N-связанное гликозилирование - N-linked glycosylation

N-связанное гликозилирование, является прикреплением олигосахарид, углевод, состоящий из нескольких молекул сахара, иногда также называемый гликан, к атому азота ( амид азот аспарагин (Asn) остаток белок ) в процессе, называемом N-гликозилирование, учился в биохимия.[1] Этот тип связи важен как для структуры[2] и функция[3] некоторых эукариотических белков. В N-связанный гликозилирование процесс происходит в эукариоты и широко в археи, но очень редко в бактерии. Природа N-связанные гликаны, присоединенные к гликопротеину, определяются белком и клеткой, в которой он экспрессируется.[4] Он также варьируется в зависимости от разновидность. Разные виды синтезируют разные типы N-связанный гликан.
Энергетика образования связи
В гликопротеине участвуют два типа связей: связи между сахариды остатки в гликане и связь между цепью гликана и молекулой белка.
Сахар части связаны друг с другом в гликановой цепи через гликозидные связи. Эти связи обычно образуются между угли 1 и 4 молекулы сахара. Образование гликозидной связи энергетически невыгодно, поэтому реакция сопряжена с гидролиз из двух АТФ молекулы.[4]
С другой стороны, присоединение остатка гликана к белку требует распознавания консенсусная последовательность. N-связанные гликаны почти всегда присоединены к азот атом боковой цепи аспарагина (Asn), который присутствует в составе Asn – X–Сер /Thr консенсусная последовательность, где X - любая аминокислота, кроме пролин (Pro).[4]
В клетках животных гликан, присоединенный к аспарагину, почти неизбежно N-ацетилглюкозамин (GlcNAc) в β-конфигурации.[4] Эта β-связь аналогична гликозидной связи между сахарными фрагментами в структуре гликана, как описано выше. Вместо того, чтобы быть прикрепленным к сахару гидроксил группа, аномерный углерод атом азота присоединен к амиду. Энергия, необходимая для этой связи, исходит от гидролиз из пирофосфат молекула.[4]
Биосинтез

Биосинтез N-связанные гликаны происходит через 3 основных этапа:[4]
- Синтез олигосахарида-предшественника, связанного с долихолом
- Перенос предшественника олигосахарида в белок по блоку
- Обработка олигосахарида
Синтез, блочный перенос и начальная обрезка прекурсора олигосахарид происходит в эндоплазматический ретикулум (ER). Последующая обработка и модификация олигосахаридной цепи осуществляется в аппарат Гольджи.
Таким образом, синтез гликопротеинов пространственно разделен в разных клеточных компартментах. Следовательно, тип NСинтез -гликан зависит от его доступности для различных ферментов, присутствующих в этих клеточных компартментах.
Однако, несмотря на разнообразие, все N-гликаны синтезируются общим путем с общей структурой гликанов ядра.[4]Основная структура гликана состоит из двух N-ацетилглюкозамин и три манноза остатки. Затем этот сердцевинный гликан перерабатывается и модифицируется, что приводит к разнообразию N-гликановые структуры.[4]
Синтез предшественника олигосахарида
Процесс N-связанное гликозилирование начинается с образования долихол -связанный сахар GlcNAc. Долихол - это липид молекула состоит из повторяющихся изопрен единицы. Эта молекула прикреплена к мембране ER. Молекулы сахара присоединены к долихолу через пирофосфатную связь.[4] (один фосфат изначально был связан с долихолом, а второй фосфат происходил из нуклеотид сахар). Затем цепь олигосахарида удлиняется путем добавления различных молекул сахара ступенчатым образом с образованием олигосахарида-предшественника.
Сборка этого олигосахарида-предшественника происходит в две фазы: фаза I и фаза II.[4] Фаза I проходит на цитоплазматический сторона ER и Фаза II проходит на просвет сторона ER.
Молекула-предшественник, готовая к переносу в белок, состоит из 2 GlcNAc, 9 маннозы и 3 глюкоза молекулы.

| |
| Фаза II | |
является донором остатка маннозы (образование: Dol-P + GDP-Man → Dol-P-Man + GDP), а Dol-P-Gluc является донором остатка глюкозы (образование: Dol-P + UDP-Glc → Dol-P- Glc + UDP).
| |
Передача гликана в белок
Как только олигосахарид-предшественник образуется, готовый гликан переносится в формирующийся полипептид в просвете мембраны ER. Эта реакция вызывается энергией, высвобождающейся при разрыве пирофосфатной связи между молекулой долихол-гликан. Перед переносом гликана на формирующийся полипептид необходимо выполнить три условия:[4]
- Аспарагин должен находиться в определенной согласованной последовательности в первичная структура (Asn – X – Ser или Asn – X – Thr или в редких случаях Asn – X – Cys).[5]
- Аспарагин должен соответствующим образом располагаться в трехмерной структуре белка (сахара полярные молекулы и, следовательно, должен быть прикреплен к аспарагину, расположенному на поверхности белка, а не похороненному внутри белка)
- Аспарагин должен находиться на просветной стороне эндоплазматического ретикулума. N-связанное гликозилирование должно быть инициировано. Целевые остатки находятся либо в секреторных белках, либо в областях трансмембранный белок который обращен к просвету.
Олигосахарилтрансфераза представляет собой фермент, ответственный за распознавание консенсусной последовательности и перенос гликана-предшественника на акцептор полипептида, который транслируется в просвете эндоплазматического ретикулума. N-связанное гликозилирование, следовательно, является ко-трансляционным событием
Обработка гликана

N-гликановая обработка осуществляется в эндоплазматическом ретикулуме и теле Гольджи. Первоначальная обрезка молекулы-предшественника происходит в ER, а последующий процессинг происходит в Golgi.
После переноса завершенного гликана на формирующийся полипептид из структуры удаляются два остатка глюкозы. Ферменты, известные как гликозидазы, удаляют некоторые остатки сахара. Эти ферменты могут разрушать гликозидные связи с помощью молекулы воды. Эти ферменты являются экзогликозидазами, поскольку действуют только на моносахарид остатки, расположенные на невосстанавливающем конце гликана.[4] Предполагается, что этот начальный этап обрезки действует как этап контроля качества в ER для мониторинга сворачивание белка.
Как только белок сложен правильно, два остатка глюкозы удаляются глюкозидаза I и II. Удаление последнего третьего остатка глюкозы сигнализирует о том, что гликопротеин готов к переходу из ER в СНГ-Гольджи.[4] . ER маннозидаза катализирует удаление этой конечной глюкозы. Однако, если белок не свернут должным образом, остатки глюкозы не удаляются, и, следовательно, гликопротеин не может покинуть эндоплазматический ретикулум. А сопровождающий белок (калнексин /кальретикулин ) связывается с развернутым или частично свернутым белком, чтобы способствовать сворачиванию белка.
Следующий шаг включает дальнейшее добавление и удаление остатков сахара в цис-Гольджи. Эти модификации катализируются гликозилтрансферазами и гликозидазами соответственно. в СНГ-Гольджи, серия маннозидаз удаляет некоторые или все четыре остатка маннозы в α-1,2 связях.[4] В то время как в медиальной части Гольджи гликозилтрансферазы добавляют остатки сахара к структуре гликанов ядра, давая начало трем основным типам гликанов: высокоманнозным, гибридным и сложным гликанам.
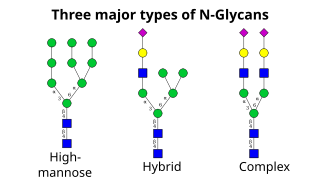
- Высокоманнозы, по сути, всего два N-ацетилглюкозамины со многими остатками маннозы, часто почти такими же, как в олигосахаридах-предшественниках до того, как они присоединятся к белку.
- Сложные олигосахариды названы так потому, что они могут содержать почти любое количество других типов сахаридов, включая более двух исходных N-ацетилглюкозамины.
- Гибридные олигосахариды содержат остатки маннозы на одной стороне ветви, а на другой стороне - остатки маннозы. N-ацетилглюкозамин инициирует сложную ветвь.
Порядок добавления сахаров к растущим гликановым цепям определяется субстратной специфичностью ферментов и их доступом к субстрату по мере их прохождения. секреторный путь. Таким образом, организация этого механизма внутри клетки играет важную роль в определении того, какие гликаны производятся.
Ферменты в Гольджи
Ферменты Гольджи играют ключевую роль в определении синтеза различных типов гликанов. Порядок действия ферментов отражается в их положении в стеке Гольджи:
| Ферменты | Расположение в Гольджи |
|---|---|
| Маннозидаза I | СНГ-Гольджи |
| GlcNAc трансферазы | медиальный Гольджи |
| Галактозилтрансфераза и Сиалилтрансфераза | транс-Гольджи |
У архей и прокариот
Похожий NПути биосинтеза -гликанов были обнаружены у прокариот и архей.[6] Однако по сравнению с эукариотами конечная структура гликанов у эубактерий и архей, по-видимому, не сильно отличается от исходного предшественника, образующегося в эндоплазматическом ретикулуме. У эукариот исходный олигосахарид-предшественник значительно модифицируется на пути к поверхности клетки.[4]
Функция
N-связанные гликаны имеют внутренние и внешние функции.[4]
В иммунной системе N-связанные гликаны на поверхности иммунной клетки помогут диктовать эту модель миграции клетки, например иммунные клетки, которые мигрируют в кожу, имеют специфические гликозилирования, которые способствуют перемещению в этот участок.[7] Паттерны гликозилирования различных иммуноглобулинов, включая IgE, IgM, IgD, IgA и IgG, наделяют их уникальными эффекторными функциями, изменяя их сродство к Fc и другим иммунным рецепторам.[7] Гликаны также могут быть вовлечены в «само» и «несамо» дискриминацию, что может иметь отношение к патофизиологии различных аутоиммунных заболеваний.[7]
| Внутренний |
|
| Внешний |
|
Клиническое значение
Изменения в N-связанное гликозилирование было связано с различными заболеваниями, включая ревматоидный артрит,[9] диабет 1 типа,[10] болезнь Крона,[11] и раковые заболевания.[12][13]
Мутации в восемнадцати генах, участвующих в N-связанное гликозилирование приводит к множеству заболеваний, большинство из которых связано с нервная система.[3][13]
Важность терапевтических белков
Много терапевтический белки на рынке антитела, которые N-связанные гликопротеины. Например, Этанерцепт, Инфликсимаб и Ритуксимаб находятся N-гликозилированные терапевтические белки.

Важность N-связанное гликозилирование становится все более очевидным в области фармацевтические препараты.[14] Хотя бактериальный или дрожжевой производство белка системы обладают значительными потенциальными преимуществами, такими как высокий выход и низкая стоимость, проблемы возникают, когда интересующий белок представляет собой гликопротеин. Большинство прокариотических систем экспрессии, таких как Кишечная палочка не может выполнять посттрансляционные модификации. С другой стороны, эукариотические экспрессирующие хозяева, такие как клетки дрожжей и животных, имеют разные паттерны гликозилирования. Белки, продуцируемые в этих экспрессионных хозяевах, часто не идентичны человеческим белкам и, следовательно, вызывают иммуногенный реакции у пациентов. Например, S.cerevisiae (дрожжи) часто производят гликаны с высоким содержанием маннозы, которые являются иммуногенными.
Системы экспрессии млекопитающих, не относящиеся к человеку, такие как CHO или же Ячейки NS0 иметь оборудование, необходимое для добавления сложных гликанов человеческого типа. Однако гликаны, продуцируемые в этих системах, могут отличаться от гликанов, продуцируемых в организме человека, поскольку они могут быть блокированы обоими N-гликолилнейраминовая кислота (Neu5Gc) и N-ацетилнейраминовая кислота (Neu5Ac), тогда как клетки человека производят только гликопротеины, содержащие N-ацетилнейраминовая кислота. Кроме того, клетки животных могут также продуцировать гликопротеины, содержащие галактоза-альфа-1,3-галактоза эпитоп, который может вызывать серьезные аллергические реакции, в том числе анафилактический шок, у людей, у которых Альфа-гал аллергия.
Эти недостатки были устранены с помощью нескольких подходов, таких как устранение путей, которые производят эти гликановые структуры, путем генетического нокаута. Кроме того, с помощью генной инженерии были созданы другие системы экспрессии для получения терапевтических гликопротеинов с человеческими N-связанные гликаны. К ним относятся дрожжи, такие как Pichia pastoris,[15] линии клеток насекомых, зеленые растения,[16] и даже бактерии.
Смотрите также
Рекомендации
- ^ «Гликозилирование». UniProt: последовательность белков и функциональная информация.
- ^ Imperiali B, O'Connor SE (декабрь 1999 г.). "Эффект N-связанное гликозилирование гликопептида и гликопротеиновой структуры ». Современное мнение в области химической биологии. 3 (6): 643–9. Дои:10.1016 / S1367-5931 (99) 00021-6. PMID 10600722.
- ^ а б Паттерсон М.С. (сентябрь 2005 г.). «Метаболические мимики: нарушения N-связанное гликозилирование ». Семинары по детской неврологии. 12 (3): 144–51. Дои:10.1016 / j.spen.2005.10.002. PMID 16584073.
- ^ а б c d е ж грамм час я j k л м п о п Дрикамер К., Тейлор М.Е. (2006). Введение в гликобиологию (2-е изд.). Oxford University Press, США. ISBN 978-0-19-928278-4.
- ^ Mellquist JL, Kasturi L, Spitalnik SL, Shakin-Eshleman SH (май 1998 г.). «Аминокислота, следующая за секвоном Asn – X – Ser / Thr, является важным фактором, определяющим Nэффективность гликозилирования связанного ядра ". Биохимия. 37 (19): 6833–7. Дои:10.1021 / bi972217k. PMID 9578569.
- ^ Делл А, Галадари А, Састре Ф, Хитчен П. (2010). «Сходства и различия в механизмах гликозилирования у прокариот и эукариот». Международный журнал микробиологии. 2010: 1–14. Дои:10.1155/2010/148178. ЧВК 3068309. PMID 21490701.
- ^ а б c Маверакис Э., Ким К., Шимода М., Гершвин М.Э., Патель Ф., Уилкен Р. и др. (Февраль 2015 г.). «Гликаны в иммунной системе и теория аутоиммунитета с измененными гликанами: критический обзор». Журнал аутоиммунитета. 57 (6): 1–13. Дои:10.1016 / j.jaut.2014.12.002. ЧВК 4340844. PMID 25578468.
- ^ Синклер AM, Эллиотт S (август 2005 г.). «Гликоинженерия: влияние гликозилирования на свойства терапевтических белков». Журнал фармацевтических наук. 94 (8): 1626–35. Дои:10.1002 / jps.20319. PMID 15959882.
- ^ Накагава Х., Хато М., Такегава Й., Дегучи К., Ито Х., Такахата М. и др. (Июнь 2007 г.). "Обнаружение измененных N-гликановые профили в цельной сыворотке пациентов с ревматоидным артритом ». Журнал хроматографии B. 853 (1–2): 133–7. Дои:10.1016 / j.jchromb.2007.03.003. HDL:2115/28276. PMID 17392038.
- ^ Бермингем М.Л., Коломбо М., МакГурнаган С.Дж., Блэкборн Л.А., Вучкович Ф., Пучич Бакович М. и др. (Январь 2018). "N-Гликановый профиль и заболевание почек при диабете 1 типа ». Уход за диабетом. 41 (1): 79–87. Дои:10.2337 / dc17-1042. PMID 29146600.
- ^ Трбоевич Акмачич I, Вентам Н.Т., Теодорату Э., Вучкович Ф., Кеннеди Н.А., Криштич Дж. И др. (Июнь 2015 г.). «Воспалительное заболевание кишечника связано с провоспалительным потенциалом иммуноглобулина G гликома». Воспалительные заболевания кишечника. 21 (6): 1237–47. Дои:10.1097 / MIB.0000000000000372. ЧВК 4450892. PMID 25895110.
- ^ Кодар К., Штадлманн Дж., Клаамас К., Сергеев Б., Куртенков О. (январь 2012 г.). "Иммуноглобулин G Fc N-Гликановое профилирование у пациентов с раком желудка с помощью LC-ESI-MS: связь с прогрессированием опухоли и выживаемостью ». Журнал гликоконъюгатов. 29 (1): 57–66. Дои:10.1007 / s10719-011-9364-z. PMID 22179780.
- ^ а б Чен Г, Ван И, Цинь Икс, Ли Х, Го И, Ван И и др. (Август 2013). "Изменение в IgG1 Fc N-связанное гликозилирование при раке легких человека: диагностический потенциал, связанный с возрастом и полом ». Электрофорез. 34 (16): 2407–16. Дои:10.1002 / elps.201200455. PMID 23766031.
- ^ Далзил М., Криспин М., Сканлан К.Н., Зицманн Н., Двек Р.А. (январь 2014 г.). «Новые принципы терапевтического использования гликозилирования». Наука. 343 (6166): 1235681. Дои:10.1126 / наука.1235681. PMID 24385630.
- ^ Гамильтон С.Р., Бобрович П., Бобрович Б., Дэвидсон Р.К., Ли Х., Митчелл Т. и др. (Август 2003 г.). «Производство сложных гликопротеинов человека в дрожжах». Наука. 301 (5637): 1244–6. Дои:10.1126 / science.1088166. PMID 12947202.
- ^ Штрассер Р., Альтманн Ф., Стейнкеллнер Х. (декабрь 2014 г.). «Контролируемое гликозилирование рекомбинантных белков растительного происхождения». Текущее мнение в области биотехнологии. 30: 95–100. Дои:10.1016 / j.copbio.2014.06.008. PMID 25000187.
внешняя ссылка
- ГликоЭП: Платформа In silico для прогнозирования N-, О- и C-Гликозиты в последовательностях эукариотических белков
- Маверакис Э., Ким К., Шимода М., Гершвин М.Э., Патель Ф., Уилкен Р., Райчаудхури С., Рухак Л. Р., Лебрилла CB (февраль 2015 г.). «Гликаны в иммунной системе и теория аутоиммунитета с измененными гликанами: критический обзор». Журнал аутоиммунитета. 57: 1–13. Дои:10.1016 / j.jaut.2014.12.002. ЧВК 4340844. PMID 25578468.