CYP3A4 - CYP3A4




Цитохром P450 3A4 (сокращенно CYP3A4) (ЕС 1.14.13.97 ) является важным фермент в организме, в основном в печени и кишечнике. Это окисляет небольшие инородные органические молекулы (ксенобиотики ), такие как токсины или лекарства, чтобы их можно было вывести из организма.
Хотя CYP3A4 дезактивирует многие препараты, есть также некоторые препараты, которые активирован ферментом. Некоторые вещества, такие как грейпфрут сок и некоторые лекарства препятствуют действию CYP3A4. Следовательно, эти вещества будут либо усиливать, либо ослаблять действие тех лекарств, которые модифицированы CYP3A4.
CYP3A4 является членом цитохром P450 семейство окислительных ферментов. Некоторые другие члены этого семейства также участвуют в метаболизме лекарств, но CYP3A4 является наиболее распространенным и универсальным. Как и все члены этой семьи, это гемопротеин, т.е. белок содержащий гем группа с атомом железа. У человека белок CYP3A4 кодируется CYP3A4 ген.[3] Этот ген является частью кластера цитохрома P450 гены на хромосома 7q22.1.[4]
Функция
CYP3A4 является членом цитохром P450 надсемейство из ферменты. Белки цитохрома P450 являются монооксигеназы которые катализируют многие реакции, участвующие в метаболизм лекарств и синтез холестерина, стероидов и других липидных компонентов.
Белок CYP3A4 локализуется в эндоплазматический ретикулум, а его выражение индуцируется глюкокортикоиды и некоторые фармакологические средства. Ферменты цитохрома P450 метаболизируют примерно 60% назначенных лекарств, при этом CYP3A4 отвечает примерно за половину этого метаболизма;[5] субстраты включают ацетаминофен, кодеин, циклоспорин (циклоспорин), диазепам и эритромицин. Фермент также метаболизирует некоторые стероиды и канцерогены.[6] Большинство лекарств деактивируются CYP3A4 либо напрямую, либо при содействии выделение от тела. Также многие вещества биоактивированный CYP3A4 с образованием своих активных соединений, а многие протоксины отравленный в их токсичные формы (примеры - см. таблицу ниже).
CYP3A4 также обладает эпоксигеназа активность в том, что он метаболизирует арахидоновая кислота к эпоксиэйкозатриеновые кислоты (EET), то есть (±) -8,9-, (±) -11,12- и (±) -14,15-эпоксиэйкозатриеновые кислоты.[7] EETs проводят широкий спектр мероприятий, включая популяризацию определенных типов рака (см. эпоксиэйкозатетраеновая кислота # рак ). CYP3A4 способствует росту различных типов линий раковых клеток человека в культуре, продуцируя (±) -14,15-эпоксиэйкозатриеновые кислоты, которые стимулируют рост этих клеток.[8] Также сообщается, что цитохром P450 обладает активностью монооксгеназы жирных кислот для метаболизма арахидоновой кислоты до 20-гидроксиэйкозатетраеновая кислота (20-НЕТЕ).[9] 20-HETE имеет широкий спектр действий, которые также включают стимуляцию роста груди и другие виды рака (см. 12-гидроксиэйкозатетраеновая кислота # рак ).
Эволюция
Ген CYP3A4 имеет гораздо более сложную регуляторную область выше по сравнению с его паралоги.[10] Эта повышенная сложность делает ген CYP3A4 более чувствительным к эндогенным и экзогенным лигандам PXR и CAR вместо того, чтобы полагаться на варианты гена для более широкой специфичности.[10] Шимпанзе и человеческий CYP3A4 высоко консервативны в метаболизме многих лиганды, хотя четыре аминокислоты, положительно выбранные у людей, привели к 5-кратному бензилирование из 7-BFC в присутствии гепатотоксичный вторичный желчная кислота литохолевая кислота.[11] Это изменение, как следствие, способствует усилению защиты человека от холестаз.[11]
Распределение тканей
Плоды склонны не экспрессировать CYP3A4 в тканях печени,[требуется разъяснение ] скорее CYP3A7 (ЕС 1.14.14.1 ), который действует на аналогичный набор подложек. CYP3A4 отсутствует в печени плода, но увеличивается примерно до 40% от взрослого уровня на четвертом месяце жизни и до 72% к 12 месяцам.[12][13]
Хотя CYP3A4 преимущественно находится в печени, он также присутствует в других органах и тканях тела, где может играть важную роль в метаболизме. CYP3A4 в кишечнике играет важную роль в метаболизме некоторых лекарств. Часто это позволяет пролекарства активироваться и впитываться - как в случае гистамин H1антагонист рецепторов терфенадин.
Недавно CYP3A4 также был идентифицирован в головном мозге, однако его роль в Центральная нервная система пока неизвестно.[14]
Механизмы
Цитохром P450 Ферменты осуществляют ряд модификаций различных лигандов, используя его большой активный центр и его способность связывать более одного субстрата за раз, чтобы выполнять сложные химические изменения в метаболизме эндогенных и экзогенных соединений. Они включают гидроксилирование, эпоксидирование олефинов, ароматических окисление, окисления гетероатомов, реакции N- и O-деалкилирования, окисления альдегидов, дегидрирование реакции и активность ароматазы.[15][16]
Гидроксилирование зр.3 Связь C-H является одним из способов воздействия CYP3A4 (и оксигеназы цитохрома P450) на свой лиганд.[17] Фактически, за гидроксилированием иногда следует дегидрирование, приводящее к более сложным метаболитам.[16] Пример молекулы, которая претерпевает более одной реакции из-за CYP3A4, включает: тамоксифен, который гидроксилируется до 4-гидрокси-тамоксифена, а затем дегидратируется до 4-гидрокситамоксифенхинонметида.[16] Было предложено два механизма в качестве основного пути гидроксилирования ферментов P450.
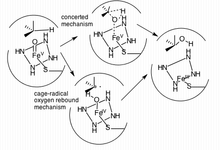
Первый предложенный путь - это радикальный метод, контролируемый клеткой («возврат кислорода»), а второй включает согласованный механизм, который не использует радикальный промежуточный продукт, а вместо этого действует очень быстро черезрадикальные часы ".[17]
Подавление при употреблении фруктов
В 1998 году различные исследователи показали, что грейпфрут сок и грейпфрут в целом являются мощным ингибитором CYP3A4, который может влиять на метаболизм различных препаратов, увеличивая их биодоступность.[18][19][20][21][22] В некоторых случаях это может привести к летальному исходу от таких лекарств, как астемизол или терфенадин.[19] Эффект грейпфрутового сока в отношении абсорбции лекарств был первоначально обнаружен в 1989 году. Первый опубликованный отчет о взаимодействии грейпфрута с лекарственными средствами был опубликован в 1991 году в Ланцет озаглавленный «Взаимодействие цитрусовых соков с фелодипином и нифедипином», и был первым клиническим описанием взаимодействия пищевых продуктов и лекарств. Эффект от грейпфрута длится 3–7 дней, с наибольшим эффектом, когда сок принимается за час до приема препарата.[23]
Помимо грейпфрута, другие фрукты обладают аналогичным действием. Нони (М. citrifolia), например, является пищевая добавка обычно употребляется в виде сока, а также подавляет CYP3A4;[24] гранат сок тоже имеет такой эффект.[25]
Изменчивость
Хотя старше 28 однонуклеотидный полиморфизм (SNP) были идентифицированы в CYP3A4 ген, было обнаружено, что это не приводит к значительной межиндивидуальной вариабельности in vivo. Можно предположить, что это может быть связано с индукцией CYP3A4 при воздействии на субстраты.
Аллели CYP3A4, которые, как сообщается, обладают минимальной функцией по сравнению с аллелями дикого типа, включают CYP3A4 * 6 (вставка A17776) и CYP3A4 * 17 (F189S). Оба этих SNP приводили к снижению каталитической активности с определенными лигандами, включая тестостерон и нифедипин по сравнению с метаболизмом дикого типа.[26]
Вариабельность функции CYP3A4 может быть определена неинвазивно с помощью дыхательный тест на эритромицин (ERMBT). Оценки ERMBT in vivo Активность CYP3A4 путем измерения радиоактивно меченного углекислого газа, выдыхаемого после внутривенного введения (14C-N-метил)-эритромицин.[27]
Индукция
CYP3A4 - это индуцированный широким спектром лиганды. Эти лиганды связываются с рецептор прегнана X (PXR). Активированный комплекс PXR образует гетеродимер с рецептор ретиноида X (RXR), который связывается с XREM регион CYP3A4 ген. XREM - это регулирующая область CYP3A4 ген, и связывание вызывает кооперативное взаимодействие с проксимальными областями промотора гена, что приводит к усилению транскрипции и экспрессии CYP3A4. Активация гетеродимера PXR / RXR инициирует транскрипция промоторной области и гена CYP3A4. Связывание лиганда увеличивается в присутствии лигандов CYP3A4, например, в присутствии афлатоксин B1, M1 и G1. Действительно, из-за большого и гибкого активного сайта фермента возможно связывание нескольких лигандов одновременно, что приводит к потенциально вредным побочным эффектам.[28]
Было показано, что индукция CYP3A4 варьируется у людей в зависимости от пола. Доказательства показывают увеличение очистка от наркотиков CYP3A4 у женщин, даже с учетом различий в массе тела. Исследование Wolbold et al. (2003) обнаружили, что медианные уровни CYP3A4, измеренные в образцах печени, удаленных хирургическим путем, у случайной выборки женщин превышали уровни CYP3A4 в печени мужчин на 129%. CYP3A4 мРНК транскрипты были обнаружены в аналогичных пропорциях, что позволяет предположить пре-трансляционный механизм активации CYP3A4 у женщин. Точная причина этого повышенного уровня фермента у женщин все еще обсуждается, однако исследования выяснили другие механизмы (такие как компенсация CYP3A5 или CYP3A7 пониженного уровня CYP3A4), которые влияют на клиренс лекарств как у мужчин, так и у женщин.[29]
Активация субстрата CYP3A4 варьируется у разных видов животных. Некоторые лиганды активируют PXR человека, который способствует транскрипции CYP3A4, но не проявляет активации у других видов. Например, PXR мыши не активируется рифампицин и человеческий PXR не активируется прегненалоном 16α-карбонитрилом[30] Для облегчения изучения функциональных путей CYP3A4 in vivo, линии мышей были разработаны с использованием трансгены для получения скрещиваний нулевого / человеческого CYP3A4 и PXR. Хотя гуманизированные мыши hCYP3A4 успешно экспрессировали фермент в кишечном тракте, низкие уровни hCYP3A4 были обнаружены в печени.[30] Этот эффект был приписан регуляции CYP3A4 гормон роста путь передачи сигнала.[30] Помимо предоставления in vivo Модель гуманизированных мышей CYP3A4 (hCYP3A4) использовались для дальнейшего подчеркивания гендерных различий в активности CYP3A4.[30]
Уровни активности CYP3A4 также связаны с диетой и факторами окружающей среды, такими как продолжительность воздействия ксенобиотических веществ.[31] Из-за обширного присутствия фермента в слизистой оболочке кишечника, фермент показал чувствительность к симптомам голодания и активируется для защиты от побочных эффектов. Действительно, у толстоголовых гольянов было показано, что голодные самки рыб имеют повышенную экспрессию PXR и CYP3A4 и проявляют более выраженную реакцию на ксенобиотические факторы после воздействия после нескольких дней голодания.[31] Изучая модели на животных и учитывая врожденные различия в активации CYP3A4, исследователи могут лучше предсказать метаболизм лекарств и побочные эффекты в путях CYP3A4 человека.
Оборот
Оценки оборот уровень человеческого CYP3A4 широко варьируется. Для печеночного CYP3A4, in vivo методы дают оценки фермента период полураспада в основном в диапазоне от 70 до 140 часов, тогда как in vitro методы дают оценки от 26 до 79 часов.[32] Обмен CYP3A4 кишечника, вероятно, будет функцией скорости энтероцит обновление; косвенный подход, основанный на восстановлении активности после воздействия грейпфрутового сока, дает измерения в диапазоне от 12 до 33 часов.[32]
Технологии
Из-за естественной склонности мембраносвязанного CYP3A4 к конгломерации исторически было трудно изучить связывание лекарств как в растворе, так и на поверхностях. Совместная кристаллизация затруднена, так как субстраты имеют тенденцию к низкой Kd (от 5 до 150 мкМ) и низкая растворимость в водных растворах.[33] Успешной стратегией выделения связанного фермента является функциональная стабилизация мономерного CYP3A4 на наночастицах серебра, полученных из наносферная литография и проанализированы с помощью локализованных поверхностный плазмонный резонанс спектроскопия (LSPR).[34] Эти анализы могут использоваться в качестве высокочувствительного анализа связывания лекарств и могут стать неотъемлемой частью дальнейших высокопроизводительных анализов, используемых при первоначальном тестировании открытия лекарств. В дополнение к LSPR, комплексы CYP3A4-Nanodisc были признаны полезными в других приложениях, включая твердотельный ЯМР, редокс-потенциометрия и стационарная кинетика ферментов.[34]
Лиганды
Ниже приводится таблица избранных субстраты, индукторы и ингибиторы из CYP3A4. Если классы агентов перечислены, внутри класса могут быть исключения.
Ингибиторы CYP3A4 можно классифицировать по их потенция, такие как:
- Сильный ингибитор тот, который вызывает как минимум 5-кратное увеличение плазмы Значения AUC, или снижение более чем на 80% оформление.[35]
- Умеренный ингибитор это тот, который вызывает по крайней мере 2-кратное увеличение значений AUC в плазме или снижение клиренса на 50-80%.[35]
- Слабый ингибитор это тот, который вызывает по крайней мере 1,25-кратное, но менее чем 2-кратное увеличение значений AUC в плазме или снижение клиренса на 20-50%.[35]
| Субстраты | Ингибиторы | Индукторы |
|---|---|---|
| Сильный
Умеренный
Слабый
Неустановленная потенция
| Сильная потенция Неустановленная потенция
|
Интерактивная карта проезда
Нажмите на гены, белки и метаболиты ниже, чтобы ссылки на соответствующие статьи. [§ 1]

- ^ Интерактивную карту путей можно редактировать на WikiPathways: "IrinotecanPathway_WP46359".
Смотрите также
использованная литература
- ^ а б c ГРЧ38: Ансамбль выпуск 89: ENSG00000160868 - Ансамбль, Май 2017
- ^ "Справочник человека по PubMed:". Национальный центр биотехнологической информации, Национальная медицинская библиотека США.
- ^ Хашимото Х., Тойде К., Китамура Р., Фудзита М., Тагава С., Ито С., Каматаки Т. (декабрь 1993 г.). «Генная структура CYP3A4, взрослой специфической формы цитохрома P450 в печени человека и его транскрипционный контроль». Европейский журнал биохимии / FEBS. 218 (2): 585–95. Дои:10.1111 / j.1432-1033.1993.tb18412.x. PMID 8269949.
- ^ Иноуэ К., Инадзава Дж., Накагава Х., Шимада Т., Ямадзаки Х., Генгерих Ф.П., Абэ Т. (июнь 1992 г.). «Отнесение гена нифедипиноксидазы человеческого цитохрома P-450 (CYP3A4) к хромосоме 7 в полосе q22.1 с помощью флуоресцентной гибридизации in situ». Японский журнал генетики человека. 37 (2): 133–8. Дои:10.1007 / BF01899734. PMID 1391968.
- ^ Zanger UM, Schwab M (апрель 2013 г.). «Ферменты цитохрома P450 в метаболизме лекарств: регуляция экспрессии генов, активности ферментов и влияние генетической изменчивости». Фармакология и терапия. 138 (1): 103–41. Дои:10.1016 / j.pharmthera.2012.12.007. PMID 23333322.
- ^ EntrezGene 1576
- ^ Бишоп-Бейли Д., Томсон С., Аскари А., Фолкнер А., Уиллер-Джонс С. (2014). «Липид-метаболизирующие CYP в регуляции и нарушении регуляции метаболизма». Ежегодный обзор питания. 34: 261–79. Дои:10.1146 / annurev-nutr-071813-105747. PMID 24819323.
- ^ Флеминг I (октябрь 2014 г.). «Фармакология оси цитохром Р450 эпоксигеназа / растворимая эпоксидгидролаза в сосудистой сети и сердечно-сосудистых заболеваниях». Фармакологические обзоры. 66 (4): 1106–40. Дои:10.1124 / пр.113.007781. PMID 25244930.
- ^ Мията Н., Танигучи К., Секи Т., Ишимото Т., Сато-Ватанабэ М., Ясуда Ю., Дои М., Каметани С., Томишима Ю., Уэки Т., Сато М., Камео К. (июнь 2001 г.). «HET0016, мощный и селективный ингибитор фермента, синтезирующего 20-HETE». Британский журнал фармакологии. 133 (3): 325–9. Дои:10.1038 / sj.bjp.0704101. ЧВК 1572803. PMID 11375247.
- ^ а б Qiu H, Mathäs M, Nestler S, Bengel C, Nem D, Gödtel-Armbrust U, Lang T, Taudien S, Burk O, Wojnowski L (март 2010 г.). «Уникальная сложность вышележащего региона CYP3A4 предполагает негенетическое объяснение вариабельности его экспрессии». Фармакогенетика и геномика. 20 (3): 167–78. Дои:10.1097 / FPC.0b013e328336bbeb. PMID 20147837. S2CID 205602787.
- ^ а б Кумар С., Цю Х., Озгуен Н., Херлин Х., Хальперт Дж. Р., Войновски Л. (июнь 2009 г.). «Разнообразие лигандов CYP3A4 человека и шимпанзе: активация человеческого CYP3A4 литохолевой кислотой в результате положительного отбора». Метаболизм и утилизация лекарств. 37 (6): 1328–33. Дои:10.1124 / dmd.108.024372. ЧВК 2683693. PMID 19299527.
- ^ Джонсон Т.Н., Ростами-Ходжеган А, Такер GT (2006). «Прогноз клиренса одиннадцати лекарств и связанной с ними вариабельности у новорожденных, младенцев и детей». Клиническая фармакокинетика. 45 (9): 931–56. Дои:10.2165/00003088-200645090-00005. PMID 16928154. S2CID 25596506.
- ^ Johnson TN, Tucker GT, Rostami-Hodjegan A (май 2008 г.). «Развитие CYP2D6 и CYP3A4 на первом году жизни». Клиническая фармакология и терапия. 83 (5): 670–1. Дои:10.1038 / sj.clpt.6100327. PMID 18043691. S2CID 9714442.
- ^ Робертсон Г.Р., Филд Дж., Гудвин Б., Бирач С., Тран М., Ленерт А., Лиддл С. (июль 2003 г.). «Трансгенные мышиные модели регуляции человеческого гена CYP3A4». Молекулярная фармакология. 64 (1): 42–50. Дои:10,1124 / моль. 64.1.42. PMID 12815159. S2CID 17209434.
- ^ Schmiedlin-Ren P, Edwards DJ, Fitzsimmons ME, He K, Lown KS, Woster PM, Rahman A, Thummel KE, Fisher JM, Hollenberg PF, Watkins PB (ноябрь 1997 г.). «Механизмы повышенной пероральной доступности субстратов CYP3A4 за счет компонентов грейпфрута. Снижение концентрации CYP3A4 в энтероцитах и основанная на механизмах инактивация фуранокумаринами». Метаболизм и утилизация лекарств. 25 (11): 1228–33. PMID 9351897.
- ^ а б c Шахрох К., Читам Т.Е., Йост Г.С. (октябрь 2012 г.). «Конформационная динамика CYP3A4 демонстрирует важную роль Arg212 в сочетании с открытием входных, выходных каналов и каналов растворителя для дегидрирования 4-гидрокситамоксифена». Biochimica et Biophysica Acta (BBA) - Общие предметы. 1820 (10): 1605–17. Дои:10.1016 / j.bbagen.2012.05.011. ЧВК 3404218. PMID 22677141.
- ^ а б Менье Б., де Виссер С. П., Шайк С. (сентябрь 2004 г.). «Механизм реакций окисления, катализируемых ферментами цитохрома p450». Химические обзоры. 104 (9): 3947–80. Дои:10.1021 / cr020443g. PMID 15352783. S2CID 33927145.
- ^ Хе К., Айер К.Р., Хейс Р.Н., Синз М.В., Вульф Т.Ф., Холленберг П.Ф. (апрель 1998 г.). «Инактивация цитохрома P450 3A4 бергамоттином, компонентом грейпфрутового сока». Химические исследования в токсикологии. 11 (4): 252–9. Дои:10.1021 / tx970192k. PMID 9548795.
- ^ а б Бейли Д.Г., Малкольм Дж., Арнольд О., Спенс Дж. Д. (август 1998 г.). «Взаимодействие грейпфрутового сока и лекарств». Британский журнал клинической фармакологии. 46 (2): 101–10. Дои:10.1046 / j.1365-2125.1998.00764.x. ЧВК 1873672. PMID 9723817.
- ^ Гарг С. К., Кумар Н., Бхаргава В. К., Прабхакар С. К. (сентябрь 1998 г.). «Влияние грейпфрутового сока на биодоступность карбамазепина у пациентов с эпилепсией». Клиническая фармакология и терапия. 64 (3): 286–8. Дои:10.1016 / S0009-9236 (98) 90177-1. PMID 9757152. S2CID 27490726.
- ^ Бейли Д.Г., Комод Г.К. (2004). «Взаимодействие грейпфрутового сока и сердечно-сосудистых препаратов». Американский журнал сердечно-сосудистых препаратов. 4 (5): 281–97. Дои:10.2165/00129784-200404050-00002. PMID 15449971. S2CID 11525439.
- ^ Бресслер Р. (ноябрь 2006 г.). «Грейпфрутовый сок и лекарственные взаимодействия. Изучение механизмов этого взаимодействия и потенциальной токсичности для определенных лекарств». Гериатрия. 61 (11): 12–8. PMID 17112309.
- ^ Lilja JJ, Kivistö KT, Neuvonen PJ (октябрь 2000 г.). «Продолжительность действия грейпфрутового сока на фармакокинетику субстрата CYP3A4 симвастатина». Клиническая фармакология и терапия. 68 (4): 384–90. Дои:10.1067 / mcp.2000.110216. PMID 11061578. S2CID 29029956.
- ^ «Интегративная медицина, Нони». Мемориальный онкологический центр Слоуна-Кеттеринга. Получено 2013-06-27.
- ^ Хидака М., Окумура М., Фудзита К., Огикубо Т., Ямасаки К., Ивакири Т., Сетогучи Н., Аримори К. (май 2005 г.). «Влияние сока граната на цитохром p450 3A (CYP3A) человека и фармакокинетику карбамазепина у крыс». Метаболизм и утилизация лекарств. 33 (5): 644–8. Дои:10.1124 / dmd.104.002824. PMID 15673597. S2CID 7997718.
- ^ Ли SJ, Goldstein JA (июнь 2005 г.). «Функционально дефектные или измененные однонуклеотидные полиморфизмы CYP3A4 и CYP3A5 и их обнаружение с помощью тестов генотипирования». Фармакогеномика. 6 (4): 357–71. Дои:10.1517/14622416.6.4.357. PMID 16004554.
- ^ Уоткинс ПБ (август 1994 г.). «Неинвазивные тесты ферментов CYP3A». Фармакогенетика. 4 (4): 171–84. Дои:10.1097/00008571-199408000-00001. PMID 7987401.
- ^ Ратаевски М., Вальчак-Джевецка А., Салковская А., Дастыч Ю. (август 2011 г.). «Афлатоксины повышают экспрессию мРНК CYP3A4 в процессе, в котором участвует фактор транскрипции PXR». Письма токсикологии. 205 (2): 146–53. Дои:10.1016 / j.toxlet.2011.05.1034. PMID 21641981.
- ^ Wolbold R, Klein K, Burk O, Nüssler AK, Neuhaus P, Eichelbaum M, Schwab M, Zanger UM (октябрь 2003 г.). «Пол - главный детерминант экспрессии CYP3A4 в печени человека». Гепатология. 38 (4): 978–88. Дои:10.1053 / jhep.2003.50393. PMID 14512885.
- ^ а б c d Гонсалес FJ (2007). «Гуманизированные мыши с рецептором CYP3A4 и прегнана X». Журнал биохимической и молекулярной токсикологии. 21 (4): 158–62. Дои:10.1002 / jbt.20173. PMID 17936928. S2CID 21501739.
- ^ а б Краго Дж., Клапер Р.Д. (сентябрь 2011 г.). «Влияние пола, режима кормления и продолжительности воздействия на экспрессию генов, связанных с метаболизмом ксенобиотиков у толстоголовых гольянов (Pimephales promelas)». Сравнительная биохимия и физиология. Токсикология и фармакология. 154 (3): 208–12. Дои:10.1016 / j.cbpc.2011.05.016. PMID 21664292.
- ^ а б Ян Дж., Ляо М., Шоу М., Джамей М., Йео К. Р., Такер Г. Т., Ростами-Ходжеган А. (июнь 2008 г.). «Оборот цитохрома p450: регуляция синтеза и деградации, методы определения скорости и значения для прогнозирования лекарственных взаимодействий». Текущий метаболизм лекарств. 9 (5): 384–94. Дои:10.2174/138920008784746382. PMID 18537575.
- ^ Севрюкова И.Ф., Поулос Т.Л. (январь 2012 г.). «Структурные и механистические сведения о взаимодействии цитохрома P4503A4 с бромэргокриптином, лигандом типа I». Журнал биологической химии. 287 (5): 3510–7. Дои:10.1074 / jbc.M111.317081. ЧВК 3271004. PMID 22157006.
- ^ а б Дас А., Чжао Дж., Шац Г.С., Слигар С.Г., Ван Дайн Р.П. (май 2009 г.). «Скрининг связывания лекарственных средств типа I и II с цитохромом P450-3A4 человека на нанодисках с помощью спектроскопии локального поверхностного плазмонного резонанса». Аналитическая химия. 81 (10): 3754–9. Дои:10.1021 / ac802612z. ЧВК 4757437. PMID 19364136.
- ^ а б c d е ж г час я j k л м п о п q р s т ты v ш Икс у z аа ab ac объявление ае аф аг ах ай эй ак аль я ан ао ap водный ар так как в au средний ау топор ай az ба bb до н.э bd быть парень bg бх би Ъ bk бл бм млрд бо бп бк br bs bt бу bv чб bx от bz ок cb cc компакт диск ce ср cg ch ci cj ск cl см сп co cp cq cr cs ct у.е. резюме cw сх Сай cz да db Округ Колумбия дд де df dg dh ди диджей Флокхарт Д.А. (2007). «Лекарственное взаимодействие: цитохром P450 Таблица взаимодействия лекарств ». Медицинский факультет Университета Индианы. Проверено 25 декабря, 2008.
- ^ Если классы агентов перечислены, внутри класса могут быть исключения.
- ^ а б c d е ж г час я j k л м п о п q р s т ты v ш Икс у z аа ab ac объявление ае аф аг ах ай эй ак аль я ан ао ap водный ар так как в au средний ау топор ай az ба bb до н.э bd быть парень bg бх би Ъ bk бл бм млрд бо бп бк FASS (фармакологический справочник): Шведская экологическая классификация фармацевтических препаратов Факты для врачей (Fakta för förskrivare). Проверено июль 2011 г.
- ^ «Эрлотиниб».
Метаболизируется главным образом CYP3A4 и, в меньшей степени, CYP1A2 и внепеченочной изоформой CYP1A1.
- ^ «Циклобензаприн». DrugBank.
- ^ Moody DE, Fang WB, Lin SN, Weyant DM, Strom SC, Omiecinski CJ (декабрь 2009 г.). «Влияние рифампицина и нелфинавира на метаболизм метадона и бупренорфина в первичных культурах гепатоцитов человека». Метаболизм и утилизация лекарств. 37 (12): 2323–9. Дои:10.1124 / dmd.109.028605. ЧВК 2784702. PMID 19773542.
- ^ Hutchinson MR, Menelaou A, Foster DJ, Coller JK, Somogyi AA (март 2004 г.). «Участие CYP2D6 и CYP3A4 в первичном окислительном метаболизме гидрокодона микросомами печени человека». Британский журнал клинической фармакологии. 57 (3): 287–97. Дои:10.1046 / j.1365-2125.2003.02002.x. ЧВК 1884456. PMID 14998425.
- ^ Танака, Э. (октябрь 1999 г.). «Клинически значимые фармакокинетические лекарственные взаимодействия с бензодиазепинами». Журнал клинической фармации и терапии. 24 (5): 347–355. Дои:10.1046 / j.1365-2710.1999.00247.x. ISSN 0269-4727. PMID 10583697. S2CID 22229823.
- ^ Саттон Д., Батлер А.М., Надин Л., Мюррей М. (июль 1997 г.). «Роль CYP3A4 в N-деметилировании дилтиазема в печени человека: ингибирование активности CYP3A4 окисленными метаболитами дилтиазема». Журнал фармакологии и экспериментальной терапии. 282 (1): 294–300. PMID 9223567.
- ^ «Разработка лекарств и их взаимодействия: таблица субстратов, ингибиторов и индукторов». Домашняя страница Управления по санитарному надзору за качеством пищевых продуктов и медикаментов США. 2009-06-25. Получено 2019-02-01.
- ^ Лаун К.С., Бейли Д.Г., Фонтана Р.Дж., Джанардан С.К., Адаир СН, Фортлаге Л.А., Браун М.Б., Гуо В., Уоткинс П.Б. (май 1997 г.). «Грейпфрутовый сок увеличивает пероральную доступность фелодипина для людей за счет снижения экспрессии белка CYP3A в кишечнике». Журнал клинических исследований. Американское общество клинических исследований. 99 (10): 2545–53. Дои:10.1172 / jci119439. ЧВК 508096. PMID 9153299.
- ^ Бейли Д.Г., Бенд-младший, Арнольд Дж. М., Тран Л. Т., Спенс Дж. Д. (июль 1996 г.). «Взаимодействие эритромицина и фелодипина: величина, механизм и сравнение с грейпфрутовым соком». Клиническая фармакология и терапия. Springer Nature. 60 (1): 25–33. Дои:10.1016 / с0009-9236 (96) 90163-0. PMID 8689808. S2CID 1246705.
- ^ Guengerich FP, Brian WR, Iwasaki M, Sari MA, Bäärnhielm C, Berntsson P (июнь 1991 г.). «Окисление дигидропиридиновых блокаторов кальциевых каналов и аналогов цитохромом печени человека P-450 IIIA4». Журнал медицинской химии. 34 (6): 1838–44. Дои:10.1021 / jm00110a012. PMID 2061924.
- ^ Като М., Накадзима М., Ямадзаки Х., Ёкои Т. (февраль 2001 г.). «Подавляющее действие субстратов CYP3A4 и их метаболитов на опосредованный Р-гликопротеином транспорт». Европейский журнал фармацевтических наук. 12 (4): 505–13. Дои:10.1016 / s0928-0987 (00) 00215-3. PMID 11231118.
- ^ Фоти RS, Rock DA, Wienkers LC, Wahlstrom JL (июнь 2010 г.). «Выбор альтернативных субстратов зондов CYP3A4 для клинических исследований взаимодействия лекарственных средств с использованием данных in vitro и моделирования in vivo». Метаболизм и утилизация лекарств. Американское общество фармакологии и экспериментальной терапии (ASPET). 38 (6): 981–7. Дои:10.1124 / dmd.110.032094. PMID 20203109. S2CID 6823063.
- ^ Odou P, Ferrari N, Barthélémy C, Brique S, Lhermitte M, Vincent A, Libersa C, Robert H (апрель 2005 г.). «Взаимодействие грейпфрутового сока и нифедипина: возможное участие нескольких механизмов». Журнал клинической фармации и терапии. 30 (2): 153–8. Дои:10.1111 / j.1365-2710.2004.00618.x. PMID 15811168. S2CID 30463290.
- ^ «NIFEDIPINE EXTENDED RELEASE- таблетка нифедипина, пролонгированного высвобождения». DailyMed. 2012-11-29. Получено 2019-02-01.
Взаимодействие с лекарствами: Нифедипин в основном выводится путем метаболизма и является субстратом CYP3A. Ингибиторы и индукторы CYP3A могут влиять на воздействие нифедипина и, следовательно, на его желательные и нежелательные эффекты. Данные in vitro и in vivo показывают, что нифедипин может ингибировать метаболизм препаратов, являющихся субстратами CYP3A, тем самым увеличивая воздействие других препаратов. Нифедипин является вазодилататором, и одновременный прием других препаратов, влияющих на артериальное давление, может привести к фармакодинамическим взаимодействиям.
- ^ Чжан И, Го Х, Линь Е.Т., Бенет Л.З. (апрель 1998 г.). «Перекрывающиеся субстратные специфичности цитохрома P450 3A и Р-гликопротеина для нового ингибитора цистеиновой протеазы». Метаболизм и утилизация лекарств. 26 (4): 360–6. PMID 9531525.
- ^ Стрингер К.А., Маллет Дж., Кларк М., Линденфельд Дж. А. (1992). «Влияние трех различных пероральных доз верапамила на расположение теофиллина». Европейский журнал клинической фармакологии. 43 (1): 35–8. Дои:10.1007 / bf02280751. PMID 1505606. S2CID 8942097.
- ^ Нильсен-Кудск Дж. Э., Буль Дж. С., Йоханнесен А. С. (февраль 1990 г.). «Верапамил-индуцированное ингибирование выведения теофиллина у здоровых людей». Фармакология и токсикология. 66 (2): 101–3. Дои:10.1111 / j.1600-0773.1990.tb00713.x. PMID 2315261.
- ^ Gin AS, Stringer KA, Welage LS, Wilton JH, Matthews GE (август 1989 г.). «Влияние верапамила на фармакокинетическое расположение теофиллина у курильщиков сигарет». Журнал клинической фармакологии. 29 (8): 728–32. Дои:10.1002 / j.1552-4604.1989.tb03407.x. PMID 2778093. S2CID 20446675.
- ^ Сирманс С.М., Пипер Дж. А., Лалонд Р. Л., Смит Д. Г., Self TH (июль 1988 г.). «Влияние блокаторов кальциевых каналов на расположение теофиллина». Клиническая фармакология и терапия. 44 (1): 29–34. Дои:10.1038 / clpt.1988.108. PMID 3391002. S2CID 39570845.
- ^ Робсон Р.А., Шахтеры Дж. О., Биркетт Д. Д. (март 1988 г.). «Селективное подавляющее действие нифедипина и верапамила на окислительный метаболизм: влияние на теофиллин». Британский журнал клинической фармакологии. 25 (3): 397–400. Дои:10.1111 / j.1365-2125.1988.tb03319.x. ЧВК 1386365. PMID 3358901.
- ^ Абернети Д.Р., Иган Дж.М., Дикинсон Т.Х., Каррам Дж. (Март 1988 г.). «Субстрат-селективное ингибирование верапамилом и дилтиаземом: дифференциальное расположение антипирина и теофиллина у людей». Журнал фармакологии и экспериментальной терапии. 244 (3): 994–9. PMID 3252045.
- ^ а б Като М., Накадзима М., Ямадзаки Х., Ёкои Т. (октябрь 2000 г.). «Ингибирующие способности 1,4-дигидропиридиновых антагонистов кальция к транспорту, опосредованному P-гликопротеином: сравнение с эффектами на CYP3A4». Фармацевтические исследования. 17 (10): 1189–97. Дои:10.1023 / а: 1007568811691. PMID 11145223. S2CID 24304693.
- ^ Druglib.com[требуется полная цитата ]
- ^ Cockshott ID (2004 г.). «Бикалутамид: клиническая фармакокинетика и метаболизм». Клиническая фармакокинетика. 43 (13): 855–78. Дои:10.2165/00003088-200443130-00003. PMID 15509184.
- ^ Мацумото С., Ямазо Ю. (февраль 2001 г.). «Участие нескольких цитохромов человека P450 в микросомальном метаболизме астемизола в печени и сравнение с терфенадином». Британский журнал клинической фармакологии. 51 (2): 133–42. Дои:10.1046 / j.1365-2125.2001.01292.x. ЧВК 2014443. PMID 11259984.
- ^ Леджер, Томас; Тонг, Винни (1 августа 2019 г.). «Лечебный сбой: ятрогенный синдром Кушинга с ингаляционным флутиказоном». Австралийский врач. 42 (4): 139–140. Дои:10.18773 / austprescr.2019.040. ЧВК 6698236. PMID 31427846.
- ^ Фермент 1.14.13.32 в КЕГГ
- ^ «Показан белок цитохрома P450 3A4 (HMDBP01018)». База данных метаболома человека. Получено 2017-08-05.
- ^ Дейли А.К., King BP (май 2003 г.). «Фармакогенетика пероральных антикоагулянтов». Фармакогенетика. 13 (5): 247–52. Дои:10.1097/00008571-200305000-00002. PMID 12724615.
- ^ Лау В.К., Васкелл Л.А., Уоткинс П.Б., Нир С.Дж., Горовиц К., Хопп А.С., Тейт А.Р., Карвилл Д.Г., Гуйер К.Э., Бейтс ER (январь 2003 г.). «Аторвастатин снижает способность клопидогреля ингибировать агрегацию тромбоцитов: новое лекарственное средство взаимодействия». Тираж. 107 (1): 32–7. Дои:10.1161 / 01.CIR.0000047060.60595.CC. PMID 12515739.
- ^ Meyer MR, Bach M, Welter J, Bovens M, Turcant A, Maurer HH (июль 2013 г.). «Кетаминовый дизайнерский препарат метоксетамин: метаболизм, включая кинетику изоферментов и токсикологическую обнаруживаемость с использованием ГХ-МС и ЖХ- (HR-) МСн». Аналитическая и биоаналитическая химия. 405 (19): 6307–21. Дои:10.1007 / s00216-013-7051-6. PMID 23774830. S2CID 27966043.
- ^ «ЛОСАРТАН - лозартан калия в таблетках, покрытых пленочной оболочкой». DailyMed. 2018-12-26. Получено 2019-02-06.
- ^ а б c d е ж «Разработка лекарств и их взаимодействия: таблица субстратов, ингибиторов и индукторов».
- ^ а б c d е ж Флауэр Р., Ранг HP, Дейл М.М., Риттер Дж. М. (2007). Фармакология Рэнга и Дейла. Эдинбург: Черчилль Ливингстон. ISBN 978-0-443-06911-6.[страница нужна ]
- ^ Пак Дж.Й., Ким К.А., Ким С.Л. (ноябрь 2003 г.). «Хлорамфеникол является сильным ингибитором изоформ цитохрома P450 CYP2C19 и CYP3A4 в микросомах печени человека». Противомикробные препараты и химиотерапия. 47 (11): 3464–9. Дои:10.1128 / AAC.47.11.3464-3469.2003. ЧВК 253795. PMID 14576103.
- ^ а б c d е ж г час я j k л м п о п q Исследования, Центр оценки лекарственных средств и. «Взаимодействие с лекарствами и маркировка - Разработка лекарств и взаимодействия с ними: таблица субстратов, ингибиторов и индукторов». www.fda.gov. Получено 2018-08-06.
- ^ Каракурт, С. (2016). «Модулирующие эффекты рутина на экспрессию цитохрома P450s и антиоксидантных ферментов в клетках гепатомы человека». Acta Pharmaceutica (Загреб, Хорватия). 66 (4): 491–502. Дои:10.1515 / acph-2016-0046. PMID 27749250. S2CID 20274417.
- ^ Ashour, M. L .; Youssef, F. S .; Gad, H.A .; Винк, М. (2017). «Ингибирование активности цитохрома P450 (CYP3A4) экстрактами 57 растений, используемых в традиционной китайской медицине (ТКМ)». Журнал Pharmacognosy. 13 (50): 300–308. Дои:10.4103/0973-1296.204561. ЧВК 5421430. PMID 28539725.
- ^ Информация о продукте: Буккальные таблетки ORAVIG®, буккальные таблетки миконазола. Praelia Pharmaceuticals, Inc (согласно FDA), Кэри, Северная Каролина, 2013 г.
- ^ http://www.rxlist.com/valerian-page3/supplements.htm#Interactions[требуется полная цитата ]
- ^ Чжан В., Рамамурти Ю., Тиндейл РФ, Селлерс Е.М. (июнь 2003 г.). «Взаимодействие бупренорфина и его метаболита норбупренорфина с цитохромами p450 in vitro». Метаболизм и утилизация лекарств. 31 (6): 768–72. Дои:10.1124 / dmd.31.6.768. PMID 12756210. S2CID 16229370.
- ^ http://www.aapsj.org/abstracts/AM_2009/AAPS2009-001235.PDF[требуется полная цитата ] В архиве 21 июля 2011 г. Wayback Machine
- ^ Вен X, Ван Дж. С., Кивисто К. Т., Неувонен П. Дж., Бэкман Дж. Т. (2001). «Оценка in vitro вальпроевой кислоты как ингибитора изоформ цитохрома P450 человека: преимущественное ингибирование цитохрома P450 2C9 (CYP2C9)». Br J Clin Pharmacol. 52 (5): 547–53. Дои:10.1046 / j.0306-5251.2001.01474.x. ЧВК 2014611. PMID 11736863.
- ^ [1] Сильное ингибирование изоформ цитохрома P450 3A человека каннабидиолом: роль фенольных гидроксильных групп в фрагменте резорцина
- ^ а б Было показано, что ненуклеозидные ингибиторы обратной транскриптазы как индуцируют, так и ингибируют CYP3A4.
- ^ Хидака М., Фудзита К., Огикубо Т., Ямасаки К., Ивакири Т., Окумура М., Кодама Х., Аримори К. (июнь 2004 г.). «Сильное ингибирование карамболой активности человеческого цитохрома P450 3A (CYP3A)». Метаболизм и утилизация лекарств. 32 (6): 581–3. Дои:10.1124 / dmd.32.6.581. PMID 15155547. S2CID 17392051.
- ^ HCVadvocate.org В архиве 2010-03-05 на Wayback Machine[требуется полная цитата ]
- ^ Годино К., Оклер К. (май 2004 г.). «Ингибирование человеческих ферментов P450 никотиновой кислотой и никотинамидом». Сообщения о биохимических и биофизических исследованиях. 317 (3): 950–6. Дои:10.1016 / j.bbrc.2004.03.137. PMID 15081432.
- ^ Кимура Ю., Ито Х, Охниши Р., Хатано Т. (январь 2010 г.). «Ингибирующее действие полифенолов на активность цитохрома P450 3A4 и 2C9 человека». Пищевая и химическая токсикология. 48 (1): 429–35. Дои:10.1016 / j.fct.2009.10.041. PMID 19883715.
Было показано, что гинко билоба содержит мощный ингибитор аментофлавон.
- ^ Lim, Y. P .; Ma, C. Y .; Liu, C.L .; Lin, Y.H .; Hu, M. L .; Chen, J. J .; Hung, D. Z .; Hsieh, W. T .; Хуанг, Дж. Д. (2012). «Сезамин: естественный лигнан ингибирует CYP3A4, противодействуя активации рецептора прегнана X». Доказательная дополнительная и альтернативная медицина: Ecam. 2012: 242810. Дои:10.1155/2012/242810. ЧВК 3356939. PMID 22645625.
- ^ Бхардвадж Р.К., Глэзер Х., Бекмонт Л., Клотц Ю., Гупта С.К., Фромм М.Ф. (август 2002 г.). «Пиперин, основной компонент черного перца, подавляет человеческий P-гликопротеин и CYP3A4». Журнал фармакологии и экспериментальной терапии. 302 (2): 645–50. Дои:10.1124 / jpet.102.034728. PMID 12130727. S2CID 7398172.
- ^ Вен X, Ван Дж. С., Неувонен П. Дж., Бэкман Дж. Т. (январь 2002 г.). «Изониазид - это механизм, основанный на ингибиторе изоформ цитохрома P450 1A2, 2A6, 2C19 и 3A4 в микросомах печени человека». Европейский журнал клинической фармакологии. 57 (11): 799–804. Дои:10.1007 / s00228-001-0396-3. PMID 11868802. S2CID 19299097.
- ^ https://www.accessdata.fda.gov/drugsatfda_docs/label/2012/203415lbl.pdf
- ^ Йоханнесен С.И., Landmark CJ (сентябрь 2010 г.). «Противоэпилептические лекарственные взаимодействия - принципы и клиническое значение». Современная нейрофармакология. 8 (3): 254–67. Дои:10.2174/157015910792246254. ЧВК 3001218. PMID 21358975.
- ^ Nallani SC, Glauser TA, Hariparsad N, Setchell K, Buckley DJ, Buckley AR, Desai PB (декабрь 2003 г.). «Дозозависимая индукция цитохрома P450 (CYP) 3A4 и активация рецептора прегнана X топираматом». Эпилепсия. 44 (12): 1521–8. Дои:10.1111 / j.0013-9580.2003.06203.x. PMID 14636322. S2CID 6915760.
- ^ Хан Э.Х., Ким Х.Г., Чхве Дж.Х., Чан Й.Дж., Ли С.С., Квон К.И., Ким И, Но К, Чжон ТК, Хван Й.П., Чунг Ю.К., Кан В., Чжон Х.Г. (май 2012 г.). «Капсаицин индуцирует экспрессию CYP3A4 через рецептор прегнана X и β-активацию CCAAT / энхансер-связывающего белка». Молекулярное питание и пищевые исследования. 56 (5): 797–809. Дои:10.1002 / mnfr.201100697. PMID 22648626. S2CID 26584141.
внешние ссылки
- PharmGKB: аннотированная информация о гене PGx для CYP3A4
- Прогнозирование субстрата CYP3A4
- Человек CYP3A4 расположение генома и CYP3A4 страница сведений о генах в Браузер генома UCSC.
- Обзор всей структурной информации, доступной в PDB для UniProt: P08684 (Цитохром P450 3A4) на PDBe-KB.
Эта статья включает текст из Национальная медицинская библиотека США, который находится в всеобщее достояние.
Галерея PDB | |
|---|---|
|