Супероксиддисмутаза - Superoxide dismutase
| Супероксиддисмутаза | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Структура тетрамера супероксиддисмутазы 2 человека Mn.[1] | |||||||||
| Идентификаторы | |||||||||
| Номер ЕС | 1.15.1.1 | ||||||||
| Количество CAS | 9054-89-1 | ||||||||
| Базы данных | |||||||||
| IntEnz | Просмотр IntEnz | ||||||||
| БРЕНДА | BRENDA запись | ||||||||
| ExPASy | Просмотр NiceZyme | ||||||||
| КЕГГ | Запись в KEGG | ||||||||
| MetaCyc | метаболический путь | ||||||||
| ПРИАМ | профиль | ||||||||
| PDB структуры | RCSB PDB PDBe PDBsum | ||||||||
| Генная онтология | AmiGO / QuickGO | ||||||||
| |||||||||
Супероксиддисмутаза (SOD, EC 1.15.1.1 ) является фермент что поочередно катализирует дисмутация (или разделение) супероксид (O2−) радикальный в обычные молекулярные кислород (O2) и пероксид водорода (ЧАС2О2). Супероксид образуется как побочный продукт метаболизма кислорода и, если его не регулировать, вызывает многие типы повреждения клеток.[2] Перекись водорода также разрушает и разрушается другими ферментами, такими как каталаза. Таким образом, СОД является важным антиоксидант защита почти всех живых клеток, подвергающихся воздействию кислорода. Одно исключение Lactobacillus plantarum и связанные лактобациллы, которые используют другой механизм для предотвращения повреждений от реактивного O2−.
Химическая реакция
SOD катализируют непропорциональность супероксида:
- 2 НО2 → O2 + H2О2
Таким образом, O2− превращается в два менее вредных вида.
Путь, по которому катализируется SOD дисмутация из супероксид для Cu, Zn СОД можно записать со следующими реакциями:
- Cu2+-SOD + O2− → Cu+-SOD + O2 (восстановление меди; окисление супероксида)
- Cu+-SOD + O2− + 2H+ → Cu2+-SOD + H2О2 (окисление меди; восстановление супероксида)
Общая форма, применимая ко всем различным формам SOD, скоординированным с металлами, может быть записана следующим образом:
- M(п + 1) +-SOD + O2− → Mп +-SOD + O2
- Mп +-SOD + O2− + 2H+ → M(п + 1) +-SOD + H2О2.
где M = Cu (n = 1); Mn (n = 2); Fe (n = 2); Ni (n = 2).
В серии таких реакций степень окисления и заряд металла катион колеблется от n до n + 1: от +1 до +2 для меди или от +2 до +3 для других металлов.
Типы
Общий
Ирвин Фридович и Джо МакКорд в Университет Дьюка открыл ферментативную активность супероксиддисмутазы в 1968 году.[3] SOD ранее были известны как группа металлопротеины с неизвестной функцией; например, CuZnSOD был известен как эритрокупреин (или гемокупреин, или цито-купреин) или как ветеринарный противовоспалительный препарат «Орготеин».[4] Точно так же Брюэр (1967) идентифицировал белок, который позже стал известен как супероксиддисмутаза как индофенолоксидаза, путем анализа белков крахмальных гелей с использованием феназин-тетразолиевого метода.[5]
Существует три основных семейства супероксиддисмутазы, в зависимости от складки белка и металла. кофактор: тип Cu / Zn (который связывает оба медь и цинк ), Fe и Mn (которые связывают либо утюг или же марганец ), и типа Ni (связывающего никель ).
 Ленточная диаграмма бычьей Cu-Zn-субъединицы СОД[6] |  Активный центр человеческого марганца SOD, марганец показан фиолетовым цветом[7] |  Mn-SOD против димеров Fe-SOD |
- Медь и цинк - чаще всего используются эукариоты, включая людей. В цитозоли практически всех эукариотический клетки содержат фермент СОД с медь и цинк (Cu-Zn-SOD). Например, коммерчески доступный Cu-Zn-SOD обычно очищается от бычьих красных кровяных телец. Фермент Cu-Zn крупного рогатого скота представляет собой гомодимер с молекулярной массой 32 500. Это была первая СОД, кристаллическая структура которой была решена в 1975 году.[8] Это 8-ниточный "Греческий ключ «бета-цилиндр, с активным центром, удерживаемым между цилиндром и двумя поверхностными петлями. Две субъединицы плотно соединены спина к спине, в основном за счет гидрофобных и некоторых электростатических взаимодействий. Лигандов меди и цинка шесть гистидин и один аспартат боковые цепи; один гистидин связан между двумя металлами.[9]
- Железо или марганец - используется прокариоты и протисты, И в митохондрии и хлоропласты
 Активный центр супероксиддисмутазы железа
Активный центр супероксиддисмутазы железа- Железо. Многие бактерии содержат фермент в форме утюг (Fe-СОД); некоторые бактерии содержат Fe-SOD, другие Mn-SOD, а некоторые (например, Кишечная палочка ) содержат оба. Fe-SOD также можно найти в хлоропласты растений. Трехмерные структуры гомологичных супероксиддисмутаз Mn и Fe имеют одинаковое расположение альфа-спиралей, а их активные центры содержат одинаковый тип и расположение боковых цепей аминокислот. Обычно это димеры, но иногда и тетрамеры.
- Марганец - почти все митохондрии, и много бактерии, содержат форму с марганец (Mn-SOD): например, Mn-SOD, обнаруженный в митохондриях человека. Лиганды ионов марганца - 3 гистидин боковые цепи, аспартат боковая цепь и молекула воды или гидрокси лиганд в зависимости от степени окисления Mn (соответственно II и III).[10]
- Никель - прокариотический. Он имеет гексамерную (6-копийную) структуру, построенную из правых 4-спиральных пучков, каждый из которых содержит N-концевые крючки, которые хелатируют ион Ni. Ni-крючок содержит мотив His-Cys-X-X-Pro-Cys-Gly-X-Tyr; он обеспечивает большинство взаимодействий, критических для связывания металлов и катализа, и, следовательно, является вероятным диагностическим средством NiSOD.[11][12]

|
|
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

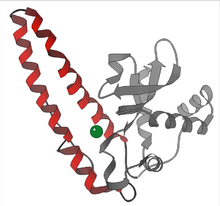


У высших растений изоферменты СОД локализованы в разных клеточных компартментах. Mn-SOD присутствует в митохондриях и пероксисомы. Fe-SOD был обнаружен в основном в хлоропластах, но также был обнаружен в пероксисомах, а CuZn-SOD был локализован в цитозоль, хлоропласты, пероксисомы и апопласт.[14][15]
Человек
У человека присутствуют три формы супероксиддисмутазы, во всех остальных млекопитающие, и большинство хордовые. SOD1 расположен в цитоплазма, SOD2 в митохондрии, и SOD3 является внеклеточный. Первый - это димер (состоит из двух единиц), а остальные - тетрамеры (четыре субъединицы). SOD1 и SOD3 содержат медь и цинк, тогда как SOD2, митохондриальный фермент, имеет марганец в его реактивном центре. В гены расположены на хромосомах 21, 6 и 4 соответственно (21q22.1, 6q25.3 и 4p15.3-p15.1).
|
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
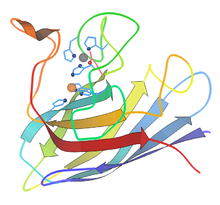


Растения
У высших растений ферменты супероксиддисмутазы (СОД) действуют как антиоксиданты и защищают клеточные компоненты от окисления. активные формы кислорода (ROS).[18] АФК могут образовываться в результате засухи, травм, гербицидов и пестицидов, озона, метаболической активности растений, дефицита питательных веществ, фотоингибирования, температуры над и под землей, токсичных металлов, а также ультрафиолетовых или гамма-лучей.[19][20] Если быть точным, молекулярный O2 сводится к O2− (АФК, называемая супероксидом), когда она поглощает возбужденный электрон, высвобождаемый из соединений электронной транспортной цепи. Известно, что супероксид денатурирует ферменты, окисляет липиды и фрагментирует ДНК.[19] SOD катализируют производство O2 и H2О2 из супероксида (O2−), что приводит к менее вредным реагентам.
При адаптации к повышенным уровням окислительного стресса концентрация СОД обычно увеличивается с увеличением степени стрессовых условий. Разделение различных форм СОД по всему растению позволяет им очень эффективно противодействовать стрессу. Есть три хорошо известных и изученных класса металлических коферментов СОД, которые существуют в растениях. Во-первых, СОД Fe состоят из двух видов, одного гомодимера (содержащего 1-2 г Fe) и одного тетрамера (содержащего 2-4 г Fe). Считается, что они являются самыми древними металлоферментами СОД и обнаруживаются как у прокариот, так и у эукариот. Fe-СОД чаще всего локализуются внутри хлоропластов растений, где они являются коренными. Во-вторых, СОД Mn состоят из гомодимеров и гомотетрамеров, каждая из которых содержит один атом Mn (III) на субъединицу. Они находятся преимущественно в митохондриях и пероксисомах. В-третьих, Cu-Zn SOD имеют электрические свойства, сильно отличающиеся от свойств двух других классов. Они сосредоточены в хлоропласт, цитозоль, а в некоторых случаях внеклеточное пространство. Обратите внимание, что Cu-Zn SOD обеспечивают меньшую защиту, чем Fe SOD, когда они локализованы в хлоропласте.[18][19][20]
Бактерии
Лейкоциты человека используют такие ферменты, как НАДФН оксидаза для выработки супероксида и других активных форм кислорода для уничтожения бактерий. Во время инфекции некоторые бактерии (например, Burkholderia pseudomallei ) поэтому продуцируют супероксиддисмутазу, чтобы защитить себя от гибели.[21]
Биохимия
СОД превосходит повреждающие реакции супероксида, защищая клетку от токсичности супероксида. Реакция супероксида с нерадикалами происходит запрещено вращать. В биологических системах это означает, что его основные реакции происходят с самим собой (дисмутация) или с другим биологическим радикалом, таким как оксид азота (НЕТ) или с металлом переходного ряда. Супероксид-анион-радикал (O2−) самопроизвольно уходит на O2 и перекись водорода (H2О2) довольно быстро (~ 105 M−1s−1 при pH 7).[нужна цитата ] СОД необходима, потому что супероксид реагирует с чувствительными и критическими клеточными мишенями. Например, он реагирует с радикалом NO и делает токсичный пероксинитрит.
Поскольку некаталитическая реакция дисмутации супероксида требует, чтобы две молекулы супероксида реагировали друг с другом, скорость дисмутации является вторым порядком по отношению к начальной концентрации супероксида. Таким образом, период полураспада супероксида, хотя и очень короткий при высоких концентрациях (например, 0,05 секунды при 0,1 мМ), на самом деле довольно велик при низких концентрациях (например, 14 часов при 0,1 нМ). Напротив, реакция супероксида с SOD имеет первый порядок по концентрации супероксида. Кроме того, супероксиддисмутаза имеет наибольшую kКот/KM (приблизительная каталитическая эффективность) любого известного фермента (~ 7 x 109 M−1s−1),[22] эта реакция ограничивается только частотой столкновений между ним и супероксидом. То есть скорость реакции «ограничена диффузией».
Высокая эффективность супероксиддисмутазы кажется необходимой: даже при субнаномолярных концентрациях, достигаемых высокими концентрациями SOD в клетках, супероксид инактивирует фермент цикла лимонной кислоты. аконитаза, может отравить энергетический обмен и высвобождать потенциально токсичное железо. Аконитаза - одна из нескольких железосеросодержащих (де) гидратаз в метаболических путях, которые, как было показано, инактивируются супероксидом.[23]
Механизм стабилизации и складывания
SOD1 - чрезвычайно стабильный белок. В голо-форме (связанная как с медью, так и с цинком) температура плавления составляет> 90 ° C. В апо-форме (не связанной с медью или цинком) температура плавления составляет ~ 60 ° C.[24] К дифференциальная сканирующая калориметрия (DSC), голо SOD1 разворачивается по механизму двух состояний: от димера к двум развернутым мономерам.[24] В химической денатурация В экспериментах holo SOD1 разворачивается по механизму трех состояний с наблюдением свернутого мономерного интермедиата.[25]
Физиология
Супероксид - один из основных активные формы кислорода в камере. Как следствие, СОД выполняет ключевую антиоксидантную роль. Физиологическое значение SOD иллюстрируется серьезными патологиями, очевидными у мышей, генетически модифицированных без этих ферментов. Мыши, лишенные SOD2, умирают через несколько дней после рождения на фоне массового окислительный стресс.[26] У мышей, лишенных SOD1, развивается широкий спектр патологий, включая гепатоцеллюлярную карциному,[27] ускорение возрастной потери мышечной массы,[28] более ранняя заболеваемость катарактой и сокращение продолжительности жизни. Мыши, лишенные SOD3, не обнаруживают каких-либо явных дефектов и демонстрируют нормальную продолжительность жизни, хотя они более чувствительны к гипероксическим повреждениям.[29] Нокаут-мыши любого фермента СОД более чувствительны к летальному исходу от соединений, вырабатывающих супероксид, таких как паракват и дикват (гербициды ).
Дрозофила Без SOD1 продолжительность жизни резко сокращается, тогда как мухи без SOD2 умирают до рождения. Истощение SOD1 и SOD2 в нервной системе и мышцах Дрозофила связано с сокращением продолжительности жизни.[30] Накопление нервных и мышечных ROS по-видимому, способствует возрастным нарушениям. Когда индуцируется сверхэкспрессия митохондриальной SOD2, продолжительность жизни взрослых Дрозофила расширяется.[31]
Среди черные садовые муравьи (Ласиус нигер), продолжительность жизни королевы на порядок больше, чем у рабочих, несмотря на отсутствие систематических различий в нуклеотидных последовательностях между ними.[32] В SOD3 Было обнаружено, что ген наиболее дифференциально сверхэкспрессируется в мозге королевы и рабочих муравьев. Это открытие указывает на возможность важной роли антиоксидантной функции в модулировании продолжительности жизни.[32]
Нокдауны SOD в черве C. elegans не вызывают серьезных физиологических нарушений. Однако продолжительность жизни C. elegans может быть расширен супероксидом /каталаза миметики, предполагающие, что окислительный стресс является основным фактором, определяющим скорость старение.[33]
Нокаут или нулевые мутации в SOD1 очень вредны для аэробного роста почкующихся дрожжей Saccharomyces cerevisiae и привести к резкому сокращению продолжительности жизни в постдиуксийском периоде. У дикого типа С. cerevisiae, Повреждение ДНК частота увеличивалась в 3 раза с возрастом, но более чем в 5 раз у мутантов, удаленных либо по SOD1 или же SOD2 гены.[34] Активные формы кислорода уровни повышаются с возрастом у этих мутантных штаммов и демонстрируют модель, аналогичную структуре Повреждение ДНК увеличиваются с возрастом. Таким образом, оказывается, что супероксиддисмутаза играет существенную роль в сохранении целостности генома во время старение в С. cerevisiaeНокаут или нулевые мутации .SOD2 вызывают ингибирование роста респираторных источников углерода в дополнение к сокращению продолжительности жизни в постдиуксийном периоде.
В делящихся дрожжах Schizosaccharomyces pombe, дефицит митохондриальной супероксиддисмутазы SOD2 ускоряет хронологическое старение.[35]
Было создано несколько прокариотических нулевых мутантов SOD, включая Кишечная палочка. Утрата периплазматического CuZnSOD вызывает потерю вирулентности и может быть привлекательной мишенью для новых антибиотиков.
Роль в болезни
Мутации в первом ферменте СОД (SOD1 ) может вызвать семейную боковой амиотрофический склероз (ALS, форма заболевание двигательного нейрона ).[36][37][38][39] Самая распространенная мутация в США - это A4V, а наиболее изученным является G93A. Две другие изоформы SOD не были связаны со многими заболеваниями человека, однако у мышей инактивация SOD2 вызывает перинатальную летальность.[26] и инактивация SOD1 вызывает гепатоцеллюлярная карцинома.[27] Мутации в SOD1 может вызывать семейный БАС (несколько доказательств также показывают, что SOD1 дикого типа в условиях клеточного стресса участвует в значительной части спорадических случаев БАС, которые составляют 90% пациентов с БАС),[40] по механизму, который в настоящее время не изучен, но не из-за потери ферментативной активности или снижения конформационной стабильности белка SOD1. Избыточная экспрессия SOD1 связана с нервными расстройствами, наблюдаемыми при Синдром Дауна.[41] У пациентов с талассемией СОД будет увеличиваться как форма механизма компенсации. Однако на хронической стадии СОД кажется недостаточным и имеет тенденцию к снижению из-за разрушения белков в результате массивной реакции окислитель-антиоксидант.[42]
У мышей внеклеточная супероксиддисмутаза (SOD3, ecSOD) способствует развитию гипертонии.[43][44] Снижение активности SOD3 было связано с заболеваниями легких, такими как острый респираторный дистресс-синдром (ARDS) или хроническая обструктивная болезнь легких (ХОБЛ).[45][46][47]
Супероксиддисмутаза также не экспрессируется в клетках нервного гребня в развивающихся плод. Следовательно, высокий уровень свободных радикалов может вызвать их повреждение и вызвать дисрафические аномалии (дефекты нервной трубки).[нужна цитата ]
Фармакологическая активность
СОД обладает мощным противовоспалительным действием. Например, СОД - это высокоэффективное экспериментальное лечение хронического воспаления у колит.[нужна цитата ] Лечение СОД уменьшается активные формы кислорода поколение и окислительный стресс и, таким образом, подавляет активацию эндотелия. Следовательно, такие антиоксиданты могут быть важными новыми препаратами для лечения воспалительное заболевание кишечника.[48]
Точно так же СОД обладает множеством фармакологических активностей. Например, улучшается цис-платина -индуцированный нефротоксичность у грызунов.[49] Как "Орготеин" или "онцеин", фармакологически активная очищенная СОД бычьей печени, он также эффективен при лечении воспалительного заболевания мочевыводящих путей у человека.[50] Какое-то время СОД из бычьей печени даже имела одобрение регулирующих органов в нескольких европейских странах для такого использования. Это было прервано опасениями по поводу прионная болезнь.[нужна цитата ]
An СОД-миметик агент TEMPOL, в настоящее время проходит клинические испытания радиозащиты и предотвращения радиационно-индуцированного дерматит.[51] TEMPOL и аналогичные SOD-миметики нитроксилов обладают множеством действий при заболеваниях, связанных с окислительным стрессом.[52]
Косметическое использование
СОД может уменьшить повреждение кожи свободными радикалами - например, уменьшить фиброз после облучения при раке груди. Однако исследования такого рода следует рассматривать как предварительные, поскольку в исследовании не было адекватного контроля, включая отсутствие рандомизации, двойного ослепления или плацебо.[53] Известно, что супероксиддисмутаза обращает фиброз, возможно, через де-дифференциация из миофибробласты вернуться к фибробласты.[54][требуется дальнейшее объяснение ]
Коммерческие источники
СОД коммерчески получают из морских фитопланктон, бычья печень, хрен, мускусная дыня, и некоторые бактерии. В терапевтических целях СОД обычно вводят местно. Нет никаких доказательств того, что употребление незащищенных SOD или продуктов, богатых SOD, может иметь какие-либо физиологические эффекты, поскольку вся проглоченная SOD является сломан в аминокислоты перед поглощается. Однако прием СОД, связанной с белками пшеницы, может улучшить ее терапевтическую активность, по крайней мере, теоретически.[55]
Смотрите также
- Каталаза
- Глутатион пероксидаза
- Джиаогулан
- НАДФН оксидаза, фермент, который производит супероксид
- Пероксидаза
Рекомендации
- ^ а б PDB: 1VAR; Borgstahl GE, Parge HE, Hickey MJ, Johnson MJ, Boissinot M, Hallewell RA, Lepock JR, Cabelli DE, Tainer JA (апрель 1996 г.). «Полиморфный вариант митохондриальной супероксиддисмутазы марганца человека Ile58Thr снижает активность, дестабилизируя тетрамерный интерфейс». Биохимия. 35 (14): 4287–97. Дои:10.1021 / bi951892w. PMID 8605177. S2CID 7450190.
- ^ Хайян М., Хашим М.А., Аль-Нашеф И.М. (2016). «Ион супероксида: образование и химические последствия». Chem. Rev. 116 (5): 3029–3085. Дои:10.1021 / acs.chemrev.5b00407. PMID 26875845.
- ^ МакКорд Дж. М., Фридович И. (ноябрь 1969 г.). «Супероксиддисмутаза. Ферментативная функция эритрокупреина (гена)». Журнал биологической химии. 244 (22): 6049–55. PMID 5389100.
- ^ МакКорд Дж. М., Фридович И. (1988). «Супероксиддисмутаза: первые двадцать лет (1968-1988)». Свободная радикальная биология и медицина. 5 (5–6): 363–9. Дои:10.1016/0891-5849(88)90109-8. PMID 2855736.
- ^ Брюэр Дж. Дж. (Сентябрь 1967 г.). «Ахроматические области крахмальных гелей, окрашенных тетразолием: унаследованные электрофоретические вариации». Американский журнал генетики человека. 19 (5): 674–80. ЧВК 1706241. PMID 4292999.
- ^ PDB: 2SOD;Тайнер Ж.А., Getzoff ED, Бим К.М., Ричардсон Дж. С., Ричардсон, округ Колумбия (сентябрь 1982 г.). «Определение и анализ 2 A-структуры меди, супероксиддисмутазы цинка». J. Mol. Биол. 160 (2): 181–217. Дои:10.1016/0022-2836(82)90174-7. PMID 7175933.
- ^ Квинт П., Реутцель Р., Микульски Р., МакКенна Р., Сильверман Д. Н. (февраль 2006 г.). «Кристаллическая структура нитрированной супероксиддисмутазы марганца человека: механизм инактивации». Свободная радикальная биология и медицина. 40 (3): 453–8. Дои:10.1016 / j.freeradbiomed.2005.08.045. PMID 16443160.
- ^ Ричардсон Дж., Томас К.А., Рубин Б.Х., Ричардсон, округ Колумбия (апрель 1975 г.). «Кристаллическая структура бычьей Cu, Zn супероксиддисмутазы при разрешении 3 A: отслеживание цепи и металлические лиганды». Труды Национальной академии наук Соединенных Штатов Америки. 72 (4): 1349–53. Дои:10.1073 / pnas.72.4.1349. ЧВК 432531. PMID 1055410..
- ^ Тайнер Дж. А., Гетцофф Э. Д., Ричардсон Дж. С., Ричардсон округ Колумбия (1983). «Строение и механизм действия медь, супероксиддисмутазы цинка». Природа. 306 (5940): 284–7. Bibcode:1983Натура 306..284Т. Дои:10.1038 / 306284a0. PMID 6316150. S2CID 4266810.
- ^ а б c PDB: 1N0J; Borgstahl GE, Parge HE, Hickey MJ, Beyer WF, Hallewell RA, Tainer JA (октябрь 1992 г.). «Структура митохондриальной супероксиддисмутазы марганца человека обнаруживает новый тетрамерный интерфейс двух 4-спиральных пучков». Клетка. 71 (1): 107–18. Дои:10.1016 / 0092-8674 (92) 90270-М. PMID 1394426. S2CID 41611695.
- ^ Barondeau DP, Kassmann CJ, Bruns CK, Tainer JA, Getzoff ED (июнь 2004 г.). «Структура и механизм супероксиддисмутазы никеля». Биохимия. 43 (25): 8038–47. Дои:10.1021 / bi0496081. PMID 15209499. S2CID 10700340.
- ^ а б PDB: 1 квартал; Вургес Дж., Ли Дж. У., Йим Й., Йим Х. С., Кан СО, Джинович Каруго К. (июнь 2004 г.). «Кристаллическая структура никельсодержащей супероксиддисмутазы выявляет другой тип активного центра». Труды Национальной академии наук Соединенных Штатов Америки. 101 (23): 8569–74. Bibcode:2004PNAS..101.8569W. Дои:10.1073 / pnas.0308514101. ЧВК 423235. PMID 15173586.
- ^ PDB: 1SDY; Джинович К., Гатти Дж., Кода А., Антолини Л., Пелоси Дж., Дезидери А., Фалькони М., Мармоччи Ф, Ролилио Дж., Болоньези М. (декабрь 1991 г.). «Уточнение структуры раствора и молекулярной динамики дрожжевого фермента супероксиддисмутазы Cu, Zn». Acta Crystallogr. B. 47 (6): 918–27. Дои:10.1107 / S0108768191004949. PMID 1772629.
- ^ Corpas FJ, Barroso JB, дель Рио, Лос-Анджелес (апрель 2001 г.). «Пероксисомы как источник активных форм кислорода и сигнальных молекул оксида азота в клетках растений». Тенденции в растениеводстве. 6 (4): 145–50. Дои:10.1016 / S1360-1385 (01) 01898-2. PMID 11286918.
- ^ Corpas FJ, Fernández-Ocaña A, Carreras A, Valderrama R, Luque F, Esteban FJ, Rodríguez-Serrano M, Chaki M, Pedrajas JR, Sandalio LM, del Río LA, Barroso JB (июль 2006 г.). «Экспрессия различных форм супероксиддисмутазы зависит от типа клеток в листьях оливы (Olea europaea L.)». Физиология растений и клеток. 47 (7): 984–94. Дои:10.1093 / pcp / pcj071. PMID 16766574.
- ^ PDB: 3CQQ; Цао X, Антонюк С.В., Ситараман С.В., Уитсон Л.Дж., Тейлор А.Б., Холлоуэй С.П., Стрэндж Р.В., Дусетт П.А., Валентайн Д.С., Тивари А., Хейворд Л.Дж., Падуя С., Колберг Д.А., Хаснаин С.С., Харт П.Дж. (июнь 2008 г.) «Структуры варианта G85R SOD1 при семейном боковом амиотрофическом склерозе». J. Biol. Chem. 283 (23): 16169–77. Дои:10.1074 / jbc.M801522200. ЧВК 2414278. PMID 18378676.
- ^ PDB: 2JLP; Антонюк С.В., Стрэндж РВ, Марклунд С.Л., Хаснаин С.С. (май 2009 г.). «Структура внеклеточной супероксиддисмутазы меди-цинка человека при разрешении 1,7 A: понимание связывания гепарина и коллагена». J. Mol. Биол. 388 (2): 310–26. Дои:10.1016 / j.jmb.2009.03.026. PMID 19289127.
- ^ а б Альшер Р.Г., Эртурк Н., Хит Л.С. (май 2002 г.). «Роль супероксиддисмутаз (СОД) в контроле окислительного стресса у растений». Журнал экспериментальной ботаники. 53 (372): 1331–41. Дои:10.1093 / jexbot / 53.372.1331. PMID 11997379.
- ^ а б c Смирнов, Николай (1993). "Обзор Тэнсли № 52 Роль активного кислорода в реакции растений на дефицит воды и высыхание". Новый Фитолог. 125: 27–58. Дои:10.1111 / j.1469-8137.1993.tb03863.x.
- ^ а б Райчаудхури СС, Дэн XW (2008). «Роль супероксиддисмутазы в борьбе с окислительным стрессом у высших растений». Ботанический обзор. 66 (1): 89–98. Дои:10.1007 / BF02857783. S2CID 7663001.
- ^ Vanaporn M, Wand M, Michell SL, Sarkar-Tyson M, Ireland P, Goldman S, Kewcharoenwong C, Rinchai D, Lertmemongkolchai G, Titball RW (август 2011 г.). «Супероксиддисмутаза С необходима для внутриклеточного выживания и вирулентности Burkholderia pseudomallei». Микробиология. 157 (Pt 8): 2392–400. Дои:10.1099 / мик ..0.050823-0. PMID 21659326.
- ^ Генрих П.К., Лёффлер Г., Petrifies PE (2006). Biochemie und Pathobiochemie (Springer-Lehrbuch) (Нем. Ред.). Берлин: Springer. п. 123. ISBN 978-3-540-32680-9.
- ^ Гарднер П.Р., Райнери И., Эпштейн Л.Б., Белый CW (июнь 1995 г.). «Супероксидный радикал и железо модулируют активность аконитазы в клетках млекопитающих». Журнал биологической химии. 270 (22): 13399–405. Дои:10.1074 / jbc.270.22.13399. PMID 7768942.
- ^ а б Статопулос П.Б., Рамфельдт Дж. А., Карбасси Ф., Сиддалл, Калифорния, Лепок Дж. Р., Мейринг Е. М. (март 2006 г.). «Калориметрический анализ термодинамической стабильности и агрегации для мутантов супероксиддисмутазы, ассоциированных с апо и голоамиотрофическим боковым склерозом, Gly-93». Журнал биологической химии. 281 (10): 6184–93. Дои:10.1074 / jbc.M509496200. PMID 16407238.
- ^ Рамфельдт Дж. А., Статопулос П. Б., Чакрабаррти А., Лепок Дж. Р., Мейринг Е. М. (январь 2006 г.). «Механизм и термодинамика индуцированной гуанидиния хлорид денатурации ALS-ассоциированных мутантных Cu, Zn супероксиддисмутаз». Журнал молекулярной биологии. 355 (1): 106–23. Дои:10.1016 / j.jmb.2005.10.042. PMID 16307756.
- ^ а б Ли Ю., Хуанг Т.Т., Карлсон Э.Дж., Мелов С., Урселл П.С., Олсон Д.Л., Благородный Л.Дж., Йошимура М.П., Бергер С., Чан П.Х., Уоллес, округ Колумбия, Эпштейн СиДжей (декабрь 1995 г.). «Дилатационная кардиомиопатия и неонатальная летальность у мутантных мышей, лишенных супероксиддисмутазы марганца». Природа Генетика. 11 (4): 376–81. Дои:10.1038 / ng1295-376. PMID 7493016. S2CID 10900822.
- ^ а б Эльчури С., Оберли Т.Д., Ци В., Эйзенштейн Р.С., Джексон Робертс Л., Ван Реммен Х., Эпштейн С.Дж., Хуанг Т.Т. (январь 2005 г.). «Дефицит CuZnSOD приводит к стойкому и широко распространенному окислительному повреждению и гепатоканцерогенезу в более позднем возрасте». Онкоген. 24 (3): 367–80. Дои:10.1038 / sj.onc.1208207. PMID 15531919.
- ^ Мюллер Флорида, Сонг В., Лю Й., Чаудхури А., Пике-Даль С., Стронг Р., Хуанг Т.Т., Эпштейн С.Дж., Робертс Л.Дж., Чете М., Фолкнер Д.А., Ван Реммен Х. (июнь 2006 г.). «Отсутствие супероксиддисмутазы CuZn приводит к повышенному окислительному стрессу и ускорению возрастной атрофии скелетных мышц». Свободная радикальная биология и медицина. 40 (11): 1993–2004. Дои:10.1016 / j.freeradbiomed.2006.01.036. PMID 16716900.
- ^ Sentman ML, Granström M, Jakobson H, Reaume A, Basu S, Marklund SL (март 2006 г.). «Фенотипы мышей, лишенных внеклеточной супероксиддисмутазы и медь- и цинксодержащей супероксиддисмутазы». Журнал биологической химии. 281 (11): 6904–9. Дои:10.1074 / jbc.M510764200. PMID 16377630.
- ^ Ока С., Хираи Дж, Ясукава Т., Накахара И., Иноуэ Ю.Х. (август 2015 г.). «Корреляция накопления активных форм кислорода за счет истощения супероксиддисмутазы с возрастными нарушениями в нервной системе и мышцах взрослых дрозофил». Биогеронтология. 16 (4): 485–501. Дои:10.1007 / s10522-015-9570-3. PMID 25801590. S2CID 18050827.
- ^ Sun J, Folk D, Брэдли TJ, Tower J (июнь 2002 г.). «Вызванная сверхэкспрессия митохондриальной Mn-супероксиддисмутазы увеличивает продолжительность жизни взрослых особей Drosophila melanogaster». Генетика. 161 (2): 661–72. ЧВК 1462135. PMID 12072463.
- ^ а б Лукас Э.Р., Келлер Л. (июль 2018 г.). «Повышенная экспрессия генов старения и иммунитета у королев черного садового муравья». Экспериментальная геронтология. 108: 92–98. Дои:10.1016 / j.exger.2018.03.020. PMID 29625209. S2CID 5045743.
- ^ Мелов С., Рэйвенскрофт Дж., Малик С., Гилл М.С., Уокер Д.В., Клейтон П.И., Уоллес, округ Колумбия, Малфрой Б., Доктроу С.Р., Литгоу Дж. Дж. (Сентябрь 2000 г.). «Увеличение продолжительности жизни с помощью миметиков супероксиддисмутазы / каталазы». Наука. 289 (5484): 1567–9. Bibcode:2000Sci ... 289.1567M. Дои:10.1126 / science.289.5484.1567. PMID 10968795. S2CID 21519801.
- ^ Муид К.А., Каракая Хо, Коц А. (февраль 2014 г.). «Отсутствие активности супероксиддисмутазы вызывает фрагментацию ядерной ДНК в процессе старения» (PDF). Biochem. Биофиз. Res. Сообщество. 444 (2): 260–3. Дои:10.1016 / j.bbrc.2014.01.056. HDL:11147/5542. PMID 24462872.
- ^ Огата Т., Сено Т., Кавано С., Икеда С. (январь 2016 г.). «Дефицит митохондриальной супероксиддисмутазы ускоряет хронологическое старение у делящихся дрожжей Schizosaccharomyces pombe». Cell Biol. Int. 40 (1): 100–6. Дои:10.1002 / cbin.10556. PMID 26507459. S2CID 205563521.
- ^ Милани П., Гальярди С., Кова Е., Середа С. (2011). «Регуляция транскрипции и посттранскрипции SOD1 и ее потенциальные последствия при БАС». Neurology Research International. 2011: 1–9. Дои:10.1155/2011/458427. ЧВК 3096450. PMID 21603028.
- ^ Дэн Х.Х., Хентати А., Тайнер Дж. А., Икбал З., Каябиаб А., Хунг В.Й., Гетцофф Э.Д., Ху П., Херцфельдт Б., Роос Р.П. (август 1993 г.). «Боковой амиотрофический склероз и структурные дефекты в Cu, Zn супероксиддисмутазе». Наука. 261 (5124): 1047–51. Bibcode:1993Научный ... 261.1047D. Дои:10.1126 / science.8351519. PMID 8351519.
- ^ Conwit RA (декабрь 2006 г.). «Предотвращение семейного БАС: клиническое испытание возможно, но оправдано ли испытание эффективности?». Журнал неврологических наук. 251 (1–2): 1–2. Дои:10.1016 / j.jns.2006.07.009. PMID 17070848. S2CID 33105812.
- ^ Аль-Чалаби А., Ли П.Н. (август 2000 г.). «Последние достижения в области бокового амиотрофического склероза». Текущее мнение в неврологии. 13 (4): 397–405. Дои:10.1097/00019052-200008000-00006. PMID 10970056. S2CID 21577500.
- ^ Гальярди С., Кова Е., Давин А., Гуарески С., Абель К., Алвиси Е., Лафоренца Ю., Гидони Р., Кашман Дж. Р., Серони М., Середа С. (август 2010 г.). «Экспрессия мРНК SOD1 при спорадическом боковом амиотрофическом склерозе». Нейробиология болезней. 39 (2): 198–203. Дои:10.1016 / j.nbd.2010.04.008. PMID 20399857. S2CID 207065284.
- ^ Гронер Y, Элрой-Стейн О., Авраам К.Б., Шиклер М., Ноблер Х., Минк-Голомб Д., Бар-Пелед О., Яром Р., Ротшенкер С. (1994). «Повреждение клеток избытком CuZnSOD и синдром Дауна». Биомедицина и фармакотерапия. 48 (5–6): 231–40. Дои:10.1016/0753-3322(94)90138-4. PMID 7999984.
- ^ Рухито Л., Мулатсих С., Софро А.С. (май 2015 г.). «Статус супероксиддисмутазы при трансфузионной талассемии». Североамериканский журнал медицинских наук. 7 (5): 194–8. Дои:10.4103/1947-2714.157480. ЧВК 4462814. PMID 26110130.
- ^ Gongora MC, Qin Z, Laude K, Kim HW, McCann L, Folz JR, Dikalov S, Fukai T., Harrison DG (сентябрь 2006 г.). «Роль внеклеточной супероксиддисмутазы при гипертонии». Гипертония. 48 (3): 473–81. Дои:10.1161 / 01.HYP.0000235682.47673.ab. PMID 16864745.
- ^ Лоб Х.Е., Марвар П.Дж., Гузик Т.Дж., Шарма С., Макканн Л.А., Вейанд С., Гордон Ф.Дж., Харрисон Д.Г. (февраль 2010 г.). «Индукция гипертонии и периферического воспаления за счет снижения внеклеточной супероксиддисмутазы в центральной нервной системе». Гипертония. 55 (2): 277–83, 6 стр. После 283. Дои:10.1161 / ГИПЕРТЕНЗИЯAHA.109.142646. ЧВК 2813894. PMID 20008675.
- ^ Янг Р.П., Хопкинс Р., Блэк П.Н., Эдди К., Ву Л., Гэмбл Г.Д., Миллс Г.Д., Гарретт Дж. Э., Итон Т. Э., Рис М. И. (май 2006 г.). «Функциональные варианты антиоксидантных генов у курильщиков с ХОБЛ и с нормальной функцией легких». Грудная клетка. 61 (5): 394–9. Дои:10.1136 / thx.2005.048512. ЧВК 2111196. PMID 16467073.
- ^ Гангули К., Депнер М., Фаттман К., Бейн К., Ури Т.Д., Весселкампер С.К., Борхерс М.Т., Шрайбер М., Гао Ф., фон Мутиус Э., Кабеш М., Лейкауф Г.Д., Шульц Х. (май 2009 г.). «Супероксиддисмутаза 3, внеклеточные (SOD3) варианты и функция легких». Физиологическая геномика. 37 (3): 260–7. Дои:10.1152 / физиолгеномика.90363.2008. ЧВК 2685504. PMID 19318538.
- ^ Gongora MC, Lob HE, Landmesser U, Guzik TJ, Martin WD, Ozumi K, Wall SM, Wilson DS, Murthy N, Gravanis M, Fukai T., Harrison DG (октябрь 2008 г.). «Потеря внеклеточной супероксиддисмутазы приводит к острому повреждению легких в присутствии окружающего воздуха: потенциальный механизм, лежащий в основе респираторного дистресс-синдрома взрослых». Американский журнал патологии. 173 (4): 915–26. Дои:10.2353 / ajpath.2008.080119. ЧВК 2543061. PMID 18787098.
- ^ Seguí J, Gironella M, Sans M, Granell S, Gil F, Gimeno M, Coronel P, Piqué JM, Panés J (сентябрь 2004 г.). «Супероксиддисмутаза облегчает TNBS-индуцированный колит за счет снижения окислительного стресса, экспрессии молекул адгезии и рекрутирования лейкоцитов в воспаленный кишечник». Журнал биологии лейкоцитов. 76 (3): 537–44. Дои:10.1189 / jlb.0304196. PMID 15197232. S2CID 15028921.
- ^ Макгиннесс Дж. Э., Проктор PH, Демопулос Х. Б., Хокансон Дж. А., Киркпатрик Д. С. (1978). «Снижение цис-платиновой нефротоксичности с помощью орготеина (супероксиддисмутазы)». Физиологическая химия и физика. 10 (3): 267–77. PMID 733940.
- ^ Марбергер Х., Хубер В., Бартч Г., Шульте Т., Свобода П. (1974). «Орготеин: новый противовоспалительный металлопротеиновый препарат для оценки клинической эффективности и безопасности при воспалительных заболеваниях мочевыводящих путей». Международная урология и нефрология. 6 (2): 61–74. Дои:10.1007 / bf02081999. PMID 4615073. S2CID 23880216.
- ^ Номер клинического исследования NCT01324141 на "Актуальный MTS-01 при дерматите во время лучевой и химиотерапии рака анального канала" на ClinicalTrials.gov
- ^ Wilcox CS (май 2010 г.). «Эффекты темпол и окислительно-восстановительных нитроксидов в моделях окислительного стресса». Фармакология и терапия. 126 (2): 119–45. Дои:10.1016 / j.pharmthera.2010.01.003. ЧВК 2854323. PMID 20153367.
- ^ Кампана Ф, Зервудис С., Пердеро Б., Гез Э., Фурке А., Бадиу С., Цакирис Г., Кулалоглоу С. (2004). «Местная супероксиддисмутаза снижает фиброз рака молочной железы после облучения». Журнал клеточной и молекулярной медицины. 8 (1): 109–16. CiteSeerX 10.1.1.336.8033. Дои:10.1111 / j.1582-4934.2004.tb00265.x. ЧВК 6740277. PMID 15090266.
- ^ Возенин-Бротонс М.С., Сиван В., Голт Н., Ренар С., Геффротин С., Деланиан С., Лефаикс Дж. Л., Мартин М. (январь 2001 г.). «Антифибротическое действие Cu / Zn SOD опосредуется репрессией TGF-бета1 и фенотипической реверсией миофибробластов». Свободная радикальная биология и медицина. 30 (1): 30–42. Дои:10.1016 / S0891-5849 (00) 00431-7. PMID 11134893.
- ^ Romao S (март 2015 г.). «Терапевтическая ценность пероральных добавок с комбинацией супероксиддисмутазы дыни и пшеничного глиадина». Питание. 31 (3): 430–6. Дои:10.1016 / j.nut.2014.10.006. PMID 25701330.
внешняя ссылка
- Онлайн-менделевское наследование в человеке (OMIM): 105400 (ALS)
- Онлайн-база данных ALS
- Краткий, но содержательный обзор SOD и литературы по нему.
- Теории старения, основанные на повреждениях Включает обсуждение роли SOD1 и SOD2 в старении.
- Комм. Врачей. For Responsible Med.
- Изображение пути SOD и окислительного стресса
- Историческая справка по исследованию СОД "Развитие Свободная радикальная биология и медицина: 20-летняя история "и"Свободная радикальная биология и медицина Последние 20 лет: самые цитируемые статьи »
- Дж. М. МакКорд обсуждает открытие SOD
- PDBe-KB предоставляет обзор всей структурной информации, доступной в PDB для супероксиддисмутазы человека [Cu-Zn]
- PDBe-KB предоставляет обзор всей структурной информации, доступной в PDB для супероксиддисмутазы человека [Mn], митохондриальная
- PDBe-KB предоставляет обзор всей структурной информации, доступной в PDB для внеклеточной супероксиддисмутазы человека [Cu-Zn]