Стероид - Википедия - Steroid

А стероидный препарат является биологически активным органическое соединение с четырьмя кольцами, расположенными в определенном молекулярная конфигурация. Стероиды выполняют две основные биологические функции: как важные компоненты клеточные мембраны которые изменяют текучесть мембраны; и, как сигнальные молекулы. Сотни стероидов находятся в растения, животные и грибы. Все стероиды производятся в клетках из стеролы ланостерин (опистоконцы ) или же циклоартенол (растения). Ланостерин и циклоартенол являются производными циклизация из тритерпен сквален.[2]
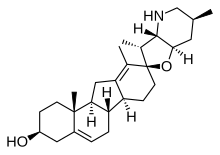
Стероид основная структура обычно состоит из семнадцати углерод атомы, соединенные в четыре "сплавлен "кольца: три шестичленных циклогексан кольца (кольца A, B и C на первой иллюстрации) и одно пятичленное циклопентан кольцо (кольцо D). Стероиды различаются функциональные группы прикрепленный к этому четырехкольцевому сердечнику и степень окисления колец. Стерины формы стероидов с гидроксильная группа в позиции три и скелет, полученный из холестан.[1]:1785f[3] Стероиды также могут быть изменены более радикально, например, путем изменения кольцевой структуры, резка одно из колец. Режущее кольцо B производит секостероиды один из которых Витамин Д3.
Примеры включают липид холестерин, половые гормоны эстрадиол и тестостерон,[4]:10–19 и противовоспалительное средство препарат, средство, медикамент дексаметазон.[5]


Номенклатура


Gonane, также известный как стеран или циклопентанопергидрофенантрен, простейший стероид и ядро всех стероидов и стеринов,[6][7] состоит из семнадцати углерод атомы в углерод-углеродных связях, образующих четыре сплавленные кольца в трехмерная форма. Три циклогексан кольца (A, B и C на первой иллюстрации) образуют скелет Perhydro производная от фенантрен. Кольцо D имеет циклопентан структура. Когда две метильные группы и восемь атомов углерода боковые цепи (в C-17, как показано для холестерина) стероид, как говорят, имеет холестановый каркас. Две общие 5α и 5β стереоизомерные формы стероидов существуют из-за различий в стороне в основном плоской кольцевой системы, к которой присоединен атом водорода (H) у углерода-5, что приводит к изменению конформации стероидного A-кольца. Изомеризация боковой цепи C-21 дает параллельный ряд соединений, называемых изостероидами.[8]
Примеры стероидных структур:

Тестостерон, главный мужчина половой гормон и анаболический стероид

Холевая кислота, а желчная кислота, показывая карбоновая кислота и дополнительные гидроксильные группы часто присутствует

Дексаметазон синтетический кортикостероид препарат, средство, медикамент

Ланостерол, то биосинтетический предшественник стероидов животного происхождения. Число атомов углерода (30) указывает на его тритерпеноид классификация.

Прогестерон, стероидный гормон, участвующий в женском менструальном цикле, беременности и эмбриогенезе

Медрогестон, синтетический препарат с действием, аналогичным прогестерону

β-ситостерин, растение или фитостерин, с полностью разветвленной углеводородной боковой цепью у C-17 и гидроксильной группой у C-3







Помимо кольцевых разрывов (расщеплений), расширения и схватки (расщепление и повторное замыкание с образованием большего или меньшего кольца) - все варианты каркаса углерод-углеродной связи - стероиды также могут варьироваться:
- в поручения на облигации внутри колец,
- в количестве метильных групп, присоединенных к кольцу (и, если они присутствуют, на заметной боковой цепи у C17),
- в функциональных группах, присоединенных к кольцам и боковой цепи, и
- в конфигурация групп, прикрепленных к кольцам и цепочке.[4]:2–9
Например, стеролы такие как холестерин и ланостерин имеют гидроксильная группа прикреплен в позиции C-3, а тестостерон и прогестерон иметь карбонил (оксозаместитель) у С-3; из этих, ланостерин сам по себе имеет две метильные группы у C-4, и холестерин (с двойной связью C-5-C-6) отличается от тестостерона и прогестерона (которые имеют двойную связь C-4-C-5).
 Холестерин, а прототип стерин животного происхождения. Эта структурная липид и ключевой стероид биосинтетический предшественник.[1]:1785f |  5α-холестан, общий стероидный сердечник |
Распространение и функции видов
Эта секция требует внимания специалиста по фармакологии. Конкретная проблема: для изучения этого и следующего раздела (и повсюду), а также для удаления избыточности перечисленного содержимого и обеспечения поиска источников для указанного содержимого, которое остается в любом разделе. (Март 2017 г.) |
В эукариоты, стероиды содержатся в грибах, животных и растениях.
Грибковые стероиды
Грибковые стероиды включают эргостерины, которые участвуют в поддержании целостности клеточной мембраны грибка. Разные противогрибковые препараты, Такие как амфотерицин B и азольные противогрибковые препараты, используйте эту информацию, чтобы убить патогенный грибы.[9] Грибы могут изменять содержание эргостерина (например, из-за мутаций потери функций в ферментах ERG3 или же ERG6, вызывающие истощение эргостерина или мутации, снижающие содержание эргостерина) для развития устойчивости к лекарствам, нацеленным на эргостерин.[10] Эргостерол аналогичен холестерин обнаружены в клеточных мембранах животных (включая человека), или фитостерины содержится в клеточных мембранах растений.[10] Все грибы содержат большое количество эргостерина, от десятков до сотен миллиграммов на 100 граммов сухого веса.[10] Кислород необходим для синтеза эргостерин в грибах.[10] Эргостерол отвечает за Витамин Д содержание в грибах; эргостерин химически превращается в провитамин D2 под воздействием ультрафиолетовый свет.[10] Провитамин D2 спонтанно образует витамин D2.[10] Однако не все грибы используют эргостерин в своих клеточных мембранах; например, патогенные виды грибов Pneumocystis jirovecii нет, что имеет важное клиническое значение (учитывая механизм действия многих противогрибковых препаратов).[10] Использование грибка Saccharomyces cerevisiae например, другие основные стероиды включают эргоста-5,7,22,24 (28) -тетраен-3β-ол, зимостерин, и ланостерин.[10] С. cerevisiae использует 5,6 ‐ дигидроэргостерол вместо эргостерола в его клеточной мембране.[10]
Стероиды для животных
Стероиды для животных включают соединения позвоночное животное и насекомое происхождение, последний в том числе экдистероиды Такие как экдистерон (контроль линьки у некоторых видов). Примеры позвоночных включают стероидные гормоны и холестерин; последний является структурным компонентом клеточные мембраны что помогает определить текучесть клеточные мембраны и является основным компонентом бляшка (причастен к атеросклероз ). Стероидные гормоны включают:
- Половые гормоны, которые влияют половые различия и поддержка воспроизведение. К ним относятся андрогены, эстрогены, и прогестагены.
- Кортикостероиды, включая большинство синтетических стероидных препаратов, с натуральный продукт классы глюкокортикоиды (которые регулируют многие аспекты метаболизм и иммунная функция ) и минералокортикоиды (которые помогают поддерживать объем крови и контролировать почечный выделение электролиты )
- Анаболические стероиды, естественный и синтетические, которые взаимодействуют с рецепторами андрогенов для увеличения синтеза мышц и костей. В популярном использовании термин «стероиды» часто относится к анаболическим стероидам.
Растительные стероиды
Растительные стероиды включают стероидные алкалоиды нашел в Пасленовые[11] и Мелантиевые (особенно род Veratrum ),[12] сердечные гликозиды,[13] то фитостерины и брассиностероиды (в состав которых входит несколько гормонов растений).
Прокариоты
В прокариоты, биосинтетические пути существуют для тетрациклического стероидного каркаса (например, в микобактерии )[14] - откуда его происхождение эукариоты предполагается[15] - и более распространенный пентациклический тритерпиноид гопаноид рамки.[16]
Типы
По функциям
Эта секция нуждается в расширении с: В этом списке не обсуждаются липидные стероиды. Также было бы полезно более подробное объяснение функции. Вы можете помочь добавляя к этому. (Январь 2019) |
Основные классы стероидные гормоны, с известными членами и примерами связанных функций:[нужна цитата ]
- Кортикостероиды:
- Глюкокортикоиды:
- Кортизол, а глюкокортикоид чьи функции включают иммуносупрессия
- Минералокортикоиды:
- Альдостерон, а минералокортикоид что помогает регулировать артериальное давление за счет водно-электролитного баланса
- Глюкокортикоиды:
- Половые стероиды:
- Гестагены:
- Прогестерон, регулирующий циклические изменения эндометрий из матка и поддерживает беременность
- Андрогены:
- Тестостерон, что способствует развитию и сохранению мужского вторичные половые признаки
- Эстрогены:
- Эстрадиол, что способствует развитию и поддержанию женских вторичных половых признаков
- Гестагены:
Дополнительные классы стероидов включают:
- Нейростероиды Такие как DHEA и аллопрегнанолон
- Аминостероид нервно-мышечные блокаторы Такие как бромид панкурония
А также следующий класс секостероиды (стероиды открытого кольца):
- Витамин Д формы, такие как эргокальциферол, холекальциферол, и кальцитриол
По структуре
Неповрежденная кольцевая система
Эта секция нуждается в расширении с: более полное обсуждение этого наиболее известного структурного типа. Вы можете помочь добавляя к этому. (Март 2017 г.) |
Стероиды можно классифицировать по химическому составу.[17] Один пример того, как MeSH выполняет эту классификацию. каталог Wikipedia MeSH. Примеры этой классификации включают:

| Учебный класс | Пример | Количество атомов углерода |
|---|---|---|
| Холестаны | Холестерин | 27 |
| Холаны | Холевая кислота | 24 |
| Прегнанес | Прогестерон | 21 |
| Андростаны | Тестостерон | 19 |
| Estranes | Эстрадиол | 18 |
Гонан (стероидное ядро) является родительской молекулой 17-углеродного тетрациклического углеводорода без алкил сайдчейны.[18]
Расщепленные, сжатые и расширенные кольца
Секостероиды (лат. seco, "резать") являются подклассом стероидных соединений, в результате чего, биосинтетически или концептуально от расщепления (расщепления) родительских стероидных колец (обычно одного из четырех). Основные подклассы секостероидов определяются атомами углерода стероидов, в которых произошло это расщепление. Например, прототипный секостероид холекальциферол, Витамин Д3 (показано), относится к подклассу 9,10-секостероидов и происходит от расщепления атомов углерода С-9 и С-10 стероидного В-кольца; 5,6-секостероиды и 13,14-стероиды аналогичны.[19]
Норстероиды (ни-, Л. Норма; "нормальный" по химии, указывающий на удаление углерода)[20] и гомостероиды (гомо-, греч. гомо; «то же самое», что означает добавление углерода) представляют собой структурные подклассы стероидов, образованные на этапах биосинтеза. Первый включает ферментативные расширение-сжатие кольца реакции, и последнее осуществляется (биомиметически ) или (чаще) через кольца из ациклический предшественники с большим (или меньшим) числом кольцевых атомов, чем у исходного стероидного каркаса.[21]
Комбинации этих изменений кольца известны в природе. Например, овцы кто пасется на кукурузная лилия глотать циклопамин (показано) и вератрамин, два из подсемейства стероидов, в которых C- и D-кольца сжимаются и расширяются соответственно через биосинтетический миграция исходного атома C-13. Прием этих C-nor-D-гомостероидов приводит к врожденным дефектам у ягнят: циклопия из циклопамин и деформация ног из-за вератрамина.[22] Еще один C-нор-D-гомостероид (накитерпиозин) выводится с помощью Окинавский цианобактериальные губки. например., Терпиос Hoshinota, что приводит к гибели кораллов от болезни черных кораллов.[23] Стероиды накитерпиозинового типа активны в отношении сигнального пути, включающего сглаженный и Ежик белки, путь, который является гиперактивным при ряде видов рака.[нужна цитата ]
Биологическое значение
Стероиды и их метаболиты часто действуют как сигнализация молекулы (наиболее яркими примерами являются стероидные гормоны), а также стероиды и фосфолипиды компоненты клеточные мембраны.[24] Стероиды, такие как холестерин, снижают текучесть мембраны.[25]Похожий на липиды, стероиды представляют собой высококонцентрированные запасы энергии. Однако обычно они не являются источниками энергии; у млекопитающих они обычно метаболизируются и выводятся из организма.
Стероиды играют решающую роль в ряде заболеваний, включая злокачественные новообразования, такие как: рак простаты, где производство стероидов внутри и вне опухоли способствует агрессивности раковых клеток.[26]
Биосинтез и обмен веществ
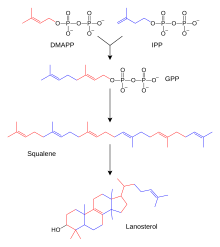
Сотни стероидов, содержащихся в животных, грибах и растения сделаны из ланостерин (у животных и грибов; см. примеры выше) или циклоартенол (в растениях). Ланостерин и циклоартенол происходят из циклизация из тритерпеноид сквален.[2]
Биосинтез стероидов - это анаболический путь, который производит стероиды из простых предшественников. У животных наблюдается уникальный биосинтетический путь (по сравнению со многими другими организмы ), что делает этот путь общей целью для антибиотики и другие противоинфекционные препараты. Метаболизм стероидов у людей также является мишенью для снижения холестерина, таких как статины.
У людей и других животных биосинтез стероидов происходит по мевалонатному пути, который использует ацетил-КоА как строительные блоки для диметилаллил пирофосфат (DMAPP) и изопентенилпирофосфат (IPP).[27][нужен лучший источник ] На последующих этапах DMAPP и IPP объединяются, чтобы сформировать геранилпирофосфат (GPP), который синтезирует стероидный ланостерин. Модификации ланостерола другими стероидами классифицируются как трансформации стероидогенеза.[28]
Мевалонатный путь

Путь мевалоната (также называемый путем HMG-CoA редуктазы) начинается с ацетил-КоА и заканчивается диметилаллил пирофосфат (DMAPP) и изопентенилпирофосфат (IPP).
DMAPP и IPP жертвуют изопрен единицы, которые собираются и модифицируются, чтобы сформировать терпены и изопреноиды[29] (большой класс липидов, в который входят каротиноиды и образуют самый большой класс растений натуральные продукты.[30] Здесь изопреновые звенья соединяются для получения сквален и сложенный в набор колец, чтобы сделать ланостерин.[31] Затем ланостерин может быть преобразован в другие стероиды, такие как холестерин и эргостерин.[31][32]
Два класса наркотики нацелить мевалонатный путь: статины (подобно розувастатин ), которые используются для уменьшения повышенный уровень холестерина,[33] и бисфосфонаты (подобно золедронат ), которые используются для лечения ряда костно-дегенеративных заболеваний.[34]
Стероидогенез
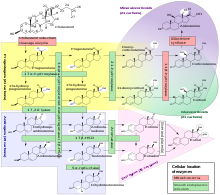
Стероидогенез - это биологический процесс, при котором стероиды производятся из холестерина и превращаются в другие стероиды.[36] В пути стероидогенеза различаются между видами. Как отмечалось выше, основными классами стероидных гормонов (с их важными членами и функциями) являются: Прогестаген, Кортикостероиды (кортикоиды), Андрогены, и Эстрогены.[37][нужна цитата ] Человеческий стероидогенез этих классов происходит в нескольких местах:
- Прогестагены являются предшественниками всех других человеческих стероидов, и все ткани человека, вырабатывающие стероиды, должны сначала преобразовать холестерин в прегненолон. Это преобразование является лимитирующим этапом синтеза стероидов, который происходит внутри митохондрия соответствующей ткани.[38][37][нужен лучший источник ]
- Кортизол, кортикостерон, альдостерон и тестостерон производятся в кора надпочечников.[37][нужна цитата ]
- Эстрадиол, эстрон и прогестерон производятся в основном в яичник, эстриол в плацента во время беременности и тестостерон прежде всего в яички (часть тестостерона также вырабатывается в коре надпочечников).[37][нужна цитата ]
- Эстрадиол преобразуется из тестостерона напрямую (у мужчин) или через первичный путь DHEA - андростендион - эстрон и вторично - через тестостерон (у женщин).[37][нужна цитата ]
- Стромальные клетки было показано, что они вырабатывают стероиды в ответ на передачу сигналов, производимых голодными по андрогенам рак простаты клетки.[39][нужен лучший источник ]
- Немного нейроны и глия в Центральная нервная система (CNS) выражают ферменты необходим для местного синтеза прегненолона, прогестерона, DHEA и DHEAS, de novo или из периферийных источников.[37][нужна цитата ]
| Секс | Половой гормон | Репродуктивный фаза | Кровь дебит | Гонад скорость секреции | Метаболический скорость оформления | Референсный диапазон (уровни сыворотки) | |
|---|---|---|---|---|---|---|---|
| SI единицы | Не-SI единицы | ||||||
| Мужчины | Андростендион | – | 2,8 мг / день | 1,6 мг / день | 2200 л / сутки | 2,8-7,3 нмоль / л | 80–210 нг / дл |
| Тестостерон | – | 6,5 мг / день | 6,2 мг / день | 950 л / сутки | 6,9–34,7 нмоль / л | 200–1000 нг / дл | |
| Estrone | – | 150 мкг / день | 110 мкг / день | 2050 л / сутки | 37–250 пмоль / л | 10–70 пг / мл | |
| Эстрадиол | – | 60 мкг / день | 50 мкг / день | 1600 л / сутки | <37–210 пмоль / л | 10–57 пг / мл | |
| Эстрона сульфат | – | 80 мкг / день | Незначительный | 167 л / сутки | 600–2500 пмоль / л | 200–900 пг / мл | |
| Женщины | Андростендион | – | 3,2 мг / день | 2,8 мг / день | 2000 л / сутки | 3,1–12,2 нмоль / л | 89–350 нг / дл |
| Тестостерон | – | 190 мкг / день | 60 мкг / день | 500 л / сутки | 0,7–2,8 нмоль / л | 20–81 нг / дл | |
| Estrone | Фолликулярная фаза | 110 мкг / день | 80 мкг / день | 2200 л / сутки | 110–400 пмоль / л | 30–110 пг / мл | |
| Лютеиновой фазы | 260 мкг / день | 150 мкг / день | 2200 л / сутки | 310–660 пмоль / л | 80–180 пг / мл | ||
| Постменопауза | 40 мкг / день | Незначительный | 1610 л / сутки | 22–230 пмоль / л | 6–60 пг / мл | ||
| Эстрадиол | Фолликулярная фаза | 90 мкг / день | 80 мкг / день | 1200 л / сутки | <37–360 пмоль / л | 10–98 пг / мл | |
| Лютеиновой фазы | 250 мкг / день | 240 мкг / день | 1200 л / сутки | 699–1250 пмоль / л | 190–341 пг / мл | ||
| Постменопауза | 6 мкг / день | Незначительный | 910 л / сутки | <37–140 пмоль / л | 10–38 пг / мл | ||
| Эстрона сульфат | Фолликулярная фаза | 100 мкг / день | Незначительный | 146 л / сутки | 700–3600 пмоль / л | 250–1300 пг / мл | |
| Лютеиновой фазы | 180 мкг / день | Незначительный | 146 л / сутки | 1100–7300 пмоль / л | 400–2600 пг / мл | ||
| Прогестерон | Фолликулярная фаза | 2 мг / день | 1,7 мг / день | 2100 л / сутки | 0,3–3 нмоль / л | 0,1–0,9 нг / мл | |
| Лютеиновой фазы | 25 мг / день | 24 мг / день | 2100 л / сутки | 19–45 нмоль / л | 6–14 нг / мл | ||
Примечания и источники Примечания: "The концентрация Количество стероида в кровотоке определяется скоростью, с которой он секретируется железами, скоростью метаболизма предшественника или прегормонов в стероид и скоростью, с которой он извлекается тканями и метаболизируется. В скорость секреции стероида относится к общей секреции соединения железой за единицу времени. Скорость секреции оценивалась путем отбора проб венозного стока из железы с течением времени и вычитания концентрации артериальных и периферических венозных гормонов. В скорость метаболического клиренса стероида определяется как объем крови, который полностью очищен от гормона за единицу времени. В дебит стероидного гормона относится к поступлению в кровь соединения из всех возможных источников, включая секрецию желез и превращение прогормонов в интересующий стероид. В устойчивом состоянии количество гормона, поступающего в кровь из всех источников, будет равно скорости, с которой он очищается (скорость метаболического клиренса), умноженной на концентрацию в крови (скорость продукции = скорость метаболического клиренса × концентрация). Если метаболизм прогормонов вносит небольшой вклад в циркулирующий пул стероидов, то скорость производства будет приблизительно соответствовать скорости секреции ». Источники: См. Шаблон. | |||||||
Альтернативные пути
У растений и бактерий немевалонатный путь использует пируват и глицеральдегид-3-фосфат в качестве подложек.[29][40]
Во время болезней могут быть задействованы пути, которые иначе не значимы для здоровых людей. Например, в одной из форм врожденная гиперплазия надпочечников а дефицит ферментативного пути 21-гидроксилазы приводит к превышению 17α-гидроксипрогестерон (17-ОНР) - этот патологический избыток 17-ОНР, в свою очередь, может быть преобразован в дигидротестостерон (DHT, мощный андроген), среди прочего 17,20 Лиазе (член цитохром P450 семейство ферментов), 5α-редуктаза и 3α-гидроксистероид дегидрогеназа.[41]
Катаболизм и выведение
Стероиды в первую очередь окисляются оксидаза цитохрома P450 ферменты, такие как CYP3A4. Эти реакции вводят кислород в стероидное кольцо, позволяя другим ферментам расщеплять холестерин на желчные кислоты.[42] Эти кислоты затем могут быть удалены путем секреции из печень в желчь.[43] Выражение оксидаза ген может быть усиленный датчиком стероидов PXR при высокой концентрации стероидов в крови.[44] Стероидные гормоны, лишенные боковой цепи холестерина и желчных кислот, обычно гидроксилированный в различных положениях звонка или окислен по 17 позиции, сопряженный с сульфатом или глюкуроновая кислота и выводится с мочой.[45]
Изоляция, определение структуры и методы анализа
Стероидный препарат изоляция, в зависимости от контекста, требуется ли выделение химического вещества для химическая структура выяснение, дериватизация или химия разложения, биологические испытания и другие исследовательские потребности (обычно от миллиграммов до граммов, но часто и больше[46] или выделение «аналитических количеств» интересующего вещества (где основное внимание уделяется идентификации и количественному определению вещества (например, в биологической ткани или жидкости). Выделенное количество зависит от аналитического метода, но обычно меньше единицы микрограмм.[47][страница нужна ] Методы изоляции для достижения двух масштабов продукта различны, но включают: добыча, осадки, адсорбция, хроматография, и кристаллизация. В обоих случаях изолированное вещество очищают до химической однородности; комбинированные методы разделения и анализа, такие как ЖХ-МС, выбираются так, чтобы быть «ортогональными» - достигая их разделения на основе различных способов взаимодействия между веществом и изолирующей матрицей - для обнаружения одного вида в чистом образце. Определение структуры относится к методам определения химической структуры изолированного чистого стероида с использованием постоянно развивающегося набора химических и физических методов, которые включают ЯМР и малая молекула кристаллография.[4]:10–19 Методы анализа перекрывают обе вышеупомянутые области, делая упор на аналитических методах определения наличия стероида в смеси и определения его количества.[47]
Химический синтез
Микробный катаболизм из фитостерин боковые цепи дает стероиды C-19, стероиды C-22 и 17-кетостероиды (т.е. предшественники к адренокортикальные гормоны и контрацептивы ).[48][49][50] Добавление и модификация функциональные группы является ключевым при производстве широкого спектра лекарств, доступных в рамках этой химической классификации. Эти модификации выполняются с использованием обычных органический синтез и / или биотрансформация техники.[51][52]
Прекурсоры
Полусинтез
В полусинтез стероидов часто начинается с предшественников, таких как холестерин,[50] фитостерины,[49] или же сапогенины.[53] Усилия Syntex, компания, занимающаяся Мексиканская торговля барбаско, использовал Диоскорея мексиканская производить сапогенин диосгенин в первые дни синтетического стероида фармацевтическая индустрия.[46]
Полный синтез
Некоторые стероидные гормоны экономически доступны только полный синтез из нефтехимия (например, 13-алкил стероиды).[50] Например, фармацевтическая Норгестрел начинается с Метокси -1-тетралон, нефтехимический продукт, полученный из фенол.
Награды за исследования
Номер Нобелевские премии были награждены за исследования стероидов, в том числе:
- 1927 (Химия ) Генрих Отто Виланд - Состав желчных кислот и стеролов и их связь с витаминами[54]
- 1928 (химия) Адольф Отто Рейнхольд Виндаус - Состав стеринов и их связь с витаминами[55]
- 1939 (химия) Адольф Бутенандт и Леопольд Ружичка - Изоляция и структурные исследования стероидных половых гормонов и связанные с ними исследования высших терпены[56]
- 1950 (Физиология или медицина ) Эдвард Кэлвин Кендалл, Тадеус Райхштейн, и Филип Хенч - Структура и биологические эффекты гормоны надпочечников[57]
- 1965 (химия) Роберт Бернс Вудворд - Отчасти для синтеза холестерина, кортизон, и ланостерин[58]
- 1969 (химия) Дерек Бартон и Odd Hassel - Развитие концепции конформации в химии с акцентом на стероидное ядро[59]
- 1975 (химия) Владимир Прелог - В частности, для разработки методов определения стереохимического течения биосинтеза холестерина из мевалоновая кислота через сквален[60]
Смотрите также
Рекомендации
- ^ а б c d Moss GP, рабочая группа Совместной комиссии IUPAC-IUB по биохимической номенклатуре (1989). «Номенклатура стероидов, рекомендации 1989 г.» (PDF). Pure Appl. Chem. 61 (10): 1783–1822. Дои:10.1351 / pac198961101783. Также доступно с теми же авторами на Карлсон П., Бык-младший, Энгель К, Фрид Дж., Кирхер Х.В., Ссуды К.Л., Мосс Г.П., Попьяк Г., Ускокович М.Р. (декабрь 1989 г.). «Совместная комиссия IUPAC-IUB по биохимической номенклатуре (JCBN). Номенклатура стероидов. Рекомендации 1989». Европейский журнал биохимии / FEBS. 186 (3): 429–58. Дои:10.1111 / j.1432-1033.1989.tb15228.x. PMID 2606099.; Также доступно на сайте «Номенклатура стероидов». Лондон, Великобритания: Лондонский университет королевы Марии. п. 3S – 1.4. Получено 10 мая 2014.
- ^ а б «Биосинтез ланостерола». Рекомендации по биохимической и органической номенклатуре, символам и терминологии. Международный союз биохимии и молекулярной биологии. Архивировано из оригинал на 2011-03-08. Получено 2006-11-28.
- ^ Также доступно в печати на Хилл Р.А., Макин Х.Л., Кирк Д.Н., Мерфи Г.М. (1991). Словарь стероидов. Лондон, Великобритания: Чепмен и Холл. стр. xxx – lix. ISBN 978-0412270604. Получено 20 июн 2015.
- ^ а б c Ледничер Д. (2011). Краткий обзор химии стероидов. Хобокен: Вайли. ISBN 978-0-470-66084-3.
- ^ Рен Т., Цидловски Дж. А. (октябрь 2005 г.). «Противовоспалительное действие глюкокортикоидов - новые механизмы для старых препаратов» (PDF). Медицинский журнал Новой Англии. 353 (16): 1711–23. Дои:10.1056 / NEJMra050541. PMID 16236742.
- ^ Виктор Анатольевич Рогозкин (14 июня 1991 г.). Метаболизм анаболических андрогенных стероидов. CRC Press. стр. 1–. ISBN 978-0-8493-6415-0.
Структурная основа стероида представляет собой стерановое ядро, полициклический стерановый каркас C17, состоящий из трех конденсированных циклогексановых колец в нелинейных или фенантреновых соединениях (A, B и C) и циклопентанового кольца (D) .1,2
- ^ Клаус Урих (16 сентября 1994 г.). Сравнительная биохимия животных. Springer Science & Business Media. стр. 624–. ISBN 978-3-540-57420-0.
- ^ Greep 2013.
- ^ Бхетария П.Дж., Шарма Н., Сингх П., Трипати П., Упадхьяй С.К., Гаутам П. (21 марта 2017 г.). «Человеческие грибковые патогены и лекарственная устойчивость к азольным препаратам». В Arora C, Sajid A, Kalia V (ред.). Устойчивость к лекарствам бактерий, грибов, малярии и рака. Springer. ISBN 978-3-319-48683-3.
- ^ а б c d е ж грамм час я Кавана, Кевин, изд. (8 сентября 2017 г.). Грибы: биология и применение. John Wiley & Sons, Inc. ISBN 9781119374312.
- ^ Wink M (сентябрь 2003 г.). «Эволюция вторичных метаболитов с экологической и молекулярно-филогенетической точки зрения». Фитохимия. 64 (1): 3–19. Дои:10.1016 / S0031-9422 (03) 00300-5. PMID 12946402.
- ^ Подмигнуть, Майкл; ван Вик, Бен-Эрик (2008). Изменяющие сознание и ядовитые растения мира. Портленд (Орегон, США) и Салусбери (Лондон, Англия): Timber press inc. С. 252, 253 и 254. ISBN 978-0-88192-952-2.
- ^ Подмигнуть, Майкл; ван Вик, Бен-Эрик (2008). Изменяющие сознание и ядовитые растения мира. Портленд (Орегон, США) и Салусбери (Лондон, Англия): Timber press inc. С. 324, 325 и 326. ISBN 978-0-88192-952-2.
- ^ Боде HB, Зеггель Б., Силаковски Б., Венцель С.К., Райхенбах Х., Мюллер Р. (январь 2003 г.). «Биосинтез стероидов у прокариот: идентификация миксобактериальных стероидов и клонирование первой бактериальной 2,3 (S) -оксидоскваленциклазы из миксобактерий Stigmatella aurantiaca». Молекулярная микробиология. 47 (2): 471–81. Дои:10.1046 / j.1365-2958.2003.03309.x. PMID 12519197.
- ^ Десмонд Э, Грибальдо С (2009). «Филогеномика синтеза стеролов: понимание происхождения, эволюции и разнообразия ключевой эукариотической особенности». Геномная биология и эволюция. 1: 364–81. Дои:10.1093 / gbe / evp036. ЧВК 2817430. PMID 20333205.
- ^ Зиденбург Г., Ендроссек Д. (июнь 2011 г.). «Сквален-гопенциклазы». Прикладная и экологическая микробиология. 77 (12): 3905–15. Дои:10.1128 / AEM.00300-11. ЧВК 3131620. PMID 21531832.
- ^ Зорея, Аарон (2014). Стероиды (здоровье и медицинские проблемы сегодня). Вестпорт, Коннектикут: Greenwood Press. С. 10–12. ISBN 978-1440802997.
- ^ Эдгрен Р.А., Станчик Ф.З. (декабрь 1999 г.). «Номенклатура гонановых прогестинов». Контрацепция. 60 (6): 313. Дои:10.1016 / S0010-7824 (99) 00101-8. PMID 10715364.
- ^ Hanson JR (июнь 2010 г.). «Стероиды: частичный синтез в медицинской химии». Отчеты о натуральных продуктах. 27 (6): 887–99. Дои:10.1039 / c001262a. PMID 20424788.
- ^ «Рекомендации ИЮПАК: Модификация скелета в пересмотренном разделе F: Натуральные продукты и родственные соединения (Рекомендации ИЮПАК 1999 г.)». Международный союз теоретической и прикладной химии (IUPAC). 1999 г.
- ^ Вольфинг Дж. (2007). «Последние разработки в области выделения и синтеза D-гомостероидов и родственных соединений». Аркивок: 210–230.
- ^ Гао Г, Чен С. (2012). «Накитерпиозин». В Corey E, Li JJ (ред.). Полный синтез натуральных продуктов: на переднем крае органической химии. Берлин: Springer. Дои:10.1007/978-3-642-34065-9. ISBN 978-3-642-34064-2.
- ^ Уэмура Э, Кита М, Аримото Х, Китамура М (2009). «Последние аспекты химической экологии: природные токсины, коралловые сообщества и симбиотические отношения». Pure Appl. Chem. 81 (6): 1093–1111. Дои:10.1351 / PAC-CON-08-08-12.
- ^ Сильверторн, Ди Англауб, 1948- (2016). Физиология человека: комплексный подход. Джонсон, Брюс Р., Обер, Уильям К., Обер, Клэр Э., Силверторн, Эндрю С. (седьмое изд.). [Сан-Франциско]. ISBN 9780321981226. OCLC 890107246.CS1 maint: несколько имен: список авторов (связь)
- ^ Садава Д., Хиллис Д.М., Хеллер Х.С., Беренбаум М.Р. (2011). Жизнь: Наука биологии (9-е изд.). Сан-Франциско: Фриман. С. 105–114. ISBN 978-1-4292-4646-0.
- ^ Любик А.А., Нури М., Чыонг С., Гаффари М., Адомат Х.Х., Кори Э., Кокс М.Э., Ли Н., Ганз Э.С., Йенки П., Фам С., Буттян Р. (2016). «Паракринная передача сигналов Sonic Hedgehog значительно способствует приобретенному стероидогенезу в микросреде опухоли простаты». Int. J. Рак. 140 (2): 358–369. Дои:10.1002 / ijc.30450. PMID 27672740.
- ^ Гроховски Л.Л., Сюй Х., Уайт Р.Х. (май 2006 г.). «Methanocaldococcus jannaschii использует модифицированный мевалонатный путь для биосинтеза изопентенилдифосфата». Журнал бактериологии. 188 (9): 3192–8. Дои:10.1128 / JB.188.9.3192-3198.2006. ЧВК 1447442. PMID 16621811.
- ^ Чатуфонпрасерт, Варанья; Джарукамджорн, Канокван; Эллингер, Изабелла (12.09.2018). «Физиология и патофизиология биосинтеза, транспорта и метаболизма стероидов в плаценте человека». Границы фармакологии. 9: 1027. Дои:10.3389 / fphar.2018.01027. ISSN 1663-9812. ЧВК 6144938. PMID 30258364.
- ^ а б Кузуяма Т., Сето Х. (апрель 2003 г.). «Разнообразие биосинтеза изопреновых единиц». Отчеты о натуральных продуктах. 20 (2): 171–83. Дои:10.1039 / b109860h. PMID 12735695.
- ^ Дубей В.С., Бхалла Р., Лутра Р. (сентябрь 2003 г.). «Обзор немевалонатного пути биосинтеза терпеноидов в растениях» (PDF). Журнал биологических наук. 28 (5): 637–46. Дои:10.1007 / BF02703339. PMID 14517367. Архивировано из оригинал (PDF) на 2007-04-15.
- ^ а б Schroepfer GJ (1981). «Биосинтез стеролов». Ежегодный обзор биохимии. 50: 585–621. Дои:10.1146 / annurev.bi.50.070181.003101. PMID 7023367.
- ^ Лис Н.Д., Скэггс Б., Кирш Д.Р., Бард М. (март 1995 г.). «Клонирование поздних генов пути биосинтеза эргостерола Saccharomyces cerevisiae - обзор». Липиды. 30 (3): 221–6. Дои:10.1007 / BF02537824. PMID 7791529.
- ^ Kones R (декабрь 2010 г.). «Розувастатин, воспаление, С-реактивный белок, ЮПИТЕР и первичная профилактика сердечно-сосудистых заболеваний - перспективы». Дизайн, разработка и терапия лекарств. 4: 383–413. Дои:10.2147 / DDDT.S10812. ЧВК 3023269. PMID 21267417.
- ^ Рулофс А.Дж., Томпсон К., Гордон С., Роджерс М.Дж. (октябрь 2006 г.). «Молекулярные механизмы действия бисфосфонатов: современное состояние». Клинические исследования рака. 12 (20, часть 2): 6222с – 6230с. Дои:10.1158 / 1078-0432.CCR-06-0843. PMID 17062705.
- ^ Хэггстрём М, Ричфилд Д. (2014). «Схема путей стероидогенеза человека». WikiJournal of Медицина. 1 (1). Дои:10.15347 / wjm / 2014.005. ISSN 2002-4436.
- ^ Ханукоглу I (декабрь 1992 г.). «Стероидогенные ферменты: структура, функция и роль в регуляции биосинтеза стероидных гормонов». Журнал стероидной биохимии и молекулярной биологии. 43 (8): 779–804. Дои:10.1016/0960-0760(92)90307-5. PMID 22217824.
- ^ а б c d е ж Миллер WL, Auchus RJ (февраль 2011 г.). «Молекулярная биология, биохимия и физиология стероидогенеза человека и его нарушений». Эндокринные обзоры. 32 (1): 81–151. Дои:10.1210 / er.2010-0013. ЧВК 3365799. PMID 21051590.
- ^ Россье MF (август 2006 г.). «Т-каналы и биосинтез стероидов: в поисках связи с митохондриями». Клеточный кальций. 40 (2): 155–64. Дои:10.1016 / j.ceca.2006.04.020. PMID 16759697.
- ^ Любик А.А., Нури М., Чыонг С., Гаффари М., Адомат Х.Х., Кори Э., Кокс М.Э., Ли Н., Ганз Э.С., Йенки П., Фам С., Буттян Р. (2016). «Паракринная передача сигналов Sonic Hedgehog значительно способствует приобретенному стероидогенезу в микросреде опухоли простаты». Международный журнал рака. 140 (2): 358–369. Дои:10.1002 / ijc.30450. PMID 27672740.[неосновной источник необходим ]
- ^ Lichtenthaler HK (июнь 1999 г.). «1-дезокси-d-ксилулозо-5-фосфатный путь биосинтеза изопреноидов в растениях». Ежегодный обзор физиологии растений и молекулярной биологии растений. 50: 47–65. Дои:10.1146 / annurev.arplant.50.1.47. PMID 15012203.
- ^ Витчел С.Ф., Аззиз Р. (2010). «Неклассическая врожденная гиперплазия надпочечников». Международный журнал детской эндокринологии. 2010: 1–11. Дои:10.1155/2010/625105. ЧВК 2910408. PMID 20671993.
- ^ Пикулева И.А. (декабрь 2006 г.). «Цитохром P450s и гомеостаз холестерина». Фармакология и терапия. 112 (3): 761–73. Дои:10.1016 / j.pharmthera.2006.05.014. PMID 16872679.
- ^ Zollner G, Marschall HU, Wagner M, Trauner M (2006). «Роль ядерных рецепторов в адаптивном ответе на желчные кислоты и холестаз: патогенетические и терапевтические соображения». Молекулярная фармацевтика. 3 (3): 231–51. Дои:10.1021 / mp060010s. PMID 16749856.
- ^ Кливер С.А., Гудвин Б., Уилсон TM (октябрь 2002 г.). «Ядерный рецептор прегнана X: ключевой регулятор метаболизма ксенобиотиков». Эндокринные обзоры. 23 (5): 687–702. Дои:10.1210 / er.2001-0038. PMID 12372848.
- ^ Штаймер Т. «Метаболизм стероидных гормонов». Сотрудничающий центр ВОЗ в области образования и исследований в области репродукции человека. Женевский фонд медицинского образования и исследований.
- ^ а б «Рассел Маркер, создатель мексиканской индустрии стероидных гормонов». Международная историческая химическая достопримечательность. Американское химическое общество.
- ^ а б Макин Х.Л., Хонор JW, Шеклтон СН, Гриффитс WJ (2010). «Общие методы экстракции, очистки и измерения стероидов с помощью хроматографии и масс-спектрометрии». В Makin HL, Gower DB (ред.). Стероидный анализ. Дордрехт; Нью-Йорк: Спрингер. С. 163–282. ISBN 978-1-4020-9774-4.
- ^ Коннер А.Х., Нагаока М., Роу Дж. У., Перлман Д. (август 1976 г.). «Микробное преобразование стеролов таллового масла в стероиды C19» (PDF). Прикладная и экологическая микробиология. 32 (2): 310–1. ЧВК 170056. PMID 987752.
- ^ а б Hesselink PG, Vliet Sv, Vries Hd, Witholt B (1989). "Оптимизация расщепления боковой цепи стероидов путем Mycobacterium sp. в присутствии циклодекстринов ». Ферментные и микробные технологии. 11 (7): 398–404. Дои:10.1016/0141-0229(89)90133-6.
- ^ а б c Сандов Дж., Шайффеле Э., Харинг М., Ниф Дж., Прежевовски К., Стах У. (2000). Гормоны. Энциклопедия промышленной химии Ульмана. Дои:10.1002 / 14356007.a13_089. ISBN 978-3527306732.
- ^ Ли HM, Meister PD, Weintraub A, Reineke LM, Eppstein SH, Murray HC, Peterson DH (1952). «Микробиологические превращения стероидов.1 I. Введение кислорода в углерод-11 прогестерона». Журнал Американского химического общества. 73 (23): 5933–5936. Дои:10.1021 / ja01143a033.
- ^ Чапек М, Олдрих Х, Алоис С (1966). Микробные превращения стероидов. Прага: Издательство Academia Чехословацкой Академии наук. Дои:10.1007/978-94-011-7603-3. ISBN 9789401176057.
- ^ Маркер RE, Rohrmann E (1939). «Стерины. LXXXI. Превращение сарсаса-погенина в прегнанедиал - 3 (α), 20 (α)». Журнал Американского химического общества. 61 (12): 3592–3593. Дои:10.1021 / ja01267a513.
- ^ "Нобелевская премия по химии 1927 г.". Нобелевский фонд.
- ^ "Нобелевская премия по химии 1928 г.". Нобелевский фонд.
- ^ "Нобелевская премия по химии 1939 г.". Нобелевский фонд.
- ^ "Нобелевская премия по физиологии и медицине 1950 г.". Нобелевский фонд.
- ^ "Нобелевская премия по химии 1965 г.". Нобелевский фонд.
- ^ "Нобелевская премия по химии 1969 г.". Нобелевский фонд.
- ^ «Нобелевская премия по химии 1975 года». Нобелевский фонд.
Библиография
- Рассел CA (2005). «Органическая химия: натуральные продукты, стероиды». В Russell CA, Roberts GK (ред.). Химическая история: обзоры новейшей литературы. Кембридж: RSC Publ. ISBN 978-0-85404-464-1.
- "Рассел Маркер: создание мексиканской индустрии стероидных гормонов - ориентир -". Американское химическое общество. 1999 г.
- Ледничер Д. (2011). Краткий обзор химии стероидов. Хобокен: Вайли. Дои:10.1002/9780470973639. ISBN 978-0-470-66085-0. Краткая история изучения стероидов.
- Йодер Р.А., Джонстон Дж. Н. (декабрь 2005 г.). «Пример полного биомиметического синтеза: карбоциклизация полиолефинов в терпены и стероиды». Химические обзоры. 105 (12): 4730–56. Дои:10.1021 / cr040623l. ЧВК 2575671. PMID 16351060. Обзор истории синтеза стероидов, особенно биомиметик.
- Хан Т.С., Уокер Б.Р., Арльт У., Росс Р.Дж. (февраль 2014 г.). «Лечение и результаты здоровья взрослых с врожденной гиперплазией надпочечников». Обзоры природы. Эндокринология. 10 (2): 115–24. Дои:10.1038 / nrendo.2013.239. PMID 24342885. Путь стероидогенеза надпочечников.
- Греп, Рой О., изд. (22 октября 2013 г.). «Кортоевые кислоты». Недавний прогресс в исследованиях гормонов: Материалы конференции по Лаврентьеву гормонам 1979 г.. Elsevier Science. С. 345–391. ISBN 978-1-4832-1956-1.
- Боуэн Р.А. (20 октября 2001 г.). «Стероидогенез». Патофизиология эндокринной системы. Государственный университет Колорадо. Архивировано из оригинал 28 февраля 2009 г.
Метаболизм карта | ||
|---|---|---|
Одиночные линии: пути, общие для большинства форм жизни. Двойные линии: пути не у человека (встречаются, например, у растений, грибов, прокариот). | ||
