каудальнее грудных и супракоракоидных мышц находятся внутренние и внешние косые мышцы живота, которые сжимают брюшную полость. Кроме того, присутствуют и другие мышцы живота, которые расширяют и сокращают грудную клетку и удерживают грудную клетку. Мышцы крыла, как видно на обозначенных изображениях, в основном выполняют функцию разгибания или сгибания локтя, движения крыла в целом или разгибания или сгибания отдельных пальцев. Эти мышцы работают, чтобы настроить крылья для полета и всех других действий.[34] Состав мышц действительно различается у разных видов и даже внутри семей.[35]

Птицы имеют уникальные шеи, удлиненные со сложной мускулатурой, поскольку они должны позволять голове выполнять функции, для которых другие животные могут использовать грудные конечности.[34]

Кожные мышцы помогают птице в ее полете, регулируя перья, которые прикреплены к кожной мышце, и помогают птице в ее маневрах в полете, а также помогают в брачных ритуалах.
На туловище и хвосте всего несколько мышц, но они очень сильные и необходимы птице. К ним относятся lateralis caudae и levator caudae, которые контролируют движение хвоста и разгибание прямых мышц, придавая хвосту большую площадь поверхности, что помогает птице держаться в воздухе, а также помогает при повороте.[34]
Состав мышц и адаптация различаются в зависимости от теорий адаптации мышц в зависимости от того, произошла ли эволюция полета сначала от взмахов руками или от скольжения.[36]
Покровная система

Напольные весы
В напольные весы птиц состоят из кератина, как клювы, когти и шпоры. Они встречаются в основном на пальцах ног и лапках (голени птиц), обычно до тибио-предплюсневого сустава, но у некоторых птиц их можно найти и выше. Во многих орлы и совы ноги оперены до пальцев ног (но не включая).[37][38][39] У большинства птиц чешуйки существенно не перекрываются, за исключением случаев зимородки и дятлы. Первоначально считалось, что чешуя и щитки птиц гомологичный рептилиям;[40] однако более поздние исследования показывают, что чешуя у птиц заново эволюционировала после эволюции перьев.[41][42][43]
Эмбрионы птиц начинают развитие с гладкой кожи. На ногах роговой или самый внешний слой этой кожи может ороговевать, утолщаться и образовывать чешуйки. Эти шкалы можно разделить на:
- Канчелла - мелкие чешуйки, которые на самом деле представляют собой просто утолщение и уплотнение кожи, пересеченную мелкими бороздками.
- Scutella - чешуя, которая не такая большая, как щитки, такие как те, что встречаются на каудальный или задняя часть курицы плюсна.
- Щитки - самые крупные чешуи, обычно на передний поверхность плюсны и спинной поверхность пальцев ног.
Ряды щитков на передней части плюсны можно назвать «акрометатарзием» или «акротарзием».
Ретикулы расположены на латеральный и медиальный поверхности (стороны) стопы и изначально считались отдельными чешуйками. Однако гистологические и эволюционное развитие работа в этой области показала, что в этих структурах отсутствует бета-кератин (признак чешуи рептилий) и они полностью состоят из альфа-кератина.[42][44] Это, наряду с их уникальной структурой, привело к предположению, что на самом деле это перьевые почки, задержанные на раннем этапе развития.[42]
Рамфотека и подотека
Купюры многих куликов имеют Тельца Хербста которые помогают им находить добычу, спрятанную под мокрым песком, путем обнаружения незначительных перепадов давления в воде.[45] Все современные птицы могут двигать частями верхней челюсти относительно корпуса мозга. Однако это более заметно у некоторых птиц и может быть легко обнаружено у попугаев.[46]
Область между глазом и клювом сбоку от головы птицы называется знания. Эта область иногда бывает без перьев, а кожа может быть окрашена, как у многих видов баклан семья.
Чешуйчатый покров, имеющийся на лапах птиц, называется подотеками.
Клюв
Клюв, клюв или рострум - это внешняя анатомическая структура птиц, которая используется для еды и для прихорашиваться, манипулирование объектами, убийство добычи, борьба, поиск пищи, ухаживание и кормление молодняка. Хотя клювы значительно различаются по размеру, форме и цвету, у них схожая основная структура. Два костных выступа - верхняя и нижняя челюсти - покрыты тонким ороговевшим слоем эпидермиса, известным как рамфотека. У большинства видов два отверстия, известные как ноздри, ведут к дыхательной системе.
Дыхательная система



Из-за высокого скорость метаболизма требуется для полета, птицы имеют высокую потребность в кислороде. Их высокоэффективные дыхательная система помогает им удовлетворить этот спрос.
Хотя у птиц есть легкие, они представляют собой довольно жесткие структуры, которые не расширяются и не сжимаются, как у млекопитающих, рептилий и многих земноводных. Вместо этого структуры, которые действуют как сильфоны, проветривать легкие - это воздушные мешочки, которые распространены по большей части тела птиц.[47] Воздушные мешки перемещают воздух в одном направлении через парабронхи жестких легких.[48][49] Хотя легкие птиц меньше, чем у млекопитающих сопоставимого размера, воздушные мешочки составляют 15% от общего объема тела, в то время как у млекопитающих легкие альвеолы, которые действуют как сильфоны, составляют всего 7% от общего объема корпуса.[50] Стенки воздушных мешочков не имеют хорошего кровоснабжения и поэтому не играют прямой роли в газообмен.
Птицам не хватает диафрагма, и поэтому используют их межреберный и Пресс расширять и сокращать всю свою торако-брюшную полость, таким образом ритмично изменяя объемы всех своих воздушных мешочков в унисон (иллюстрация справа). Активной фазой дыхания у птиц является выдох, требующий сокращения их дыхательных мышц.[49] Расслабление этих мышц вызывает вдох.
Три различных группы органов выполняют дыхание: передний воздушные мешки (межключичный, шейный и передний грудной), легкие, а задний воздушные мешки (задние грудные и брюшные полости). Обычно в системе девять воздушных мешков;[49] однако это число может варьироваться от семи до двенадцати, в зависимости от вида птиц. Воробьиные имеют семь воздушных мешков, так как ключичные воздушные мешки могут соединяться или сливаться с передними грудными мешками.
При вдыхании воздух окружающей среды сначала попадает в птицу через ноздри откуда он нагревается, увлажняется и фильтруется в носовых проходах и верхних частях трахеи.[50] Оттуда воздух попадает в нижнюю трахея и продолжает чуть дальше сиринкс, после чего трахея разветвляется на две главные бронхи, идя к двум легким. Первичные бронхи входят в легкие, чтобы стать внутрилегочными бронхами, которые выделяют набор параллельных ветвей, называемых вентробронхами, и, чуть дальше, эквивалентный набор дорсобронхов.[51] Концы внутрилегочных бронхов выпускают воздух в задние воздушные мешки на каудальный конец птицы. Каждая пара дорсо-вентробронхов связана большим количеством параллельных микроскопических воздушных капилляров (или парабронхов), где газообмен происходит.[51] Когда птица вдыхает, трахейный воздух течет через внутрилегочные бронхи в задние воздушные мешки, а также в дорсобронхи (но не в вентробронхи, отверстия которых во внутрилегочные бронхи ранее считались плотно закрытыми при вдохе).[51] Однако более поздние исследования показали, что аэродинамика бронхиальной архитектуры направляет вдыхаемый воздух от отверстий вентробронхов в продолжение внутрилегочного бронха в направлении дорсобронхов и задних воздушных мешков.[48][52]). Из дорсобронхов воздух проходит через парабронхи (и, следовательно, через газообменник) в вентробронхи, откуда воздух может выходить только в расширяющиеся передние воздушные мешочки. Итак, во время вдоха расширяются и задний, и передний воздушные мешки,[51] задние воздушные мешки заполняются свежим вдыхаемым воздухом, а передние воздушные мешочки - «отработанным» (бедным кислородом) воздухом, только что прошедшим через легкие.
Во время выдоха считалось, что внутрилегочные бронхи сильно сужены между областью ответвления вентробронхов и областью ответвления дорсобронхов.[51] Но теперь считается, что более сложные аэродинамические характеристики имеют тот же эффект.[48][52] Таким образом, сокращающиеся задние воздушные мешки могут опорожняться только в дорсобронхи. Оттуда свежий воздух из задних воздушных мешочков проходит через парабронхи (в том же направлении, что и при вдохе) в вентробронхи. Дыхательные пути, соединяющие вентробронхи и передние воздушные мешки с внутрилегочными бронхами, открываются во время выдоха, позволяя воздуху с низким содержанием кислорода выходить из этих двух органов через трахею наружу.[51] Таким образом, насыщенный кислородом воздух постоянно (в течение всего цикла дыхания) проходит через парабронхи в одном направлении.[1]

Кровоток через легкие птицы направлен под прямым углом к потоку воздуха через парабронхи, образуя перекрестно-проточную систему обмена (см. Иллюстрацию слева).[51][53] В парциальное давление кислорода в парабронках уменьшается по длине как O2 проникает в кровь. Кровеносные капилляры, выходящие из теплообменника около входа воздушного потока, потребляют больше O2 чем капилляры, уходящие около выходного конца парабронхов. Когда содержимое всех капилляров смешивается, конечное парциальное давление кислорода смешанной легочной венозной крови выше, чем у выдыхаемого воздуха,[51][53] но тем не менее составляет менее половины вдыхаемого воздуха,[51] таким образом достигается примерно такое же системное парциальное давление кислорода в артериальной крови, что и млекопитающие делают с их легкими типа мехов.[51]
Трахея - это область мертвый космос: воздух с низким содержанием кислорода, который он содержит в конце выдоха, является первым воздухом, который повторно входит в задние воздушные мешочки и легкие. По сравнению с дыхательные пути млекопитающих объем мертвого пространства у птицы в среднем в 4,5 раза больше, чем у млекопитающих того же размера.[51][50] Птицы с длинной шеей неизбежно будут иметь длинные трахеи, и поэтому должны делать более глубокий вдох, чем млекопитающие, чтобы сделать поправку на больший объем мертвого пространства. У некоторых птиц (например, лебедь-кликун, Лебедь Cygnus, то белая колпица, Platalea leucorodia, то Американский журавль, Grus americana, а в шлеме curassow, Pauxi pauxi) трахея, длина которой у некоторых кранов может достигать 1,5 м,[51] скручивается взад и вперед внутри тела, резко увеличивая вентиляцию мертвого пространства.[51] Назначение этой необычной особенности неизвестно.
Воздух проходит через легкие в одном направлении как во время выдоха, так и во время вдоха, в результате чего, за исключением бедного кислородом воздуха мертвого пространства, оставшегося в трахее после выдоха и вдыхаемого в начале вдоха, смешивание нового богатого кислородом воздуха с отработанным воздухом практически отсутствует. воздух с низким содержанием кислорода (как в легкие млекопитающих ), изменяясь только (от богатого кислородом до бедного кислородом), когда он движется (в одном направлении) через парабронхи.
В легких птиц нет альвеолы как это делают легкие млекопитающих. Вместо этого они содержат миллионы узких проходов, известных как парабронхи, соединяющие дорсобронхи с вентробронхами на обоих концах легких. Воздух течет вперед (каудально к краниальному) через параллельные парабронхи. Эти парабронхи имеют сотовые стены. Ячейки сот - это тупиковые воздушные пузырьки, называемые предсердие, какой проект радиально от парабронхов. В предсердие являются сайтом газообмен путем простого распространения.[54] Кровоток вокруг парабронхов (и их предсердий) образует перекрестно-текущий газообменник (см. Диаграмму слева).[51][53]
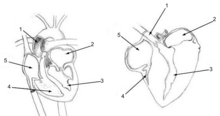
У всех видов птиц, за исключением пингвина, есть небольшая область легких, посвященная «неопульмоническим парабронхам». Эта неорганизованная сеть микроскопических трубок ответвляется от задних воздушных мешков и беспорядочно открывается как в дорсо-, так и в вентробронхи, а также непосредственно во внутрилегочные бронхи. В отличие от парабронхов, в которых воздух движется однонаправленно, поток воздуха в неопульмональных парабронхах является двунаправленным. Неопульмональные парабронхи никогда не составляют более 25% от общей поверхности газообмена птиц.[50]
В сиринкс - звуковой голосовой орган птиц, расположенный у основания трахеи птицы. Как и у млекопитающих гортань, звук создается вибрацией воздуха, проходящего через орган. Сиринкс позволяет некоторым видам птиц издавать чрезвычайно сложные вокализации, даже имитируя человеческую речь. У некоторых певчих птиц сиринкс может издавать более одного звука одновременно.
Сердечно-сосудистая система
Птицы имеют четырехкамерный сердце,[55] вместе с млекопитающими и некоторыми рептилиями (в основном крокодилы ). Эта адаптация обеспечивает эффективный перенос питательных веществ и кислорода по всему телу, обеспечивая птицам энергию для полета и поддержания высокого уровня активности. А колибри с рубиновым горлом Сердце бьется до 1200 ударов в минуту (около 20 ударов в секунду).[56]
Пищеварительная система


Обрезать

Многие птицы обладают мускулистым мешочком вдоль пищевод называется обрезать. Урожай предназначен как для смягчения пищи, так и для регулирования ее прохождения через систему путем временного хранения. Размер и форма урожая у птиц весьма различны.[57] Члены семьи Columbidae, Такие как голуби, производят питательные урожай молока который скармливается их молодым регургитация.[58]
Провентрикулюс
Птичий желудок состоит из двух органов, преджелудок и желудок которые работают вместе во время пищеварение. Провентрикулус представляет собой трубку в форме стержня, которая находится между пищеводом и желудком и выделяет соляная кислота и пепсиноген в пищеварительный тракт.[58] Кислота превращает неактивный пепсиноген в активный протеолитический фермент, пепсин, который разбивает конкретные пептидные связи нашел в белки, чтобы произвести набор пептиды, которые аминокислотные цепи которые короче, чем исходный диетический белок.[59][60] В желудочный сок (соляная кислота и пепсиноген) смешиваются с содержимым желудка в результате мышечных сокращений желудка.[61]
Желудок
Мышечный желудок состоит из четырех мышечных полос, которые вращают и измельчают пищу, перемещая пищу из одной области в другую внутри желудка. Желудок некоторых видов травоядных птиц, например индейки и перепелов,[57] содержит мелкие частицы песка или камня, называемые гастролиты которые глотает птица, чтобы помочь в процессе измельчения, выполняя функцию зубы. Камни в желудке похожи между птицами и динозавры, оставившие гастролиты как следы окаменелостей.[58]
Кишечник
Частично переваренное и измельченное содержимое желудка, теперь называемое болюсом, передается в кишечник, куда панкреатический и кишечные ферменты завершить переваривание легкоусвояемой пищи. Затем продукты пищеварения всасываются через кишечник. слизистая оболочка в кровь. Кишечник заканчивается через толстую кишку в вентиляционном отверстии или клоака который служит общим выходом для почечных и кишечных экскрементов, а также для откладки яиц.[62] Однако, в отличие от млекопитающих, многие птицы не выделяют большие части (грубые корма) непереваренной пищи (например, перья, мех, фрагменты костей и шелуху семян) через клоаку, а отрыгивают их, как пищевые гранулы.[63][64]
Пьянство
Птицы пьют тремя основными способами: под действием силы тяжести, сосанием и использованием языка. Жидкость также получают из пищи.
Большинство птиц неспособны глотать из-за «всасывающего» или «откачивающего» действия перистальтика в их пищевод (как это делают люди) и пить, многократно поднимая голову после наполнения рта, чтобы позволить жидкости течь под действием силы тяжести, метод, обычно описываемый как «потягивание» или «опрокидывание».[65]Заметным исключением является Columbidae; на самом деле, согласно Конрад Лоренц в 1939 г .:
порядок распознается по единственной поведенческой характеристике, а именно, что при питье вода накачивается перистальтикой пищевода, которая происходит без исключения внутри отряда. Однако единственная другая группа, которая демонстрирует такое же поведение, - это Pteroclidae, находится рядом с голубями именно по этой, несомненно, очень старой характеристике.[66]
Хотя это общее правило все еще остается в силе, с того времени было сделано несколько исключений в обоих направлениях.[65][67]
Кроме того, специализированные нектар кормушки типа солнечных птиц (Нектаринииды ) и колибри (Trochilidae ) пить, используя выступающие желобчатые или желобчатые языки, а попугаи (Psittacidae кипятком воды.[65]
У многих морских птиц есть железы возле глаз, которые позволяют им пить морскую воду. Из ноздрей выводится лишняя соль. Многие пустынные птицы получают воду, в которой они нуждаются, исключительно из пищи. Удаление азотистых отходов в виде мочевой кислоты снижает физиологическую потребность в воде,[68] поскольку мочевая кислота не очень токсична, и поэтому ее не нужно разбавлять большим количеством воды.[69]
Репродуктивная и мочеполовая системы


У самцов два яички которые становятся в сотни раз больше за время сезон размножения производить сперма.[70] Яички у птиц обычно асимметричны, у большинства птиц левое яичко больше.[71] Самки птиц в большинстве семейств имеют только одну функциональную яичник (левый), соединенный с яйцеводом - хотя на эмбриональной стадии у каждой самки птицы присутствуют два яичника. У некоторых видов птиц функционируют два яичника, и порядок Apterygiformes всегда сохраняйте оба яичника.[72][73]
У большинства самцов птиц нет фаллос. У самцов видов без фаллоса сперма хранится в семенные клубочки в пределах клоакальный выпуклость перед совокуплением. В течение совокупление, самка отводит хвост в сторону, а самец садится на самку сзади или спереди (как в птица ) или движется очень близко к ней. В клоаки затем коснитесь, чтобы сперма попала в репродуктивные пути женщины. Это может произойти очень быстро, иногда менее чем за полсекунды.[74]
Сперма хранится в хранилище женской спермы трубочки на срок от недели до более 100 дней,[75] в зависимости от вида. Затем яйца будут оплодотворены индивидуально по мере их выхода из яичников, прежде чем скорлупа кальцинируется в яйцевод. После того, как яйцо откладывает самка, эмбрион продолжает развиваться в яйце вне женского тела.

Много водоплавающая птица и некоторые другие птицы, такие как страус и индюк, обладать фаллос.[76] Это, по-видимому, наследственное состояние птиц; большинство птиц потеряли фаллос.[77] Считается, что длина связана с конкуренция спермы у видов, которые обычно спариваются много раз за сезон размножения; сперма, отложенная ближе к яичникам, с большей вероятностью достигнет оплодотворения.[78][79] The longer and more complicated phalli tend to occur in waterfowl whose females have unusual anatomical features of the vagina (such as dead end sacs and clockwise coils). These vaginal structures may be used to prevent penetration by the male phallus (which coils counter-clockwise). In these species, copulation is often violent and female co-operation is not required; the female ability to prevent fertilization may allow the female to choose the father for her offspring.[79][80][81][82] When not copulating, the phallus is hidden within the proctodeum compartment within the cloaca, just inside the vent.
After the eggs hatch, parents provide varying degrees of care in terms of food and protection. Ранний birds can care for themselves independently within minutes of hatching; альтриальный hatchlings are helpless, blind, and naked, and require extended parental care. The chicks of many ground-nesting birds such as куропатки и кулики are often able to run virtually immediately after hatching; such birds are referred to as очаговый. The young of hole-nesters, though, are often totally incapable of unassisted survival. The process whereby a chick acquires feathers until it can fly is called "fledging".
Some birds, such as pigeons, geese, and red-crowned cranes, remain with their mates for life and may produce offspring on a regular basis.
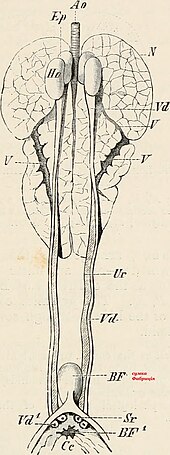
Почка
Птичий почки function in almost the same way as the more extensively studied mammalian kidney, but with a few important adaptations; while much of the anatomy remains unchanged in design, some important modifications have occurred during their evolution. A bird has paired kidneys which are connected to the lower gastrointestinal tract through the мочеточники. Depending on the bird species, the кора makes up around 71-80% of the kidney's mass, while the мозговое вещество is much smaller at about 5-15% of the mass. Blood vessels and other tubes make up the remaining mass. Unique to birds is the presence of two different types of нефроны (the functional unit of the kidney) both reptilian-like nephrons located in the cortex and mammalian-like nephrons located in the medulla. Reptilian nephrons are more abundant but lack the distinctive loops of Henle seen in mammals. The urine collected by the kidney is emptied into the клоака through the ureters and then to the colon by reverse перистальтика.

Нервная система
Birds have acute eyesight—raptors (хищные птицы ) have vision eight times sharper than humans—thanks to higher densities of photoreceptors in the retina (up to 1,000,000 per square mm in Buteos, compared to 200,000 for humans), a high number of нейроны в optic nerves, a second set of eye muscles not found in other animals, and, in some cases, an indented ямка which magnifies the central part of the visual field. Many species, including колибри и альбатросы, have two foveas in each eye. Many birds can detect polarised light.
The avian ear is adapted to pick up on slight and rapid changes of pitch found in bird song. General avian tympanic membrane form is ovular and slightly conical. Morphological differences in the middle ear are observed between species. Ossicles within green finches, blackbirds, song thrushes, and house sparrows are proportionately shorter to those found in pheasants, Mallard ducks, and sea birds. In song birds, a syrinx allows the respective possessors to create intricate melodies and tones. The middle avian ear is made up of three semicircular canals, each ending in an ampulla and joining to connect with the macula sacculus and lagena, of which the cochlea, a straight short tube to the external ear, branches from.[83]
Birds have a large brain to body mass ratio. This is reflected in the advanced and complex интеллект птиц.
Immune system
В иммунная система of birds resembles that of other animals. Birds have both врожденный и адаптивный immune systems. Birds are susceptible to опухоли, immune deficiency and autoimmune diseases.
Bursa of fabricius
Функция
В bursa of fabricius, also known as the cloacal bursa, is a lymphoid organ which aids in the production of В-лимфоциты в течение гуморальный иммунитет. The bursa of fabricius is present during juvenile stages but curls up, and in the воробей is not visible after the sparrow reaches sexual maturity.[84]
Анатомия
The bursa of fabricius is a circular pouch connected to the superior dorsal side of the клоака . The bursa is composed of many folds, known as plica, which are lined by more than 10,000 follicles encompassed by connective tissue and surrounded by мезенхима. Each follicle consists of a cortex that surrounds a medulla. The cortex houses the highly compacted В-лимфоциты, whereas the medulla houses лимфоциты loosely.[85] The medulla is separated from the lumen by the эпителий and this aids in the transport of эпителиальные клетки into the lumen of the bursa. There are 150,000 B lymphocytes located around each follicle.[86]
Смотрите также
Рекомендации
- ^ а б Ritchison, Gary. "Ornithology (Bio 554/754):Bird Respiratory System". Университет Восточного Кентукки. Получено 2007-06-27.
- ^ Gier, H. T. (1952). "The air sacs of the loon" (PDF). Аук. 69 (1): 40–49. Дои:10.2307/4081291. JSTOR 4081291. Получено 2014-01-21.
- ^ Smith, Nathan D. (2011). "Body mass and foraging ecology predict evolutionary patterns of skeletal pneumaticity in the diverse "waterbird" clade". Эволюция. 66 (4): 1059–1078. Дои:10.1111/j.1558-5646.2011.01494.x. PMID 22486689. S2CID 42793145.
- ^ Fastovsky, David E.; Weishampel, David B. (2005). The Evolution and Extinction of the Dinosaurs (второе изд.). Cambridge, New York, Melbourne, Madrid, Cape Town, Singapore, São Paulo: Cambridge University Press. ISBN 978-0-521-81172-9. Получено 2014-01-21.
- ^ Bezuidenhout, A.J.; Groenewald, H.B.; Soley, J.T. (1999). "An anatomical study of the respiratory air sacs in ostriches" (PDF). Ондерстепортский журнал ветеринарных исследований. The Onderstepoort Veterinary Institute. 66 (4): 317–325. PMID 10689704. Получено 2014-01-21.
- ^ Wedel, Mathew J. (2003). "Vertebral pneumaticity, air sacs, and the physiology of sauropod dinosaurs" (PDF). Палеобиология. 29 (2): 243–255. Дои:10.1666/0094-8373(2003)029<0243:vpasat>2.0.co;2. Получено 2014-01-21.
- ^ а б c "Скелет". fsc.fernbank.edu. Получено 2018-03-16.
- ^ Telecommunications, Interactive Media - Nebraska Educational. "Project Beak: Adaptations: Skeletal System: Neck Vertebrae". projectbeak.org. Получено 2018-03-16.
- ^ Hogenboom, Melissa. "How birds see straight". Получено 2018-04-14.
- ^ "Why do pigeons bob their heads when they walk? Everyday Mysteries: Fun Science Facts from the Library of Congress". www.loc.gov. Получено 2018-04-14.
- ^ а б Troje, Nikolaus; Frost, Barrie (February 2000). "Head-Bobbing in pigeons: How stable is the hold phase?" (PDF). Журнал экспериментальной биологии. 203 (Pt 5): 935–940. PMID 10667977.
- ^ Frost, B.J (1978). "The optokinetic basis of head-bobbing in the pigeon". Журнал экспериментальной биологии. 74: 187–195. CiteSeerX 10.1.1.556.8783.
- ^ "Flightless Birds". SKELETONS: Museum of Osteology (Oklahoma City).
- ^ DÜZLER, A.; ÖZGEL, Ö.; DURSUN, N. (2006). "Morphometric analysis of the sternum in avian species" (PDF). Turkish Journal of Veterinary and Animal Sciences. 30: 311–314. ISSN 1303-6181.
- ^ а б Bhullar, Bhart-Anjan S .; Маруган-Лобон, Хесус; Расимо, Фернандо; Бевер, Гейб С .; Rowe, Timothy B.; Norell, Mark A.; Abzhanov, Arhat (2012-05-27). "Birds have paedomorphic dinosaur skulls". Природа. 487 (7406): 223–226. Bibcode:2012Natur.487..223B. Дои:10.1038/nature11146. ISSN 1476-4687. PMID 22722850. S2CID 4370675.
- ^ Лушар, Антуан; Viriot, Laurent (2011). "From snout to beak: the loss of teeth in birds". Тенденции в экологии и эволюции. 26 (12): 663–673. Дои:10.1016/j.tree.2011.09.004. PMID 21978465.
- ^ а б c d Bhullar, Bhart-Anjan S .; Хэнсон, Майкл; Фаббри, Маттео; Pritchard, Adam; Бевер, Гейб С .; Hoffman, Eva (2016-09-01). "How to Make a Bird Skull: Major Transitions in the Evolution of the Avian Cranium, Paedomorphosis, and the Beak as a Surrogate Hand". Интегративная и сравнительная биология. 56 (3): 389–403. Дои:10.1093/icb/icw069. ISSN 1540-7063. PMID 27371392.
- ^ Huang, Jiandong; Wang, Xia; Hu, Yuanchao; Лю, Цзя; Peteya, Jennifer A.; Clarke, Julia A. (2016-03-15). «Новый орнитюрин из раннего мелового периода Китая проливает свет на эволюцию раннего экологического и черепного разнообразия птиц». PeerJ. 4: e1765. Дои:10.7717 / peerj.1765. ISSN 2167-8359. ЧВК 4806634. PMID 27019777.
- ^ LAUDER, GEORGE V. (1982-05-01). "Patterns of Evolution in the Feeding Mechanism of Actinopterygian Fishes". Американский зоолог. 22 (2): 275–285. Дои:10.1093/icb/22.2.275. ISSN 1540-7063.
- ^ Шеффер, Бобб; Rosen, Donn Eric (1961). "Major Adaptive Levels in the Evolution of the Actinopterygian Feeding Mechanism". Американский зоолог. 1 (2): 187–204. Дои:10.1093/icb/1.2.187. JSTOR 3881250.
- ^ Simonetta, Alberto M. (1960-09-01). "On the Mechanical Implications of the Avian Skull and Their Bearing on the Evolution and Classification of Birds". Ежеквартальный обзор биологии. 35 (3): 206–220. Дои:10.1086/403106. ISSN 0033-5770. S2CID 85091693.
- ^ Lingham-Soliar, Theagarten (1995-01-30). "Anatomy and functional morphology of the largest marine reptile known, Mosasaurus hoffmanni (Mosasauridae, Reptilia) from the Upper Cretaceous, Upper Maastrichtian of The Netherlands". Фил. Пер. R. Soc. Лондон. B. 347 (1320): 155–180. Bibcode:1995RSPTB.347..155L. Дои:10.1098 / рстб.1995.0019. ISSN 0962-8436.
- ^ Холлидей, Кейси М .; Witmer, Lawrence M. (2008). "Cranial kinesis in dinosaurs: intracranial joints, protractor muscles, and their significance for cranial evolution and function in diapsids". Журнал палеонтологии позвоночных. 28 (4): 1073–1088. Дои:10.1671/0272-4634-28.4.1073. S2CID 15142387.
- ^ Wing, Leonard W. (1956) Natural History of Birds. Компания Рональда Пресс.
- ^ Proctor, N. S. & Lynch, P. J. (1998) Manual of Ornithology: Avian Structure & Function. Издательство Йельского университета. ISBN 0300076193
- ^ Lockley, M. G.; Li, R .; Harris, J.D .; Matsukawa, M.; Liu, M. (2007). "Earliest zygodactyl bird feet: Evidence from Early Cretaceous roadrunner-like tracks". Naturwissenschaften. 94 (8): 657–665. Bibcode:2007NW.....94..657L. Дои:10.1007/s00114-007-0239-x. PMID 17387416. S2CID 15821251.
- ^ Gatesy S. M. Locomotor evolution on the line to modern birds // Mesozoic Birds: above the heads of dinosaurs/ Eds L. M. Chiappe, L. M. Witmer. — Berkeley : Univ. California Press, 2002. — P. 432–447
- ^ Du Brul, E. Lloyd (1962). "The general phenomenon of bipedalism". Американский зоолог. 2 (2): 205–208. Дои:10.1093/icb/2.2.205.
- ^ Kurochkin E. N. Synopsis of Mesozoic Birds and Early evolution of Class Aves // Archaeopteryx. — 1995 b. —13. — P. 47–66.
- ^ Long, C. A.; Zhang, G. P.; George, T. F.; Long, C. F. (2003). "Physical theory, origin of flight, and synthesis proposed for birds". Журнал теоретической биологии. 224 (1): 9–26. Дои:10.1016/S0022-5193(03)00116-4. PMID 12900201.
- ^ Long, C. A.; Zhang, G. P.; George, T. F.; Long, C. F. (2003). "Physical theory, origin of flight, and synthesis proposed for birds". Журнал теоретической биологии. 224 (1): 9–26. Дои:10.1016/S0022-5193(03)00116-4. PMID 12900201.
- ^ Kurochkin E. N. Synopsis of Mesozoic Birds and Early evolution of Class Aves // Archaeopteryx. — 1995 b. —13. — P. 47–66.
- ^ Bogdanovich I. A. Once more about origin of birds and fl ight: “cursorial” or “arboreal”? // Vestnik zoologii. — 2007. — 41, N 3. — P. 283–284.
- ^ а б c d Proctor, Noble S., Lynch, Patrick J. (1993). Manual of Ornithology. Нью-Хейвен и Лондон: Издательство Йельского университета. С. 149–170. ISBN 978-0-300-07619-6.
- ^ Picasso, Mariana B. J.; Mosto, María C. (2018). "Wing myology of Caracaras (Aves, Falconiformes): muscular features associated with flight behavior". Зоология позвоночных. 68 (2): 177–190.
- ^ Tobalske, Bret W (2016). "Evolution of avian flight: muscles and constraints on performance". Философские труды Королевского общества B: биологические науки. 371 (1704): 20150383. Дои:10.1098/rstb.2015.0383. ЧВК 4992707. PMID 27528773.
- ^ Ferguson-Lees, James; Кристи, Дэвид А. (2001). Хищники мира. Лондон: Кристофер Хелм. С. 67–68. ISBN 978-0-7136-8026-3.
- ^ Tarboton, Warwick; Erasmus, Rudy (1998). Owls & Owling in Southern Africa. Кейптаун: Издательство Struik. п. 10. ISBN 1-86872-104-3.
- ^ Oberprieler, Ulrich; Cillie, Burger (2002). Raptor Identification Guide for Southern Africa. Parklands: Random House. п. 8. ISBN 978-0-9584195-7-4.
- ^ Lucas, Alfred M. (1972). Avian Anatomy - integument. East Lansing, Michigan, USA: USDA Avian Anatomy Project, Michigan State University. pp. 67, 344, 394–601.
- ^ Sawyer, R.H., Knapp, L.W. 2003. Avian Skin Development and the Evolutionary Origin of Feathers. J.Exp.Zool. (Mol.Dev.Evol) Vol.298B:57-72.
- ^ а б c Dhouailly, D. 2009. A New Scenario for the Evolutionary Origin of Hair, Feather, and Avian Scales. J.Anat. Vol.214:587-606
- ^ Чжэн, X .; Чжоу, З .; Ван, X .; Zhang, F .; Чжан, X .; Wang, Y .; Сюй, X. (2013). "Hind wings in basal birds and the evolution of leg feathers". Наука. 339 (6125): 1309–1312. Bibcode:2013Научный ... 339.1309Z. CiteSeerX 10.1.1.1031.5732. Дои:10.1126 / science.1228753. PMID 23493711. S2CID 206544531.
- ^ Stettenheim, Peter R (2000). "The Integumentary Morphology of Modern Birds—An Overview". Американский зоолог. 40 (4): 461–477. Дои:10.1093/icb/40.4.461.
- ^ Piersma, Theunis; van Aelst, Renee; Kurk, Karin; Berkhoudt, Herman; Leo R. M. Maas (1998). "A New Pressure Sensory Mechanism for Prey Detection in Birds: The Use of Principles of Seabed Dynamics?". Труды: Биологические науки.. 265 (1404): 1377–1383. Дои:10.1098/rspb.1998.0445. ЧВК 1689215.
- ^ Zusi, R L (1984). "A Functional and Evolutionary Analysis of Rhynchokinesis in Birds". Вклад Смитсоновского института в зоологию. 395. HDL:10088/5187.
- ^ Calder, William A. (1996). Size, Function, and Life History. Mineola, New York: Courier Dove Publications. п. 91. ISBN 978-0-486-69191-6.
- ^ а б c Maina, John N. (2005). The lung air sac system of birds development, structure, and function; with 6 tables. Берлин: Springer. pp. 3.2–3.3 "Lung", "Airway (Bronchiol) System" 66–82. ISBN 978-3-540-25595-6.
- ^ а б c Krautwald-Junghanns, Maria-Elisabeth; и другие. (2010). Diagnostic Imaging of Exotic Pets: Birds, Small Mammals, Reptiles. Germany: Manson Publishing. ISBN 978-3-89993-049-8.
- ^ а б c d Whittow, G. Causey (2000). Птичья физиология Стурки. Сан-Диего, Калифорния: Academic Press. стр.233 –241. ISBN 978-0-12-747605-6.
- ^ а б c d е ж грамм час я j k л м п о Ritchson, G. "BIO 554/754 – Ornithology: Avian respiration". Department of Biological Sciences, Eastern Kentucky University. Получено 2009-04-23.
- ^ а б Sturkie, P.D. (1976). Sturkie, P. D (ed.). Avian Physiology. Нью-Йорк: Springer Verlag. п. 201. Дои:10.1007/978-1-4612-4862-0. ISBN 978-1-4612-9335-4. S2CID 36415426.
- ^ а б c d Scott, Graham R. (2011). "Commentary: Elevated performance: the unique physiology of birds that fly at high altitudes". Журнал экспериментальной биологии. 214 (15): 2455–2462. Дои:10.1242/jeb.052548. PMID 21753038.
- ^ "Bird lung". Архивировано из оригинал 11 марта 2007 г.
- ^ Sinn-Hanlon, Janet. "Comparative Anatomy of the Chicken Heart". University Of Illinois.
- ^ Osborne, June (1998). The Ruby-Throated Hummingbird. Техасский университет Press. п.14. ISBN 978-0-292-76047-9.
- ^ а б Ornithology, The Cornell Lab of. "All About Bird Anatomy from Bird Academy". academy.allaboutbirds.org. Получено 2018-05-11.
- ^ а б c Zaher, Mostafa (2012). "Anatomical, histological and histochemical adaptations of the avian alimentary canal to their food habits: I-Coturnix coturnix". Life Science Journal. 9: 253–275.
- ^ Stryer, Lubert (1995). In: Biochemistry (Четвертое изд.). Нью-Йорк: W.H. Фримен и компания. С. 250–251. ISBN 0-7167-2009-4.
- ^ Moran, Edwin (2016). "Gastric digestion of protein through pancreozyme action optimizes intestinal forms for absorption, mucin formation and villus integrity". Наука и технология кормов для животных. 221: 284–303. Дои:10.1016/j.anifeedsci.2016.05.015.[постоянная мертвая ссылка ]
- ^ Svihus, Birger (2014). "Function of the digestive system". The Journal of Applied Poultry Research. 23 (2): 306–314. Дои:10.3382/japr.2014-00937.
- ^ Storer, Tracy I.; Usinger, R. L.; Стеббинс, Роберт С .; Nybakken, James W. (1997). General Zoology (шестое изд.). Нью-Йорк: Макгроу-Хилл. стр.750–751. ISBN 978-0-07-061780-3.
- ^ Tarboton, Warwick; Erasmus, Rudy (1998). Owls & Owling in Southern Africa. Кейптаун: Издательство Struik. С. 28–29. ISBN 1-86872-104-3.
- ^ Kemp, Alan; Kemp, Meg (1998). Sasol Birds of Prey of Africa and its Islands. London: New Holland Publishers (UK) Ltd. p. 332. ISBN 1-85974-100-2.
- ^ а б c Cade, Tom J. & Greenwald, Lewis I. (1966). "Drinking Behavior of Mousebirds in the Namib Desert, Southern Africa" (PDF). Аук. 83 (1).
- ^ K. Lorenz, Verhandl. Deutsch. Zool. Ges., 41 [Zool. Anz. Дополнение 12]: 69-102, 1939
- ^ Cade, Tom J.; Willoughby, Ernest J. & Maclean, Gordon L. (1966). "Drinking Behavior of Sandgrouse in the Namib and Kalahari Deserts, Africa" (PDF). Аук. 83 (1).
- ^ Gordon L. Maclean (1996) The Ecophysiology of Desert Birds. Springer. ISBN 3-540-59269-5
- ^ Elphick, Jonathan (2016). Birds: A Complete Guide to their Biology and Behavior. Буффало, Нью-Йорк: Книги Светлячка. С. 53–54. ISBN 978-1-77085-762-9.
- ^ A study of the seasonal changes in avian testes В архиве 2013-06-20 at WebCite Alexander Watson, J. Physiol. 1919;53;86-91, 'greenfinch (Carduelis chloris)', "In early summer (May and June) they are as big as a whole pea and in early winter (November) they are no bigger than a pin head"
- ^ Lake, PE (1981). "Male genital organs". In King AS, McLelland J (ed.). Form and function in birds. 2. Нью-Йорк: Академ. С. 1–61.
- ^ Kinsky, FC (1971). "The consistent presence of paired ovaries in the Kiwi(Apteryx) with some discussion of this condition in other birds". Журнал орнитологии. 112 (3): 334–357. Дои:10.1007/BF01640692. S2CID 28261057.
- ^ Fitzpatrick, FL (1934). "Unilateral and bilateral ovaries in raptorial birds" (PDF). Бюллетень Уилсона. 46 (1): 19–22.
- ^ Lynch, Wayne; Lynch, photographs by Wayne (2007). Owls of the United States and Canada : a complete guide to their biology and behavior. Балтимор: Издательство Университета Джона Хопкинса. п. 151. ISBN 978-0-8018-8687-4.
- ^ Birkhead, TR; A. P. Moller (1993). "Sexual selection and the temporal separation of reproductive events: sperm storage data from reptiles, birds and mammals". Биологический журнал Линнеевского общества. 50 (4): 295–311. Дои:10.1111/j.1095-8312.1993.tb00933.x.
- ^ Jamieson, Barrie G M (14 October 2011). Reproductive Biology and Phylogeny of Birds, Part A: Phylogeny, Morphology, Hormones and Fertilization. CRC Press. ISBN 978-1-4398-4275-1.
- ^ Herrera, A. M; S. G. Shuster; C. L. Perriton; M. J. Cohn (2013). "Developmental Basis of Phallus Reduction during Bird Evolution". Текущая биология. 23 (12): 1065–1074. Дои:10.1016/j.cub.2013.04.062. PMID 23746636.
- ^ McCracken, KG (2000). "The 20-cm Spiny Penis of the Argentine Lake Duck (Oxyura vittata)" (PDF). Аук. 117 (3): 820–825. Дои:10.1642/0004-8038(2000)117[0820:TCSPOT]2.0.CO;2.
- ^ а б Arnqvist, G.; I. Danielsson (1999). "Copulatory Behavior, Genital Morphology, and Male Fertilization Success in Water Striders". Эволюция. 53 (1): 147–156. Дои:10.2307/2640927. JSTOR 2640927. PMID 28565197.
- ^ Eberhard, W (2010). "Evolution of genitalia: theories, evidence, and new directions". Genetica. 138 (1): 5–18. Дои:10.1007/s10709-009-9358-y. PMID 19308664. S2CID 1409845.
- ^ Hosken, D.J.; P. Stockley (2004). "Sexual selection and genital evolution" (PDF). Тенденции в экологии и эволюции. 19 (2): 87–93. CiteSeerX 10.1.1.509.2660. Дои:10.1016/j.tree.2003.11.012. PMID 16701234. Архивировано из оригинал (PDF) на 2017-10-12. Получено 2018-08-26.
- ^ Brennan, P. L. R.; R. O. Prum; K. G. McCracken; M. D. Sorenson; R. E. Wilson; T. R. Birkhead (2007). "Coevolution of Male and Female Genital Morphology in Waterfowl". PLOS ONE. 2 (5): e418. Bibcode:2007PLoSO...2..418B. Дои:10.1371/journal.pone.0000418. ЧВК 1855079. PMID 17476339.
- ^ Mills, Robert (March 1994). "Applied comparative anatomy of the avian middle ear". Журнал Королевского медицинского общества. 87 (3): 155–6. ЧВК 1294398. PMID 8158595.
- ^ R., Anderson, Ted (2006-01-01). Biology of the Ubiquitous House Sparrow : From Genes to Populations. Oxford University Press, США. ISBN 9780198041351. OCLC 922954367.
- ^ Anderson, Ted (2006). Biology of the Ubiquitous House Sparrow: From Genes to Populations. Нью-Йорк: Издательство Оксфордского университета. стр.390. ISBN 978-0-19-530411-4.
- ^ Nagy, N; Magyar, A (March 1, 2001). "Development of the follicle-associated epithelium and the secretory dendritic cell in the bursa of fabricius of the guinea fowl (Numida meleagris) studied by novel monoclonal antibodies". The Anatomical Record. 262 (3): 279–292. Дои:10.1002/1097-0185(20010301)262:3<279::aid-ar1038>3.0.co;2-i. PMID 11241196.
внешняя ссылка
| Поля |  | |
|---|---|---|
| Бактерии и грибы | ||
| Протисты |
| |
| Растения | ||
| Беспозвоночные | ||
| Млекопитающие | ||
| Другие позвоночные | ||
| Глоссарии | ||
| похожие темы | ||
| ||