Дыхательная система - Respiratory system
| Дыхательная система | |
|---|---|
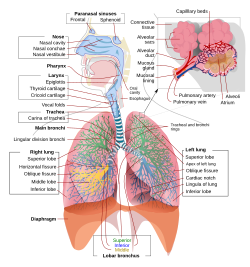 Полное схематическое изображение дыхательной системы человека с их частями и функциями. | |
| подробности | |
| Идентификаторы | |
| латинский | systema респиратор |
| MeSH | D012137 |
| TA98 | A06.0.00.000 |
| TA2 | 3133 |
| FMA | 7158 |
| Анатомическая терминология | |
В дыхательная система (также дыхательный аппарат, вентиляционная система) это биологическая система состоящий из конкретных органы и конструкции, используемые для газообмен в животные и растения. Анатомия и физиология, благодаря которым это происходит, сильно различаются в зависимости от размера организма, среды, в которой он живет, и его эволюционной истории. В наземные животные респираторная поверхность интернализируется как подкладка легкие.[1] Газообмен в легких встречается в миллионах маленьких воздушных мешочков, называемых альвеолы у млекопитающих и рептилий, но предсердие у птиц. Эти микроскопические воздушные мешочки имеют очень богатое кровоснабжение, благодаря чему воздух находится в тесном контакте с кровью.[2] Эти воздушные мешки сообщаются с внешней средой через систему дыхательных путей или полых трубок, самая большая из которых трахея, который в середине груди разветвляется на два основных бронхи. Они попадают в легкие, где разветвляются на все более узкие вторичные и третичные бронхи, которые разветвляются на множество более мелких трубок, бронхиолы. В птицы бронхиолы называются парабронхи. Это бронхиолы или парабронхи, которые обычно открываются в микроскопический альвеолы у млекопитающих и предсердие у птиц. Воздух должен закачиваться из окружающей среды в альвеолы или предсердия в процессе дыхание который включает мышцы дыхания.
В большинстве рыбы, и ряд других водные животные (и то и другое позвоночные и беспозвоночные ) дыхательная система состоит из жабры, которые частично или полностью являются внешними органами, купающимися в водной среде. Эта вода течет по жабрам различными активными или пассивными способами. Газообмен происходит в жабрах, которые состоят из тонких или очень плоских нити и Ламмелы которые открывают очень большую площадь сильно васкуляризированный ткань в воду.
Другие животные, такие как насекомые, имеют дыхательные системы с очень простыми анатомическими особенностями и амфибии даже кожа играет жизненно важную роль в газообмене. Растения также есть дыхательные системы, но направление газообмена может быть противоположным таковому у животных. Дыхательная система растений включает анатомические особенности, такие как: устьица, которые находятся в различных частях растения.[3]
Млекопитающие
Анатомия

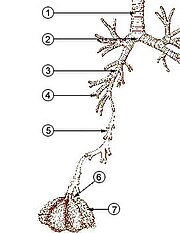
В люди и другие млекопитающие, анатомия типичной дыхательной системы - это дыхательные пути. Урочище разделено на верхний и нижние дыхательные пути. Верхний тракт включает нос, носовые полости, пазухи, глотка и часть гортань выше голосовые складки. Нижний тракт (рис. 2) включает нижнюю часть гортани, трахея, бронхи, бронхиолы и альвеолы.
Ветвящиеся дыхательные пути нижних отделов тракта часто называют дыхательное дерево или трахеобронхиальное дерево (рис. 2).[4] Интервалы между последовательными точками ветвления вдоль различных ветвей «дерева» часто называют ветвящимися «поколениями», которых у взрослого человека около 23. Ранние поколения (приблизительно поколения 0–16), состоящие из трахеи и бронхов, а также более крупных бронхиол, которые просто действуют как воздуховоды, доставляя воздух в респираторные бронхиолы, альвеолярные протоки и альвеолы (примерно поколения 17–23), где газообмен происходит.[5][6] Бронхиолы определяются как небольшие дыхательные пути без какой-либо хрящевой поддержки.[4]
Первые бронхи, ответвившиеся от трахея правый и левый главные бронхи. Вторые только по диаметру после трахеи (1,8 см), эти бронхи (1–1,4 см в диаметре)[5] введите легкие на каждом хилум, где они разветвляются на более узкие вторичные бронхи, известные как долевые бронхи, а они - на более узкие третичные бронхи, известные как сегментарные бронхи. Дальнейшие отделы сегментарных бронхов (диаметром от 1 до 6 мм)[7] известны как сегментарные бронхи 4-го, 5-го и 6-го порядка или сгруппированы вместе как субсегментарные бронхи.[8][9]
По сравнению с 23 количеством (в среднем) ветвлений дыхательного дерева у взрослого человека мышь таких разветвлений всего около 13.
Альвеолы - это тупиковые точки «дерева», а это означает, что любой воздух, попадающий в них, должен выходить по тому же маршруту. Такая система создает мертвый космос - объем воздуха (около 150 мл для взрослого человека), который заполняет дыхательные пути после выдоха и вдыхается обратно в альвеолы, прежде чем воздух из окружающей среды достигнет их.[10][11] В конце вдоха дыхательные пути заполняются воздухом окружающей среды, который выдыхается, не контактируя с газообменником.[10]
Вентиляционные объемы
Легкие расширяются и сжимаются во время дыхательного цикла, втягивая воздух в легкие и из них. Объем воздуха, входящего в легкие или выходящего из них при нормальных условиях покоя ( дыхательный объем около 500 мл), а объемы, перемещаемые во время максимально форсированного вдоха и максимально форсированного выдоха, измеряются у людей с помощью спирометрия.[12] Типичная спирограмма взрослого человека с названиями различных изменений объема, которые могут пройти легкие, проиллюстрирована ниже (рис. 3):

Не весь воздух из легких может быть удален при максимально форсированном выдохе. Это остаточный объем примерно 1,0–1,5 литра, что невозможно измерить спирометрией. Объемы, включающие остаточный объем (т.е. функциональная остаточная емкость около 2,5-3,0 литров, и общая емкость легких около 6 литров), следовательно, также нельзя измерить спирометрией. Их измерение требует специальных приемов.[12]
Скорости, с которыми воздух вдыхается или выдыхается через рот или нос, в альвеолы или выходят из них, указаны в таблице ниже вместе с тем, как они рассчитываются. Количество циклов дыхания в минуту известно как частота дыхания.
| Измерение | Уравнение | Описание |
|---|---|---|
| Минутная вентиляция | дыхательный объем * частота дыхания | общий объем воздуха, поступающего или выходящего из носа или рта в минуту. |
| Альвеолярная вентиляция | (дыхательный объем - мертвое пространство) * частота дыхания | объем воздуха, входящего или выходящего из альвеол в минуту. |
| Вентиляция мертвого пространства | мертвое пространство * частота дыхания | объем воздуха, который не достигает альвеол во время вдоха, а остается в дыхательных путях за минуту. |
Механика дыхания



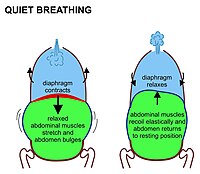

В млекопитающие вдох в покое в первую очередь связан с сокращением диафрагма. Это выступающий вверх куполообразный лист мышцы, который отделяет грудную полость от брюшной полости. Когда он сжимается, лист становится плоским (т.е. движется вниз, как показано на рис. 7), увеличивая объем грудной полости. Сокращающаяся диафрагма толкает органы брюшной полости вниз. Но поскольку тазовое дно препятствует движению самых нижних органов брюшной полости в этом направлении, податливое содержимое брюшной полости заставляет живот выпирать наружу вперед и в стороны, потому что расслабленные мышцы живота не сопротивляются этому движению (рис. 7). Это полностью пассивное выпячивание (и сжатие во время выдоха) живота при нормальном дыхании иногда называют «брюшным дыханием», хотя на самом деле это «диафрагмальное дыхание», которое не видно снаружи тела. Млекопитающие задействуют мышцы живота только во время сильного выдоха (см. Рис. 8 и обсуждение ниже). Ни при каких обстоятельствах не вдыхайте.
Когда диафрагма сжимается, грудная клетка одновременно увеличивается за счет того, что ребра тянут вверх межреберные мышцы как показано на рис. 4. Все ребра наклонены вниз от задней части к передней (как показано на рис. 4); но самые нижние ребра также наклонены вниз от средней линии наружу (рис. 5). Таким образом, поперечный диаметр грудной клетки может быть увеличен таким же образом, как и переднезадний диаметр, за счет так называемого движение рукоятки насоса показано на рис.4.
Увеличение вертикального размера грудной полости за счет сокращения диафрагмы и двух ее горизонтальных размеров за счет подъема передней и боковых сторон ребер вызывает падение внутригрудного давления. Внутренние части легких открыты для внешнего воздуха и, будучи эластичными, расширяются, заполняя увеличившееся пространство. Приток воздуха в легкие происходит через дыхательные пути (Рис. 2). В здоровье эти дыхательные пути начни с носа.[13][14] Можно начать с рта, который является резервной системой дыхания. Однако хронический дыхание через рот приводит к болезни или является ее признаком.[15][16][17] Они заканчиваются микроскопическими тупиковыми мешочками, называемыми альвеолы ) всегда открыты, хотя диаметры различных секций могут быть изменены симпатичный и парасимпатическая нервная система. Следовательно, альвеолярное давление воздуха всегда близко к атмосферному давлению воздуха (около 100кПа на уровне моря) в состоянии покоя, при этом градиенты давления, заставляющие воздух входить и выходить из легких во время дыхания, редко превышают 2–3 кПа.[18][19]
Во время выдоха расслабляются диафрагма и межреберные мышцы. Это возвращает грудную клетку и живот в положение, определяемое их анатомической эластичностью. Это «среднее положение покоя» грудной клетки и живота (рис. 7), когда легкие содержат их функциональная остаточная емкость воздуха (голубая область на рисунке справа справа), который у взрослого человека имеет объем около 2,5–3,0 литров (рис. 3).[6] Выдох в состоянии покоя длится примерно вдвое дольше, чем вдох, потому что диафрагма расслабляется более мягко, чем активно сокращается во время вдоха.

Объем воздуха, который движется в или наружу (через нос или рот) в течение одного дыхательного цикла называется дыхательный объем. У взрослого человека в состоянии покоя она составляет около 500 мл на вдох. В конце выдоха в дыхательных путях содержится около 150 мл альвеолярного воздуха, который является первым воздухом, который вдыхается обратно в альвеолы во время вдоха.[10][20] Этот объем воздуха, который выдыхается из альвеол и возвращается обратно, известен как мертвый космос вентиляция, следствием которой является то, что из 500 мл, вдыхаемых в альвеолы при каждом вдохе, только 350 мл (500 мл - 150 мл = 350 мл) свежего теплого и влажного воздуха.[6] Поскольку эти 350 мл свежего воздуха тщательно перемешиваются и разбавляются воздухом, который остается в альвеолах после нормального выдоха (т. Е. функциональная остаточная емкость примерно 2,5–3,0 л), очевидно, что состав альвеолярного воздуха очень мало изменяется во время цикла дыхания (см. рис. 9). Кислород напряжение (или парциальное давление) остается близким к 13-14 кПа (около 100 мм рт. ст.), а давление диоксида углерода очень близко к 5,3 кПа (или 40 мм рт. ст.). Это контрастирует с составом сухого наружного воздуха на уровне моря, где парциальное давление кислорода составляет 21 кПа (или 160 мм рт. Ст.), А парциальное давление углекислого газа составляет 0,04 кПа (или 0,3 мм рт. Ст.).[6]
При тяжелом дыхании (гиперпноэ ), как, например, во время упражнения вдох вызывается более мощным и большим ходом сокращающейся диафрагмы, чем в состоянии покоя (рис. 8). В дополнение "вспомогательные мышцы вдоха «преувеличивают действие межреберных мышц (рис. 8). Эти вспомогательные мышцы вдоха являются мышцами, которые отходят от шейные позвонки и основание черепа до верхних ребер и грудина, иногда через посредническую привязку к ключицы.[6] Когда они сокращаются, внутренний объем грудной клетки увеличивается в гораздо большей степени, чем может быть достигнуто сокращением одних только межреберных мышц. Если смотреть снаружи на тело, подъем ключиц во время напряженного или затрудненного вдоха иногда называют ключичное дыхание, особенно во время астма нападения и у людей с хроническая обструктивная болезнь легких.
При тяжелом дыхании выдох вызывается расслаблением всех мышц вдоха. Но теперь мышцы живота вместо того, чтобы оставаться расслабленными (как в состоянии покоя), с силой сокращаются, вытягивая нижние края грудная клетка вниз (спереди и по бокам) (рис. 8). Это не только резко уменьшает размер грудной клетки, но и толкает органы брюшной полости вверх к диафрагме, которая, следовательно, глубоко вдается в грудную клетку (рис. 8). Объем легких в конце выдоха теперь значительно ниже среднего положения в состоянии покоя и содержит гораздо меньше воздуха, чем «функциональная остаточная емкость» в состоянии покоя. Однако у нормального млекопитающего легкие не могут быть полностью опорожнены. У взрослого человека после максимального выдоха в легких всегда остается не менее 1 литра остаточного воздуха.[6]
Автоматический ритмичный вдох и выдох может прерываться кашлем, чиханием (формы очень сильного выдоха), выражением широкого спектра эмоций (смех, вздох, крик от боли, раздраженные вдохи) и т. произвольные действия в виде речи, пения, свиста и игры на духовых инструментах. Все эти действия зависят от мышц, описанных выше, и их воздействия на движение воздуха в легких и из легких.
Хотя это и не форма дыхания, Маневр Вальсальвы задействует дыхательные мышцы. На самом деле это очень сильное выдыхательное усилие против плотно закрытого голосовая щель, чтобы воздух не выходил из легких.[21] Вместо этого содержимое брюшной полости эвакуируется в противоположном направлении через отверстия в тазовом дне. Мышцы живота сокращаются очень сильно, в результате чего давление внутри живота и грудной клетки возрастает до чрезвычайно высокого уровня. Маневр Вальсальвы может выполняться добровольно, но, как правило, это рефлекс, возникающий при попытке опорожнить брюшную полость, например, во время трудной дефекации или во время родов. Во время этого маневра дыхание прекращается.
Газообмен
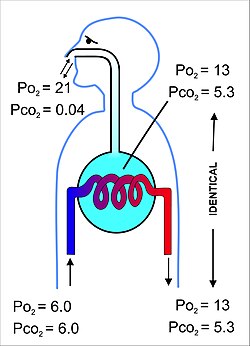


Основное назначение дыхательной системы - уравновешивание парциальных давлений дыхательных газов в альвеолярном воздухе и в легочной капиллярной крови (рис. 11). Этот процесс происходит простым распространение,[22] через очень тонкую мембрану (известную как гемато-воздушный барьер ), образующий стенки легочные альвеолы (Рис.10). Он состоит из альвеолярные эпителиальные клетки, их подвальные мембраны и эндотелиальные клетки альвеолярных капилляров (рис. 10).[23] Этот газовый барьер для крови чрезвычайно тонкий (у людей в среднем 2,2 мкм). Он сложен примерно в 300 миллионов маленьких воздушных мешочков, называемых альвеолы[23] (каждый диаметром от 75 до 300 мкм), отходящие от дыхательных путей бронхиолы в легкие, таким образом обеспечивая чрезвычайно большую площадь поверхности (примерно 145 м2) для газообмена.[23]
Воздух, содержащийся в альвеолах, имеет полупостоянный объем около 2,5-3,0 литров, который полностью окружает альвеолярную капиллярную кровь (рис. 12). Это гарантирует, что уравновешивание парциальных давлений газов в двух отсеках будет очень эффективным и происходит очень быстро. Кровь, покидающая альвеолярные капилляры и в конечном итоге распределяющаяся по телу, имеет частичное давление кислорода 13-14 кПа (100 мм рт. ст.), а парциальное давление диоксида углерода 5,3 кПа (40 мм рт. ст.) (то есть такое же, как напряжения кислорода и углекислого газа, как в альвеолах).[6] Как упоминалось в раздел выше соответствующие парциальные давления кислорода и углекислого газа в окружающем (сухом) воздухе на уровне моря составляют 21 кПа (160 мм рт. ст.) и 0,04 кПа (0,3 мм рт. ст.) соответственно.[6]
Эта заметная разница между составом альвеолярного воздуха и окружающего воздуха может сохраняться, поскольку функциональная остаточная емкость содержится в тупиковых мешочках, связанных с наружным воздухом довольно узкими и относительно длинными трубками (дыхательные пути: нос, глотка, гортань, трахея, бронхи и их ветви вниз к бронхиолы ), через которую воздух должен вдыхаться и выдыхаться (т.е. нет однонаправленного сквозного потока, как в легкое птицы ). Эта типичная анатомия млекопитающих в сочетании с тем фактом, что легкие не опорожняются и повторно надуваются при каждом вдохе (оставляя значительный объем воздуха, примерно 2,5-3,0 литра, в альвеолах после выдоха), гарантирует, что состав альвеол Воздух почти не нарушается, если при каждом вдохе подмешивать 350 мл свежего воздуха. Таким образом, животному предоставляется совершенно особая «портативная атмосфера», состав которой существенно отличается от современный атмосферный воздух.[24] Именно эта портативная атмосфера ( функциональная остаточная емкость ), которому подвергается кровь и, следовательно, ткани тела, а не наружный воздух.
Результирующие артериальные парциальные давления кислорода и углекислого газа равны гомеостатически управляемый. Повышение артериального парциального давления CO2 и, в меньшей степени, падение артериального парциального давления O2, рефлекторно вызовет более глубокое и быстрое дыхание, пока напряжение газов крови в легких и, следовательно, в артериальной крови нормализуется. Обратное происходит, когда давление углекислого газа падает или, опять же, в меньшей степени, повышается давление кислорода: частота и глубина дыхания снижаются до тех пор, пока не восстанавливается нормальность газов крови.
Поскольку кровь, поступающая в альвеолярные капилляры, имеет парциальное давление O2 в среднем 6 кПа (45 мм рт. ст.), в то время как давление в альвеолярном воздухе составляет 13-14 кПа (100 мм рт. ст.), будет чистая диффузия кислорода в капиллярную кровь, изменяющую состав 3 литров альвеолярный воздух слегка. Точно так же, поскольку кровь, поступающая в альвеолярные капилляры, имеет парциальное давление CO2 также около 6 кПа (45 мм рт. ст.), в то время как альвеолярный воздух составляет 5,3 кПа (40 мм рт. ст.), происходит чистое перемещение углекислого газа из капилляров в альвеолы. Изменения, вызываемые этими чистыми потоками отдельных газов в альвеолярный воздух и из него, требуют замены примерно 15% альвеолярного воздуха окружающим воздухом каждые 5 секунд или около того. Это очень жестко контролируется контролем газов артериальной крови (которые точно отражают состав альвеолярного воздуха) с помощью аортальный и каротидные тела, а также датчик газов крови и pH на передней поверхности продолговатый мозг в мозгу. В легких также есть датчики кислорода и углекислого газа, но они в первую очередь определяют диаметр бронхиолы и легочные капилляры, и поэтому несут ответственность за направление потока воздуха и крови к разным частям легких.
Только в результате точного поддержания состава 3 литров альвеолярного воздуха с каждым вдохом некоторое количество углекислого газа выбрасывается в атмосферу, а некоторое количество кислорода забирается из внешнего воздуха. Если за короткий период времени было потеряно больше углекислого газа, чем обычно. гипервентиляция дыхание будет замедлено или остановлено до тех пор, пока альвеолярное парциальное давление углекислого газа не вернется к 5,3 кПа (40 мм рт. ст.). Поэтому, строго говоря, неверно, что основная функция дыхательной системы - избавлять организм от «отходов» углекислого газа. Углекислый газ, который выдыхается при каждом вдохе, вероятно, правильнее рассматривать как побочный продукт внеклеточной жидкости организма. углекислый газ и pH-гомеостаты
Если эти гомеостаты скомпрометированы, тогда респираторный ацидоз, или респираторный алкалоз произойдет. В конечном итоге это может быть компенсировано почечной коррекцией ЧАС+ и HCO3− концентрации в плазме; но поскольку это требует времени, синдром гипервентиляции может, например, возникать, когда возбуждение или тревога заставляют человека дышать быстро и глубоко, вызывая таким образом неприятные ощущения. респираторный алкалоз через продувку слишком большого количества CO2 из крови в наружный воздух.[25]
Кислород имеет очень низкую растворимость в воде и поэтому переносится в крови в слабом сочетании с гемоглобин. Кислород удерживается в гемоглобине четырьмя двухвалентное железо -содержащий гем групп на молекулу гемоглобина. Когда все группы гема содержат один O2 Каждая молекула крови считается «насыщенной» кислородом, и никакое дальнейшее повышение парциального давления кислорода не приведет к значительному увеличению концентрации кислорода в крови. Большая часть углекислого газа в крови переносится в виде ионов бикарбоната (HCO3−) в плазме. Однако конверсия растворенного CO2 в HCO3− (за счет добавления воды) слишком медленна для скорости, с которой кровь циркулирует через ткани, с одной стороны, и через альвеолярные капилляры, с другой. Следовательно, реакция катализируется карбоангидраза, фермент внутри красные кровяные клетки.[26] Реакция может идти в обоих направлениях в зависимости от преобладающего парциального давления CO.2.[6] Небольшое количество диоксида углерода переносится белковой частью молекул гемоглобина в виде карбамино группы. Общая концентрация углекислого газа (в виде ионов бикарбоната, растворенного CO2, и карбаминогруппы) в артериальной крови (т.е. после того, как она уравновесится с альвеолярным воздухом) составляет около 26 мМ (или 58 мл / 100 мл),[27] по сравнению с концентрацией кислорода в насыщенной артериальной крови около 9 мМ (или 20 мл / 100 мл крови).[6]
Контроль вентиляции
Вентиляция легких у млекопитающих происходит через дыхательные центры в продолговатый мозг и мосты из мозговой ствол.[6] Эти области образуют серию нервные пути которые получают информацию о парциальные давления кислорода и углекислого газа в артериальная кровь. Эта информация определяет среднюю скорость вентиляции альвеолы из легкие, чтобы сохранить эти постоянное давление. Дыхательный центр делает это через двигательные нервы которые активируют диафрагма и другие мышцы дыхания.
Частота дыхания увеличивается, когда парциальное давление диоксида углерода в крови увеличивается. Это обнаруживается центральные хеморецепторы газов крови на передней поверхности продолговатый мозг.[6] В аортальный и каротидные тела, являются хеморецепторы газов периферической крови которые особенно чувствительны к артериальному парциальное давление O2 хотя они тоже реагируют, но менее сильно, на парциальное давление CO2.[6] На уровне моря в нормальных условиях частота и глубина дыхания определяются в первую очередь парциальным давлением углекислого газа в артериальной крови, а не артериальным давлением. парциальное давление кислорода, который может изменяться в довольно широком диапазоне, прежде чем дыхательные центры в продолговатом мозге и мосте отреагируют на него, изменив частоту и глубину дыхания.[6]
Упражнение увеличивает частоту дыхания за счет дополнительного количества углекислого газа, образующегося в результате ускоренного метаболизма тренирующихся мышц.[28] Кроме того, пассивные движения конечностей также рефлекторно увеличивают частоту дыхания.[6][28]
Информация получена от рецепторы растяжения в пределах легких дыхательный объем (глубина вдоха и выдоха).
Реакция на низкое атмосферное давление
В альвеолы открыты (через дыхательные пути) для атмосферы, в результате чего альвеолярное давление воздуха точно такое же, как давление окружающего воздуха на уровне моря, на высоте или в любой искусственной атмосфере (например, в водолазной камере или декомпрессионной камере) в которым человек дышит свободно. С участием расширение легких альвеолярный воздух занимает больший объем, а его давление падает пропорционально, заставляя воздух проходить через дыхательные пути, пока давление в альвеолах снова не станет равным давлению окружающего воздуха. Во время выдоха происходит обратное. Эта обработать (вдоха и выдоха) точно так же на уровне моря, как и на вершине Mt. Эверест, или в камера для дайвинга или декомпрессионная камера.
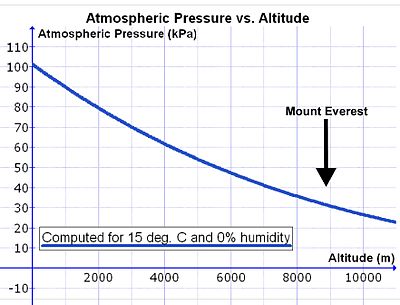
Однако по мере того, как человек поднимается над уровнем моря, плотность воздуха уменьшается экспоненциально (см. рис. 14), уменьшив примерно вдвое с каждым подъемом на 5500 м..[29] Поскольку состав атмосферного воздуха на глубине ниже 80 км практически постоянен, в результате постоянного перемешивающего воздействия погоды концентрация кислорода в воздухе (ммоль O2 на литр окружающего воздуха) уменьшается с той же скоростью, что и падение атмосферного давления с высотой.[30] Следовательно, чтобы вдыхать такое же количество кислорода в минуту, человек должен вдыхать пропорционально больший объем воздуха в минуту на высоте, чем на уровне моря. Это достигается за счет более глубокого и быстрого дыхания (т.е. гиперпноэ ), чем на уровне моря (см. ниже).

Однако есть осложнение, которое увеличивает объем воздуха, который необходимо вдыхать в минуту (минутный объем дыхания ), чтобы обеспечить легким такое же количество кислорода на высоте, как и на уровне моря. При вдохе воздух нагревается и насыщается водяным паром при прохождении через носовые ходы и глотка. Давление насыщенного водяного пара зависит только от температуры. При внутренней температуре тела 37 ° C это 6,3.кПа (47,0 мм рт. Ст.), Независимо от любых других факторов, включая высоту.[31] Таким образом, на уровне моря, где окружающее атмосферное давление составляет около 100 кПа, влажный воздух, поступающий в легкие из трахея состоит из водяного пара (6,3 кПа), азота (74,0 кПа), кислорода (19,7 кПа) и следовых количеств диоксида углерода и других газов (всего 100 кПа). В сухом воздухе частичное давление из O2 на уровне моря составляет 21,0 кПа (т.е. 21% от 100 кПа) по сравнению с 19,7 кПа кислорода, поступающего в альвеолярный воздух. (Парциальное давление кислорода в трахее составляет 21% от [100 кПа - 6,3 кПа] = 19,7 кПа). На вершине Mt. Эверест (на высоте 8 848 м или 29 029 футов) общая атмосферное давление 33,7 кПа, из которых 7,1 кПа (или 21%) приходится на кислород.[29] Воздух, попадающий в легкие, также имеет общее давление 33,7 кПа, из которых 6,3 кПа неизбежно составляют водяной пар (как на уровне моря). Это снижает парциальное давление кислорода, поступающего в альвеолы, до 5,8 кПа (или 21% от [33,7 кПа - 6,3 кПа] = 5,8 кПа). Таким образом, снижение парциального давления кислорода во вдыхаемом воздухе существенно больше, чем можно было бы предположить по снижению общего атмосферного давления на высоте (на горе Эверест: 5,8 кПа vs. 7,1 кПа).
Еще одна небольшая сложность существует на высоте. Если бы объем легких мгновенно увеличился вдвое в начале вдоха, давление воздуха внутри легких уменьшилось бы вдвое. Это происходит независимо от высоты. Таким образом, уменьшение вдвое давления воздуха на уровне моря (100 кПа) приводит к внутрилегочному давлению воздуха 50 кПа. То же самое на высоте 5500 м, где атмосферное давление всего 50 кПа, внутрилегочное давление воздуха падает до 25 кПа. Таким образом, такое же изменение объема легких на уровне моря приводит к разнице давления между окружающим воздухом и внутрилегочным воздухом на 50 кПа, тогда как на высоте 5500 м разница составляет всего 25 кПа. Таким образом, на этой высоте движущее давление, заставляющее воздух поступать в легкие во время вдоха, уменьшается вдвое. В показатель приток воздуха в легкие при вдохе на уровне моря вдвое больше, чем на высоте 5500 м. Однако в действительности вдох и выдох происходят гораздо мягче и менее резко, чем в приведенном примере. Разница между атмосферным и внутрилегочным давлением, при котором воздух поступает в легкие и выходит из них во время дыхательного цикла, составляет всего 2–3 кПа.[18][19] Удвоение или больше этих небольших перепадов давления может быть достигнуто только за счет очень значительных изменений дыхательного усилия на больших высотах.
Все вышеупомянутые влияния низкого атмосферного давления на дыхание компенсируются в первую очередь более глубоким и быстрым дыханием (гиперпноэ ). Точная степень гиперпноэ определяется по гомеостат газов крови, который регулирует парциальные давления кислорода и углекислого газа в артериальной крови. Эта гомеостат уделяет приоритетное внимание регуляции артериальной частичное давление диоксида углерода над кислородом на уровне моря.[6] Иными словами, на уровне моря парциальное артериальное давление CO2 поддерживается на уровне, очень близком к 5,3 кПа (или 40 мм рт. ст.) в широком диапазоне обстоятельств, за счет артериального парциального давления O2, который может изменяться в очень широком диапазоне значений, прежде чем вызвать корректирующую респираторную реакцию. Однако, когда атмосферное давление (и, следовательно, парциальное давление O2 в атмосферном воздухе) падает ниже 50-75% от его значения на уровне моря, кислород гомеостаз имеет приоритет над гомеостазом углекислого газа.[6] Это переключение происходит на высоте около 2500 м (или около 8000 футов). Если это переключение происходит относительно внезапно, гиперпноэ на большой высоте вызовет резкое падение артериального парциального давления углекислого газа с последующее повышение pH артериальной плазмы. Это один из авторов высокогорная болезнь. С другой стороны, если переключение на кислородный гомеостаз неполное, то гипоксия может осложнить клиническую картину с потенциально летальным исходом.
В меньшей бронхи и бронхиолы. В ответ на низкое парциальное давление кислорода во вдыхаемом воздухе эти датчики рефлекторно вызывают сужение легочных артериол.[32] (Это полная противоположность соответствующего рефлекса в тканях, где низкое артериальное парциальное давление O2 вызывают расширение артериолярных сосудов.) На высоте это вызывает повышение легочного артериального давления в результате кровоток в легких распределяется гораздо более равномерно, чем на уровне моря. На уровне моря давление в легочной артерии очень низкое, в результате чего верхушки легких получают гораздо меньше крови, чем основания, которые относительно перенасыщены кровью. Только в середине легких приток крови и воздуха к альвеолам идеально согласован. На высоте это изменение соотношение вентиляция / перфузия альвеол от верхушки легких до низа удаляется, при этом все альвеолы перфузируются и вентилируются более или менее физиологически идеальным образом. Это еще один важный фактор акклиматизация к большим высотам и низкое давление кислорода.
Почки измеряют кислород содержание (ммоль O2/ литр крови, а не парциальное давление O2) артериальной крови. Когда содержание кислорода в крови хронически низкое, например на большой высоте, чувствительные к кислороду клетки почек выделяют эритропоэтин (ЭПО) в кровь.[33][34] Этот гормон стимулирует красный костный мозг для увеличения скорости производства эритроцитов, что приводит к увеличению гематокрит крови и, как следствие, увеличение ее способности переносить кислород (из-за ныне высокого гемоглобин содержание крови). Другими словами, при том же парциальном давлении O2, человек с высоким гематокритом переносит больше кислорода на литр крови, чем человек с более низким гематокритом. Следовательно, у жителей высокогорья гематокрит выше, чем у жителей, живущих на уровне моря.[34][35]
Другие функции легких
Местная оборона
Раздражение нервных окончаний внутри носовые проходы или дыхательные пути, может вызвать кашлевой рефлекс и чихание. Эти реакции вызывают насильственное вытеснение воздуха из трахея или нос соответственно. Таким образом, раздражители попадают в слизь какие линии дыхательных путей выводятся или перемещаются в рот где они могут быть проглотил.[6] Во время кашля сокращение гладких мышц стенок дыхательных путей сужает трахею, стягивая концы хрящевых пластин вместе и проталкивая мягкие ткани в просвет. Это увеличивает скорость выдыхаемого воздуха, чтобы вытеснить и удалить любые раздражающие частицы или слизь.
Респираторный эпителий может выделять различные молекулы, которые помогают защитить легкие. К ним относятся секреторные иммуноглобулины (IgA), Colleins, дефенсины и другие пептиды и протеазы, активные формы кислорода, и активные формы азота. Эти выделения могут действовать как противомикробные средства, защищая дыхательные пути от инфекции. Разнообразие хемокины и цитокины также секретируются, которые привлекают традиционные иммунные клетки и другие клетки к очагу инфекции.
Поверхностно-активное вещество Иммунная функция в первую очередь связана с двумя белками: SP-A и SP-D. Эти белки могут связываться с сахарами на поверхности патогенов и тем самым опсонизировать их для поглощения фагоцитами. Он также регулирует воспалительные реакции и взаимодействует с адаптивным иммунным ответом. Разложение или инактивация поверхностно-активного вещества может способствовать повышению восприимчивости к воспалению и инфекции легких.[36]
Большая часть дыхательной системы выстлана слизистыми оболочками, содержащими лимфоидная ткань, связанная со слизистой оболочкой, который производит белые кровяные клетки такие как лимфоциты.
Профилактика альвеолярного коллапса
Легкие производят поверхностно-активное вещество, поверхностно-активный липопротеин комплекс (фосфолипопротеин), образованный альвеолярные клетки II типа. Он плавает на поверхности тонкого водянистого слоя, выстилающего внутреннюю поверхность альвеол, снижая поверхностное натяжение воды.
Поверхностное натяжение водной поверхности (граница раздела вода-воздух) приводит к усадке этой поверхности.[6] Когда эта поверхность изогнута, как в альвеолах легких, усадка поверхности уменьшает диаметр альвеол. Чем острее кривизна границы раздела вода-воздух тем больше тенденция к разрушению альвеол.[6] Это имеет три эффекта. Во-первых, поверхностное натяжение внутри альвеол препятствует расширению альвеол во время вдоха (то есть делает легкие жесткими или неподатливыми). Поверхностно-активное вещество снижает поверхностное натяжение и, следовательно, улучшает легкие. послушный или менее жесткий, чем если бы его не было. Во-вторых, диаметры альвеол увеличиваются и уменьшаются во время дыхательного цикла. Это означает, что альвеолы имеют большая склонность к коллапсу (т.е. причина ателектаз ) в конце выдоха, что в конце вдоха. Поскольку поверхностно-активное вещество плавает на водной поверхности, его молекулы более плотно упакованы вместе, когда альвеолы сжимаются во время выдоха.[6] Это заставляет их иметь больший эффект снижения поверхностного натяжения, когда альвеолы маленькие, чем когда они большие (как в конце вдоха, когда молекулы поверхностно-активного вещества расположены более широко). Поэтому тенденция к сжатию альвеол в конце выдоха почти такая же, как и в конце вдоха. В-третьих, поверхностное натяжение изогнутого водянистого слоя, выстилающего альвеолы, имеет тенденцию втягивать воду из тканей легких в альвеолы. Поверхностно-активное вещество снижает эту опасность до незначительного уровня и сохраняет альвеолы сухими.[6][37]
Недоношенные дети у тех, кто не может производить сурфактант, есть легкие, которые имеют тенденцию разрушаться при каждом выдохе. Если не лечить, это состояние называется респираторный дистресс-синдром, смертельно. Основные научные эксперименты, проведенные с использованием клеток из легких цыплят, подтверждают возможность использования стероиды как средство дальнейшего развития альвеолярных клеток II типа.[38] Фактически, однажды преждевременные роды находится под угрозой, прилагаются все усилия, чтобы отсрочить роды, и ряд стероидный препарат Во время этой задержки матери часто делают инъекции, чтобы способствовать созреванию легких.[39]
Вклад в функции всего тела
Сосуды легких содержат фибринолитическая система что растворяется сгустки которые могли попасть в малый круг кровообращения эмболия, часто из-за глубоких вен ног. Они также выделяют различные вещества, которые попадают в системную артериальную кровь, и удаляют другие вещества из системной венозной крови, которые достигают их через легочную артерию. Немного простагландины выводятся из кровотока, а другие синтезируются в легких и попадают в кровь при растяжении легочной ткани.
Легкие активируют один гормон. Физиологически неактивный декапептид ангиотензин I преобразуется в альдостерон -выделяющий октапептид, ангиотензин II, в малом круге кровообращения. Реакция возникает и в других тканях, но особенно заметна в легких. Ангиотензин II также оказывает прямое влияние на стенки артериол, вызывая артериолярный вазоконстрикция, и, как следствие, рост артериальное давление.[40] Большое количество ангиотензин-превращающий фермент ответственные за эту активацию расположены на поверхностях эндотелиальные клетки альвеолярных капилляров. Конвертирующий фермент также инактивирует брадикинин. Время циркуляции через альвеолярные капилляры составляет менее одной секунды, но 70% ангиотензина I, достигающего легких, превращается в ангиотензин II за один проход через капилляры. Четыре других пептидазы были идентифицированы на поверхности легочных эндотелиальных клеток.
Вокализация
Движение газа через гортань, глотка и рот позволяет людям разговаривать, или фонат. Вокализация или пение у птиц происходит через сиринкс, орган, расположенный в основании трахеи. Вибрация воздуха, проходящего через гортань (голосовые связки ) у людей и сиринкс у птиц вызывает звук. Из-за этого движение газа жизненно важно для общение целей.
Контроль температуры
Задыхаясь у собак, кошек, птиц и некоторых других животных обеспечивает средство для снижения температуры тела за счет испарения слюны во рту (вместо испарения пота на коже).
Клиническое значение
Заболевания дыхательной системы можно разделить на несколько общих групп:
- Обструктивные состояния дыхательных путей (например, эмфизема, бронхит, астма )
- Легочные ограничительные состояния (например, фиброз, саркоидоз, альвеолярное повреждение, плевральный выпот )
- Сосудистые заболевания (например, отек легких, легочная эмболия, легочная гипертензия )
- Инфекционные, экологические и другие «болезни» (например, пневмония, туберкулез, асбестоз, твердые загрязнители )
- Первичный рак (например, бронхиальная карцинома, мезотелиома )
- Вторичный рак (например, рак, который возник в других частях тела, но посеял себя в легких)
- Недостаточно поверхностно-активного вещества (например, респираторный дистресс-синдром у недоношенных детей).
Заболевания дыхательной системы обычно лечатся пульмонолог и респираторный терапевт.
Если есть неспособность дышать или недостаточное дыхание аппарат ИВЛ может быть использовано.
Исключительные млекопитающие
Лошади
Лошади облигатные носовые дыхатели Это означает, что они отличаются от многих других млекопитающих, потому что у них нет возможности дышать ртом, и они должны вдыхать воздух через нос.
Слонов
В слон это единственное известное млекопитающее, у которого нет плевральная полость. Скорее теменный и висцеральная плевра оба состоят из плотных соединительная ткань и соединены друг с другом через рыхлую соединительную ткань.[41] Это отсутствие плевральной полости наряду с необычно толстым диафрагма, считаются эволюционные адаптации позволяя слону оставаться под водой в течение длительного периода времени, дыша через ствол который появляется как трубка.[42]
У слона легкие прикреплены к диафрагме, и дыхание зависит в основном от диафрагмы, а не от расширения грудной клетки.[43]
Птицы



Ключ:
1. череп; 2. шейные позвонки; 3. фуркула; 4. коракоидный; 5. позвоночные ребра; 6. грудина и ее киль; 7. надколенник; 8. предплюсна; 9. цифры; 10. большеберцовая кость (большеберцовая кость ); 11. малоберцовая кость (большеберцовая кость ); 12. бедренная кость; 13. седалищная кость (безымянный ); 14. лобок (безымянный); 15. подвздошная кость (безымянный); 16. хвостовые позвонки; 17. пигостиль; 18. синсакрум; 19. лопатка; 20. спинные позвонки; 21. плечевая кость; 22. локтевая кость; 23. радиус; 24. запястье (запястный сустав ); 25. пясть (запястный сустав ); 26. цифры; 27. Alula

Дыхательная система птиц значительно отличается от таковой у млекопитающих. Во-первых, у них жесткие легкие, которые не расширяются и не сжимаются во время дыхательного цикла. Вместо обширной системы воздушные мешки (Рис. 15), распределенные по их телам, действуют как сильфоны, втягивающие воздух из окружающей среды в мешочки и вытесняющие отработанный воздух после того, как он прошел через легкие (Рис. 18).[44] Птицы тоже не имеют диафрагмы или плевральные полости.
Легкие птиц меньше, чем у млекопитающих сопоставимого размера, но воздушные мешки составляют 15% от общего объема тела, по сравнению с 7%, отведенными на легкие. альвеолы которые действуют как мехи у млекопитающих.[45]
Вдох и выдох вызываются попеременным увеличением и уменьшением объема всей торако-брюшной полости (или целомудрие ) с использованием как брюшных, так и реберных мышц.[46][47][48] Во время вдоха мышцы, прикрепленные к ребрам позвонков (рис. 17), сокращаются, наклоняя их вперед и наружу. Это толкает грудные ребра, к которым они прикреплены почти под прямым углом, вниз и вперед, грудина (с его выдающимся киль ) в том же направлении (рис. 17). Это увеличивает как вертикальный, так и поперечный диаметр грудной части туловища. Движение вперед и вниз, в частности, задний конец грудины тянет брюшную стенку вниз, увеличивая объем этой области туловища.[46] Увеличение объема всей полости туловища снижает давление воздуха во всех торако-брюшных воздушных мешках, заставляя их заполняться воздухом, как описано ниже.
На выдохе внешняя косая мышца, прикрепляющаяся к грудине и позвоночным ребрам. спереди, и к тазу (лобковая и подвздошная кость на рис.17) сзади (составляющая часть брюшной стенки) меняет направление вдоха, одновременно сжимая содержимое брюшной полости, увеличивая, таким образом, давление во всех воздушных мешочках. Таким образом, во время выдоха воздух удаляется из дыхательной системы.[46]

При вдохе воздух попадает в трахея через ноздри и рот и продолжает сиринкс в этот момент трахея разветвляется на две главные бронхи, идущий к двум легким (рис. 16). Первичные бронхи входят в легкие, чтобы стать внутрилегочными бронхами, которые выделяют набор параллельных ветвей, называемых вентробронхами, а чуть дальше - эквивалентный набор дорсобронхов (рис. 16).[46] Концы внутрилегочных бронхов выпускают воздух в задние воздушные мешки на каудальный конец птицы. Каждая пара дорсо-вентробронхов связана большим количеством параллельных микроскопических воздушных капилляров (или парабронхи ) где газообмен происходит (рис. 16).[46] Когда птица вдыхает, трахейный воздух течет через внутрилегочные бронхи в задние воздушные мешки, а также в дорсобронхи, но не в Вентробронхи (рис.18). Это связано с архитектурой бронхов, которая направляет вдыхаемый воздух от отверстий вентробронхов в продолжение внутрилегочного бронха к дорсобронхам и задним воздушным мешочкам.[50][51][52] Из дорсобронхов вдыхаемый воздух проходит через парабронхи (и, следовательно, через газообменник) к вентробронкам, откуда воздух может выходить только в расширяющиеся передние воздушные мешочки. Итак, во время вдоха расширяются и задний, и передний воздушные мешки,[46] задние воздушные мешки заполняются свежим вдыхаемым воздухом, а передние воздушные мешочки - «отработанным» (бедным кислородом) воздухом, только что прошедшим через легкие.
Во время выдоха давление в задних воздушных мешочках (которые были заполнены свежим воздухом во время вдоха) увеличивается из-за описанного выше сокращения косой мышцы. Аэродинамика соединительных отверстий от задних воздушных мешков до дорсобронхов и внутрилегочных бронхов гарантирует, что воздух покидает эти мешочки в направлении легких (через дорсобронхи), а не возвращается вниз по внутрилегочным бронхам (рис. 18).[50][52] Из дорсобронхов свежий воздух из задних воздушных мешочков проходит через парабронхи (в том же направлении, что и при вдохе) в вентробронхи. Дыхательные пути, соединяющие вентробронхи и передние воздушные мешки с внутрилегочными бронхами, направляют «отработанный», бедный кислородом воздух из этих двух органов в трахею, откуда он выходит наружу.[46] Таким образом, насыщенный кислородом воздух постоянно (в течение всего цикла дыхания) проходит через парабронхи в одном направлении.[53]
Кровоток через легкие птицы направлен под прямым углом к потоку воздуха через парабронхи, образуя перекрестно-проточную систему обмена (рис. 19).[44][46][49] В парциальное давление кислорода в парабронках уменьшается по длине как O2 проникает в кровь. Кровеносные капилляры, выходящие из теплообменника около входа воздушного потока, потребляют больше O2 чем капилляры, уходящие около выходного конца парабронхов. Когда содержимое всех капилляров смешивается, конечное парциальное давление кислорода смешанной легочной венозной крови выше, чем у выдыхаемого воздуха,[46][49] но тем не менее составляет менее половины вдыхаемого воздуха,[46] таким образом достигается примерно такое же системное парциальное давление кислорода в артериальной крови, что и млекопитающие делают с их легкими типа мехов.[46]
Трахея - это область мертвый космос: воздух с низким содержанием кислорода, который он содержит в конце выдоха, является первым воздухом, который повторно входит в задние воздушные мешочки и легкие. По сравнению с дыхательные пути млекопитающих объем мертвого пространства у птицы в среднем в 4,5 раза больше, чем у млекопитающих того же размера.[45][46] Птицы с длинной шеей неизбежно будут иметь длинные трахеи, и поэтому должны делать более глубокий вдох, чем млекопитающие, чтобы сделать поправку на больший объем мертвого пространства. У некоторых птиц (например, лебедь-кликун, Лебедь Cygnus, то белая колпица, Platalea leucorodia, то Американский журавль, Grus americana, а в шлеме curassow, Pauxi pauxi) трахея, длина которой у некоторых кранов может достигать 1,5 м,[46] скручивается взад и вперед внутри тела, резко увеличивая вентиляцию мертвого пространства.[46] Назначение этой необычной особенности неизвестно.
Рептилии

В анатомическая структура из легкие менее сложен в рептилии чем в млекопитающие, у рептилий отсутствует очень обширная древовидная структура дыхательных путей, обнаруженная в легких млекопитающих. Газообмен у рептилий все еще встречается в альвеолы Однако.[44] Рептилии не обладают диафрагма. Таким образом, дыхание происходит за счет изменения объема полости тела, которое контролируется сокращением межреберные мышцы у всех рептилий кроме черепахи. У черепах определяется сокращение определенных пар боковых мышц. вдыхание и выдох.[54]
Амфибии
И легкие, и кожа служат в качестве органов дыхания в амфибии. Вентиляция легких у амфибий зависит от вентиляция с положительным давлением. Мышцы опускают дно полости рта, увеличивая его и втягивая воздух через ноздри в полость рта. ротовая полость. Когда ноздри и рот закрыты, дно ротовой полости выталкивается вверх, в результате чего воздух по трахее попадает в легкие. Кожа этих животных сильно васкуляризована и влажная, влага поддерживается за счет секреции слизь из специализированных клеток, и участвует в кожное дыхание. Хотя легкие являются основными органами газообмена между кровью и окружающим воздухом (когда они находятся вне воды), уникальные свойства кожи способствуют быстрому газообмену, когда земноводные погружаются в воду, богатую кислородом.[55]У некоторых амфибий есть жабры либо на ранних стадиях своего развития (например, головастики из лягушки ), в то время как другие сохраняют их в зрелом возрасте (например, некоторые саламандры ).[44]
Рыбы


Кислород плохо растворяется в воде. Полностью вентилируемый пресная вода поэтому содержит только 8–10 мл O2/ литр по сравнению с O2 концентрация 210 мл / л в воздухе на уровне моря.[59] Кроме того, коэффициент диффузии (то есть скорость, с которой вещество диффундирует из области с высокой концентрацией в область с низкой концентрацией при стандартных условиях) дыхательных газов равна обычно на 10 000 быстрее в воздухе, чем в воде.[59] Так, например, кислород имеет коэффициент диффузии 17,6 мм.2/ с в воздухе, но всего 0,0021 мм2/ с в воде.[60][61][62][63] Соответствующие значения для диоксида углерода 16 мм.2/ с в воздухе и 0,0016 мм2/ с в воде.[62][63] Это означает, что когда кислород забирается из воды, контактирующей с газообменником, он значительно медленнее замещается кислородом из богатых кислородом областей, расположенных на небольшом расстоянии от теплообменника, чем это могло бы происходить в воздухе. Рыбы развились жабры решать эти проблемы. Жабры - это специализированные органы, содержащие нити, которые далее делятся на ламели. Ламели содержат плотный тонкостенная капиллярная сеть что подвергает большую площадь поверхности газообмена очень большим объемам воды, проходящей через них.[64]
Жабры используют встречный обмен система, повышающая эффективность поглощения кислорода из воды.[56][57][58] Свежая насыщенная кислородом вода, попадающая через рот, непрерывно «прокачивается» через жабры в одном направлении, в то время как кровь в ламеллах течет в противоположном направлении, создавая противоток крови и воды (рис. 22), на котором рыба выживает. зависит.[58]
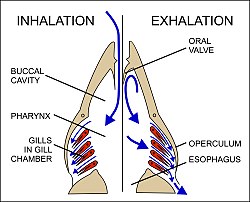
Вода всасывается через рот, закрывая крышка (жаберная крышка) и увеличение ротовой полости (рис. 23). Одновременно с этим расширяются жаберные камеры, создавая там более низкое давление, чем во рту, заставляя воду течь по жабрам.[58] Затем ротовая полость сжимается, вызывая закрытие пассивных оральных клапанов, тем самым предотвращая обратный поток воды изо рта (рис. 23).[58][65] Вместо этого вода во рту вытесняется через жабры, в то время как жаберные камеры сокращаются, опорожняя содержащуюся в них воду через глазничные отверстия (рис. 23). Обратный поток в жаберную камеру во время фазы вдоха предотвращается мембраной вдоль вентро-задний граница жаберной крышки (диаграмма слева на рис. 23). Таким образом, ротовая полость и жаберные камеры действуют попеременно как всасывающий насос и нагнетательный насос, чтобы поддерживать постоянный поток воды через жабры в одном направлении.[58] Поскольку кровь в ламеллярных капиллярах течет в направлении, противоположном направлению воды, последующее противоток поток крови и воды поддерживает резкие градиенты концентрации кислорода и углекислого газа по всей длине каждого капилляра (нижняя диаграмма на рис. 22). Таким образом, кислород может непрерывно диффундировать вниз по градиенту в кровь, а углекислый газ - вниз по градиенту в воду.[57] Хотя противоточные системы обмена теоретически позволяют почти полностью переносить респираторный газ с одной стороны теплообменника на другую, у рыб обычно менее 80% кислорода в воде, протекающей по жабрам, передается в кровь.[56]
В некоторых активных пелагический У акул вода проходит через рот и жабры во время движения в процессе, известном как «вентиляция барана».[66] В состоянии покоя большинство акул, как и большинство костистых рыб, перекачивают воду через жабры, чтобы насыщенная кислородом вода продолжала течь по их жабрам. Но небольшое количество видов потеряли способность качать воду через жабры и вынуждены плавать без отдыха. Эти виды облигатные вытяжные вентиляторы и предположительно задыхаться если не может двигаться. Облигатная вентиляция барана также характерна для некоторых пелагических костных видов рыб.[67]
Есть несколько рыб, которые могут получать кислород в течение коротких периодов времени из воздуха, проглоченного над поверхностью воды. Таким образом Двоякодышащая рыба обладают одним или двумя легкими, а лабиринт рыбы разработали особый «лабиринтный орган», который характеризует этот подотряд рыб. Орган лабиринта представляет собой сильно сложенную надстройку.жаберный аксессуар орган дыхания. Он образован васкуляризированный расширение наджаберной кости первой жаберной дуги и используется для дыхание в воздухе.[68]
Этот орган позволяет лабиринтной рыбе ловить кислород непосредственно из воздуха, вместо того, чтобы брать его из воды, в которой они живут, с помощью жабры. Орган лабиринта помогает кислороду во вдыхаемом воздухе поглощаться кровоток. В результате лабиринтные рыбы могут выжить в течение короткого периода времени без воды, поскольку они могут вдыхать воздух вокруг себя, если они остаются влажными.
Лабиринтные рыбы не рождаются с функциональными лабиринтными органами. Развитие органа происходит постепенно, и большинство молодых лабиринтных рыб дышат полностью жабрами и развивают органы лабиринта, когда становятся старше.[68]
Беспозвоночные
Членистоногие
Некоторые виды краб использовать дыхательный орган, называемый жаберное легкое.[69] Его жаберная структура увеличивает площадь поверхности для газообмена, который больше подходит для приема кислорода из воздуха, чем из воды. Одни из самых маленьких пауки и клещи могут дышать, просто обмениваясь газом через поверхность тела. Более крупные пауки, скорпионы и другие членистоногие использовать примитив книга легкое.
Насекомые
Большинство насекомых пассивно дышат через дыхальца (специальные отверстия в экзоскелет ), и воздух достигает каждой части тела через серию все меньших и меньших трубок, называемых «трахеями», когда их диаметр относительно велик, итрахеолы 'когда их диаметр очень мал. Трахеолы контактируют с отдельными клетками по всему телу.[44] Они частично заполнены жидкостью, которая может выводиться из отдельных трахеол, когда ткани, такие как мышцы, активны и имеют высокую потребность в кислороде, приближая воздух к активным клеткам.[44] Это, вероятно, вызвано накоплением молочной кислоты в активных мышцах, вызывая осмотический градиент, перемещающий воду из трахеол в активные клетки. Распространение газов эффективно на небольших расстояниях, но не на больших - это одна из причин, по которой все насекомые относительно малы. Насекомые, у которых нет дыхалец и трахей, например некоторые Collembola, дышат непосредственно через кожу, в том числе путем диффузии газов.[70]
Число дыхалец у насекомого варьируется между видами, однако они всегда идут парами, по одному с каждой стороны тела, и обычно по одной паре на сегмент. У некоторых Diplura их одиннадцать, с четырьмя парами на грудной клетке, но у большинства древних форм насекомых, таких как стрекозы и кузнечики, есть два грудных и восемь брюшных дыхалец. Однако у большинства оставшихся насекомых их меньше. Именно на уровне трахеол кислород доставляется к клеткам для дыхания.
Когда-то считалось, что насекомые непрерывно обмениваются газами с окружающей средой. простая диффузия газов в трахеальную систему. Однако совсем недавно были задокументированы большие различия в характере вентиляции у насекомых, и дыхание насекомых, по-видимому, сильно варьируется. Некоторые мелкие насекомые не демонстрируют непрерывных дыхательных движений и могут не контролировать дыхательные пути мышцами. Однако другие используют мышечное сокращение из брюшная полость наряду с скоординированным сокращением и расслаблением дыхательных путей для создания циклических схем газообмена и уменьшения потерь воды в атмосферу. Наиболее крайняя форма этих паттернов называется прерывистый газообмен циклы.[71]
Моллюски
Моллюски обычно имеют жабры, которые позволяют газообмену между водной средой и их кровеносными системами. У этих животных также есть сердце, которое перекачивает кровь, содержащую гемоцианин как его молекула, захватывающая кислород.[44] Следовательно, эта дыхательная система похожа на дыхательную систему позвоночных рыб. В дыхательная система брюхоногих моллюсков может включать жабры или легкое.
Растения
Растения использовать углекислый газ газ в процессе фотосинтез, и выдох кислород газ как отходы. Химическое уравнение фотосинтеза - 6 CO.2 (диоксид углерода) и 6 H2O (вода), что в присутствии солнечного света делает C6ЧАС12О6 (глюкоза) и 6 O2 (кислород). Фотосинтез использует электроны на атомах углерода в качестве хранилища энергии, получаемой от солнечного света.[72] Дыхание противоположно фотосинтезу. Он восстанавливает энергию для химических реакций в клетках. При этом атомы углерода и их электроны соединяются с кислородом, образуя CO.2 который легко удаляется как из клеток, так и из организма. Растения используют оба процесса: фотосинтез для захвата энергии и окислительный метаболизм использовать это.
Дыхание растений ограничивается процессом распространение. Растения поглощают углекислый газ через отверстия, известные как устьица, которые могут открываться и закрываться на нижней стороне их уходит а иногда и другие части их анатомии. Большинству растений требуется кислород для катаболический процессы (реакции разрушения, высвобождающие энергию). Но количество O2 используется в час, мало, поскольку они не задействованы в деятельности, требующей высоких показателей аэробный метаболизм. Однако их потребность в воздухе очень высока, поскольку они нуждаются в CO.2 для фотосинтеза, который составляет всего 0,04% окружающего воздуха. Таким образом, для получения 1 г глюкозы требуется удаление всего CO.2 от по крайней мере 18,7 литров воздуха на уровне моря. Но из-за неэффективности фотосинтетического процесса используются значительно большие объемы воздуха.[72][73]
Смотрите также
- Великое окислительное событие - Палеопротерозойский выброс атмосферного кислорода
- Дыхательная адаптация
- Спирометрия
- Исследование легочной функции (PFT)
использованная литература
- ^ Кэмпбелл, Нил А. (1990). Биология (2-е изд.). Редвуд-Сити, Калифорния: Benjamin / Cummings Pub. Co., стр. 834–835. ISBN 0-8053-1800-3.
- ^ Hsia, CC; Гайд, DM; Вейбель, ER (15 марта 2016 г.). «Структура легких и внутренние проблемы газообмена». Комплексная физиология. 6 (2): 827–95. Дои:10.1002 / cphy.c150028. ЧВК 5026132. PMID 27065169.
- ^ Запад, Джон Б. (1995). Физиология дыхания - основы. Балтимор: Уильямс и Уилкинс. стр.1–10. ISBN 0-683-08937-4.
- ^ а б Гилрой, Энн М .; Макферсон, Брайан Р .; Росс, Лоуренс М. (2008). Атлас анатомии. Штутгарт: Тиме. С. 108–111. ISBN 978-1-60406-062-1.
- ^ а б Покок, Джиллиан; Ричардс, Кристофер Д. (2006). Физиология человека: основы медицины (3-е изд.). Оксфорд: Издательство Оксфордского университета. С. 315–317. ISBN 978-0-19-856878-0.
- ^ а б c d е ж г час я j k л м п о п q р s т ты v Тортора, Джерард Дж .; Анагностакос, Николас П. (1987). Основы анатомии и физиологии (Пятое изд.). Нью-Йорк: Harper & Row, Publishers. стр.556–586. ISBN 0-06-350729-3.
- ^ Kacmarek, Роберт М .; Димас, Стивен; Мак, Крейг В. (13 августа 2013 г.). Основы респираторной помощи - электронная книга. Elsevier Health Sciences. ISBN 9780323277785.
- ^ Неттер, Фрэнк Х. (2014). Атлас анатомии человека, включая интерактивные вспомогательные материалы и руководства для студентов (6-е изд.). Филадельфия, Пенсильвания: W B Saunders Co. p. 200. ISBN 978-1-4557-0418-7.
- ^ Матон, Антея; Жан Хопкинс; Чарльз Уильям Маклафлин; Сьюзан Джонсон; Марианна Куон Уорнер; Дэвид ЛаХарт; Джилл Д. Райт (1993). Биология человека и здоровье. Вуд Клиффс, Нью-Джерси, США: Prentice Hall. ISBN 0-13-981176-1.[страница нужна ]
- ^ а б c Фаулер У.С. (1948). «Исследования функции легких. II. Мертвое пространство дыхательных путей». Am. J. Physiol. 154 (3): 405–416. Дои:10.1152 / ajplegacy.1948.154.3.405. PMID 18101134.
- ^ "анатомическое мертвое пространство". TheFreeDictionary.com.
- ^ а б Тортора, Джерард Дж .; Анагностакос, Николас П. (1987). Основы анатомии и физиологии (Пятое изд.). Нью-Йорк: Harper & Row, Publishers. стр.570–572. ISBN 0-06-350729-3.
- ^ Туровски, Джейсон (2016-04-29). "Следует ли дышать через рот или нос?". Кливлендская клиника. Получено 2020-06-28.
- ^ «Твой нос, хранитель твоих легких». Бостонский медицинский центр. Получено 2020-06-29.
- ^ Даль, Мелисса (11.01.2011). "'Грубое дыхание через рот вредно для вашего здоровья ». Новости NBC. Получено 2020-06-28.
- ^ Валчева, Зорница (январь 2018). «РОЛЬ РТОВОГО ДЫХАНИЯ В РАЗВИТИИ И ФОРМИРОВАНИИ ЗУБОВ» (PDF). Журнал IMAB. Получено 2020-05-31.
- ^ Гросс, Терри (27.05.2020). «Как« утраченное искусство »дыхания может повлиять на сон и устойчивость». Национальное общественное радио (NPR) /Свежий воздух. Получено 2020-06-23.
- ^ а б Koen, Chrisvan L .; Коэслаг, Йохан Х. (1995). «О стабильности субатмосферного внутриплеврального и внутричерепного давления». Новости физиологических наук. 10 (4): 176–178. Дои:10.1152 / Physiologyonline.1995.10.4.176.
- ^ а б Уэст, Дж. Б. (1985). Физиология дыхания: основы. Балтимор: Уильямс и Уилкинс. С. 21–30, 84–84, 98–101.
- ^ Берк, ТВ; Кюнг, М; Бурки, Н.К. (1989). «Легочный газообмен при бронхоспазме, вызванной гистамином, у пациентов с астмой». Грудь. 96 (4): 752–6. Дои:10.1378 / сундук.96.4.752. PMID 2791669. S2CID 18569280.
- ^ Тейлор, Д. (1996). «Маневр Вальсальвы: критический обзор». Журнал Южнотихоокеанского общества подводной медицины. 26 (1). ISSN 0813-1988. OCLC 16986801. Получено 14 марта 2016.
- ^ Матон, Антея; Хопкинс, Джин Сьюзен; Джонсон, Чарльз Уильям; Маклафлин, Марианна Куон; Уорнер, Дэвид; ЛаХарт Райт, Джилл (2010). Биология человека и здоровье. Энглвудские скалы: Прентис-холл. С. 108–118. ISBN 978-0134234359.
- ^ а б c Уильямс, Питер Л .; Уорик, Роджер; Дайсон, Мэри; Баннистер, Лоуренс Х. (1989). Анатомия Грея (Тридцать седьмое изд.). Эдинбург: Черчилль Ливингстон. С. 1278–1282. ISBN 0443-041776.
- ^ Лавлок, Джеймс (1991). Healing Gaia: практическая медицина для планеты. Нью-Йорк: Книги Гармонии. С. 21–34, 73–88. ISBN 0-517-57848-4.
- ^ Шу, Британская Колумбия; Чанг, ГГ; Ли, ФГ; Tzeng, DS; Lin, HY; Легкое, FW (2007-10-31). «Родительская привязанность, преморбидная личность и психическое здоровье у молодых мужчин с синдромом гипервентиляции». Психиатрические исследования. 153 (2): 163–70. Дои:10.1016 / j.psychres.2006.05.006. PMID 17659783. S2CID 3931401.
- ^ Генри Р.П., Свенсон ER (июнь 2000 г.). «Распределение и физиологическое значение карбоангидразы в органах газообмена позвоночных». Физиология дыхания. 121 (1): 1–12. Дои:10.1016 / S0034-5687 (00) 00110-9. PMID 10854618.
- ^ Diem, K .; Лентнер, К. (1970). «Кровь - неорганические вещества». в: Научные таблицы (Седьмое изд.). Базель, Швейцария: CIBA-GEIGY Ltd. стр. 571.
- ^ а б "Дыхание". Харви Проект. Получено 27 июля 2012.
- ^ а б «Высотный онлайн-калькулятор кислорода». altitude.org. Архивировано из оригинал 29 июля 2012 г.. Получено 15 августа 2007.
- ^ Тайсон, П.Д .; Престон-Уайт, Р.А. (2013). Погода и климат юга Африки. Кейптаун: Издательство Оксфордского университета. С. 3–10, 14–16, 360. ISBN 9780195718065.
- ^ Diem, K .; Лентер, К. (1970). Научные таблицы (Седьмое изд.). Базель, Швейцария: Ciba-Geigy. С. 257–258.
- ^ Фон Эйлер, США; Лильестранд, Г. (1946). «Наблюдения за артериальным давлением в легочной артерии у кошек». Acta Physiologica Scandinavica. 12 (4): 301–320. Дои:10.1111 / j.1748-1716.1946.tb00389.x.
- ^ «Обнаружение EPO». Всемирное антидопинговое агентство. Получено 7 сентября 2017.
- ^ а б Тортора, Джерард Дж .; Анагностакос, Николас П. (1987). Основы анатомии и физиологии (Пятое изд.). Нью-Йорк: Harper & Row, Publishers. стр.444–445. ISBN 0-06-350729-3.
- ^ Фишер Дж. В., Кури С., Дьюси Т., Мендель С. (1996). «Продукция эритропоэтина интерстициальными клетками почек гипоксических обезьян». Британский журнал гематологии. 95 (1): 27–32. Дои:10.1046 / j.1365-2141.1996.d01-1864.x. PMID 8857934. S2CID 38309595.
- ^ Райт, Джо Рэй (2004). «Защитные функции организма легочного сурфактанта». Биология новорожденного. 85 (4): 326–32. Дои:10.1159/000078172. PMID 15211087. S2CID 25469141.
- ^ Запад, Джон Б. (1994). Физиология дыхания - основы. Балтимор: Уильямс и Уилкинс. стр.21–30, 84–84, 98–101. ISBN 0-683-08937-4.
- ^ Салливан, LC; Оргейг, S (2001). «Дексаметазон и адреналин стимулируют секрецию сурфактанта в клетках типа II эмбриональных цыплят». Американский журнал физиологии. Регуляторная, интегративная и сравнительная физиология. 281 (3): R770–7. Дои:10.1152 / ajpregu.2001.281.3.r770. PMID 11506991.
- ^ Недоношенные дети, развитие легких и респираторный дистресс-синдром. Pregnancy-facts.com.
- ^ Канайде, Хидео; Ичики, Тошихиро; Нисимура, Дзюнджи; Хирано, Кацуя (28 ноября 2003). «Клеточный механизм сужения сосудов, вызванный ангиотензином II, еще предстоит определить». Циркуляционные исследования. 93 (11): 1015–1017. Дои:10.1161 / 01.RES.0000105920.33926.60. ISSN 0009-7330. PMID 14645130.
- ^ West, John B .; Равичандран (1993). «Дыхание слона с трубкой объясняет уникальную анатомию его плевры». Физиология дыхания. 126 (1): 1–8. Дои:10.1016 / S0034-5687 (01) 00203-1. PMID 11311306.
- ^ Запад, Джон Б. (2002). «Почему у слона нет плевральной полости?». Новости Physiol Sci. 17 (2): 47–50. Дои:10.1152 / nips.01374.2001. PMID 11909991. S2CID 27321751.
- ^ Шошани, Джеескель (декабрь 1998 г.). «Понимание эволюции хоботка: трудная задача». Тенденции в экологии и эволюции. 13 (12): 480–487. Дои:10.1016 / S0169-5347 (98) 01491-8. PMID 21238404.
- ^ а б c d е ж г Кэмпбелл, Нил А. (1990). Биология (2-е изд.). Редвуд-Сити, Калифорния: Benjamin / Cummings Pub. Co., стр. 836–844. ISBN 0-8053-1800-3.
- ^ а б Уиттоу, Дж. Кози (2000). Птичья физиология Стурки. Сан-Диего, Калифорния: Academic Press. С. 233–241. ISBN 978-0-12-747605-6.
- ^ а б c d е ж г час я j k л м п о Ритчсон, Г. «BIO 554/754 - Орнитология: дыхание птиц». Департамент биологических наук Университета Восточного Кентукки. Получено 2009-04-23.
- ^ Сторер, Трейси I .; Usinger, R.L .; Стеббинс, Роберт С .; Нибаккен, Джеймс У. (1997). Общая зоология (шестое изд.). Нью-Йорк: Макгроу-Хилл. стр.752–753. ISBN 0-07-061780-5.
- ^ Ромер, Альфред Шервуд (1970). Тело позвоночного (Четвертое изд.). Филадельфия: W.B. Сондерс. стр.323–324. ISBN 0-7216-7667-7.
- ^ а б c Скотт, Грэм Р. (2011). «Комментарий: Повышенная производительность: уникальная физиология птиц, летающих на больших высотах». Журнал экспериментальной биологии. 214 (Pt 15): 2455–2462. Дои:10.1242 / jeb.052548. PMID 21753038.
- ^ а б Майна, Джон Н. (2005). Система развития, строения и функции легочного воздушного мешка птиц; с 6 столами. Берлин: Springer. С. 3.2–3.3 «Легкое», «Система дыхательных путей (бронхиальная)» 66–82. ISBN 978-3-540-25595-6.
- ^ Краутвальд-Юнгханс, Мария-Элизабет; и другие. (2010). Диагностическая визуализация экзотических домашних животных: птиц, мелких млекопитающих, рептилий. Германия: Manson Publishing. ISBN 978-3-89993-049-8.
- ^ а б Стурки, П. (1976). Стурки, П. Д. (ред.). Птичья физиология. Нью-Йорк: Springer Verlag. п. 201. Дои:10.1007/978-1-4612-4862-0. ISBN 978-1-4612-9335-4. S2CID 36415426.
- ^ Ричисон, Гэри. «Орнитология (Bio 554/754): дыхательная система птиц». Университет Восточного Кентукки. Проверено 27 июня 2007.
- ^ Дыхательная система. Encyclopdia Britannica.
- ^ Готтлиб, G; Джексон, округ Колумбия (1976). «Важность легочной вентиляции в контроле дыхания лягушки-быка». Am J Physiol. 230 (3): 608–13. Дои:10.1152 / ajplegacy.1976.230.3.608. PMID 4976.
- ^ а б c Кэмпбелл, Нил А. (1990). Биология (Второе изд.). Редвуд-Сити, Калифорния: Бенджамин / Каммингс Паблишинг Компани, Инк., Стр. 836–838. ISBN 0-8053-1800-3.
- ^ а б c Хьюз GM (1972). «Морфометрия жабр рыб». Физиология дыхания. 14 (1–2): 1–25. Дои:10.1016 / 0034-5687 (72) 90014-х. PMID 5042155.
- ^ а б c d е ж Сторер, Трейси I .; Usinger, R.L .; Стеббинс, Роберт С .; Нюбаккен, Джеймс У. (1997). Общая зоология (шестое изд.). Нью-Йорк: Макгроу-Хилл. стр.668–670. ISBN 0-07-061780-5.
- ^ а б М. б. против Робертса; Майкл Рейсс; Грейс Монгер (2000). Продвинутая биология. Лондон, Великобритания: Нельсон. С. 164–165.
- ^ Касслер, Э. Л. (1997). Диффузия: массоперенос в жидкостных системах (2-е изд.). Нью-Йорк: Издательство Кембриджского университета. ISBN 0-521-45078-0.
- ^ Велти, Джеймс Р .; Уикс, Чарльз Э .; Уилсон, Роберт Э .; Роррер, Грегори (2001). Основы переноса количества движения, тепла и массы. Вайли. ISBN 978-0-470-12868-8.
- ^ а б CRC Press Online: Справочник CRC по химии и физике, раздел 6, 91-е издание
- ^ а б Распространение
- ^ Ньюстед Джеймс Д. (1967). «Тонкое строение дыхательных пластинок жабр костистых животных». Исследования клеток и тканей. 79 (3): 396–428. Дои:10.1007 / bf00335484. PMID 5598734. S2CID 20771899.
- ^ Ромер, Альфред Шервуд; Парсонс, Томас С. (1977). Тело позвоночного. Филадельфия, Пенсильвания: Holt-Saunders International. С. 316–327. ISBN 0-03-910284-X.
- ^ Гилбертсон, Ланс (1999). Руководство зоологической лаборатории. Нью-Йорк: Макгроу-Хилл. ISBN 0-07-237716-X.
- ^ Уильям Дж. Беннетта (1996). "Глубокое дыхание". Получено 2007-08-28.
- ^ а б Пинтер, Х. (1986). Лабиринт Рыбка. Образовательная серия Barron's, Inc., ISBN 0-8120-5635-3
- ^ Гальперин Дж, Ансальдо М, Пеллерано Г.Н., Люке С.М. (июль 2000 г.). «Бимодальное дыхание у эстуарного краба Chasmagnathus granulatus Dana 1851 - физиологические и морфологические исследования». Сравнительная биохимия и физиология. Часть A, Молекулярная и интегративная физиология. 126 (3): 341–9. Дои:10.1016 / S1095-6433 (00) 00216-6. PMID 10964029.
- ^ Сеть Earth Life, морфология и анатомия насекомых. Earthlife.net. Проверено 21 апреля 2013.
- ^ Лайтон, JRB (январь 1996 г.). «Прерывистый газообмен у насекомых». Анну Рев Энтомол. 41: 309–324. Дои:10.1146 / annurev.en.41.010196.001521. PMID 8546448.
- ^ а б Страйер, Люберт (1995). «Фотосинтез». В: Биохимия. (Четвертое изд.). Нью-Йорк: W.H. FreeMan and Company. С. 653–680. ISBN 0-7167-2009-4.
- ^ Кэмпбелл, Нил А. (1990). Биология (Второе изд.). Редвуд-Сити, Калифорния: Бенджамин / Каммингс Паблишинг Компани, Инк., Стр. 206–223. ISBN 0-8053-1800-3.
внешние ссылки
- Описание дыхательной системы на уровне средней школы
- Введение в дыхательную систему
- Научное пособие: Дыхательная система Простое руководство для старшеклассников
- Дыхательная система Уровень университета (документ Microsoft Word)
- Лекции по физиологии дыхания от известного респираторного физиолога Джон Б. Уэст (также на YouTube )
| Библиотечные ресурсы около Дыхательная система |
| Авторитетный контроль |
|---|