Рецептор эстрогена - Estrogen receptor
| рецептор эстрогена 1 (ER-альфа) | |||||||
|---|---|---|---|---|---|---|---|
 | |||||||
| Идентификаторы | |||||||
| Символ | ESR1 | ||||||
| Альт. символы | ER-α, NR3A1 | ||||||
| Ген NCBI | 2099 | ||||||
| HGNC | 3467 | ||||||
| OMIM | 133430 | ||||||
| PDB | 1ERE | ||||||
| RefSeq | NM_000125 | ||||||
| UniProt | P03372 | ||||||
| Прочие данные | |||||||
| Locus | Chr. 6 q24-q27 | ||||||
| |||||||
| рецептор эстрогена 2 (ER-бета) | |||||||
|---|---|---|---|---|---|---|---|
 | |||||||
| Идентификаторы | |||||||
| Символ | ESR2 | ||||||
| Альт. символы | ER-β, NR3A2 | ||||||
| Ген NCBI | 2100 | ||||||
| HGNC | 3468 | ||||||
| OMIM | 601663 | ||||||
| PDB | 1QKM | ||||||
| RefSeq | NM_001040275 | ||||||
| UniProt | Q92731 | ||||||
| Прочие данные | |||||||
| Locus | Chr. 14 q21-q22 | ||||||
| |||||||
Рецепторы эстрогенов (ER) являются группой белки найдено внутри клетки. Они есть рецепторы которые активируются гормон эстроген (17β-эстрадиол ).[1] Существует два класса ER: ядерные рецепторы эстрогена (ERα и ERβ ), которые являются членами ядерный рецептор семья внутриклеточный рецепторы и мембранные рецепторы эстрогена (МСК) (GPER (GPR30), ER-X, и гq-mER ), которые в основном G-белковые рецепторы. Эта статья относится к первому (ER).
После активации эстрогеном ER может перемещать в ядро и связываются с ДНК, чтобы регулировать активность различных генов (т.е. это ДНК-связывающая фактор транскрипции ). Однако он также выполняет дополнительные функции, независимые от связывания ДНК.[2]
Так как рецепторы гормонов для половые стероиды (рецепторы стероидных гормонов ), ER, рецепторы андрогенов (AR), и рецепторы прогестерона (PR) важны в половое созревание и беременность.
Протеомика
Существует две разные формы рецептора эстрогена, обычно называемые α и β, каждый из которых кодируется отдельным геном (ESR1 и ESR2 соответственно). Гормонально-активируемые рецепторы эстрогена димеры и, поскольку две формы коэкспрессируются во многих типах клеток, рецепторы могут образовывать гомодимеры ERα (αα) или ERβ (ββ) или гетеродимеры ERαβ (αβ).[3]Рецепторы эстрогена альфа и бета демонстрируют значительную общую гомологию последовательностей, и оба состоят из пяти домены обозначены от A / B до F (перечислены от N- до C-конца; аминокислота порядковые номера относятся к человеческому ER).
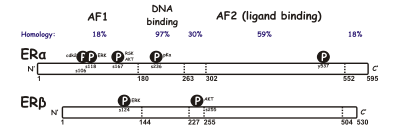
В N-концевой Домен A / B может трансактивировать транскрипция гена в отсутствие связанного лиганд (например, гормон эстроген). Хотя эта область способна активировать транскрипцию гена без лиганда, эта активация является слабой и более избирательной по сравнению с активацией, обеспечиваемой доменом E. Домен C, также известный как ДНК-связывающий домен связывается с эстрогеном элементы ответа в ДНК. Домен D - это шарнирная область, которая соединяет домены C и E. Домен E содержит полость для связывания лиганда, а также сайты связывания для коактиватор и корепрессор белки. E-домен в присутствии связанного лиганда способен активировать транскрипцию гена. В C-терминал Функция F-области не совсем ясна и имеет переменную длину.
|
| ||||||||||||||||||||||||||||||||||||||||
Известно, что из-за альтернативного сплайсинга РНК существует несколько изоформ ER. Идентифицированы по крайней мере три изоформы ERα и пять ERβ. Подтипы рецепторов изоформ ERβ могут трансактивировать транскрипцию только тогда, когда образуется гетеродимер с функциональным рецептором ERβ1 массой 59 кДа. Рецептор ERβ3 был обнаружен на высоком уровне в яичках. Две другие изоформы ERα имеют размер 36 и 46 кДа.[4][5]
Рецептор ERγ описан только у рыб, но не у человека.[6]
Генетика
У человека две формы рецептора эстрогена кодируются разными гены, ESR1 и ESR2 на шестом и четырнадцатом хромосома (6q25.1 и 14q23.2) соответственно.
Распределение
Оба ER широко экспрессируются в разных типах тканей, однако есть некоторые заметные различия в паттернах их экспрессии:[7]
- В ERα находится в эндометрий, рак молочной железы клетки, стромальные клетки яичников и гипоталамус.[8] У мужчин ERα белок находится в эпителии отводящие протоки.[9]
- Выражение ERβ белок был зарегистрирован в яичниках клетки гранулезы, почка, мозг, кость, сердце,[10] легкие, кишечный слизистая оболочка, простата, и эндотелиальный клетки.
Считается, что ER являются цитоплазматическими рецепторами в их нелигандованном состоянии, но исследования визуализации показали, что только небольшая часть ER находится в цитоплазме, при этом большая часть ER постоянно находится в ядре.[11]Первичный транскрипт «ERα» дает начало нескольким альтернативно сплайсированным вариантам неизвестной функции.[12]
Лиганды
Агонисты
- Эндогенный эстрогены (например., эстрадиол, эстрон, эстриол, эстетрол )
- Натуральный эстрогены (например., конъюгированные эстрогены )
- Синтетический эстрогены (например., этинилэстрадиол, диэтилстильбестрол )
Смешанный (механизм действия агониста и антагониста)
- Фитоэстрогены (например., куместрол, Daidzein, геништейн, мироэстрол )
- Селективные модуляторы рецепторов эстрогена (например., тамоксифен, кломифен, ралоксифен )
Антагонисты
- Антиэстрогены (например., фулвестрант, ICI-164384, этамокситрифетол )
Родство
| Лиганд | Другие имена | Относительное сродство связывания (РБА,%)а | Абсолютное связывающее сродство (Kя, нМ)а | Действие | ||
|---|---|---|---|---|---|---|
| ERα | ERβ | ERα | ERβ | |||
| Эстрадиол | E2; 17β-эстрадиол | 100 | 100 | 0.115 (0.04–0.24) | 0.15 (0.10–2.08) | Эстроген |
| Estrone | E1; 17-кетоэстрадиол | 16.39 (0.7–60) | 6.5 (1.36–52) | 0.445 (0.3–1.01) | 1.75 (0.35–9.24) | Эстроген |
| Эстриол | E3; 16α-OH-17β-E2 | 12.65 (4.03–56) | 26 (14.0–44.6) | 0.45 (0.35–1.4) | 0.7 (0.63–0.7) | Эстроген |
| Эстетрол | E4; 15α, 16α-Di-OH-17β-E2 | 4.0 | 3.0 | 4.9 | 19 | Эстроген |
| Альфатрадиол | 17α-эстрадиол | 20.5 (7–80.1) | 8.195 (2–42) | 0.2–0.52 | 0.43–1.2 | Метаболит |
| 16-эпиестриол | 16β-гидрокси-17β-эстрадиол | 7.795 (4.94–63) | 50 | ? | ? | Метаболит |
| 17-эпиестриол | 16α-гидрокси-17α-эстрадиол | 55.45 (29–103) | 79–80 | ? | ? | Метаболит |
| 16,17-эпиестриол | 16β-гидрокси-17α-эстрадиол | 1.0 | 13 | ? | ? | Метаболит |
| 2-гидроксиэстрадиол | 2-ОН-E2 | 22 (7–81) | 11–35 | 2.5 | 1.3 | Метаболит |
| 2-метоксиэстрадиол | 2-MeO-E2 | 0.0027–2.0 | 1.0 | ? | ? | Метаболит |
| 4-гидроксиэстрадиол | 4-ОН-E2 | 13 (8–70) | 7–56 | 1.0 | 1.9 | Метаболит |
| 4-метоксиэстрадиол | 4-MeO-E2 | 2.0 | 1.0 | ? | ? | Метаболит |
| 2-гидроксиэстрон | 2-ОН-E1 | 2.0–4.0 | 0.2–0.4 | ? | ? | Метаболит |
| 2-метоксиэстрон | 2-MeO-E1 | <0.001–<1 | <1 | ? | ? | Метаболит |
| 4-гидроксиэстрон | 4-ОН-E1 | 1.0–2.0 | 1.0 | ? | ? | Метаболит |
| 4-метоксиэстрон | 4-MeO-E1 | <1 | <1 | ? | ? | Метаболит |
| 16α-гидроксиэстрон | 16α-OH-E1; 17-кетоэстриол | 2.0–6.5 | 35 | ? | ? | Метаболит |
| 2-гидроксиэстриол | 2-ОН-E3 | 2.0 | 1.0 | ? | ? | Метаболит |
| 4-метоксиэстриол | 4-MeO-E3 | 1.0 | 1.0 | ? | ? | Метаболит |
| Эстрадиола сульфат | E2S; Эстрадиол 3-сульфат | <1 | <1 | ? | ? | Метаболит |
| Дисульфат эстрадиола | Эстрадиол 3,17β-дисульфат | 0.0004 | ? | ? | ? | Метаболит |
| Эстрадиол 3-глюкуронид | E2-3G | 0.0079 | ? | ? | ? | Метаболит |
| Эстрадиол 17β-глюкуронид | E2-17G | 0.0015 | ? | ? | ? | Метаболит |
| Эстрадиол 3-глюк. 17β-сульфат | Э2-3Г-17С | 0.0001 | ? | ? | ? | Метаболит |
| Эстрона сульфат | E1S; Эстрон 3-сульфат | <1 | <1 | >10 | >10 | Метаболит |
| Бензоат эстрадиола | EB; Эстрадиол 3-бензоат | 10 | ? | ? | ? | Эстроген |
| Эстрадиол 17β-бензоат | E2-17B | 11.3 | 32.6 | ? | ? | Эстроген |
| Эстрон метиловый эфир | Эстрон 3-метиловый эфир | 0.145 | ? | ? | ? | Эстроген |
| Ent-Эстрадиол | 1-эстрадиол | 1.31–12.34 | 9.44–80.07 | ? | ? | Эстроген |
| Equilin | 7-дегидроэстрон | 13 (4.0–28.9) | 13.0–49 | 0.79 | 0.36 | Эстроген |
| Эквиленин | 6,8-дидегидроэстрон | 2.0–15 | 7.0–20 | 0.64 | 0.62 | Эстроген |
| 17β-дигидроэкилин | 7-дегидро-17β-эстрадиол | 7.9–113 | 7.9–108 | 0.09 | 0.17 | Эстроген |
| 17α-дигидроэкилин | 7-дегидро-17α-эстрадиол | 18.6 (18–41) | 14–32 | 0.24 | 0.57 | Эстроген |
| 17β-дигидроэквиленин | 6,8-дидегидро-17β-эстрадиол | 35–68 | 90–100 | 0.15 | 0.20 | Эстроген |
| 17α-дигидроэквиленин | 6,8-дидегидро-17α-эстрадиол | 20 | 49 | 0.50 | 0.37 | Эстроген |
| Δ8-Эстрадиол | 8,9-дегидро-17β-эстрадиол | 68 | 72 | 0.15 | 0.25 | Эстроген |
| Δ8-Эстроне | 8,9-дегидроэстрон | 19 | 32 | 0.52 | 0.57 | Эстроген |
| Этинилэстрадиол | EE; 17α-Этинил-17β-E2 | 120.9 (68.8–480) | 44.4 (2.0–144) | 0.02–0.05 | 0.29–0.81 | Эстроген |
| Местранол | EE 3-метиловый эфир | ? | 2.5 | ? | ? | Эстроген |
| Моксестрол | RU-2858; 11β-метокси-EE | 35–43 | 5–20 | 0.5 | 2.6 | Эстроген |
| Метилэстрадиол | 17α-метил-17β-эстрадиол | 70 | 44 | ? | ? | Эстроген |
| Диэтилстильбестрол | DES; Стилбестрол | 129.5 (89.1–468) | 219.63 (61.2–295) | 0.04 | 0.05 | Эстроген |
| Гексэстрол | Дигидродиэтилстильбестрол | 153.6 (31–302) | 60–234 | 0.06 | 0.06 | Эстроген |
| Диенестрол | Дегидростильбестрол | 37 (20.4–223) | 56–404 | 0.05 | 0.03 | Эстроген |
| Бензэстрол (B2) | – | 114 | ? | ? | ? | Эстроген |
| Хлортрианизен | ТАСЕ | 1.74 | ? | 15.30 | ? | Эстроген |
| Трифенилэтилен | TPE | 0.074 | ? | ? | ? | Эстроген |
| Трифенилбромэтилен | TPBE | 2.69 | ? | ? | ? | Эстроген |
| Тамоксифен | ICI-46,474 | 3 (0.1–47) | 3.33 (0.28–6) | 3.4–9.69 | 2.5 | SERM |
| Афимоксифен | 4-гидрокситамоксифен; 4-ОНТ | 100.1 (1.7–257) | 10 (0.98–339) | 2.3 (0.1–3.61) | 0.04–4.8 | SERM |
| Торемифен | 4-хлоротамоксифен; 4-CT | ? | ? | 7.14–20.3 | 15.4 | SERM |
| Кломифен | РСЗО-41 | 25 (19.2–37.2) | 12 | 0.9 | 1.2 | SERM |
| Циклофенил | F-6066; Сексовид | 151–152 | 243 | ? | ? | SERM |
| Наоксидин | U-11,000A | 30.9–44 | 16 | 0.3 | 0.8 | SERM |
| Ралоксифен | – | 41.2 (7.8–69) | 5.34 (0.54–16) | 0.188–0.52 | 20.2 | SERM |
| Арзоксифен | LY-353,381 | ? | ? | 0.179 | ? | SERM |
| Ласофоксифен | CP-336,156 | 10.2–166 | 19.0 | 0.229 | ? | SERM |
| Ормелоксифен | Centchroman | ? | ? | 0.313 | ? | SERM |
| Левормелоксифен | 6720-CDRI; NNC-460 020 | 1.55 | 1.88 | ? | ? | SERM |
| Оспемифен | Deaminohydroxytoremifene | 2.63 | 1.22 | ? | ? | SERM |
| Базедоксифен | – | ? | ? | 0.053 | ? | SERM |
| Etacstil | GW-5638 | 4.30 | 11.5 | ? | ? | SERM |
| ICI-164,384 | – | 63.5 (3.70–97.7) | 166 | 0.2 | 0.08 | Антиэстроген |
| Фулвестрант | ICI-182,780 | 43.5 (9.4–325) | 21.65 (2.05–40.5) | 0.42 | 1.3 | Антиэстроген |
| Пропилпиразолетриол | PPT | 49 (10.0–89.1) | 0.12 | 0.40 | 92.8 | Агонист ERα |
| 16α-LE2 | 16α-лактон-17β-эстрадиол | 14.6–57 | 0.089 | 0.27 | 131 | Агонист ERα |
| 16α-Йодо-E2 | 16α-йод-17β-эстрадиол | 30.2 | 2.30 | ? | ? | Агонист ERα |
| Метилпиперидинопиразол | MPP | 11 | 0.05 | ? | ? | Антагонист ERα |
| Диарилпропионитрил | ДПН | 0.12–0.25 | 6.6–18 | 32.4 | 1.7 | Агонист ERβ |
| 8β-VE2 | 8β-винил-17β-эстрадиол | 0.35 | 22.0–83 | 12.9 | 0.50 | Агонист ERβ |
| Prinaberel | ЕРБ-041; ПУТЬ-202,041 | 0.27 | 67–72 | ? | ? | Агонист ERβ |
| ЕРБ-196 | ПУТЬ-202 196 | ? | 180 | ? | ? | Агонист ERβ |
| Эртеберел | СЕРБА-1; LY-500,307 | ? | ? | 2.68 | 0.19 | Агонист ERβ |
| СЕРБА-2 | – | ? | ? | 14.5 | 1.54 | Агонист ERβ |
| Куместрол | – | 9.225 (0.0117–94) | 64.125 (0.41–185) | 0.14–80.0 | 0.07–27.0 | Ксеноэстроген |
| Геништейн | – | 0.445 (0.0012–16) | 33.42 (0.86–87) | 2.6–126 | 0.3–12.8 | Ксеноэстроген |
| Equol | – | 0.2–0.287 | 0.85 (0.10–2.85) | ? | ? | Ксеноэстроген |
| Daidzein | – | 0.07 (0.0018–9.3) | 0.7865 (0.04–17.1) | 2.0 | 85.3 | Ксеноэстроген |
| Биоханин А | – | 0.04 (0.022–0.15) | 0.6225 (0.010–1.2) | 174 | 8.9 | Ксеноэстроген |
| Кемпферол | – | 0.07 (0.029–0.10) | 2.2 (0.002–3.00) | ? | ? | Ксеноэстроген |
| Нарингенин | – | 0.0054 (<0.001–0.01) | 0.15 (0.11–0.33) | ? | ? | Ксеноэстроген |
| 8-пренилнарингенин | 8-PN | 4.4 | ? | ? | ? | Ксеноэстроген |
| Кверцетин | – | <0.001–0.01 | 0.002–0.040 | ? | ? | Ксеноэстроген |
| Иприфлавон | – | <0.01 | <0.01 | ? | ? | Ксеноэстроген |
| Мироэстрол | – | 0.39 | ? | ? | ? | Ксеноэстроген |
| Дезоксимироэстрол | – | 2.0 | ? | ? | ? | Ксеноэстроген |
| β-ситостерин | – | <0.001–0.0875 | <0.001–0.016 | ? | ? | Ксеноэстроген |
| Ресвератрол | – | <0.001–0.0032 | ? | ? | ? | Ксеноэстроген |
| α-Зеараленол | – | 48 (13–52.5) | ? | ? | ? | Ксеноэстроген |
| β-Зеараленол | – | 0.6 (0.032–13) | ? | ? | ? | Ксеноэстроген |
| Зеранол | α-Зеараланол | 48–111 | ? | ? | ? | Ксеноэстроген |
| Талеранол | β-Зеараланол | 16 (13–17.8) | 14 | 0.8 | 0.9 | Ксеноэстроген |
| Зеараленон | ZEN | 7.68 (2.04–28) | 9.45 (2.43–31.5) | ? | ? | Ксеноэстроген |
| Зеараланон | ZAN | 0.51 | ? | ? | ? | Ксеноэстроген |
| Бисфенол А | BPA | 0.0315 (0.008–1.0) | 0.135 (0.002–4.23) | 195 | 35 | Ксеноэстроген |
| Эндосульфан | EDS | <0.001–<0.01 | <0.01 | ? | ? | Ксеноэстроген |
| Кепоне | Хлордекон | 0.0069–0.2 | ? | ? | ? | Ксеноэстроген |
| о, п '-DDT | – | 0.0073–0.4 | ? | ? | ? | Ксеноэстроген |
| п, п '-DDT | – | 0.03 | ? | ? | ? | Ксеноэстроген |
| Метоксихлор | п, п '-Диметокси-ДДТ | 0.01 (<0.001–0.02) | 0.01–0.13 | ? | ? | Ксеноэстроген |
| HPTE | Гидроксихлор; п, п '-ОН-ДДТ | 1.2–1.7 | ? | ? | ? | Ксеноэстроген |
| Тестостерон | Т; 4-Андростенолон | <0.0001–<0.01 | <0.002–0.040 | >5000 | >5000 | Андроген |
| Дигидротестостерон | DHT; 5α-Андростанолон | 0.01 (<0.001–0.05) | 0.0059–0.17 | 221–>5000 | 73–1688 | Андроген |
| Нандролон | 19-нортестостерон; 19-NT | 0.01 | 0.23 | 765 | 53 | Андроген |
| Дегидроэпиандростерон | DHEA; Прастерон | 0.038 (<0.001–0.04) | 0.019–0.07 | 245–1053 | 163–515 | Андроген |
| 5-Андростендиол | A5; Андростендиол | 6 | 17 | 3.6 | 0.9 | Андроген |
| 4-Андростендиол | – | 0.5 | 0.6 | 23 | 19 | Андроген |
| 4-Андростендион | A4; Андростендион | <0.01 | <0.01 | >10000 | >10000 | Андроген |
| 3α-Андростандиол | 3α-Адиол | 0.07 | 0.3 | 260 | 48 | Андроген |
| 3β-Андростандиол | 3β-Адиол | 3 | 7 | 6 | 2 | Андроген |
| Андростандион | 5α-Андростандион | <0.01 | <0.01 | >10000 | >10000 | Андроген |
| Этиохоландион | 5β-Андростандион | <0.01 | <0.01 | >10000 | >10000 | Андроген |
| Метилтестостерон | 17α-метилтестостерон | <0.0001 | ? | ? | ? | Андроген |
| Этинил-3α-андростандиол | 17α-этинил-3α-адиол | 4.0 | <0.07 | ? | ? | Эстроген |
| Этинил-3β-андростандиол | 17α-этинил-3β-адиол | 50 | 5.6 | ? | ? | Эстроген |
| Прогестерон | P4; 4-прегненедион | <0.001–0.6 | <0.001–0.010 | ? | ? | Прогестаген |
| Норэтистерон | СЕТЬ; 17α-этинил-19-NT | 0.085 (0.0015–<0.1) | 0.1 (0.01–0.3) | 152 | 1084 | Прогестаген |
| Норэтинодрел | 5 (10) -норэтистерон | 0.5 (0.3–0.7) | <0.1–0.22 | 14 | 53 | Прогестаген |
| Тиболон | 7α-метилноретинодрел | 0.5 (0.45–2.0) | 0.2–0.076 | ? | ? | Прогестаген |
| Δ4-Тиболон | 7α-метилноэтистерон | 0.069–<0.1 | 0.027–<0.1 | ? | ? | Прогестаген |
| 3α-гидрокситиболон | – | 2.5 (1.06–5.0) | 0.6–0.8 | ? | ? | Прогестаген |
| 3β-гидрокситиболон | – | 1.6 (0.75–1.9) | 0.070–0.1 | ? | ? | Прогестаген |
| Сноски: а = (1) Связывание сродства значения имеют формат «медиана (диапазон)» (# (# - #)), «диапазон» (# - #) или «значение» (#) в зависимости от доступных значений. Полные наборы значений в пределах диапазонов можно найти в коде Wiki. (2) Аффинность связывания определяли с помощью исследований замещения в различных in vitro системы с маркированный эстрадиол и человек ERα и ERβ белки (кроме значений ERβ из Kuiper et al. (1997), которые представляют собой ERβ крысы). Источники: См. Страницу шаблона. | ||||||
Связывание и функциональная избирательность
Домен спирали 12 ER играет решающую роль в определении взаимодействий с коактиваторами и корепрессорами и, следовательно, в соответствующем агонистическом или антагонистическом эффекте лиганда.[13][14]
Другой лиганды могут различаться по своему сродству к альфа- и бета-изоформам рецептора эстрогена:
- эстрадиол одинаково хорошо связывается с обоими рецепторами[15]
- эстрон, и ралоксифен связываются преимущественно с альфа-рецептором[15]
- эстриол, и геништейн к бета-рецептору[15]
Подтип селективные модуляторы рецепторов эстрогена предпочтительно связываются либо с α-, либо с β-подтипом рецептора. Кроме того, различные комбинации рецепторов эстрогена могут по-разному реагировать на различные лиганды, что может проявляться в тканевых агонистических и антагонистических эффектах.[16] Было высказано предположение, что соотношение концентраций α- и β-подтипа играет роль при определенных заболеваниях.[17]
Концепция чего-либо селективные модуляторы рецепторов эстрогена основан на способности стимулировать взаимодействия ER с различными белками, такими как транскрипционный коактиватор или корепрессоры. Кроме того, соотношение коактиватора и корепрессорного белка варьируется в разных тканях.[18] Как следствие, один и тот же лиганд может быть агонистом в некоторых тканях (где преобладают коактиваторы), а в других тканях (где преобладают корепрессоры) - антагонистами. Тамоксифен, например, является антагонистом грудь и поэтому используется как рак молочной железы лечение[19] но агонист ER в кость (тем самым предотвращая остеопороз ) и частичный агонист в эндометрий (увеличивая риск рак матки ).
Передача сигнала
Поскольку эстроген - это стероидный гормон, он может проходить через фосфолипидные мембраны клетки, и рецепторы, следовательно, не должны быть связаны с мембраной для связывания с эстрогеном.
Геномный
В отсутствие гормона рецепторы эстрогена в основном расположены в цитозоле. Связывание гормона с рецептором запускает ряд событий, начиная с миграции рецептора из цитозоля в ядро, димеризации рецептора и последующего связывания димера рецептора со специфическими последовательностями ДНК, известными как элементы гормонального ответа. Затем комплекс ДНК / рецептор привлекает другие белки, которые отвечают за транскрипция нисходящей ДНК в мРНК и, наконец, в белок, который приводит к изменению функции клетки. Рецепторы эстрогенов также встречаются в ядро клетки, и оба подтипа рецепторов эстрогена имеют ДНК переплет домен и может функционировать как факторы транскрипции регулировать производство белки.
Рецептор также взаимодействует с протеин-активатор 1 и Сп-1 для содействия транскрипции с помощью нескольких коактиваторов, таких как ПЕЛП-1.[2]
Прямое ацетилирование рецептора эстрогена альфа по остаткам лизина в шарнирной области с помощью p300 регулирует трансактивацию и чувствительность к гормонам.[20]
Негеномный
Некоторые рецепторы эстрогенов связаны с мембрана клеточной поверхности и может быстро активироваться при воздействии на клетки эстрогена.[21][22]
Кроме того, некоторые ЭПР могут связываться с клеточными мембранами путем прикрепления к кавеолин-1 и образуют комплексы с G белки, стриатин, рецептор тирозинкиназы (например., EGFR и IGF-1 ) и нерецепторные тирозинкиназы (например, Src ).[2][21] Через стриатин некоторые из этих мембраносвязанных ЭР могут приводить к повышению уровня Ca2+ и оксид азота (НЕТ).[23] Через рецепторные тирозинкиназы сигналы передаются в ядро через митоген-активированная протеинкиназа (MAPK / ERK) путь и фосфоинозитид-3-киназа (Pl3K /AKT ) путь.[24] Гликогенсинтаза киназа-3 (GSK) -3β ингибирует транскрипцию ядерным ER путем ингибирования фосфорилирование из серин 118 ядерной ERα. Фосфорилирование GSK-3β устраняет его ингибирующий эффект, и это может быть достигнуто с помощью пути PI3K / AKT и пути MAPK / ERK через rsk.
Было показано, что 17β-эстрадиол активирует Рецептор, связанный с G-белком GPR30.[25] Однако субклеточная локализация и роль этого рецептора до сих пор остаются предметом споров.[26]
Болезнь


Рак
Рецепторы эстрогена чрезмерно экспрессируются примерно в 70% случаев. рак молочной железы случаи, называемые «ER-положительными», и могут быть продемонстрированы в таких тканях с использованием иммуногистохимия. Были предложены две гипотезы, объясняющие, почему это вызывает туморогенез, и имеющиеся данные свидетельствуют о том, что оба механизма способствуют:
- Во-первых, связывание эстрогена с ЭПР стимулирует пролиферацию клетки молочной железы, что приводит к увеличению деление клеток и Репликация ДНК, что приводит к мутациям.
- Во-вторых, метаболизм эстрогенов производит генотоксичный трата.
Результатом обоих процессов является нарушение клеточный цикл, апоптоз и Ремонт ДНК, что увеличивает вероятность образования опухоли. ERα определенно связан с более дифференцированными опухолями, в то время как доказательства участия ERβ спорны. Различные версии ESR1 ген были идентифицированы (с однонуклеотидные полиморфизмы ) и связаны с различными рисками развития рака груди.[19]
Эстроген и ЭР также участвуют в рак молочной железы, рак яичников, рак толстой кишки, рак простаты, и рак эндометрия. Распространенный рак толстой кишки связан с потерей ERβ, преобладающего ER в ткани толстой кишки, и рак толстой кишки лечится с помощью ERβ-специфических агонистов.[27]
Эндокринный терапия рака груди включает селективные модуляторы рецепторов эстрогена (SERMS), например тамоксифен, которые действуют как антагонисты ER в ткани груди, или ингибиторы ароматазы, такие как анастрозол. Статус ER используется для определения чувствительности рак молочной железы поражения тамоксифеном и ингибиторами ароматазы.[28] Еще один SERM, ралоксифен, использовался в качестве профилактической химиотерапии для женщин с высоким риском развития рака груди.[29] Еще один химиотерапевтический антиэстроген, ICI 182 780 (Faslodex), который действует как полный антагонист, также способствует деградации рецептора эстрогена.
Однако, de novo устойчивость к эндокринной терапии подрывает эффективность использования конкурентных ингибиторов, таких как тамоксифен. Гормональная депривация с помощью ингибиторов ароматазы также оказывается бесполезной.[30] Массивно параллельное секвенирование генома выявило общее присутствие точечных мутаций на ESR1 которые являются драйверами устойчивости и способствуют конформации агониста ERα без ограничения лиганд. Такая конститутивная, независимая от эстрогена активность обусловлена специфическими мутациями, такими как мутации D538G или Y537S / C / N, в лиганд-связывающем домене ESR1 и способствуют пролиферации клеток и прогрессированию опухолей без гормональной стимуляции.[31]
Менопауза
Метаболические эффекты эстрогена у женщин в постменопаузе были связаны с генетическим полиморфизмом рецептор эстрогена бета (ER-β).[32]
Старение
Исследования на самках мышей показали, что уровень рецептора эстрогена-альфа снижается в преоптической области. гипоталамус как они стареют. Самки мышей, которым давали калорийность ограничена диета на протяжении большей части жизни поддерживала более высокие уровни ERα в преоптическом гипоталамусе, чем их коллеги без ограничения калорийности.[8]
Ожирение
Драматическая демонстрация важности эстрогенов в регулировании отложения жира происходит от трансгенные мыши которые были генетически сконструированы так, чтобы не иметь функциональных ароматаза ген. У этих мышей очень низкий уровень эстрогена и они страдают ожирением.[33] Ожирение также наблюдалось у самок мышей с дефицитом эстрогена, лишенных рецептор фолликулостимулирующего гормона.[34] Влияние низкого уровня эстрогена на увеличение ожирения было связано с альфа-рецептором эстрогена.[35]
Открытие
Рецепторы эстрогенов были впервые идентифицированы Элвуд В. Дженсен на Чикагский университет в 1958 г.,[36][37] за что Дженсен был награжден Премия Ласкера.[38] Ген второго рецептора эстрогена (ERβ) был идентифицирован в 1996 году Kuiper et al. в простате и яичниках крыс с использованием вырожденных праймеров ERalpha.[39]
Смотрите также
- Мембранный рецептор эстрогена
- Синдром нечувствительности к эстрогенам
- Дефицит ароматазы
- Синдром избытка ароматазы
использованная литература
- ^ Dahlman-Wright K, Cavailles V, Fuqua SA, Jordan VC, Katzenellenbogen JA, Korach KS, Maggi A, Muramatsu M, Parker MG, Gustafsson JA (декабрь 2006 г.). «Международный фармакологический союз. LXIV. Рецепторы эстрогенов». Фармакологические обзоры. 58 (4): 773–81. Дои:10.1124 / пр.58.4.8. PMID 17132854. S2CID 45996586.
- ^ а б c Левин Е.Р. (август 2005 г.). «Интеграция внеядерного и ядерного действия эстрогена». Молекулярная эндокринология. 19 (8): 1951–9. Дои:10.1210 / я.2004-0390. ЧВК 1249516. PMID 15705661.
- ^ Ли X, Хуанг Дж, Йи П, Бамбара Р.А., Хильф Р., Муян М. (сентябрь 2004 г.). «Одноцепочечные рецепторы эстрогена (ER) показывают, что гетеродимер ERalpha / beta имитирует функции димера ERalpha в геномных путях передачи сигналов эстрогена». Молекулярная и клеточная биология. 24 (17): 7681–94. Дои:10.1128 / MCB.24.17.7681-7694.2004. ЧВК 506997. PMID 15314175.
- ^ Nilsson S, Mäkelä S, Treuter E, Tujague M, Thomsen J, Andersson G, Enmark E, Pettersson K, Warner M, Gustafsson JA (октябрь 2001 г.). «Механизмы действия эстрогенов». Физиологические обзоры. 81 (4): 1535–65. Дои:10.1152 / Physrev.2001.81.4.1535. PMID 11581496.
- ^ Люнг Ю.К., Мак П., Хассан С., Хо С.М. (август 2006 г.). «Изоформы рецептора эстрогена (ER) -бета: ключ к пониманию передачи сигналов ER-бета». Труды Национальной академии наук Соединенных Штатов Америки. 103 (35): 13162–7. Дои:10.1073 / pnas.0605676103. ЧВК 1552044. PMID 16938840.
- ^ Хокинс МБ, Торнтон Дж. У., Бригады D, Шкипер Дж. К., Дотт А., Томас П. (сентябрь 2000 г.). «Идентификация третьего отдельного рецептора эстрогена и реклассификация рецепторов эстрогена у костистых». Труды Национальной академии наук Соединенных Штатов Америки. 97 (20): 10751–6. Дои:10.1073 / pnas.97.20.10751. ЧВК 27095. PMID 11005855.
- ^ Couse JF, Lindzey J, Grandien K, Gustafsson JA, Korach KS (ноябрь 1997 г.). «Распределение в тканях и количественный анализ рибонуклеиновой кислоты-мессенджера рецептора эстрогена-альфа (ERalpha) и рецептора эстрогена-бета (ERbeta) у мышей дикого типа и мышей с нокаутом ERalpha». Эндокринология. 138 (11): 4613–21. Дои:10.1210 / en.138.11.4613. PMID 9348186.
- ^ а б Ягмайе Ф, Саид О, Гаран С.А., Фрейтаг В., Тимирас П.С., Штернберг Х. (июнь 2005 г.). «Ограничение калорийности снижает потерю клеток и поддерживает иммунореактивность рецептора эстрогена-альфа в преоптическом гипоталамусе самок мышей B6D2F1» (PDF). Письма о нейроэндокринологии. 26 (3): 197–203. PMID 15990721.
- ^ Hess RA (июль 2003 г.). «Эстроген в половом тракте взрослого мужчины: обзор». Репродуктивная биология и эндокринология. 1 (52): 52. Дои:10.1186/1477-7827-1-52. ЧВК 179885. PMID 12904263.
- ^ Бабикер Ф.А., Де Виндт Л.Дж., ван Эйкельс М., Гроэ С., Мейер Р., Довенданс ПА (февраль 2002 г.). «Действие эстрогенного гормона в сердце: регуляторная сеть и функция». Сердечно-сосудистые исследования. 53 (3): 709–19. Дои:10.1016 / S0008-6363 (01) 00526-0. PMID 11861041.
- ^ Htun H, Holth LT, Walker D, Davie JR, Hager GL (февраль 1999 г.). «Прямая визуализация альфа-рецептора эстрогена человека показывает роль лиганда в ядерном распределении рецептора». Молекулярная биология клетки. 10 (2): 471–86. Дои:10.1091 / mbc.10.2.471. ЧВК 25181. PMID 9950689.
- ^ Пфеффер У., Фекаротта Э., Видали Г (май 1995 г.). «Коэкспрессия множества мессенджерных РНК вариантов рецептора эстрогена в нормальных и неопластических тканях груди и в клетках MCF-7». Исследования рака. 55 (10): 2158–65. PMID 7743517.
- ^ Асчензи П., Бочеди А., Марино М. (август 2006 г.). «Взаимосвязь структуры и функции рецепторов эстрогена альфа и бета: влияние на здоровье человека». Молекулярные аспекты медицины. 27 (4): 299–402. Дои:10.1016 / j.mam.2006.07.001. PMID 16914190.
- ^ Bourguet W, Germain P, Gronemeyer H (октябрь 2000 г.). «Ядерные рецепторные лиганд-связывающие домены: трехмерные структуры, молекулярные взаимодействия и фармакологические последствия». Тенденции в фармакологических науках. 21 (10): 381–8. Дои:10.1016 / S0165-6147 (00) 01548-0. PMID 11050318.
- ^ а б c Zhu BT, Han GZ, Shim JY, Wen Y, Jiang XR (сентябрь 2006 г.). «Количественное соотношение структура-активность различных эндогенных метаболитов эстрогена для альфа- и бета-подтипов рецепторов эстрогена человека: понимание структурных детерминант, способствующих связыванию различных подтипов». Эндокринология. 147 (9): 4132–50. Дои:10.1210 / en.2006-0113. PMID 16728493.
- ^ Кансра С., Ямагата С., Снид Л., Фостер Л., Бен-Джонатан Н. (июль 2005 г.). «Дифференциальные эффекты антагонистов рецепторов эстрогена на пролиферацию лактотрофов гипофиза и высвобождение пролактина». Молекулярная и клеточная эндокринология. 239 (1–2): 27–36. Дои:10.1016 / j.mce.2005.04.008. PMID 15950373. S2CID 42052008.
- ^ Бакас П., Лиапис А., Влахопулос С., Гинер М., Логотети С., Креатсас Г., Мелигова А.К., Алексис М.Н., Зумпурлис В. (ноябрь 2008 г.). «Рецепторы эстрогена альфа и бета в миоме матки: основа измененной чувствительности к эстрогену». Фертильность и бесплодие. 90 (5): 1878–85. Дои:10.1016 / j.fertnstert.2007.09.019. PMID 18166184.
- ^ Шан Й., Браун М. (март 2002 г.). «Молекулярные детерминанты тканевой специфичности SERM». Наука. 295 (5564): 2465–8. Дои:10.1126 / science.1068537. PMID 11923541. S2CID 30634073.
- ^ а б Деру Б.Дж., Корах К.С. (март 2006 г.). «Рецепторы эстрогенов и болезни человека». Журнал клинических исследований. 116 (3): 561–70. Дои:10.1172 / JCI27987. ЧВК 2373424. PMID 16511588.
- ^ Ван С., Фу М., Анджелетти Р., Сиконольфи-Баез Л., Ройтенс А.Т., Альбанезе С., Лисанти М.П., Катценелленбоген Б.С., Като С., Хопп Т., Фукуа С.А., Лопес Г.Н., Кушнер П.Дж., Пестелл Р.Г. (май 2001 г.). «Прямое ацетилирование шарнирной области альфа рецептора эстрогена с помощью p300 регулирует трансактивацию и чувствительность к гормонам». Журнал биологической химии. 276 (21): 18375–83. Дои:10.1074 / jbc.M100800200. PMID 11279135.
- ^ а б Зивадинович Д., Гаметчу Б., Уотсон К.С. (2005). «Уровни мембранного рецептора-альфа эстрогена в клетках рака молочной железы MCF-7 позволяют прогнозировать реакцию на цАМФ и пролиферацию». Исследование рака груди. 7 (1): R101–12. Дои:10.1186 / bcr958. ЧВК 1064104. PMID 15642158.
- ^ Björnström L, Sjöberg M (июнь 2004 г.). «Зависимая от рецептора эстрогена активация AP-1 через негеномную передачу сигналов». Ядерный рецептор. 2 (1): 3. Дои:10.1186/1478-1336-2-3. ЧВК 434532. PMID 15196329.
- ^ Лу Кью, Паллас, округ Колумбия, Суркс, Гонконг, Баур В.Е., Мендельсон, М.Э., Карас, Р.Х. (декабрь 2004 г.). «Стриатин собирает мембранный сигнальный комплекс, необходимый для быстрой негеномной активации эндотелиальной NO-синтазы альфа-рецептором эстрогена». Труды Национальной академии наук Соединенных Штатов Америки. 101 (49): 17126–31. Дои:10.1073 / pnas.0407492101. ЧВК 534607. PMID 15569929.
- ^ Като С., Эндох Х., Масухиро Й., Китамото Т., Учияма С., Сасаки Х., Масусигэ С., Гото Й., Нисида Е., Кавасима Х., Мецгер Д., Чамбон П. (декабрь 1995 г.) «Активация рецептора эстрогена посредством фосфорилирования митоген-активированной протеинкиназой». Наука. 270 (5241): 1491–4. Дои:10.1126 / science.270.5241.1491. PMID 7491495. S2CID 4662264.
- ^ Prossnitz ER, Arterburn JB, Sklar LA (февраль 2007 г.). «GPR30: рецептор эстрогена, связанный с G-белком». Молекулярная и клеточная эндокринология. 265-266: 138–42. Дои:10.1016 / j.mce.2006.12.010. ЧВК 1847610. PMID 17222505.
- ^ Отто С., Роде-Шульц Б., Шварц Г., Фукс И., Клевер М., Бриттен Д., Лангер Г., Бадер Б., Прелле К., Нуббемейер Р., Фрицемайер К. Х. (октябрь 2008 г.). «Рецептор 30, связанный с G-белком, находится в эндоплазматическом ретикулуме и не активируется эстрадиолом». Эндокринология. 149 (10): 4846–56. Дои:10.1210 / en.2008-0269. PMID 18566127.
- ^ Харрис HA, Альберт LM, Leathurby Y, Malamas MS, Mewshaw RE, Miller CP, Kharode YP, Marzolf J, Komm BS, Winneker RC, Frail DE, Henderson RA, Zhu Y, Keith JC (октябрь 2003 г.). «Оценка агониста рецептора эстрогена-бета на животных моделях болезни человека». Эндокринология. 144 (10): 4241–9. Дои:10.1210 / en.2003-0550. PMID 14500559.
- ^ Клемонс М., Дэнсон С., Хауэлл А. (август 2002 г.). «Тамоксифен (« Нолвадекс »): обзор». Отзывы о лечении рака. 28 (4): 165–80. Дои:10.1016 / s0305-7372 (02) 00036-1. PMID 12363457.
- ^ Fabian CJ, Kimler BF (март 2005 г.). «Селективные модуляторы эстрогеновых рецепторов для первичной профилактики рака груди». Журнал клинической онкологии. 23 (8): 1644–55. Дои:10.1200 / JCO.2005.11.005. PMID 15755972.
- ^ Oesterreich S, Davidson NE (декабрь 2013 г.). «Поиск мутаций ESR1 при раке груди». Природа Генетика. 45 (12): 1415–6. Дои:10,1038 / нг.2831. ЧВК 4934882. PMID 24270445.
- ^ Ли С., Шен Д., Шао Дж., Краудер Р., Лю В., Прат А. и др. (Сентябрь 2013 г.). «Устойчивые к эндокринной терапии варианты ESR1, выявленные путем геномной характеристики ксенотрансплантатов, полученных из рака груди». Отчеты по ячейкам. 4 (6): 1116–30. Дои:10.1016 / j.celrep.2013.08.022. ЧВК 3881975. PMID 24055055.
- ^ Дараби М., Ани М., Панджепур М., Раббани М., Мовахедян А., Зареан Э (2011). «Влияние полиморфизма рецептора эстрогена β A1730G на ответ экспрессии гена ABCA1 на заместительную гормональную терапию в постменопаузе». Генетическое тестирование и молекулярные биомаркеры. 15 (1–2): 11–5. Дои:10.1089 / gtmb.2010.0106. PMID 21117950.
- ^ Hewitt KN, Boon WC, Murata Y, Jones ME, Simpson ER (сентябрь 2003 г.). «Мышь с нокаутом ароматазы демонстрирует сексуально диморфное нарушение гомеостаза холестерина». Эндокринология. 144 (9): 3895–903. Дои:10.1210 / en.2003-0244. PMID 12933663.
- ^ Данилович Н., Бабу П.С., Син В., Гердес М., Кришнамурти Н., Сайрам М.Р. (ноябрь 2000 г.). «Дефицит эстрогена, ожирение и аномалии скелета у самок мышей с нокаутом рецепторов фолликулостимулирующего гормона (FORKO)». Эндокринология. 141 (11): 4295–308. Дои:10.1210 / en.141.11.4295. PMID 11089565.
- ^ Олссон С., Хеллберг Н., Парини П., Видал О, Бохлули-Й. М., Боллули М., Рудлинг М., Линдберг М.К., Уорнер М., Анжелин Б., Густафссон Дж. А. (ноябрь 2000 г.). «Ожирение и нарушенный профиль липопротеинов у самцов мышей с дефицитом рецептора эстрогена альфа». Сообщения о биохимических и биофизических исследованиях. 278 (3): 640–5. Дои:10.1006 / bbrc.2000.3827. PMID 11095962.
- ^ Дженсен Э.В., Иордания В.К. (июнь 2003 г.). «Рецептор эстрогена: модель для молекулярной медицины» (Абстрактные). Клинические исследования рака. 9 (6): 1980–9. PMID 12796359.
- ^ Дженсен Э (2011). «Разговор с Элвудом Дженсеном. Интервью Дэвида Д. Мура». Ежегодный обзор физиологии. 74: 1–11. Дои:10.1146 / annurev-physicol-020911-153327. PMID 21888507.
- ^ Дэвид Брейси, 2004 г. "Ученый Калифорнийского университета получил американскую Нобелевскую премию за исследования. »Пресс-релиз Университета Цинциннати.
- ^ Койпер Г.Г., Энмарк Э., Пелто-Хукко М., Нильссон С., Густафссон Я.А. (июнь 1996 г.). «Клонирование нового рецептора, экспрессируемого в простате и яичнике крыс». Труды Национальной академии наук Соединенных Штатов Америки. 93 (12): 5925–30. Дои:10.1073 / пнас.93.12.5925. ЧВК 39164. PMID 8650195.
внешние ссылки
- Рецепторы эстрогена в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)
- Дэвид С. Гудселл (01.09.2003). «Рецептор эстрогена». Банк данных белков, Исследовательское сотрудничество в области структурной биоинформатики (RCSB). Получено 2008-03-15.