Антиоксидант - Antioxidant

Антиоксиданты находятся соединения которые препятствуют окисление. Окисление это химическая реакция что может произвести свободные радикалы, тем самым приводя к цепные реакции это может повредить клетки организмов. Антиоксиданты, такие как тиолы или же аскорбиновая кислота (витамин С) прекращают эти цепные реакции. Чтобы сбалансировать окислительный стресс, растения и животные поддерживают сложные системы перекрывающихся антиоксидантов, таких как глутатион и ферменты (например., каталаза и супероксиддисмутаза ), произведенные внутри, или диетический антиоксиданты Витамин С и витамин Е.
Термин «антиоксидант» в основном используется для двух совершенно разных групп веществ: промышленные химикаты которые добавляются в продукты для предотвращения окисления, а также встречающиеся в природе соединения, которые присутствуют в продуктах питания и ткань. Первые, промышленные антиоксиданты, имеют разнообразное применение: действуют как консерванты в еде и косметике, а также окисление -ингибиторы в топливах.[1]
Антиоксидант пищевые добавки не было показано, что они улучшают здоровье людей или эффективны в предотвращении болезней.[2] Добавки бета-каротин, витамин А, а витамин Е не оказывает положительного воздействия на смертность[3][4] или же рак риск.[5][нуждается в обновлении ][6] Дополнительно добавление селен или витамин Е не снижает риск сердечно-сосудистые заболевания.[7][8]
Влияние на здоровье
Отношение к диете
Хотя определенные уровни антиоксиданта витамины в диете необходимы для хорошего здоровья, до сих пор ведутся серьезные споры о том, обладают ли богатые антиоксидантами продукты или добавки противозачаточной активностью. Более того, если они действительно полезны, неизвестно, какие антиоксиданты полезны для здоровья в рационе и в каких количествах сверх обычного рациона.[9][10][11] Некоторые авторы оспаривают гипотезу о том, что витамины-антиоксиданты могут предотвращать хронические заболевания,[9][12] а другие заявляют, что гипотеза недоказана и ошибочна.[13] Полифенолы, обладающие антиоксидантными свойствами in vitro, обладают неизвестной антиоксидантной активностью in vivo из-за обширного метаболизм после пищеварения и немного клинические данные эффективности.[14]
Взаимодействия
Обычные фармацевтические препараты (и добавки) с антиоксидантными свойствами могут влиять на эффективность некоторых противораковое средство лекарства и радиационная терапия.[15]
Побочные эффекты

Относительно сильные восстанавливающие кислоты могут иметь антинутриент эффекты путем привязки к диетические минералы Такие как утюг и цинк в желудочно-кишечный тракт и предотвращая их всасывание.[16] Примеры Щавелевая кислота, дубильные вещества и фитиновая кислота, которые богаты растительными диетами.[17] Кальций и дефицит железа не редкость в диетах в развивающиеся страны где меньше мяса едят и где высокое потребление фитиновой кислоты из бобов и пресных целое зерно хлеб. Однако проращивание, замачивание или микробная ферментация - это все домашние стратегии, которые снижают содержание фитатов и полифенолов в неочищенных злаках. Сообщалось об увеличении абсорбции Fe, Zn и Ca у взрослых, которых кормили дефитинизированными злаками, по сравнению со злаками, содержащими их природный фитат.[18]
| Еда | Уменьшение присутствия кислоты |
|---|---|
| Какао бобы и шоколад, шпинат, репа и ревень[19] | Щавелевая кислота |
| Цельное зерно, кукуруза, бобовые[20] | Фитиновая кислота |
| Чай, бобы, капуста[19][21] | Танины |
Высокие дозы некоторых антиоксидантов могут иметь долгосрочные вредные последствия. В бета-каротин и Ретинол Испытание эффективности (CARET) исследование пациентов с раком легких показало, что у курильщиков, получавших добавки, содержащие бета-каротин и витамин А, повышалась частота рака легких.[22] Последующие исследования подтвердили эти побочные эффекты.[23] Эти вредные эффекты могут также наблюдаться у некурящих, поскольку один метаанализ включая данные примерно 230000 пациентов, которые показали, что добавление β-каротина, витамина A или витамина E связано с повышенной смертностью, но не наблюдалось значительного эффекта от витамина C.[24] Никакого риска для здоровья не наблюдалось, когда все рандомизированные контролируемые исследования были изучены вместе, но рост смертности был обнаружен, когда отдельно рассматривались только исследования высокого качества и с низким риском смещения.[25] Поскольку большинство этих исследований с низким уровнем систематической ошибки касались либо пожилые люди или людей с заболеваниями, эти результаты могут не относиться к населению в целом.[26] Позднее этот метаанализ был повторен и расширен теми же авторами, подтвердив предыдущие результаты.[25] Эти две публикации согласуются с некоторыми предыдущими метаанализами, которые также предположили, что добавление витамина E увеличивает смертность,[27] и что антиоксидантные добавки увеличивают риск рак толстой кишки.[28] Бета-каротин может также увеличить рак легких.[28][29] В целом, большое количество клинических испытаний антиоксидантных добавок позволяет предположить, что либо эти продукты не влияют на здоровье, либо вызывают небольшое увеличение смертности среди пожилых или уязвимых групп населения.[9][10][24]
Окислительная проблема в биологии

А парадокс в метаболизм в том, что подавляющее большинство сложных жизнь на Земле требует кислород для своего существования кислород является высокореактивным элементом, который повреждает живые организмы, производя активные формы кислорода.[30] Следовательно, организмы содержат сложную сеть антиоксидантов. метаболиты и ферменты которые работают вместе, чтобы предотвратить окислительное повреждение клеточных компонентов, таких как ДНК, белки и липиды.[31][32] В общем, антиоксидантные системы либо предотвращают образование этих реактивных частиц, либо удаляют их до того, как они могут повредить жизненно важные компоненты клетки.[30][31] Однако активные формы кислорода также выполняют полезные клеточные функции, такие как редокс-сигнализация. Таким образом, функция антиоксидантных систем заключается не в полном удалении окислителей, а в поддержании их на оптимальном уровне.[33]
Активные формы кислорода, продуцируемые в клетках, включают: пероксид водорода (ЧАС2О2), хлорноватистая кислота (HClO) и свободные радикалы такой как гидроксильный радикал (· OH) и супероксид-анион (O2−).[34] Гидроксильный радикал особенно нестабилен и будет реагировать быстро и неспецифично с большинством биологических молекул. Этот вид образуется из перекиси водорода в катализируемый металлом окислительно-восстановительные реакции, такие как Реакция Фентона.[35] Эти окислители могут повредить клетки, запуская цепные химические реакции, такие как перекисное окисление липидов или путем окисления ДНК или белков.[31] Повреждение ДНК может вызвать мутации и, возможно, рак, если его не исправить Ремонт ДНК механизмы,[36][37] при повреждении белки вызывает угнетение ферментов, денатурация и деградация белка.[38]
Использование кислорода в процессе выработки метаболической энергии приводит к образованию активных форм кислорода.[39] В этом процессе супероксид-анион образуется в виде побочный продукт нескольких шагов в электронная транспортная цепь.[40] Особенно важно снижение коэнзим Q в комплекс III, поскольку в качестве промежуточного соединения (Q·−). Этот нестабильный промежуточный продукт может привести к "утечке" электронов, когда электроны прыгают прямо на кислород и образуют супероксид-анион, вместо того, чтобы проходить через нормальную серию хорошо контролируемых реакций цепи переноса электронов.[41] Пероксид также образуется в результате окисления восстановленных флавопротеины, Такие как комплекс I.[42] Однако, хотя эти ферменты могут продуцировать окислители, относительная важность цепи переноса электронов для других процессов, которые генерируют пероксид, неясна.[43][44] У растений водоросли, и цианобактерии, активные формы кислорода также образуются во время фотосинтез,[45] особенно в условиях высоких Интенсивность света.[46] Этот эффект частично компенсируется участием каротиноиды в фотоингибирование, а у водорослей и цианобактерий - большим количеством йодид и селен,[47] который включает эти антиоксиданты, реагирующие с чрезмерно восстановленными формами фотосинтетические реакционные центры для предотвращения образования активных форм кислорода.[48][49]
Примеры биоактивных антиоксидантных соединений
Антиоксиданты подразделяются на две широкие группы в зависимости от того, растворимы ли они в воде (гидрофильный ) или в липидах (липофильный ). Обычно водорастворимые антиоксиданты реагируют с окислителями в клетке. цитозоль и плазма крови, а липидорастворимые антиоксиданты защищают клеточные мембраны из перекисное окисление липидов.[31] Эти соединения могут быть синтезированы в организме или получены с пищей.[32] Различные антиоксиданты присутствуют в широком диапазоне концентраций в телесные жидкости и ткани, с некоторыми, такими как глутатион или же убихинон в основном присутствуют внутри клеток, в то время как другие, такие как мочевая кислота распределяются более равномерно (см. таблицу ниже). Некоторые антиоксиданты обнаружены только в нескольких организмах, и эти соединения могут иметь важное значение для патогены и может быть факторы вирулентности.[50]
Относительная важность и взаимодействие между этими различными антиоксидантами - очень сложный вопрос, поскольку различные антиоксидантные соединения и системы антиоксидантных ферментов имеют синергетический и взаимозависимые эффекты друг на друга.[51][52] Следовательно, действие одного антиоксиданта может зависеть от правильного функционирования других членов антиоксидантной системы.[32] Степень защиты, обеспечиваемая любым антиоксидантом, также будет зависеть от его концентрации, его реакционной способности по отношению к конкретным рассматриваемым реактивным формам кислорода и статуса антиоксидантов, с которыми он взаимодействует.[32]
Некоторые соединения способствуют антиоксидантной защите за счет хелатирующий переходные металлы и предотвращение того, чтобы они катализировали производство свободных радикалов в клетке. Особенно важна способность связывать железо, что является функцией железосвязывающие белки Такие как трансферрин и ферритин.[44] Селен и цинк обычно называют антиоксидантные питательные вещества, но эти химические элементы сами по себе не обладают антиоксидантным действием и вместо этого необходимы для активности некоторых антиоксидантных ферментов, как обсуждается ниже.
| Антиоксидант | Растворимость | Концентрация в сыворотке крови человека (мкМ) | Концентрация в ткани печени (мкмоль / кг) |
|---|---|---|---|
| Аскорбиновая кислота (Витамин С ) | Вода | 50–60[53] | 260 (человек)[54] |
| Глутатион | Вода | 4[55] | 6,400 (человек)[54] |
| Липоевая кислота | Вода | 0.1–0.7[56] | 4–5 (крыса)[57] |
| Мочевая кислота | Вода | 200–400[58] | 1600 (человек)[54] |
| Каротины | Липид | β-каротин: 0.5–1[59] | 5 (человек, общие каротиноиды)[61] |
| α-токоферол (витамин Е) | Липид | 10–40[60] | 50 (человек)[54] |
| Убихинол (коэнзим Q) | Липид | 5[62] | 200 (человек)[63] |
Мочевая кислота
Мочевая кислота является антиоксидантом с самой высокой концентрацией в крови человека. Мочевая кислота (UA) - это оксипурин-антиоксидант, производимый из ксантин ферментом ксантиноксидаза, и является промежуточным продуктом пурин метаболизм.[64] Почти у всех наземных животных, уратоксидаза дополнительно катализирует окисление мочевой кислоты до аллантоин,[65] но у людей и большинства высших приматов ген уратоксидазы нефункционален, поэтому дальнейшее расщепление UA не происходит.[65][66] Эволюционные причины потери превращения уратов в аллантоин остаются предметом активных предположений.[67][68] Антиоксидантное действие мочевой кислоты привело исследователей к предположению, что эта мутация была полезна для первых приматов и людей.[68][69] Исследования высотной акклиматизации подтверждают гипотезу о том, что ураты действуют как антиоксидант, смягчая окислительный стресс, вызванный высокогорной гипоксией.[70]
Мочевая кислота имеет самую высокую концентрацию любого антиоксиданта в крови[58] и обеспечивает более половины общей антиоксидантной способности сыворотки крови человека.[71] Антиоксидантная активность мочевой кислоты также сложна, учитывая, что она не реагирует с некоторыми окислителями, такими как супероксид, но действует против пероксинитрит,[72] перекиси, и хлорноватистая кислота.[64] Опасения по поводу повышенного вклада UA в подагра следует рассматривать как один из многих факторов риска.[73] Сам по себе связанный с UA риск подагры на высоких уровнях (415–530 мкмоль / л) составляет всего 0,5% в год с увеличением до 4,5% в год в UA. уровни перенасыщения (535+ мкмоль / л).[74] Многие из этих вышеупомянутых исследований определили антиоксидантное действие UA в пределах нормальных физиологических уровней,[70][72] а некоторые обнаружили антиоксидантную активность на уровне 285 мкмоль / л.[75]
Витамин С
Аскорбиновая кислота или же Витамин С это моносахарид Снижение окисления (редокс ) катализатор встречается как у животных, так и у растений.[76] Поскольку один из ферментов, необходимых для производства аскорбиновой кислоты, был утерян мутация в течение примат эволюция люди должны получать его из своего рациона; поэтому это диетический витамин.[76][77] Большинство других животных способны вырабатывать это соединение в своем организме и не нуждаются в нем в своем рационе.[78] Аскорбиновая кислота необходима для превращения проколлаген к коллаген путем окисления пролин остатки гидроксипролин.[76] В других клетках он поддерживается в восстановленной форме за счет реакции с глутатионом, которая может катализироваться протеин дисульфид изомераза и глутаредоксины.[79][80] Аскорбиновая кислота представляет собой окислительно-восстановительный катализатор, который может восстанавливать и тем самым нейтрализовать активные формы кислорода, такие как перекись водорода.[76][81] Помимо прямого антиоксидантного действия аскорбиновая кислота также субстрат для окислительно-восстановительного фермента аскорбатпероксидаза, функция, которая используется для устойчивости растений к стрессу.[82] Аскорбиновая кислота присутствует в больших количествах во всех частях растений и может достигать 20%.миллимолярный в хлоропласты.[83]
Глутатион
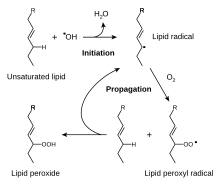
Глутатион это цистеин -содержащий пептид встречается в большинстве форм аэробной жизни.[84] Он не требуется в диете и вместо этого синтезируется в клетках из его составляющих. аминокислоты.[85] Глутатион обладает антиоксидантными свойствами, так как тиол группа в своем цистеин часть является восстановителем и может обратимо окисляться и восстанавливаться. В клетках глутатион поддерживается в восстановленной форме ферментом глутатионредуктаза и, в свою очередь, снижает другие метаболиты и ферментные системы, такие как аскорбат в глутатион-аскорбатный цикл, глутатионпероксидазы и глутаредоксины, а также непосредственно реагирует с окислителями.[79] Благодаря своей высокой концентрации и центральной роли в поддержании окислительно-восстановительного состояния клетки, глутатион является одним из важнейших клеточных антиоксидантов.[84] В некоторых организмах глутатион заменяется другими тиолами, например, микотиол в Актиномицеты, бациллитиол в некоторых Грамположительные бактерии,[86][87] или по трипанотион в Кинетопластиды.[88][89]
Витамин Е
Витамин Е - собирательное название набора из восьми связанных токоферолы и токотриенолы, которые жирорастворимый витамины с антиоксидантными свойствами.[90][91] Из них наиболее изучен альфа-токоферол, так как у него самый высокий биодоступность, причем организм преимущественно поглощает и метаболизирует эту форму.[92]
Было заявлено, что форма α-токоферола является наиболее важным жирорастворимым антиоксидантом и что она защищает мембраны от окисления, вступая в реакцию с липидными радикалами, образующимися в цепной реакции перекисного окисления липидов.[90][93] Это удаляет промежуточные продукты свободных радикалов и предотвращает продолжение реакции распространения. В результате этой реакции образуются окисленные α-токофероксильные радикалы, которые могут быть возвращены в активную восстановленную форму путем восстановления другими антиоксидантами, такими как аскорбат, ретинол или убихинол.[94] Это согласуется с результатами, показывающими, что α-токоферол, но не водорастворимые антиоксиданты, эффективно защищает глутатионпероксидазу 4 (GPX4 ) -дефицитные клетки от гибели клеток.[95] GPx4 - единственный известный фермент, который эффективно восстанавливает гидропероксиды липидов в биологических мембранах.
Однако роль и важность различных форм витамина Е в настоящее время неясны.[96][97] и даже было высказано предположение, что наиболее важная функция α-токоферола - это сигнальная молекула, причем эта молекула не играет значительной роли в метаболизме антиоксидантов.[98][99] Функции других форм витамина E еще менее изучены, хотя γ-токоферол является нуклеофил который может реагировать с электрофильный мутагены,[92] и токотриенолы могут играть важную роль в защите нейроны от повреждений.[100]
Прооксидантная активность
Антиоксиданты, являющиеся восстановителями, также могут действовать как прооксиданты. Например, витамин C обладает антиоксидантной активностью, когда восстанавливает окисляющие вещества, такие как перекись водорода;[101] однако он также уменьшит ионы металлов, которые генерируют свободные радикалы через Реакция Фентона.[35][102]
- 2 Fe3+ + Аскорбат → 2 Fe2+ + Дегидроаскорбат
- 2 Fe2+ + 2 часа2О2 → 2 Fe3+ + 2 ОН· + 2 ОН−
Относительная важность антиоксидантной и прооксидантной активности антиоксидантов является областью текущих исследований, но витамин С, который проявляет свое действие как витамин путем окисления полипептидов, по-видимому, оказывает в организме человека в основном антиоксидантное действие.[102] Однако меньше данных доступно для других пищевых антиоксидантов, таких как витамин E,[103] или полифенолы.[104][105] Точно так же патогенез заболеваний, связанных с гиперурикемией, вероятно, связан с прямыми и косвенными прооксидантными свойствами мочевой кислоты.
То есть, как это ни парадоксально, агенты, которые обычно считаются антиоксидантами, могут действовать как условные прооксиданты и фактически увеличивают окислительный стресс. Помимо аскорбата, важные с медицинской точки зрения условные прооксиданты включают мочевую кислоту и сульфгидриламинокислоты, такие как гомоцистеин. Как правило, в качестве катализатора используется какой-либо металл переходного ряда, например медь или железо. Потенциальная роль прооксидантной роли мочевой кислоты, например, при атеросклерозе и ишемическом инсульте рассмотрена выше. Другой пример - постулируемая роль гомоцистеин при атеросклерозе.
Ферментные системы
![{displaystyle {ce {{underset {Oxygen} {O2}} -> {underset {Superoxide} {* O2 ^ {-}}} -> [{ce {Супероксид на дисмутазе}}] {underset {водород на пероксиде} { H2O2}} -> [{ce {Пероксидазы на каталазе}}] {underset {Water} {H2O}}}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/8feecd26377be56b431d182ea9a398ab6b5d3b7f)
Как и в случае с химическими антиоксидантами, клетки защищены от окислительного стресса взаимодействующей сетью антиоксидантных ферментов.[30][31] Здесь супероксид, выделяемый такими процессами, как окислительного фосфорилирования сначала превращается в перекись водорода, а затем восстанавливается до воды. Этот путь детоксикации является результатом действия множества ферментов: супероксиддисмутазы катализируют первую стадию, а затем каталазы и различные пероксидазы удаляют перекись водорода. Как и в случае метаболитов антиоксидантов, вклад этих ферментов в антиоксидантную защиту может быть трудно отделить друг от друга, но образование трансгенные мыши недостаток только одного антиоксидантного фермента может быть информативным.[106]
Супероксиддисмутаза, каталаза и пероксиредоксины
Супероксиддисмутазы (SOD) - это класс близкородственных ферментов, которые катализируют распад супероксид-аниона на кислород и перекись водорода.[107][108] Ферменты СОД присутствуют почти во всех аэробных клетках и во внеклеточных жидкостях.[109] Ферменты супероксиддисмутазы содержат кофакторы ионов металлов, которые, в зависимости от изофермента, могут быть медью, цинком, марганец или утюг. У человека СОД медь / цинк присутствует в цитозоль, а марганцевая СОД присутствует в митохондрия.[108] Существует также третья форма SOD в внеклеточные жидкости, который содержит медь и цинк в своих активных центрах.[110] Митохондриальный изофермент, по-видимому, является наиболее биологически важным из этих трех, поскольку мыши, лишенные этого фермента, умирают вскоре после рождения.[111] Напротив, мыши, лишенные SOD медь / цинк (Sod1), жизнеспособны, но имеют многочисленные патологии и сокращенную продолжительность жизни (см. Статью о супероксид ), в то время как мыши без внеклеточной SOD имеют минимальные дефекты (чувствительные к гипероксия ).[106][112] У растений изоферменты СОД присутствуют в цитозоле и митохондриях, а СОД железа содержится в хлоропласты что отсутствует в позвоночные и дрожжи.[113]
Каталазы представляют собой ферменты, которые катализируют превращение перекиси водорода в воду и кислород с использованием кофактора железа или марганца.[114][115] Этот белок локализован в пероксисомы в большинстве эукариотический клетки.[116] Каталаза - необычный фермент, поскольку, хотя перекись водорода является ее единственным субстратом, она следует за механизм для пинг-понга. Здесь его кофактор окисляется одной молекулой перекиси водорода, а затем регенерируется путем переноса связанного кислорода на вторую молекулу субстрата.[117] Несмотря на очевидную важность удаления перекиси водорода, люди с генетическим дефицитом каталазы - "акаталаземия "- или мыши генно-инженерный полностью лишившись каталазы, мало страдают от побочных эффектов.[118][119]

Пероксиредоксины пероксидазы, которые катализируют восстановление перекиси водорода, органические гидропероксиды, а также пероксинитрит.[121] Они делятся на три класса: типичные пероксиредоксины 2-цистеина; атипичные пероксиредоксины 2-цистеина; и пероксиредоксины 1-цистеина.[122] Эти ферменты имеют один и тот же основной каталитический механизм, при котором окислительно-восстановительный цистеин (перекисный цистеин) в активный сайт окисляется до сульфеновая кислота пероксидным субстратом.[123] Чрезмерное окисление этого остатка цистеина в пероксиредоксинах инактивирует эти ферменты, но это можно обратить вспять действием сульфиредоксин.[124] Пероксиредоксины, по-видимому, играют важную роль в метаболизме антиоксидантов, поскольку мыши, лишенные пероксиредоксина 1 или 2, сокращают продолжительность жизни и страдают от гемолитическая анемия, в то время как растения используют пероксиредоксины для удаления перекиси водорода, образующейся в хлоропластах.[125][126][127]
Системы тиоредоксина и глутатиона
В тиоредоксин система содержит 12-кДа протеин тиоредоксин и его компаньон тиоредоксинредуктаза.[128] Белки, относящиеся к тиоредоксину, присутствуют во всех секвенированных организмах. Растения, такие как Arabidopsis thaliana, имеют особенно большое разнообразие изоформ.[129] Активный центр тиоредоксина состоит из двух соседний цистеины, как часть высококонсервативного CXXC мотив, который может циклически переключаться между активной (восстановленной) формой дитиола и окисленной дисульфид форма. В активном состоянии тиоредоксин действует как эффективный восстанавливающий агент, улавливая активные формы кислорода и поддерживая другие белки в их восстановленном состоянии.[130] После окисления активный тиоредоксин регенерируется под действием тиоредоксинредуктазы с использованием НАДФН как донор электронов.[131]
В глутатион система включает глутатион, глутатионредуктаза, глутатионпероксидазы, и глутатион S-трансферазы.[84] Эта система содержится у животных, растений и микроорганизмов.[84][132] Глутатионпероксидаза - это фермент, содержащий четыре селен -кофакторы который катализирует распад перекиси водорода и органических гидропероксидов. Существует как минимум четыре различных глутатионпероксидазы. изоферменты у животных.[133] Глутатионпероксидаза 1 является наиболее распространенным и очень эффективным поглотителем перекиси водорода, в то время как глутатионпероксидаза 4 наиболее активна с гидропероксидами липидов. Удивительно, но глутатионпероксидаза 1 не обязательна, поскольку у мышей, у которых отсутствует этот фермент, нормальная продолжительность жизни,[134] но они сверхчувствительны к индуцированному окислительному стрессу.[135] Кроме того, глутатион S-трансферазы проявляют высокую активность в отношении перекисей липидов.[136] Эти ферменты находятся на особенно высоком уровне в печени и также служат в детоксикация метаболизм.[137]
Использование в технологии
Пищевые консерванты
Антиоксиданты используются как пищевые добавки помочь предохраняться от порчи продуктов питания. Воздействие кислорода и солнечного света являются двумя основными факторами окисления пищи, поэтому пищу можно сохранить, храня в темноте и запечатывая в контейнерах или даже покрывая воском, как в случае с огурцами. Однако, поскольку кислород также важен для растений дыхание, хранение растительного сырья в анаэробный условия создают неприятный вкус и непривлекательные цвета.[138] Следовательно, упаковка свежих фруктов и овощей содержит ~ 8% кислорода в атмосфере. Антиоксиданты - особенно важный класс консервантов, поскольку, в отличие от бактериальных или грибковый порча, реакции окисления все еще протекают относительно быстро в замороженных или охлажденных пищевых продуктах.[139] Эти консерванты включают природные антиоксиданты, такие как аскорбиновая кислота (AA, E300) и токоферолы (E306), а также синтетические антиоксиданты, такие как пропилгаллат (PG, E310), третичный бутилгидрохинон (TBHQ), бутилированный гидроксианизол (BHA, E320) и бутилированный гидрокситолуол (BHT, E321).[140][141]
Наиболее распространенными молекулами, подвергающимися окислению, являются ненасыщенные жиры; окисление заставляет их превращаться протухший.[142] Поскольку окисленные липиды часто обесцвечиваются и имеют неприятный привкус, например металлический или сернистый ароматизаторов, важно избегать окисления в богатой жирами пище. Таким образом, эти продукты редко сохраняются путем сушки; вместо этого они сохраняются курение, засолка или же брожение. Даже менее жирные продукты, такие как фрукты, перед сушкой на воздухе опрыскиваются серосодержащими антиоксидантами. Окисление часто катализируется металлами, поэтому жиры, такие как масло, никогда не следует оборачивать. алюминиевая фольга или хранятся в металлических контейнерах. Некоторые жирные продукты, такие как оливковое масло, частично защищены от окисления благодаря естественному содержанию антиоксидантов, но остаются чувствительными к фотоокислению.[143] Антиоксидантные консерванты также добавляются в косметику на жировой основе, такую как помада и увлажняющие средства чтобы предотвратить прогорклость.
Промышленное использование

Антиоксиданты часто добавляют в промышленные продукты. Обычно используется как стабилизаторы в топливо и смазочные материалы для предотвращения окисления, а в бензинах - для предотвращения полимеризации, которая приводит к образованию остатков загрязнения двигателя.[144] В 2014 году мировой рынок природных и синтетических антиоксидантов составлял 2,25 миллиарда долларов США с прогнозом роста до 3,25 миллиарда долларов к 2020 году.[145]
Антиоксидант полимерные стабилизаторы широко используются для предотвращения деградации полимеры такие как каучуки, пластмассы и клеи что вызывает потерю прочности и гибкости этих материалов.[146] Полимеры, содержащие двойные связи в их основных цепочках, таких как натуральная резина и полибутадиен, особенно восприимчивы к окисление и озонолиз. Их можно защитить антиозонанты. Твердые полимерные продукты начинают трескаться на открытых поверхностях по мере разрушения материала и разрыва цепей. Режим крекинга варьируется от кислорода до озон нападение, первое вызывает эффект «сумасшедшего мощения», в то время как воздействие озона вызывает более глубокие трещины, расположенные под прямым углом к деформации растяжения в изделии. Окисление и УФ-деградация также часто связаны, в основном потому, что УФ-излучение создает свободные радикалы путем разрыва связи. Затем свободные радикалы реагируют с кислородом с образованием пероксия радикалы, которые вызывают еще больший ущерб, часто в цепная реакция. Другие полимеры, подверженные окислению, включают: полипропилен и полиэтилен. Первый более чувствителен из-за наличия вторичные атомы углерода присутствует в каждом повторяющемся блоке. Атака происходит в этот момент, потому что образующийся свободный радикал более стабилен, чем образующийся первичный атом углерода. Окисление полиэтилена обычно происходит в слабых звеньях цепи, таких как точки разветвления в полиэтилен низкой плотности.
| Присадка к топливу | Составные части[147] | Приложения[147] |
|---|---|---|
| АО-22 | N, N'-ди-2-бутил-1,4-фенилендиамин | Турбинные масла, трансформаторные масла, гидравлические жидкости, воск, и смазки |
| АО-24 | N, N'-ди-2-бутил-1,4-фенилендиамин | Низкотемпературные масла |
| АО-29 | 2,6-ди-трет-бутил-4-метилфенол | Турбинные масла, трансформаторные масла, гидравлические жидкости, воски, консистентные смазки и бензины |
| АО-30 | 2,4-диметил-6-трет-бутилфенол | Реактивное топливо и бензины, в том числе авиационные бензины |
| АО-31 | 2,4-диметил-6-трет-бутилфенол | Топливо для реактивных двигателей и бензины, в том числе авиационные бензины |
| АО-32 | 2,4-диметил-6-трет-бутилфенол и 2,6-ди-трет-бутил-4-метилфенол | Топливо для реактивных двигателей и бензины, включая авиационные бензины |
| АО-37 | 2,6-ди-трет-бутилфенол | Реактивное топливо и бензины, широко одобренные для авиационное топливо |
Уровни в пище

Витамины-антиоксиданты содержатся в овощах, фруктах, яйцах, бобовых и орехах. Витамины A, C и E могут быть разрушены при длительном хранении или продолжительном приготовлении пищи.[148] Влияние приготовления пищи и обработки пищевых продуктов сложное, поскольку эти процессы также могут увеличить биодоступность антиоксидантов, таких как некоторые каротиноиды в овощах.[149] Обработанная пища содержит меньше витаминов-антиоксидантов, чем свежие и сырые продукты, поскольку при приготовлении пища подвергается воздействию тепла и кислорода.[150]
| Витамины-антиоксиданты | Продукты с высоким содержанием витаминов-антиоксидантов[21][151][152] |
|---|---|
| Витамин С (аскорбиновая кислота) | Свежие или замороженные фрукты и овощи |
| Витамин Е (токоферолы, токотриенолы) | Растительные масла, орехи, и семена |
| Каротиноиды (каротины в качестве провитамин А ) | Фрукты, овощи и яйца |
Другие антиоксиданты не поступают с пищей, а вырабатываются в организме. Например, убихинол (коэнзим Q) плохо всасывается из кишечника и производится через мевалонатный путь.[63] Другой пример глутатион, который состоит из аминокислот. Поскольку любой глутатион в кишечнике расщепляется на свободный цистеин, глицин и глютаминовая кислота до того, как он будет поглощен, даже большой пероральный прием мало влияет на концентрацию глутатиона в организме.[153][154] Хотя большое количество серосодержащих аминокислот, таких как ацетилцистеин может увеличить глутатион,[155] Нет никаких доказательств того, что потребление большого количества этих предшественников глутатиона полезно для здоровых взрослых.[156]
Измерение и признание недействительности ORAC
Измерение содержания полифенолов и каротиноидов в пище - непростой процесс, поскольку антиоксиданты в совокупности представляют собой разнообразную группу соединений с разной реакционной способностью по отношению к различным реактивным формам кислорода. В наука о еде анализы in vitro, абсорбционная способность кислородных радикалов (ORAC) когда-то был отраслевым стандартом для оценки антиоксидантной силы цельных продуктов, соков и пищевых добавок, в основном по наличию полифенолы.[157][158] Предыдущие измерения и оценки Министерство сельского хозяйства США были отозваны в 2012 году как биологически не относящиеся к здоровью человека, ссылаясь на отсутствие физиологических доказательств того, что полифенолы обладают антиоксидантными свойствами. in vivo.[159] Следовательно, метод ORAC, полученный только из in vitro экспериментов, больше не считается имеющим отношение к рациону человека или биология, по состоянию на 2010 год.[159]
Альтернативные измерения содержания антиоксидантов в пищевых продуктах in vitro, также основанные на присутствии полифенолов, включают: Реагент Фолин-Чокальтеу, а Эквивалентная антиоксидантная способность Trolox проба.[160]
История
В рамках адаптации к морской жизни наземные растения начали производить неморские антиоксиданты, такие как аскорбиновая кислота (Витамин С ), полифенолы и токоферолы. Эволюция покрытосеменные растения между 50 и 200 миллионами лет назад привели к развитию многих антиоксидантных пигментов, особенно во время Юрский период - как химическая защита от активные формы кислорода которые являются побочными продуктами фотосинтез.[161] Первоначально термин «антиоксидант» относился к химическому веществу, предотвращающему потребление кислорода. В конце 19 - начале 20 веков обширные исследования были сосредоточены на использовании антиоксидантов в важных промышленных процессах, таких как предотвращение образования металлов. коррозия, то вулканизация резины, и полимеризация топлива в обрастание из двигатель внутреннего сгорания.[162]
Ранние исследования роли антиоксидантов в биологии были сосредоточены на их использовании для предотвращения окисления ненасыщенные жиры, что является причиной прогорклость.[163] Антиоксидантную активность можно было измерить, просто поместив жир в закрытый контейнер с кислородом и измерив скорость потребления кислорода. Однако это была идентификация витамины С и E как антиоксиданты, которые произвели революцию в этой области и привели к осознанию важности антиоксидантов в биохимии живые организмы.[164][165] Возможный механизмы действия антиоксидантов были впервые исследованы, когда было признано, что вещество с антиоксидантной активностью, вероятно, само по себе легко окисляется.[166] Исследование того, как витамин Е предотвращает процесс перекисное окисление липидов привело к идентификации антиоксидантов как восстановителей, предотвращающих окислительные реакции, часто за счет уборка мусора активные формы кислорода прежде, чем они смогут повредить клетки.[167]
Рекомендации
- ^ Дабельштейн В., Реглицкий А., Шютце А., Редерс К. (2007). «Автомобильное топливо». Энциклопедия промышленной химии Ульмана. Дои:10.1002 / 14356007.a16_719.pub2. ISBN 978-3-527-30673-2.
- ^ «Антиоксиданты: глубже». NCCIH. Июнь 2010. Архивировано с оригинал 25 августа 2018 г.. Получено 20 июн 2018.
- ^ Бжелакович Г, Николова Д, Глууд С (2013). «Мета-регрессионные анализы, мета-анализы и последовательные исследования воздействия добавок с бета-каротином, витамином А и витамином Е по отдельности или в различных комбинациях на общую смертность: есть ли у нас доказательства отсутствия вреда? ". PLOS ONE. 8 (9): e74558. Bibcode:2013PLoSO ... 874558B. Дои:10.1371 / journal.pone.0074558. ЧВК 3765487. PMID 24040282.
- ^ Абнер Е.Л., Шмитт Ф.А., Мендиондо М.С., Маркум Дж.Л., Крысцио Р.Дж. (июль 2011 г.). «Витамин Е и смертность от всех причин: метаанализ». Современная наука о старении. 4 (2): 158–70. Дои:10.2174/1874609811104020158. ЧВК 4030744. PMID 21235492.
- ^ Кортес-Жофре М., Руэда Дж. Р., Корсини-Муньос Дж., Фонсека-Кортес С., Карабальозо М., Bonfill Cosp X (2012). «Препараты для профилактики рака легких у здоровых людей». Кокрановская база данных систематических обзоров. 10: CD002141. Дои:10.1002 / 14651858.CD002141.pub2. PMID 23076895.
- ^ Цзян Л., Ян К. Х., Тиан Дж. Х., Гуань КЛ, Яо Н, Цао Н, Ми Д.Х., Ву Дж, Ма Б., Ян Ш. (2010). «Эффективность антиоксидантных витаминов и добавок селена в профилактике рака простаты: метаанализ рандомизированных контролируемых исследований». Питание и рак. 62 (6): 719–27. Дои:10.1080/01635581.2010.494335. PMID 20661819. S2CID 13611123.
- ^ Рис К., Хартли Л., День С, Флауэрс Н., Кларк А., Странджи С. (2013). «Добавка селена для первичной профилактики сердечно-сосудистых заболеваний» (PDF). Кокрановская база данных систематических обзоров. 1 (1): CD009671. Дои:10.1002 / 14651858.CD009671.pub2. ЧВК 7433291. PMID 23440843. Архивировано из оригинал (PDF) 12 августа 2017 г.. Получено 23 июля 2018.
- ^ Shekelle PG, Morton SC, Jungvig LK, Udani J, Spar M, Tu W, J Suttorp M, Coulter I, Newberry SJ, Hardy M (апрель 2004 г.). «Эффект дополнительного приема витамина Е для профилактики и лечения сердечно-сосудистых заболеваний». Журнал общей внутренней медицины. 19 (4): 380–9. Дои:10.1111 / j.1525-1497.2004.30090.x. ЧВК 1492195. PMID 15061748.
- ^ а б c Станнер С.А., Хьюз Дж., Келли С.Н., Батрисс Дж. (Май 2004 г.). "Обзор эпидемиологических доказательств" антиоксидантной гипотезы ".'". Питание для общественного здравоохранения. 7 (3): 407–22. Дои:10.1079 / PHN2003543. PMID 15153272.
- ^ а б Шенкин А. (февраль 2006 г.). «Ключевая роль микронутриентов». Клиническое питание. 25 (1): 1–13. Дои:10.1016 / j.clnu.2005.11.006. PMID 16376462.
- ^ Вудсайд СП, МакКолл Д., Макгартланд С., Янг И.С. (ноябрь 2005 г.). «Микроэлементы: потребление с пищей против использования добавок». Труды Общества питания. 64 (4): 543–53. Дои:10.1079 / PNS2005464. PMID 16313697.
- ^ Еда, питание, физическая активность и профилактика рака: глобальная перспектива. Всемирный фонд исследования рака (2007). ISBN 978-0-9722522-2-5.
- ^ Приветствую Н., Кортес М., Дрейк Е. Н., Спаллхольц Дж. Э. (июль 2008 г.). «Химиопрофилактика рака: радикальная перспектива». Свободная радикальная биология и медицина. 45 (2): 97–110. Дои:10.1016 / j.freeradbiomed.2008.04.004. PMID 18454943.
- ^ «Флавоноиды». Институт Линуса Полинга, Государственный университет Орегона, Корваллис. 2016 г.. Получено 24 июля 2016.
- ^ Леммо В. (сентябрь 2014 г.). «Возможные взаимодействия отпускаемых по рецепту и без рецепта лекарств, обладающих антиоксидантными свойствами, с лучевой и химиотерапией». Международный журнал рака. 137 (11): 2525–33. Дои:10.1002 / ijc.29208. PMID 25220632. S2CID 205951215.
- ^ Hurrell RF (сентябрь 2003 г.). «Влияние источников растительного белка на биодоступность микроэлементов и минералов». Журнал питания. 133 (9): 2973S – 7S. Дои:10.1093 / jn / 133.9.2973S. PMID 12949395.
- ^ Хант-младший (сентябрь 2003 г.). «Биодоступность железа, цинка и других микроэлементов из вегетарианских диет». Американский журнал клинического питания. 78 (3 приложения): 633S – 639S. Дои:10.1093 / ajcn / 78.3.633S. PMID 12936958.
- ^ Гибсон Р.С., Перлас Л., Хотц С. (май 2006 г.). «Повышение биодоступности питательных веществ в растительной пище на бытовом уровне». Труды Общества питания. 65 (2): 160–8. Дои:10.1079 / PNS2006489. PMID 16672077.
- ^ а б Mosha TC, Gaga HE, Pace RD, Laswai HS, Mtebe K (июнь 1995 г.). «Влияние бланширования на содержание антипитательных факторов в избранных овощах». Растительные продукты для питания человека. 47 (4): 361–7. Дои:10.1007 / BF01088275. PMID 8577655. S2CID 1118651.
- ^ Сандберг А.С. (декабрь 2002 г.). «Биодоступность минералов в бобовых». Британский журнал питания. 88 Suppl 3 (Suppl 3): S281–5. Дои:10.1079 / BJN / 2002718. PMID 12498628.
- ^ а б Бичер Г.Р. (октябрь 2003 г.). «Обзор диетических флавоноидов: номенклатура, встречаемость и потребление». Журнал питания. 133 (10): 3248S – 3254S. Дои:10.1093 / jn / 133.10.3248S. PMID 14519822.
- ^ Оменн Г.С., Гудман Г.Е., Торнквист М.Д., Бальмс Дж., Каллен М.Р., Гласс А., Кио Дж. П., Мейскенс Флорида, Валанис Б., Уильямс Дж. Х., Барнхарт С., Черняк М.Г., Бродкин К.А., Хаммар С. (ноябрь 1996 г.). «Факторы риска рака легких и воздействия вмешательства в CARET, испытании эффективности бета-каротина и ретинола» (PDF). Журнал Национального института рака. 88 (21): 1550–9. Дои:10.1093 / jnci / 88.21.1550. PMID 8901853.
- ^ Албейнс Д. (июнь 1999 г.). «Бета-каротин и рак легких: тематическое исследование». Американский журнал клинического питания. 69 (6): 1345S – 50S. Дои:10.1093 / ajcn / 69.6.1345S. PMID 10359235.
- ^ а б Белакович Г., Николова Д., Глууд Л.Л., Симонетти Р.Г., Глууд С. (февраль 2007 г.). «Смертность в рандомизированных исследованиях антиоксидантных добавок для первичной и вторичной профилактики: систематический обзор и метаанализ». JAMA. 297 (8): 842–57. Дои:10.1001 / jama.297.8.842. PMID 17327526.
- ^ а б Белакович Г., Николова Д., Глууд Л.Л., Симонетти Р.Г., Глууд С. (14 марта 2012 г.). «Антиоксидантные добавки для предотвращения смертности здоровых участников и пациентов с различными заболеваниями». Кокрановская база данных систематических обзоров. 3 (3): CD007176. Дои:10.1002 / 14651858.CD007176.pub2. HDL:10138/136201. PMID 22419320.
- ^ Эксперты утверждают, что исследование, посвященное рискам, связанным с антиоксидантами, витамином, основано на ошибочной методологии Выпуск новостей от Государственный университет Орегона опубликовано на ScienceDaily. Проверено 19 апреля 2007 г.
- ^ Миллер Э. Р., Пастор-Барриузо Р., Далал Д., Римерсма Р. А., Аппель Л. Дж., Гуаллар Э. (январь 2005 г.). «Мета-анализ: добавление высоких доз витамина Е может увеличить общую смертность». Анналы внутренней медицины. 142 (1): 37–46. Дои:10.7326/0003-4819-142-1-200501040-00110. PMID 15537682.
- ^ а б Bjelakovic G, Nagorni A, Nikolova D, Simonetti RG, Bjelakovic M, Gluud C (июль 2006 г.). «Мета-анализ: антиоксидантные добавки для первичной и вторичной профилактики колоректальной аденомы». Пищевая фармакология и терапия. 24 (2): 281–91. Дои:10.1111 / j.1365-2036.2006.02970.x. PMID 16842454. S2CID 20452618.
- ^ Кортес-Жофре, Марсела; Руэда, Хосе-Рамон; Асеньо-Лобос, Клаудиа; Мадрид, Ева; Bonfill Cosp, Ксавье (4 марта 2020 г.). «Препараты для профилактики рака легких у здоровых людей». Кокрановская база данных систематических обзоров. 3: CD002141. Дои:10.1002 / 14651858.CD002141.pub3. ISSN 1469-493X. ЧВК 7059884. PMID 32130738.
- ^ а б c Дэвис К.Дж. (1995). «Окислительный стресс: парадокс аэробной жизни». Симпозиум Биохимического общества. 61: 1–31. Дои:10.1042 / bss0610001. PMID 8660387.
- ^ а б c d е Sies H (март 1997 г.). «Окислительный стресс: оксиданты и антиоксиданты». Экспериментальная физиология. 82 (2): 291–5. Дои:10.1113 / expphysiol.1997.sp004024. PMID 9129943. S2CID 20240552.
- ^ а б c d Вертуани С., Ангусти А., Манфредини С. (2004). «Сеть антиоксидантов и про-антиоксидантов: обзор». Текущий фармацевтический дизайн. 10 (14): 1677–94. Дои:10.2174/1381612043384655. PMID 15134565.
- ^ Ри С.Г. (июнь 2006 г.). «Передача сигналов клетки. H2O2, необходимое зло для передачи сигналов клетки». Наука. 312 (5782): 1882–3. Дои:10.1126 / наука.1130481. PMID 16809515. S2CID 83598498.
- ^ Валко М., Лейбфриц Д., Монкол Дж., Кронин М.Т., Мазур М., Телсер Дж. (2007). «Свободные радикалы и антиоксиданты в нормальных физиологических функциях и болезнях человека». Международный журнал биохимии и клеточной биологии. 39 (1): 44–84. Дои:10.1016 / j.biocel.2006.07.001. PMID 16978905.
- ^ а б Stohs SJ, Bagchi D (февраль 1995 г.). «Окислительные механизмы при токсичности ионов металлов» (PDF). Свободная радикальная биология и медицина (Представлена рукопись). 18 (2): 321–36. CiteSeerX 10.1.1.461.6417. Дои:10.1016 / 0891-5849 (94) 00159-Н. PMID 7744317.
- ^ Накабеппу Ю., Сакуми К., Сакамото К., Цучимото Д., Цузуки Т., Накатсу Ю. (апрель 2006 г.). «Мутагенез и канцерогенез, вызванные окислением нуклеиновых кислот». Биологическая химия. 387 (4): 373–9. Дои:10.1515 / BC.2006.050. PMID 16606334. S2CID 20217256.
- ^ Валко М., Изакович М., Мазур М., Родос С.Дж., Телсер Дж. (Ноябрь 2004 г.). «Роль кислородных радикалов в повреждении ДНК и заболеваемости раком». Молекулярная и клеточная биохимия. 266 (1–2): 37–56. Дои:10.1023 / B: MCBI.0000049134.69131.89. PMID 15646026. S2CID 207547763.
- ^ Stadtman ER (август 1992 г.). «Окисление белков и старение». Наука. 257 (5074): 1220–4. Bibcode:1992Научный ... 257.1220S. Дои:10.1126 / science.1355616. PMID 1355616.
- ^ Раха С., Робинсон Б.Х. (октябрь 2000 г.). «Митохондрии, свободные радикалы кислорода, болезни и старение». Тенденции в биохимических науках. 25 (10): 502–8. Дои:10.1016 / S0968-0004 (00) 01674-1. PMID 11050436.
- ^ Леназ Г (2001). «Митохондриальная продукция активных форм кислорода: механизмы и последствия патологии человека». IUBMB Life. 52 (3–5): 159–64. Дои:10.1080/15216540152845957. PMID 11798028. S2CID 45366190.
- ^ Финкель Т., Холбрук, штат Нью-Джерси (ноябрь 2000 г.). «Окислители, окислительный стресс и биология старения». Природа. 408 (6809): 239–47. Bibcode:2000Натура.408..239F. Дои:10.1038/35041687. PMID 11089981. S2CID 2502238.
- ^ Херст Дж., Кинг М.С., Прайд КР (октябрь 2008 г.). «Производство активных форм кислорода комплексом I». Сделки Биохимического Общества. 36 (Pt 5): 976–80. Дои:10.1042 / BST0360976. PMID 18793173.
- ^ Seaver LC, Imlay JA (ноябрь 2004 г.). «Являются ли респираторные ферменты основным источником внутриклеточной перекиси водорода?». Журнал биологической химии. 279 (47): 48742–50. Дои:10.1074 / jbc.M408754200. PMID 15361522.
- ^ а б Имлай JA (2003). «Пути окислительного повреждения». Ежегодный обзор микробиологии. 57: 395–418. Дои:10.1146 / annurev.micro.57.030502.090938. PMID 14527285.
- ^ Деммиг-Адамс Б., Адамс В.В. (декабрь 2002 г.). «Антиоксиданты в фотосинтезе и питании человека». Наука. 298 (5601): 2149–53. Bibcode:2002Наука ... 298.2149D. Дои:10.1126 / science.1078002. PMID 12481128. S2CID 27486669.
- ^ Кригер-Лишкай А (январь 2005 г.). «Производство синглетного кислорода при фотосинтезе». Журнал экспериментальной ботаники. 56 (411): 337–46. CiteSeerX 10.1.1.327.9651. Дои:10.1093 / jxb / erh237. PMID 15310815.
- ^ Kupper FC, Carpenter LJ, McFiggans GB, Palmer CJ, Waite TJ, Boneberg EM, Woitsch S, Weiller M, Abela R, Grolimund D, Potin P, Butler A, Luther GW, Kroneck PMH, Meyer-Klaucke W, Feiters MC ( 2008 г.). «Накопление йодида обеспечивает водоросли неорганическим антиоксидантом, влияющим на химический состав атмосферы». Труды Национальной академии наук. 105 (19): 6954–6958. Bibcode:2008PNAS..105.6954K. Дои:10.1073 / pnas.0709959105. ISSN 0027-8424. ЧВК 2383960. PMID 18458346.
- ^ Сабо I, Бергантино Э., Джакометти GM (июль 2005 г.). «Световой и кислородный фотосинтез: рассеяние энергии как механизм защиты от фотоокисления». Отчеты EMBO. 6 (7): 629–34. Дои:10.1038 / sj.embor.7400460. ЧВК 1369118. PMID 15995679.
- ^ Kerfeld CA (октябрь 2004 г.). «Водорастворимые каротиноидные белки цианобактерий» (PDF). Архивы биохимии и биофизики (Представлена рукопись). 430 (1): 2–9. Дои:10.1016 / j.abb.2004.03.018. PMID 15325905.
- ^ Миллер Р.А., Бритиган Б.Е. (январь 1997 г.). «Роль оксидантов в патофизиологии микробов». Обзоры клинической микробиологии. 10 (1): 1–18. Дои:10.1128 / CMR.10.1.1. ЧВК 172912. PMID 8993856.
- ^ Шодьер Дж, Феррари-Илиу Р (1999). «Внутриклеточные антиоксиданты: от химических механизмов к биохимическим». Пищевая и химическая токсикология. 37 (9–10): 949–62. Дои:10.1016 / S0278-6915 (99) 00090-3. PMID 10541450.
- ^ Sies H (июль 1993 г.). «Стратегии антиоксидантной защиты». Европейский журнал биохимии / FEBS. 215 (2): 213–9. Дои:10.1111 / j.1432-1033.1993.tb18025.x. PMID 7688300.
- ^ Khaw KT, Woodhouse P (июнь 1995 г.). «Взаимосвязь витамина С, инфекции, гемостатических факторов и сердечно-сосудистых заболеваний». BMJ. 310 (6994): 1559–63. Дои:10.1136 / bmj.310.6994.1559. ЧВК 2549940. PMID 7787643.
- ^ а б c d Эвелсон П., Травацио М., Репетто М., Эскобар Дж., Ллезуи С., Лисси Э.А. (апрель 2001 г.). «Оценка общего реактивного антиоксидантного потенциала (TRAP) гомогенатов тканей и их цитозолей». Архивы биохимии и биофизики. 388 (2): 261–6. Дои:10.1006 / abbi.2001.2292. PMID 11368163.
- ^ Моррисон Дж. А., Якобсен Д. В., Спречер Д. Л., Робинсон К., Хури П., Дэниелс С. Р. (ноябрь 1999 г.). «Сывороточный глутатион у мужчин-подростков предсказывает у родителей ишемическую болезнь сердца». Тираж. 100 (22): 2244–7. Дои:10.1161 / 01.CIR.100.22.2244. PMID 10577998.
- ^ Тейхерт Дж., Прейсс Р. (ноябрь 1992 г.). «ВЭЖХ-методы определения липоевой кислоты и ее восстановленной формы в плазме крови человека». Международный журнал клинической фармакологии, терапии и токсикологии. 30 (11): 511–2. PMID 1490813.
- ^ Акиба С., Мацуго С., Пакер Л., Кониси Т. (май 1998 г.). «Анализ липоевой кислоты, связанной с белками, в тканях новым ферментативным методом». Аналитическая биохимия. 258 (2): 299–304. Дои:10.1006 / abio.1998.2615. PMID 9570844.
- ^ а б Гланцунис Г.К., Цимойаннис Э.С., Каппас А.М., Галарис Д.А. (2005). «Мочевая кислота и окислительный стресс». Текущий фармацевтический дизайн. 11 (32): 4145–51. Дои:10.2174/138161205774913255. PMID 16375736.
- ^ Эль-Сохеми А., Бейлин А., Кабагамбе Е., Аскерио А., Шпигельман Д., Кампос Н. (июль 2002 г.). «Индивидуальные концентрации каротиноидов в жировой ткани и плазме как биомаркеры диетического потребления». Американский журнал клинического питания. 76 (1): 172–9. Дои:10.1093 / ajcn / 76.1.172. PMID 12081831.
- ^ а б Соуэлл А. Л., Хафф Д. Л., Йегер П. Р., Кодилл С. П., Гюнтер Е. В. (март 1994 г.). «Ретинол, альфа-токоферол, лютеин / зеаксантин, бета-криптоксантин, ликопин, альфа-каротин, транс-бета-каротин и четыре ретиниловых эфира в сыворотке, определяемые одновременно обращенно-фазовой ВЭЖХ с многоволновым детектированием». Клиническая химия. 40 (3): 411–6. Дои:10.1093 / Clinchem / 40.3.411. PMID 8131277.[постоянная мертвая ссылка ]
- ^ Шталь В., Шварц В., Сандквист А. Р., Сис Н. (апрель 1992 г.). «цис-транс-изомеры ликопина и бета-каротина в сыворотке и тканях человека». Архивы биохимии и биофизики. 294 (1): 173–7. Дои:10.1016 / 0003-9861 (92) 90153-Н. PMID 1550343.
- ^ Зита С., Овервад К., Мортенсен С.А., Синдберг С.Д., Месгаард С., Хантер Д.А. (2003). «Концентрации коэнзима Q10 в сыворотке крови у здоровых мужчин с добавлением 30 или 100 мг кофермента Q10 в течение двух месяцев в рандомизированном контролируемом исследовании». БиоФакторы. 18 (1–4): 185–93. Дои:10.1002 / biof.5520180221. PMID 14695934. S2CID 19895215.
- ^ а б Турунен М., Олссон Дж., Даллнер Дж. (Январь 2004 г.). «Метаболизм и функция коэнзима Q». Biochimica et Biophysica Acta (BBA) - Биомембраны. 1660 (1–2): 171–99. Дои:10.1016 / j.bbamem.2003.11.012. PMID 14757233.
- ^ а б Эномото А, Эндоу Х (сентябрь 2005 г.). «Роль переносчиков органических анионов (ОАТ) и переносчика уратов (URAT1) в патофизиологии болезней человека». Клиническая и экспериментальная нефрология. 9 (3): 195–205. Дои:10.1007 / s10157-005-0368-5. PMID 16189627. S2CID 6145651.
- ^ а б Wu XW, Lee CC, Muzny DM, Caskey CT (декабрь 1989 г.). «Уратоксидаза: первичная структура и эволюционные последствия». Труды Национальной академии наук Соединенных Штатов Америки. 86 (23): 9412–6. Bibcode:1989PNAS ... 86.9412 Вт. Дои:10.1073 / пнас.86.23.9412. ЧВК 298506. PMID 2594778.
- ^ Wu XW, Muzny DM, Lee CC, Caskey CT (январь 1992 г.). «Два независимых мутационных события в потере уратоксидазы во время эволюции гоминоидов». Журнал молекулярной эволюции. 34 (1): 78–84. Bibcode:1992JMolE..34 ... 78 Вт. Дои:10.1007 / BF00163854. PMID 1556746. S2CID 33424555.
- ^ Альварес-Ларио Б., Макаррон-Висенте Дж. (Ноябрь 2010 г.). «Мочевая кислота и эволюция». Ревматология. 49 (11): 2010–5. Дои:10.1093 / ревматология / keq204. PMID 20627967.
- ^ а б Ватанабе С., Канг Д.Х., Фенг Л., Накагава Т., Канеллис Дж., Лан Х., Маццали М., Джонсон Р.Дж. (сентябрь 2002 г.). «Мочевая кислота, эволюция гоминоидов и патогенез солевой чувствительности». Гипертония. 40 (3): 355–60. Дои:10.1161 / 01.HYP.0000028589.66335.AA. PMID 12215479.
- ^ Джонсон Р.Дж., Эндрюс П., Беннер С.А., Оливер В. (2010). «Премия Теодора Э. Вудворда. Эволюция ожирения: выводы из среднего миоцена». Труды Американской клинической и климатологической ассоциации. 121: 295–305, обсуждение 305–8. ЧВК 2917125. PMID 20697570.
- ^ а б Бэйли Дж. К., Бейтс М. Г., Томпсон А. А., Уоринг В. С., Партридж Р. В., Шнопп М. Ф., Симпсон А., Гулливер-Слоан Ф., Максвелл С. Р., Уэбб Д. Д. (май 2007 г.). «Выработка эндогенных уратов увеличивает антиоксидантную способность плазмы у здоровых людей, живущих в низинах, находящихся на большой высоте». Грудь. 131 (5): 1473–8. Дои:10.1378 / сундук.06-2235. PMID 17494796.
- ^ Беккер Б.Ф. (июнь 1993 г.). «К физиологической функции мочевой кислоты». Свободная радикальная биология и медицина. 14 (6): 615–31. Дои:10.1016 / 0891-5849 (93) 90143-И. PMID 8325534.
- ^ а б Саутин Ю.Ю., Джонсон Р.Дж. (июнь 2008 г.). «Мочевая кислота: парадокс оксидант-антиоксидант». Нуклеозиды, нуклеотиды и нуклеиновые кислоты. 27 (6): 608–19. Дои:10.1080/15257770802138558. ЧВК 2895915. PMID 18600514.
- ^ Eggebeen AT (сентябрь 2007 г.). «Подагра: обновление». Американский семейный врач. 76 (6): 801–8. PMID 17910294.
- ^ Кэмпион EW, Глинн Р.Дж., ДеЛабри Л.О. (март 1987 г.). «Бессимптомная гиперурикемия. Риски и последствия в исследовании нормативного старения». Американский журнал медицины. 82 (3): 421–6. Дои:10.1016/0002-9343(87)90441-4. PMID 3826098.
- ^ Назаревич Р.Р., Циолковски В., Ваккаро П.С., Гафурифар П. (декабрь 2007 г.). «Влияние краткосрочной кетогенной диеты на окислительно-восстановительный статус крови человека». Исследования омоложения. 10 (4): 435–40. Дои:10.1089 / rej.2007.0540. PMID 17663642.
- ^ а б c d "Витамин С". Информационный центр по микронутриентам, Институт Линуса Полинга, Университет штата Орегон, Корваллис, Орегон. 1 июля 2018 г.. Получено 19 июн 2019.
- ^ Смирнов Н (2001). «Биосинтез L-аскорбиновой кислоты». Кофактор биосинтеза. Витамины и гормоны. 61. С. 241–66. Дои:10.1016 / S0083-6729 (01) 61008-2. ISBN 978-0-12-709861-6. PMID 11153268.
- ^ Линстер К.Л., Ван Шафтинген Э. (январь 2007 г.). «Витамин С. Биосинтез, переработка и разложение у млекопитающих». Журнал FEBS. 274 (1): 1–22. Дои:10.1111 / j.1742-4658.2006.05607.x. PMID 17222174. S2CID 21345196.
- ^ а б Мейстер А. (апрель 1994 г.). «Антиоксидантная система глутатион-аскорбиновая кислота у животных». Журнал биологической химии. 269 (13): 9397–400. PMID 8144521.
- ^ Уэллс WW, Сюй Д.П., Ян Ю.Ф., Рок, Пенсильвания (сентябрь 1990 г.). «Тиолтрансфераза (глутаредоксин) млекопитающих и протеиндисульфидизомераза обладают активностью дегидроаскорбатредуктазы». Журнал биологической химии. 265 (26): 15361–4. PMID 2394726.
- ^ Padayatty SJ, Katz A, Wang Y, Eck P, Kwon O, Lee JH, Chen S, Corpe C, Dutta A, Dutta SK, Levine M (февраль 2003 г.). «Витамин С как антиоксидант: оценка его роли в профилактике заболеваний». Журнал Американского колледжа питания. 22 (1): 18–35. Дои:10.1080/07315724.2003.10719272. PMID 12569111. S2CID 21196776.
- ^ Сигэока С., Исикава Т., Тамой М., Миягава Ю., Такеда Т., Ябута Ю., Йошимура К. (май 2002 г.). «Регуляция и функция изоферментов аскорбатпероксидазы». Журнал экспериментальной ботаники. 53 (372): 1305–19. Дои:10.1093 / jexbot / 53.372.1305. PMID 11997377.
- ^ Смирнов Н, Уиллер ГЛ (2000). «Аскорбиновая кислота в растениях: биосинтез и функции». Критические обзоры в биохимии и молекулярной биологии. 35 (4): 291–314. Дои:10.1080/10409230008984166. PMID 11005203. S2CID 85060539.
- ^ а б c d Мейстер А, Андерсон МЭ (1983). «Глутатион». Ежегодный обзор биохимии. 52: 711–60. Дои:10.1146 / annurev.bi.52.070183.003431. PMID 6137189.
- ^ Мейстер А. (ноябрь 1988 г.). «Метаболизм глутатиона и его селективная модификация». Журнал биологической химии. 263 (33): 17205–8. PMID 3053703.
- ^ Габалла А., Ньютон Г.Л., Антельманн Х., Парсонейдж Д., Аптон Х., Рават М., Клэйборн А., Фейи Р.К., Хельманн Д.Д. (апрель 2010 г.). «Биосинтез и функции бациллитиола, основного низкомолекулярного тиола в бациллах». Труды Национальной академии наук Соединенных Штатов Америки. 107 (14): 6482–6. Bibcode:2010PNAS..107.6482G. Дои:10.1073 / pnas.1000928107. ЧВК 2851989. PMID 20308541.
- ^ Newton GL, Rawat M, La Clair JJ, Jothivasan VK, Budiarto T, Hamilton CJ, Claiborne A, Helmann JD, Fahey RC (сентябрь 2009 г.). «Бацилтиол - тиол-антиоксидант, вырабатываемый бациллами». Природа Химическая Биология. 5 (9): 625–627. Дои:10.1038 / nchembio.189. ЧВК 3510479. PMID 19578333.
- ^ Fahey RC (2001). «Новые тиолы прокариот». Ежегодный обзор микробиологии. 55: 333–56. Дои:10.1146 / annurev.micro.55.1.333. PMID 11544359.
- ^ Fairlamb AH, Cerami A (1992). «Метаболизм и функции трипанотиона в кинетопластиде». Ежегодный обзор микробиологии. 46: 695–729. Дои:10.1146 / annurev.mi.46.100192.003403. PMID 1444271.
- ^ а б Эррера Э., Барбас С. (март 2001 г.). «Витамин Е: действие, метаболизм и перспективы». Журнал физиологии и биохимии. 57 (2): 43–56. Дои:10.1007 / BF03179812. PMID 11579997. S2CID 7272312.
- ^ Пакер Л., Вебер С.У., Римбах Г. (февраль 2001 г.). «Молекулярные аспекты антиоксидантного действия альфа-токотриенола и клеточной сигнализации». Журнал питания. 131 (2): 369С – 73С. Дои:10.1093 / jn / 131.2.369S. PMID 11160563.
- ^ а б Бригелиус-Флоэ Р., Трабер М.Г. (июль 1999 г.). «Витамин Е: функции и обмен веществ». Журнал FASEB. 13 (10): 1145–55. CiteSeerX 10.1.1.337.5276. Дои:10.1096 / fasebj.13.10.1145. PMID 10385606. S2CID 7031925.
- ^ Трабер М.Г., Аткинсон Дж. (Июль 2007 г.). «Витамин Е, антиоксидант и не более того». Свободная радикальная биология и медицина. 43 (1): 4–15. Дои:10.1016 / j.freeradbiomed.2007.03.024. ЧВК 2040110. PMID 17561088.
- ^ Ван Х, Куинн П.Дж. (июль 1999 г.). «Витамин Е и его функции в мембранах». Прогресс в исследованиях липидов. 38 (4): 309–36. Дои:10.1016 / S0163-7827 (99) 00008-9. PMID 10793887.
- ^ Зайлер А., Шнайдер М., Фёрстер Х, Рот С., Вирт Е. К., Калмзее С., Плеснила Н., Креммер Е., Радмарк О., Вурст В., Борнкамм Г. В., Швайцер Ю., Конрад М. (сентябрь 2008 г.). «Глутатионпероксидаза 4 воспринимает и переводит окислительный стресс в зависимую от 12/15 липоксигеназ и опосредованную AIF гибель клеток». Клеточный метаболизм. 8 (3): 237–48. Дои:10.1016 / j.cmet.2008.07.005. PMID 18762024.
- ^ Бригелиус-Флоэ Р., Дэвис К.Дж. (июль 2007 г.). «Является ли витамин E антиоксидантом, регулятором передачи сигналов и экспрессии генов, или является« нездоровой »пищей? Комментарии к двум сопроводительным документам:« Молекулярный механизм действия альфа-токоферола »А. Аззи и« Витамин E, антиоксидант и не более того "М. Трабера и Дж. Аткинсона". Свободная радикальная биология и медицина. 43 (1): 2–3. Дои:10.1016 / j.freeradbiomed.2007.05.016. PMID 17561087.
- ^ Аткинсон Дж., Эпанд РФ, Эпанд РМ (март 2008 г.). «Токоферолы и токотриенолы в мембранах: критический обзор». Свободная радикальная биология и медицина. 44 (5): 739–64. Дои:10.1016 / j.freeradbiomed.2007.11.010. PMID 18160049.
- ^ Аззи А (июль 2007 г.). «Молекулярный механизм действия альфа-токоферола». Свободная радикальная биология и медицина. 43 (1): 16–21. Дои:10.1016 / j.freeradbiomed.2007.03.013. PMID 17561089.
- ^ Zingg JM, Azzi A (май 2004 г.). «Неантиоксидантная активность витамина Е». Современная лекарственная химия. 11 (9): 1113–33. Дои:10.2174/0929867043365332. PMID 15134510. Архивировано из оригинал 6 октября 2011 г.
- ^ Сен С.К., Ханна С., Рой С. (март 2006 г.). «Токотриенолы: витамин Е помимо токоферолов». Науки о жизни. 78 (18): 2088–98. Дои:10.1016 / j.lfs.2005.12.001. ЧВК 1790869. PMID 16458936.
- ^ Дуарте Т.Л., Лунек Дж. (Июль 2005 г.). «Обзор: Когда антиоксидант не является антиоксидантом? Обзор новых действий и реакций витамина С». Свободные радикальные исследования. 39 (7): 671–86. Дои:10.1080/10715760500104025. PMID 16036346. S2CID 39962659.
- ^ а б Карр А., Фрей Б. (июнь 1999 г.). «Действует ли витамин С как прооксидант в физиологических условиях?». Журнал FASEB. 13 (9): 1007–24. Дои:10.1096 / fasebj.13.9.1007. PMID 10336883. S2CID 15426564.
- ^ Шнайдер С. (январь 2005 г.). «Химия и биология витамина Е». Молекулярное питание и пищевые исследования. 49 (1): 7–30. Дои:10.1002 / mnfr.200400049. PMID 15580660.
- ^ Холливелл Б. (август 2008 г.). «Полифенолы являются антиоксидантами или прооксидантами? Что мы узнаем из клеточных культур и исследований in vivo?». Архивы биохимии и биофизики. 476 (2): 107–112. Дои:10.1016 / j.abb.2008.01.028. PMID 18284912.
- ^ Ристоу М., Зарсе К. (июнь 2010 г.). «Как повышенный окислительный стресс способствует долголетию и метаболическому здоровью: концепция митохондриального гормезиса (митохондриального гормезиса)». Экспериментальная геронтология. 45 (6): 410–418. Дои:10.1016 / j.exger.2010.03.014. PMID 20350594. S2CID 207727334.
- ^ а б Хо Ю.С., Магненат Дж. Л., Гаргано М., Цао Дж. (Октябрь 1998 г.). «Природа механизмов антиоксидантной защиты: урок трансгенных исследований». Перспективы гигиены окружающей среды. 106 Дополнение 5 (Дополнение 5): 1219–28. Дои:10.2307/3433989. JSTOR 3433989. ЧВК 1533365. PMID 9788901.
- ^ Зелко И.Н., Мариани Т.Дж., Фольц Р.Дж. (август 2002 г.). «Мультигенное семейство супероксиддисмутазы: сравнение структур, эволюции и экспрессии генов CuZn-SOD (SOD1), Mn-SOD (SOD2) и EC-SOD (SOD3)». Свободная радикальная биология и медицина. 33 (3): 337–49. Дои:10.1016 / S0891-5849 (02) 00905-X. PMID 12126755.
- ^ а б Баннистер СП, Баннистер WH, Ротилио Дж. (1987). «Аспекты структуры, функции и применения супероксиддисмутазы». CRC Критические обзоры в биохимии. 22 (2): 111–80. Дои:10.3109/10409238709083738. PMID 3315461.
- ^ Джонсон Ф, Джуливи С (2005). «Супероксиддисмутазы и их влияние на здоровье человека». Молекулярные аспекты медицины. 26 (4–5): 340–52. Дои:10.1016 / j.mam.2005.07.006. PMID 16099495.
- ^ Нозик-Грейк Э., Сулиман Х. Б., Пиантадози, Калифорния (декабрь 2005 г.). «Внеклеточная супероксиддисмутаза». Международный журнал биохимии и клеточной биологии. 37 (12): 2466–71. Дои:10.1016 / j.biocel.2005.06.012. PMID 16087389.
- ^ Мелов С., Шнайдер Дж. А., Дэй Б. Дж., Хинерфельд Д., Коскун П., Мирра С. С., Крапо Дж. Д., Уоллес, округ Колумбия (февраль 1998 г.). «Новый неврологический фенотип у мышей, лишенных митохондриальной супероксиддисмутазы марганца». Природа Генетика. 18 (2): 159–63. Дои:10.1038 / ng0298-159. PMID 9462746. S2CID 20843002.
- ^ Reaume AG, Elliott JL, Hoffman EK, Kowall NW, Ferrante RJ, Siwek DF, Wilcox HM, Flood DG, Beal MF, Brown RH, Scott RW, Snider WD (май 1996 г.). «Моторные нейроны у мышей с дефицитом супероксиддисмутазы Cu / Zn развиваются нормально, но демонстрируют повышенную гибель клеток после повреждения аксонов». Природа Генетика. 13 (1): 43–7. Дои:10.1038 / ng0596-43. PMID 8673102. S2CID 13070253.
- ^ Ван Кэмп В., Инзе Д., Ван Монтегю М. (1997). «Регуляция и функция супероксиддисмутаз табака». Свободная радикальная биология и медицина. 23 (3): 515–20. Дои:10.1016 / S0891-5849 (97) 00112-3. PMID 9214590.
- ^ Chelikani P, Fita I, Loewen PC (январь 2004 г.). «Разнообразие структур и свойств каталаз» (PDF). Клеточные и молекулярные науки о жизни (Представлена рукопись). 61 (2): 192–208. Дои:10.1007 / s00018-003-3206-5. HDL:10261/111097. PMID 14745498. S2CID 4411482.
- ^ Замоцки М, Коллер Ф (1999). «Понимание структуры и функции каталаз: ключи от молекулярной эволюции и мутагенеза in vitro». Прогресс в биофизике и молекулярной биологии. 72 (1): 19–66. Дои:10.1016 / S0079-6107 (98) 00058-3. PMID 10446501.
- ^ дель Рио, Л. М., Сандалио Л. М., Пальма Дж. М., Буэно П., Корпас Ф. Дж. (ноябрь 1992 г.). «Метаболизм кислородных радикалов в пероксисомах и клеточные проявления». Свободная радикальная биология и медицина. 13 (5): 557–80. Дои:10.1016 / 0891-5849 (92) 90150-Ф. PMID 1334030.
- ^ Хайнер А.Н., Рэйвен Э.Л., Торнели Р.Н., Гарсия-Кановас Ф., Родригес-Лопес Дж. Н. (июль 2002 г.). «Механизмы образования соединения I в пероксидазах гема». Журнал неорганической биохимии. 91 (1): 27–34. Дои:10.1016 / S0162-0134 (02) 00390-2. PMID 12121759.
- ^ Мюллер С., Ридель HD, Стреммель В. (декабрь 1997 г.). «Прямые доказательства того, что каталаза является преобладающим ферментом, удаляющим H2O2, в эритроцитах человека». Кровь. 90 (12): 4973–8. Дои:10.1182 / кровь.V90.12.4973. PMID 9389716.
- ^ Огата М (февраль 1991 г.). «Акаталаземия». Генетика человека. 86 (4): 331–40. Дои:10.1007 / BF00201829. PMID 1999334.
- ^ Parsonage D, Youngblood D, Sarma G, Wood Z, Karplus P, Poole L (2005). «Анализ связи между ферментативной активностью и олигомерным состоянием в AhpC, бактериальном пероксиредоксине». Биохимия. 44 (31): 10583–92. Дои:10.1021 / bi050448i. ЧВК 3832347. PMID 16060667. PDB 1YEX
- ^ Ри С.Г., Чае Х.З., Ким К. (июнь 2005 г.). «Пероксиредоксины: исторический обзор и предварительный просмотр новых механизмов и новых концепций в передаче сигналов в клетке». Свободная радикальная биология и медицина. 38 (12): 1543–52. Дои:10.1016 / j.freeradbiomed.2005.02.026. PMID 15917183.
- ^ Вуд З.А., Шредер Э., Робин Харрис Дж., Пул Л.Б. (январь 2003 г.). «Структура, механизм и регуляция пероксиредоксинов». Тенденции в биохимических науках. 28 (1): 32–40. Дои:10.1016 / S0968-0004 (02) 00003-8. PMID 12517450.
- ^ Клэйборн А., Йе Дж. И., Маллет Т. К., Люба Дж., Крейн Е. Дж., Чарриер В., Парсонейдж Д. (ноябрь 1999 г.). «Белок-сульфеновые кислоты: разнообразные роли маловероятного игрока в ферментативном катализе и окислительно-восстановительном регулировании». Биохимия. 38 (47): 15407–16. Дои:10.1021 / bi992025k. PMID 10569923.
- ^ Йонссон Т.Дж., Лоутер В.Т. (2007). «Белки восстановления пероксиредоксина». Системы пероксиредоксина. Субклеточная биохимия. 44. С. 115–41. Дои:10.1007/978-1-4020-6051-9_6. ISBN 978-1-4020-6050-2. ЧВК 2391273. PMID 18084892.
- ^ Neumann CA, Krause DS, Carman CV, Das S, Dubey DP, Abraham JL, Bronson RT, Fujiwara Y, Orkin SH, Van Etten RA (июль 2003 г.). «Важная роль пероксиредоксина Prdx1 в антиоксидантной защите эритроцитов и подавлении опухоли» (PDF). Природа. 424 (6948): 561–5. Bibcode:2003Натура.424..561Н. Дои:10.1038 / природа01819. PMID 12891360. S2CID 3570549.
- ^ Lee TH, Kim SU, Yu SL, Kim SH, Park DS, Moon HB, Dho SH, Kwon KS, Kwon HJ, Han YH, Jeong S, Kang SW, Shin HS, Lee KK, Rhee SG, Yu DY (июнь 2003 г. ). «Пероксиредоксин II необходим для поддержания продолжительности жизни эритроцитов у мышей». Кровь. 101 (12): 5033–8. Дои:10.1182 / кровь-2002-08-2548. PMID 12586629.
- ^ Дитц К.Дж., Джейкоб С., Эльз М.Л., Лакса М., Тогнетти В., де Миранда С.М., Байер М., Финкемайер И. (2006). «Функция пероксиредоксинов в окислительно-восстановительном метаболизме органелл растений». Журнал экспериментальной ботаники. 57 (8): 1697–709. Дои:10.1093 / jxb / erj160. PMID 16606633.
- ^ Нордберг Дж., Арнер Э.С. (декабрь 2001 г.). «Активные формы кислорода, антиоксиданты и тиоредоксиновая система млекопитающих». Свободная радикальная биология и медицина. 31 (11): 1287–312. Дои:10.1016 / S0891-5849 (01) 00724-9. PMID 11728801.
- ^ Виейра душ Сантуш С., Рей П. (июль 2006 г.). «Тиоредоксины растений являются ключевыми участниками реакции на окислительный стресс». Тенденции в растениеводстве. 11 (7): 329–34. Дои:10.1016 / j.tplants.2006.05.005. PMID 16782394.
- ^ Арнер Э.С., Хольмгрен А. (октябрь 2000 г.). «Физиологические функции тиоредоксина и тиоредоксинредуктазы». Европейский журнал биохимии / FEBS. 267 (20): 6102–9. Дои:10.1046 / j.1432-1327.2000.01701.x. PMID 11012661.
- ^ Mustacich D, Powis G (февраль 2000 г.). «Тиоредоксинредуктаза». Биохимический журнал. 346 (1): 1–8. Дои:10.1042/0264-6021:3460001. ЧВК 1220815. PMID 10657232.
- ^ Creissen G, Broadbent P, Stevens R, Wellburn AR, Mullineaux P (май 1996). «Манипуляция метаболизмом глутатиона в трансгенных растениях». Сделки Биохимического Общества. 24 (2): 465–9. Дои:10.1042 / bst0240465. PMID 8736785.
- ^ Бригелиус-Флоэ Р. (ноябрь 1999 г.). «Тканевые функции отдельных пероксидаз глутатиона». Свободная радикальная биология и медицина. 27 (9–10): 951–65. Дои:10.1016 / S0891-5849 (99) 00173-2. PMID 10569628.
- ^ Хо Ю.С., Магненат Дж. Л., Бронсон Р. Т., Цао Дж., Гаргано М., Сугавара М., Funk CD (июнь 1997 г.). «Мыши с дефицитом клеточной глутатионпероксидазы развиваются нормально и не проявляют повышенной чувствительности к гипероксии». Журнал биологической химии. 272 (26): 16644–51. Дои:10.1074 / jbc.272.26.16644. PMID 9195979.
- ^ de Haan JB, Bladier C, Griffiths P, Kelner M, O'Shea RD, Cheung NS, Bronson RT, Silvestro MJ, Wild S, Zheng SS, Beart PM, Hertzog PJ, Kola I (август 1998 г.). «Мыши с гомозиготной нулевой мутацией по самой распространенной глутатионпероксидазе, Gpx1, демонстрируют повышенную чувствительность к параквату и перекиси водорода, вызывающим окислительный стресс». Журнал биологической химии. 273 (35): 22528–36. Дои:10.1074 / jbc.273.35.22528. PMID 9712879.
- ^ Шарма Р., Ян Й., Шарма А., Авасти С., Авасти Ю. К. (апрель 2004 г.). «Антиоксидантная роль глутатион-S-трансфераз: защита от токсичности оксидантов и регуляция стресс-опосредованного апоптоза». Антиоксиданты и редокс-сигналы. 6 (2): 289–300. Дои:10.1089/152308604322899350. PMID 15025930.
- ^ Hayes JD, Flanagan JU, Jowsey IR (2005). «Трансферазы глутатиона». Ежегодный обзор фармакологии и токсикологии. 45: 51–88. Дои:10.1146 / annurev.pharmtox.45.120403.095857. PMID 15822171.
- ^ Кадер А.А., Загоры Д., Кербель Е.Л. (1989). «Упаковка фруктов и овощей в модифицированной атмосфере». Критические обзоры в области пищевой науки и питания. 28 (1): 1–30. Дои:10.1080/10408398909527490. PMID 2647417.
- ^ Заллен Е.М., Хичкок М.Дж., Гертц Г.Е. (декабрь 1975 г.). «Системы охлажденного питания. Влияние выдержки при охлаждении на качество говяжьего хлеба». Журнал Американской диетической ассоциации. 67 (6): 552–7. PMID 1184900.
- ^ Айверсон Ф (июнь 1995 г.). «Фенольные антиоксиданты: исследования отделения здравоохранения по бутилированному гидроксианизолу». Письма о раке. 93 (1): 49–54. Дои:10.1016 / 0304-3835 (95) 03787-В. PMID 7600543.
- ^ "Индекс номера E". Британский кулинарный гид. В архиве из оригинала 4 марта 2007 г.. Получено 5 марта 2007.
- ^ Робардс К., Керр А.Ф., Патсалидес Э. (февраль 1988 г.). «Прогорклость и ее измерение в пищевых маслах и закусках. Обзор». Аналитик. 113 (2): 213–24. Bibcode:1988Ана ... 113..213R. Дои:10.1039 / an9881300213. PMID 3288002.
- ^ Дель Карло М., Саккетти Дж., Ди Маттиа С., Компаньоне Д., Мастрокола Д., Либераторе Л., Чичелли А. (июнь 2004 г.). «Вклад фенольной фракции в антиоксидантную активность и окислительную стабильность оливкового масла». Журнал сельскохозяйственной и пищевой химии. 52 (13): 4072–9. Дои:10.1021 / jf049806z. PMID 15212450.
- ^ Бузер CE, Хаммонд GS, Гамильтон CE, Сен JN (1955). «Окисление углеводородов воздухом.1II. Стехиометрия и судьба ингибиторов в бензоле и хлорбензоле». Журнал Американского химического общества. 77 (12): 3233–7. Дои:10.1021 / ja01617a026.
- ^ «Мировой рынок антиоксидантов (природных и синтетических) может вырасти с 2,25 млрд долларов США в 2014 году до 3,25 млрд долларов США к 2020 году, при этом среднегодовой рост составит 5,5%». GlobalNewswire, Эль-Сегундо, Калифорния. 19 января 2016 г.. Получено 30 января 2017.
- ^ "Зачем использовать антиоксиданты?". SpecialChem клеи. Архивировано из оригинал 11 февраля 2007 г.. Получено 27 февраля 2007.
- ^ а б «Топливные антиоксиданты». Innospec Chemicals. Архивировано из оригинал 15 октября 2006 г.. Получено 27 февраля 2007.
- ^ Родригес-Амая DB (2003). «Пищевые каротиноиды: анализ, состав и изменения при хранении и переработке пищевых продуктов». Форум питания. 56: 35–7. PMID 15806788.
- ^ Maiani G, Castón MJ, Catasta G, Toti E, Cambrodón IG, Bysted A, Granado-Lorencio F, Olmedilla-Alonso B, Knuthsen P, Valoti M, Böhm V, Mayer-Miebach E, Behsnilian D, Schlemmer U (сентябрь 2009 г.) ). «Каротиноиды: актуальные знания об источниках пищи, потреблении, стабильности и биодоступности, а также их защитной роли в организме человека». Молекулярное питание и пищевые исследования. 53 Дополнение 2: S194–218. Дои:10.1002 / mnfr.200800053. HDL:10261/77697. PMID 19035552. Архивировано из оригинал 27 сентября 2018 г.. Получено 18 апреля 2017.
- ^ Генри CJ, Heppell N (февраль 2002 г.). «Питательные потери и выгоды во время обработки: будущие проблемы и проблемы». Труды Общества питания. 61 (1): 145–8. Дои:10.1079 / PNS2001142. PMID 12002789.
- ^ «Антиоксиданты и профилактика рака: информационный бюллетень». Национальный институт рака. В архиве из оригинала 4 марта 2007 г.. Получено 27 февраля 2007.
- ^ Ортега Р. (декабрь 2006 г.). «Важность функциональных продуктов в средиземноморской диете». Питание для общественного здравоохранения. 9 (8A): 1136–40. Дои:10.1017 / S1368980007668530. PMID 17378953.
- ^ Витчи А., Редди С., Стофер Б., Лаутербург Б. Х. (1992). «Системная доступность орального глутатиона». Европейский журнал клинической фармакологии. 43 (6): 667–9. Дои:10.1007 / BF02284971. PMID 1362956. S2CID 27606314.
- ^ Флэгг EW, Коутс RJ, Эли JW, Джонс Д.П., Гюнтер EW, Байерс TE, Блок GS, Гринберг RS (1994). «Потребление глутатиона с пищей у людей и взаимосвязь между потреблением и уровнем общего глутатиона в плазме». Питание и рак. 21 (1): 33–46. Дои:10.1080/01635589409514302. PMID 8183721.
- ^ Додд С., Дин О., Кополов Д.Л., Малхи Г.С., Берк М. (декабрь 2008 г.). «N-ацетилцистеин для антиоксидантной терапии: фармакология и клиническое применение». Мнение эксперта по биологической терапии. 8 (12): 1955–62. Дои:10.1517/14728220802517901. PMID 18990082. S2CID 74736842.
- ^ ван де Полл М.С., Деджонг С.Х., Сотерс ПБ (июнь 2006 г.). «Адекватный диапазон серосодержащих аминокислот и биомаркеров их избытка: уроки энтерального и парентерального питания». Журнал питания. 136 (6 Прил.): 1694S – 1700S. Дои:10.1093 / jn / 136.6.1694S. PMID 16702341.
- ^ Цао Г., Алессио Х. М., Катлер Р. Г. (март 1993 г.). «Анализ способности антиоксидантов к поглощению кислородных радикалов». Свободная радикальная биология и медицина. 14 (3): 303–11. Дои:10.1016 / 0891-5849 (93) 90027-П. PMID 8458588.
- ^ Ou B, Hampsch-Woodill M, Prior RL (октябрь 2001 г.). «Разработка и проверка улучшенного анализа способности поглощения радикалов кислорода с использованием флуоресцеина в качестве флуоресцентного зонда». Журнал сельскохозяйственной и пищевой химии. 49 (10): 4619–26. Дои:10.1021 / jf010586o. PMID 11599998.
- ^ а б «Изъято: способность поглощать кислородные радикалы (ORAC) отобранных продуктов, выпуск 2 (2010 г.)». Министерство сельского хозяйства США, Служба сельскохозяйственных исследований. 16 мая 2012. Получено 13 июн 2012.
- ^ Prior RL, Wu X, Schaich K (май 2005 г.). «Стандартизованные методы определения антиоксидантной способности и фенольных соединений в пищевых продуктах и диетических добавках» (PDF). Журнал сельскохозяйственной и пищевой химии. 53 (10): 4290–302. Дои:10.1021 / jf0502698. PMID 15884874. Архивировано из оригинал (PDF) 29 декабря 2016 г.. Получено 24 октября 2017.
- ^ Benzie IF (сентябрь 2003 г.). «Эволюция диетических антиоксидантов». Сравнительная биохимия и физиология A. 136 (1): 113–26. Дои:10.1016 / S1095-6433 (02) 00368-9. HDL:10397/34754. PMID 14527634.
- ^ Мэттил Х.А. (1947). «Антиоксиданты». Ежегодный обзор биохимии. 16: 177–92. Дои:10.1146 / annurev.bi.16.070147.001141. PMID 20259061.
- ^ Немецкий JB (1999). «Пищевая промышленность и окисление липидов». Влияние обработки на безопасность пищевых продуктов. Успехи экспериментальной медицины и биологии. 459. С. 23–50. Дои:10.1007/978-1-4615-4853-9_3. ISBN 978-0-306-46051-7. PMID 10335367.
- ^ Джейкоб Р.А. (1996). Три эпохи открытия витамина С. Субклеточная биохимия. 25. С. 1–16. Дои:10.1007/978-1-4613-0325-1_1. ISBN 978-1-4613-7998-0. PMID 8821966.
- ^ Рыцарь JA (1998). «Свободные радикалы: их история и текущее состояние при старении и болезнях». Анналы клинической и лабораторной науки. 28 (6): 331–46. PMID 9846200.
- ^ Moureu C, Dufraisse C (1922). "Sur l'autoxydation: Les antioxygènes". Comptes Rendus des Séances et Mémoires de la Société de Biologie (На французском). 86: 321–322.
- ^ Вольф G (март 2005 г.). «Открытие антиоксидантной функции витамина Е: вклад Генри А. Маттилла». Журнал питания. 135 (3): 363–6. Дои:10.1093 / jn / 135.3.363. PMID 15735064.
дальнейшее чтение
- Холливелл, Барри. и Джон М. К. Гаттеридж, Свободные радикалы в биологии и медицине (Oxford University Press, 2007), ISBN 0-19-856869-X
- Лейн, Ник, Кислород: молекула, создавшая мир (Oxford University Press, 2003), ISBN 0-19-860783-0
- Покорный Ян, Нелли Янислива и Майкл Х. Гордон, Антиоксиданты в пищевых продуктах: практическое применение (CRC Press, 2001), ISBN 0-8493-1222-1
внешняя ссылка
 СМИ, связанные с Антиоксиданты в Wikimedia Commons
СМИ, связанные с Антиоксиданты в Wikimedia Commons