Гомеостаз - Homeostasis
В биология, гомеостаз состояние устойчивого внутреннего, физический, и химический условия поддерживаются живые системы.[1] Это условие оптимального функционирования организма, включающее множество переменных, таких как температура тела и баланс жидкости, находясь в определенных заранее заданных пределах (гомеостатический диапазон). Другие переменные включают pH из внеклеточной жидкости, концентрации натрий, калий и кальций ионы, а также уровень сахара в крови, и их необходимо регулировать, несмотря на изменения в окружающей среде, диете или уровне активности. Каждая из этих переменных контролируется одним или несколькими регуляторами или гомеостатическими механизмами, которые вместе поддерживают жизнь.
Гомеостаз обеспечивается естественным сопротивлением изменениям, когда он уже находится в оптимальных условиях,[2] а равновесие поддерживается многими регулирующими механизмами. Все механизмы гомеостатического контроля имеют по крайней мере три взаимозависимых компонента регулируемой переменной: рецептор, центр управления и эффектор.[3] Рецептор - это чувствительный компонент, который отслеживает и реагирует на изменения в окружающей среде, будь то внешние или внутренние. Рецепторы включают терморецепторы, и механорецепторы. Центры управления включают дыхательный центр, а ренин-ангиотензиновая система. Эффектор - это цель, на которую воздействуют, чтобы вернуть изменение в нормальное состояние. На клеточном уровне рецепторы включают: ядерные рецепторы которые вызывают изменения в экспрессия гена через повышающее или понижающее регулирование и действовать в негативный отзыв механизмы. Примером этого является управление желчные кислоты в печень.[4]
Некоторые центры, такие как ренин-ангиотензиновая система, управляйте более чем одной переменной. Когда рецептор ощущает стимул, он реагирует, посылая потенциалы действия в центр управления. Центр управления устанавливает диапазон обслуживания - допустимые верхний и нижний пределы - для конкретной переменной, например температуры. Центр управления реагирует на сигнал, определяя соответствующий ответ и отправляя сигналы в эффектор, который может быть одной или несколькими мышцами, органом или железа. Когда сигнал получен и на него воздействуют, рецептору предоставляется отрицательная обратная связь, которая устраняет необходимость в дальнейшей передаче сигналов.[5]
В каннабиноидный рецептор типа 1 (CB1), расположенный в пресинаптический нейрон, это рецептор что может остановить стресс нейротрансмиттер выпуск в постсинаптический нейрон; он активируется эндоканнабиноиды (EC), такие как анандамид (N-арахидоноилэтаноламид; AEA) и 2-арахидоноилглицерин (2-AG) через ретроградная сигнализация процесс, в котором эти соединения синтезируются постсинаптическими нейронами и высвобождаются из них и возвращаются к пресинаптическому окончанию, чтобы связываться с рецептором CB1 для модуляции высвобождения нейромедиатора для достижения гомеостаза.[6]
В полиненасыщенные жирные кислоты (ПНЖК) являются липид производные от Омега 3 (докозагексаеновая кислота, DHA, и эйкозапентаеновая кислота, EPA ) или омега-6 (арахидоновая кислота, ARA ) синтезируются из мембрана фосфолипиды и используемый в качестве предшественника эндоканнабиноидов (ЭК), оказывает значительное влияние на точную настройку гомеостаза тела.[7]
История
Концепция регуляции внутренней среды была описана французским физиологом. Клод Бернард в 1849 г., а слово гомеостаз был придуман Уолтер Брэдфорд Кэннон в 1926 г.[8][9] В 1932 г. Джозеф Баркрофт британский физиолог первым сказал, что выше мозг функция требовала максимально стабильной внутренней среды. Таким образом, для Баркрофта гомеостаз организовывался не только мозгом - гомеостаз служил мозгу.[10] Гомеостаз - это почти исключительно биологический термин, относящийся к концепциям, описанным Бернардом и Кэнноном, относительно постоянства внутренней среды, в которой клетки тела живут и выживают.[8][9][11] Период, термин кибернетика применяется к технологическим Системы управления Такие как термостаты, которые действуют как гомеостатические механизмы, но часто определяются гораздо шире, чем биологический термин гомеостаза.[5][12][13][14]
Этимология
Слово гомеостаз (/ˌчасoʊмяoʊˈsтeɪsɪs/[15][16]) использует комбинирование форм из гомео- и -стаз, Новая латынь из Греческий: ὅμοιος homoios, "похожие" и στάσις застой, «стоять на месте», что приводит к идее «оставаться прежним».
Обзор
В метаболические процессы всех организмов может иметь место только в очень специфических физических и химических средах. Условия различаются для каждого организма и от того, происходят ли химические процессы внутри клетка или в тканевая жидкость купание клеток. Наиболее известными гомеостатическими механизмами у людей и других млекопитающих являются регуляторы, которые поддерживают состав внеклеточной жидкости (или «внутренней среды») константа, особенно в отношении температура, pH, осмоляльность, а концентрации натрий, калий, глюкоза, углекислый газ, и кислород. Однако множество других гомеостатических механизмов, охватывающих многие аспекты физиология человека, контролировать другие сущности в теле. Если уровни переменных выше или ниже необходимых, они часто начинаются с префикса гипер- и гипо-соответственно такие как гипертермия и переохлаждение или же гипертония и гипотония.

Если объект находится под гомеостатическим контролем, это не означает, что его ценность обязательно абсолютно стабильна для здоровья. Внутренняя температура тела регулируется, например, гомеостатическим механизмом с датчиками температуры, среди прочего, в гипоталамус из мозг.[17] Тем не менее уставка регулятора регулярно сбрасывается.[18] Например, внутренняя температура тела у человека меняется в течение дня (т.е. имеет циркадный ритм ), с самыми низкими температурами в ночное время и самыми высокими во второй половине дня. Другой нормальный колебания температуры включать те, которые связаны с менструальный цикл.[19][20] Уставка регулятора температуры сбрасывается во время инфекции, чтобы вызвать повышение температуры.[17][21][22] Организмы способны приспосабливаться к различным условиям, таким как изменения температуры или уровень кислорода на высоте, с помощью процесса акклиматизация.
Гомеостаз не влияет на все действия в организме.[23][24] Например, сигнал (будь то через нейроны или же гормоны ) от датчика к исполнительному элементу обязательно сильно варьируется, чтобы передавать Информация о направлении и величине ошибки, обнаруженной датчиком.[25][26][27] Точно так же реакция эффектора должна быть легко регулируемой, чтобы обратить ошибку - на самом деле она должна быть почти пропорциональна (но в противоположном направлении) ошибке, которая угрожает внутренней среде.[13][14] Например, артериальное давление у млекопитающих контролируется гомеостатически и измеряется рецепторы растяжения в стенах дуга аорты и каротидные синусы в начале внутренние сонные артерии.[17] Датчики отправляют сообщения через сенсорные нервы к продолговатый мозг мозга, указывая, артериальное давление упал или поднялся, и на сколько. Затем продолговатый мозг распределяет сообщения по двигательные или эфферентные нервы принадлежащий к автономная нервная система к широкому спектру эффекторных органов, активность которых, следовательно, изменяется, чтобы обратить вспять ошибку кровяного давления. Одним из эффекторных органов является сердце, частота которого стимулируется к повышению (тахикардия ) при падении артериального давления или замедлении (брадикардия ), когда давление поднимается выше заданного значения.[17] Таким образом, частота сердечных сокращений (для которой нет датчика в организме) не контролируется гомеостатически, а является одной из эффекторных реакций на ошибки в артериальном кровяном давлении. Другой пример - скорость потливость. Это один из факторов, влияющих на гомеостатический контроль температуры тела, и поэтому он сильно варьируется в грубой пропорции с тепловой нагрузкой, которая угрожает дестабилизировать внутреннюю температуру тела, для чего есть датчик в гипоталамус мозга.
Управление переменными
Температура ядра

Млекопитающие регулировать их температура ядра используя ввод от терморецепторы в гипоталамус, мозг,[17][28] спинной мозг, внутренние органы, и большие вены.[29][30] Помимо внутреннего регулирования температуры, процесс, называемый аллостаз может вступить в игру, которая регулирует поведение, чтобы адаптироваться к проблеме очень жарких или холодных крайностей (и к другим проблемам).[31] Эти корректировки могут включать поиск тени и снижение активности, поиск более теплых условий и увеличение активности, или скопление людей.[32]Поведенческая терморегуляция имеет приоритет над физиологической терморегуляцией, поскольку необходимые изменения могут быть затронуты быстрее, а способность физиологической терморегуляции реагировать на экстремальные температуры ограничена.[33]
Когда внутренняя температура падает, кровоснабжение кожи уменьшается из-за интенсивного вазоконстрикция.[17] Приток крови к конечностям (которые имеют большую площадь поверхности) аналогичным образом уменьшается и возвращается в туловище через глубокие вены, расположенные рядом с артериями (формируя Venae Comitantes ).[28][32][34] Это действует как система противоточного обмена который передает тепло артериальной крови непосредственно в венозную кровь, возвращающуюся в туловище, вызывая минимальные потери тепла конечностями в холодную погоду.[28][32][35] Подкожные вены конечностей сильно сужены,[17] не только уменьшая теплопотери от этого источника, но и заставляя венозную кровь попадать в противоточную систему в глубине конечностей.
Скорость метаболизма увеличивается, сначала из-за отсутствия дрожи. термогенез,[36] с последующим дрожащий термогенез если более ранние реакции недостаточны для исправления переохлаждение.
Когда повышение внутренней температуры обнаруживается терморецепторы, то потовые железы в коже стимулируются через холинергический симпатические нервы выделять пот на кожу, которая при испарении охлаждает кожу и кровь, протекающую по ней. Одышка является альтернативным эффектором у многих позвоночных, который охлаждает тело также за счет испарения воды, но на этот раз от слизистые оболочки горла и рта.
Глюкоза в крови

Содержание сахара в крови уровни регулируемый в довольно узких пределах.[37] У млекопитающих основными датчиками для этого являются бета-клетки из островки поджелудочной железы.[38][39] Бета-клетки реагируют на повышение уровня сахара в крови, секретируя инсулин в кровь, и одновременно подавляя соседние альфа-клетки от секрета глюкагон в кровь.[38] Эта комбинация (высокий уровень инсулина в крови и низкий уровень глюкагона) действует на эффекторные ткани, главными из которых являются печень, жировые клетки и мышечные клетки. Печень не может производить глюкоза, взяв его вместо этого и преобразовав в гликоген и триглицериды. Гликоген накапливается в печени, но триглицериды секретируются в кровь в виде липопротеины очень низкой плотности (VLDL) частицы, которые поглощаются жировая ткань, там храниться в виде жиров. Жировые клетки поглощают глюкозу через специальные транспортеры глюкозы (GLUT4 ), количество которых в клеточной стенке увеличивается в результате прямого воздействия инсулина на эти клетки. Глюкоза, которая поступает в жировые клетки таким образом, превращается в триглицериды (через те же метаболические пути, которые используются в печени), а затем сохраняется в этих жировых клетках вместе с триглицеридами, производными от ЛПОНП, которые были произведены в печени. Мышечные клетки также поглощают глюкозу через инсулино-чувствительные каналы глюкозы GLUT4 и превращают ее в мышечный гликоген.
Падение уровня глюкозы в крови вызывает остановку секреции инсулина и глюкагон секретироваться альфа-клетками в кровь. Это препятствует захвату глюкозы из крови печенью, жировыми клетками и мышцами. Вместо этого печень сильно стимулируется к производству глюкозы из гликогена (через гликогенолиз ) и из неуглеводных источников (таких как лактат и деаминированный аминокислоты ) с использованием процесса, известного как глюконеогенез.[40] Произведенная таким образом глюкоза сбрасывается в кровь, исправляя обнаруженную ошибку (гипогликемия ). Гликоген, хранящийся в мышцах, остается в мышцах и расщепляется только во время упражнений на глюкозо-6-фосфат и оттуда пируват быть поданным в цикл лимонной кислоты или превратился в лактат. В кровь возвращаются только лактат и продукты жизнедеятельности цикла лимонной кислоты. Печень может поглощать только лактат, и в процессе потребления энергии глюконеогенез преобразовать обратно в глюкозу.
Уровни железа
Эта секция нуждается в расширении. Вы можете помочь добавляя к этому. (Ноябрь 2017 г.) |
Медное регулирование
Эта секция нуждается в расширении. Вы можете помочь добавляя к этому. (Апрель 2018 г.) |
Уровни газов крови
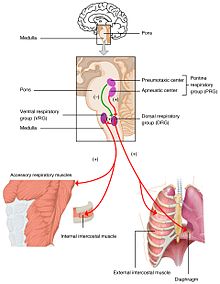
Изменения уровня кислорода, углекислого газа и pH плазмы отправляются в дыхательный центр, в мозговой ствол где они регулируются. частичное давление из кислород и углекислый газ в артериальная кровь контролируется периферические хеморецепторы (ПНС ) в сонная артерия и дуга аорты. Изменение в парциальное давление диоксида углерода определяется как изменение pH в спинномозговая жидкость к центральные хеморецепторы (ЦНС ) в продолговатый мозг из мозговой ствол. Информация от этих наборов датчиков отправляется в дыхательный центр, который активирует эффекторные органы - диафрагма и другие мышцы дыхания. Повышенный уровень углекислого газа в крови или пониженный уровень кислорода приведет к более глубокому дыханию и увеличению частота дыхания чтобы вернуть газы крови к равновесию.
Слишком мало углекислого газа и, в меньшей степени, слишком много кислорода в крови может временно остановить дыхание, состояние, известное как апноэ, который фридайверы используйте, чтобы продлить время пребывания под водой.
В парциальное давление диоксида углерода является более решающим фактором при мониторинге pH.[41] Однако на большой высоте (более 2500 м) мониторинг парциального давления кислорода имеет приоритет, и гипервентиляция поддерживает постоянный уровень кислорода. При более низком уровне углекислого газа, чтобы поддерживать pH на уровне 7,4, почки выделяют ионы водорода в кровь и выводят бикарбонат с мочой.[42][43] Это важно в акклиматизация к большой высоте.[44]
Содержание кислорода в крови
Почки измеряют содержание кислорода, а не парциальное давление кислорода в артериальной крови. Когда содержание кислорода в крови хронически низкий, чувствительные к кислороду клетки секретируют эритропоэтин (ЭПО) в кровь.[45] Эффекторная ткань - это красный костный мозг который производит красные кровяные тельца (RBCs) (эритроциты). Увеличение количества эритроцитов приводит к увеличению гематокрит в крови, и последующее повышение гемоглобин что увеличивает пропускную способность кислорода. Это механизм, благодаря которому у жителей высокогорья уровень гематокрита выше, чем у жителей уровня моря, а также почему люди с легочная недостаточность или же шунты справа налево в сердце (через которое венозная кровь проходит через легкие и попадает непосредственно в системный кровоток) имеют столь же высокий гематокрит.[46][47]
Независимо от парциального давления кислорода в крови количество переносимого кислорода зависит от содержания гемоглобина. Парциальное давление кислорода может быть достаточным, например, в анемия, но содержание гемоглобина будет недостаточным, а следовательно, и содержание кислорода. Имея достаточно железа, витамин B12 и фолиевая кислота, EPO может стимулировать производство эритроцитов, а содержание гемоглобина и кислорода восстанавливается до нормального уровня.[46][48]
Артериальное кровяное давление
Мозг может регулировать кровоток в диапазоне значений артериального давления с помощью вазоконстрикция и расширение сосудов артерий.[49]
Рецепторы высокого давления называются барорецепторы в стенах дуга аорты и каротидный синус (в начале внутренняя сонная артерия ) контролировать артериальную артериальное давление.[50] Повышение давления обнаруживается, когда стенки артерий растягиваются из-за увеличения объем крови. Это вызывает клетки сердечной мышцы выделять гормон предсердный натрийуретический пептид (ANP) в кровь. Это действует на почки, подавляя секрецию ренина и альдостерона, вызывая выброс натрия и сопутствующей воды в мочу, тем самым уменьшая объем крови.[51]Затем эта информация передается через афферентные нервные волокна, в одиночное ядро в продолговатый мозг.[52] Отсюда двигательные нервы принадлежащий к автономная нервная система стимулируются, чтобы влиять на деятельность в основном сердца и артерий наименьшего диаметра, называемых артериолы. Артериолы - главные сосуды сопротивления в артериальное дерево, а небольшие изменения диаметра вызывают большие изменения сопротивления потоку через них. Когда артериальное давление повышается, артериолы стимулируются к расширять облегчая выход крови из артерий, тем самым сдувая их и возвращая артериальное давление к норме. В то же время сердце стимулируется через холинергический парасимпатические нервы бить медленнее (называется брадикардия ), обеспечивая уменьшение притока крови в артерии, что способствует снижению давления и исправлению исходной ошибки.
Низкое давление в артериях вызывает противоположный рефлекс сужения артериол и учащение пульса (называемое тахикардия ). Если артериальное давление падает очень быстро или чрезмерно, продолговатый мозг стимулирует мозговое вещество надпочечников, через "преганглионарный" симпатические нервы, секретировать адреналин (адреналин) в кровь. Этот гормон усиливает тахикардию и вызывает тяжелые вазоконстрикция артериол ко всем, кроме основных органов тела (особенно сердца, легких и головного мозга). Эти реакции обычно корректируют низкое артериальное давление (гипотония ) очень эффективно.
Уровни кальция

Ионизированный кальций плазмы (Ca2+) концентрация очень жестко контролируется парой гомеостатических механизмов.[53] Датчик первого находится в паращитовидные железы, где главные ячейки почувствовать Ca2+ уровень с помощью специализированных рецепторов кальция в их мембранах. Датчики для второго - это парафолликулярные клетки в щитовидная железа. Главные клетки паращитовидной железы выделяют паратироидный гормон (ПТГ) в ответ на падение уровня ионизированного кальция в плазме; парафолликулярные клетки щитовидной железы выделяют кальцитонин в ответ на повышение уровня ионизированного кальция в плазме.
В эффектор органами первого гомеостатического механизма являются кости, то почка, и через гормон, выделяемый в кровь почками в ответ на высокий уровень ПТГ в крови, двенадцатиперстная кишка и тощая кишка. Гормон паращитовидной железы (в высоких концентрациях в крови) вызывает резорбция кости, высвобождая кальций в плазму. Это очень быстрое действие, которое может устранить угрозу. гипокальциемия в течение нескольких минут. Высокие концентрации ПТГ вызывают выведение фосфат-ионы через мочу. Поскольку фосфаты соединяются с ионами кальция с образованием нерастворимых солей (см. Также костный минерал ), снижение уровня фосфатов в крови, высвобождает свободные ионы кальция в ионизированный пул кальция в плазме. ПТГ оказывает на почки второе действие. Он стимулирует производство и высвобождение почками кальцитриол в кровь. Этот стероидный препарат Гормон действует на эпителиальные клетки верхнего отдела тонкой кишки, увеличивая их способность поглощать кальций из содержимого кишечника в кровь.[54]
Второй гомеостатический механизм с его датчиками в щитовидной железе высвобождает кальцитонин в кровь, когда уровень ионизированного кальция в крови повышается. Этот гормон действует в первую очередь на кости, вызывая быстрое удаление кальция из крови и откладывание его в нерастворимой форме в костях.[нужна цитата ]
Два гомеостатических механизма, действующие через ПТГ, с одной стороны, и кальцитонин, с другой, могут очень быстро исправить любую приближающуюся ошибку в уровне ионизированного кальция в плазме, либо удаляя кальций из крови и откладывая его в скелете, либо удаляя из нее кальций. . В скелет действует как чрезвычайно большой запас кальция (около 1 кг) по сравнению с запасом кальция в плазме (около 180 мг). Более длительное регулирование происходит за счет абсорбции или потери кальция из кишечника.
Другой пример - это наиболее хорошо охарактеризованные эндоканнабиноиды подобно анандамид (N-арахидоноилэтаноламид; AEA) и 2-арахидоноилглицерин (2-AG), синтез которого происходит под действием ряда внутриклеточный ферменты активируется в ответ на повышение уровня внутриклеточного кальция для введения гомеостаза и предотвращения опухоль развитие через предполагаемые защитные механизмы, которые предотвращают рост клеток и миграция путем активации CB1 и / или CB2 и прилегающий рецепторы.[55]
Концентрация натрия
Гомеостатический механизм, контролирующий концентрацию натрия в плазме, намного сложнее, чем большинство других гомеостатических механизмов, описанных на этой странице.
Датчик расположен в юкстагломерулярный аппарат почек, который определяет концентрацию натрия в плазме неожиданно косвенным образом. Вместо того, чтобы измерять его непосредственно в крови, проходящей мимо юкстагломерулярные клетки эти клетки реагируют на концентрацию натрия в почечная канальцевая жидкость после того, как он уже претерпел определенную модификацию в проксимальный извитый каналец и петля Генле.[56] Эти клетки также реагируют на скорость кровотока через юкстагломерулярный аппарат, который в нормальных условиях прямо пропорционален скорости кровотока. артериальное кровяное давление, что делает эту ткань дополнительным датчиком артериального давления.
В ответ на снижение концентрации натрия в плазме или падение артериального давления юкстагломерулярные клетки высвобождают ренин в кровь.[56][57][58] Ренин - это фермент, расщепляющий декапептид (короткая белковая цепь, длиной 10 аминокислот) из плазмы α-2-глобулин называется ангиотензиноген. Этот декапептид известен как ангиотензин I.[56] Не имеет известной биологической активности. Однако, когда кровь циркулирует в легких, легочный капилляр эндотелиальный фермент под названием ангиотензин-превращающий фермент (ACE) отщепляет еще две аминокислоты от ангиотензина I с образованием октапептида, известного как ангиотензин II. Ангиотензин II - это гормон, который действует на кора надпочечников, вызывая выброс в кровь стероидный гормон, альдостерон. Ангиотензин II также действует на гладкие мышцы стенок артериол, вызывая сужение этих сосудов малого диаметра, тем самым ограничивая отток крови из артериального дерева, вызывая повышение артериального давления. Таким образом, это усиливает меры, описанные выше (под заголовком «Артериальное кровяное давление»), которые защищают артериальное кровяное давление от изменений, особенно гипотония.
Стимулированный ангиотензином II альдостерон освобожден от зона клубочков из надпочечники оказывает влияние, в частности, на эпителиальные клетки дистальные извитые канальцы и сборные каналы почек. Здесь он вызывает реабсорбцию ионов натрия из почечная канальцевая жидкость в обмен на ионы калия, которые выделяются из плазмы крови в канальцевую жидкость и выходят из организма через мочу.[56][59] Реабсорбция ионов натрия из жидкости почечных канальцев останавливает дальнейшую потерю ионов натрия из организма и, следовательно, предотвращает ухудшение состояния. гипонатриемия. Гипонатриемия может быть только исправлено за счет потребления соли в рационе. Однако неясно, может ли «солевой голод» быть инициирован гипонатриемией и каким механизмом это могло произойти.
Когда концентрация ионов натрия в плазме выше нормы (гипернатриемия ) прекращается высвобождение ренина из юкстагломерулярного аппарата, прекращается выработка ангиотензина II и его последующее высвобождение альдостерона в кровь. Почки отвечают выделением ионов натрия с мочой, тем самым нормализуя концентрацию ионов натрия в плазме. Низкий уровень ангиотензина II в крови снижает артериальное давление как неизбежный сопутствующий ответ.
Реабсорбция ионов натрия из канальцевой жидкости в результате высокого уровня альдостерона в крови сама по себе не приводит к возврату воды из канальцев в кровь из канальцев. дистальные извитые канальцы или же сборные каналы. Это связано с тем, что натрий реабсорбируется в обмен на калий и поэтому вызывает лишь незначительное изменение осмотический градиент между кровью и трубчатой жидкостью. Кроме того, эпителий дистальных извитых канальцев и собирательных трубок непроницаем для воды в отсутствие антидиуретический гормон (АДГ) в крови. ADH является частью контроля над баланс жидкости. Его уровень в крови зависит от осмоляльность плазмы, которая измеряется в гипоталамус мозга. Действие альдостерона на почечные канальцы предотвращает потерю натрия в внеклеточной жидкости (ECF). Таким образом, не происходит изменения осмоляльности ECF и, следовательно, не изменяется концентрация ADH в плазме.Однако низкие уровни альдостерона вызывают потерю ионов натрия из ECF, что потенциально может вызвать изменение внеклеточной осмоляльности и, следовательно, уровней ADH в крови.
Концентрация калия
Высокие концентрации калия в плазме вызывают деполяризация из зона клубочков мембраны клеток во внешнем слое кора надпочечников.[60] Это вызывает высвобождение альдостерон в кровь.
Альдостерон действует прежде всего на дистальные извитые канальцы и сборные каналы почек, стимулируя выведение ионов калия с мочой.[56] Однако он делает это, активируя базолатеральный Na+/ К+ насосы эпителиальных клеток канальцев. Эти обменники натрия / калия перекачивают три иона натрия из клетки в интерстициальную жидкость и два иона калия в клетку из интерстициальной жидкости. Это создает градиент ионной концентрации что приводит к реабсорбции натрия (Na+) ионы из канальцевой жидкости в кровь и секретируют калий (K+) ионы из крови в мочу (просвет собирательного протока).[61][62]
Баланс жидкости
В общее количество воды в организме нужно поддерживать баланс. Баланс жидкости включает в себя стабилизацию объема жидкости, а также поддержание уровней электролиты во внеклеточной жидкости стабильна. Баланс жидкости поддерживается в процессе осморегуляция и по поведению. Осмотическое давление обнаруживается осморецепторы в срединное преоптическое ядро в гипоталамус. Измерение плазмы осмоляльность чтобы дать представление о содержании воды в организме, полагается на тот факт, что потери воды из организма (через неизбежная потеря воды через кожу который не является полностью водонепроницаемым и поэтому всегда слегка влажным, водяной пар в выдыхаемом воздухе, потливость, рвота, нормальный кал и особенно понос ) все гипотонический, что означает, что они менее соленые, чем жидкости организма (сравните, например, вкус слюны со вкусом слез. Последняя имеет почти такое же содержание соли, что и внеклеточная жидкость, тогда как первая является гипотонической по отношению к плазме. Слюна не имеет соленого вкуса, тогда как слезы явно соленые). Почти все нормальные и аномальные потери вода тела поэтому заставляют внеклеточную жидкость становиться гипертонический. И наоборот, чрезмерное потребление жидкости разбавляет внеклеточную жидкость, вызывая регистрацию гипоталамуса. гипотоническая гипонатриемия условия.
Когда гипоталамус обнаруживает гипертоническую внеклеточную среду, вызывает секрецию антидиуретического гормона (АДГ), называемого вазопрессин который действует на эффекторный орган, которым в данном случае является почка. Воздействие вазопрессина на почечные канальцы заключается в реабсорбции воды из дистальные извитые канальцы и сборные каналы, таким образом предотвращая ухудшение потери воды с мочой. Гипоталамус одновременно стимулирует близлежащие центр жажды вызывая почти непреодолимое (если гипертонус достаточно серьезный) позыв выпить воды. Прекращение оттока мочи предотвращает гиповолемия и гипертонус от ухудшения; питье воды исправляет дефект.
Гипоосмоляльность приводит к очень низким уровням АДГ в плазме. Это приводит к подавлению реабсорбции воды из почечных канальцев, вызывая выделение большого количества очень разбавленной мочи, тем самым избавляясь от избытка воды в организме.
Потеря воды с мочой, когда гомеостат воды в организме не нарушен, является компенсационный потеря воды, исправление любой избыток воды в организме. Однако, поскольку почки не могут вырабатывать воду, рефлекс жажды является важнейшим вторым эффекторным механизмом гомеостата воды в организме. исправление любой дефицит воды в организме.
PH крови

В pH плазмы может изменяться респираторными изменениями парциального давления углекислого газа; или изменено метаболическими изменениями в угольная кислота к бикарбонат-ион соотношение. В бикарбонатная буферная система регулирует соотношение угольной кислоты и бикарбоната, равное 1:20, при этом соотношении pH крови составляет 7,4 (как поясняется в Уравнение Хендерсона – Хассельбаха ). Изменение pH плазмы дает кислотно-щелочной дисбаланс.В кислотно-щелочной гомеостаз Есть два механизма, которые могут помочь регулировать pH. Респираторная компенсация механизм дыхательный центр, регулирует парциальное давление диоксида углерода изменяя частоту и глубину дыхания, чтобы вернуть pH в норму. Парциальное давление диоксида углерода также определяет концентрацию угольной кислоты, и буферная система бикарбоната также может иметь значение. Почечная компенсация может помочь буферной системе бикарбоната. Датчик концентрации бикарбоната плазмы точно неизвестен. Весьма вероятно, что клетки почечных канальцев дистальных извитых канальцев сами чувствительны к pH плазмы.[нужна цитата ] Метаболизм этих клеток производит двуокись углерода, которая быстро превращается в водород и бикарбонат под действием карбоангидраза.[63] Когда pH ECF падает (становится более кислым), клетки почечных канальцев выделяют ионы водорода в канальцевую жидкость, чтобы покинуть организм с мочой. Ионы бикарбоната одновременно секретируются в кровь, что снижает содержание угольной кислоты и, как следствие, повышает pH плазмы.[63] Обратное происходит, когда pH плазмы поднимается выше нормы: ионы бикарбоната выводятся с мочой, а ионы водорода выделяются в плазму.
Когда ионы водорода выводятся с мочой, а бикарбонат - в кровь, последний соединяется с избыточными ионами водорода в плазме, которые стимулировали почки к выполнению этой операции. В результате реакции в плазме образуется угольная кислота, которая находится в равновесии с парциальным давлением диоксида углерода в плазме. Это строго регулируется, чтобы исключить чрезмерное накопление угольной кислоты или бикарбоната. Таким образом, общий эффект заключается в том, что ионы водорода теряются с мочой при падении pH плазмы. Сопутствующее повышение уровня бикарбоната плазмы убирает увеличенное количество ионов водорода (вызванное падением pH плазмы), и образующийся избыток угольной кислоты удаляется в легкие в виде диоксида углерода. Это восстанавливает нормальное соотношение между бикарбонатом и парциальным давлением углекислого газа и, следовательно, pH плазмы. Обратное происходит, когда высокий pH плазмы стимулирует почки выделять ионы водорода в кровь и выводить бикарбонат с мочой. Ионы водорода объединяются с избыточными ионами бикарбоната в плазме, снова образуя избыток угольной кислоты, которая может выдыхаться в виде диоксида углерода в легкие, поддерживая концентрацию бикарбонат-иона в плазме, парциальное давление диоксида углерода и, следовательно, , pH плазмы, постоянный.
Спинномозговая жидкость
Цереброспинальная жидкость (ЦСЖ) позволяет регулировать распределение веществ между клетками головного мозга,[64] и нейроэндокринный факторы, незначительные изменения которых могут вызвать проблемы или повредить нервную систему. Например, высокий глицин концентрация нарушает температура и артериальное давление контроль, и высокий CSF pH причины головокружение и обморок.[65]
Нейротрансмиссия
Тормозящие нейроны в Центральная нервная система играют гомеостатическую роль в балансе нейрональной активности между возбуждением и торможением. Тормозящие нейроны, использующие ГАМК, вносят компенсирующие изменения в нейронные сети, предотвращая скачкообразные уровни возбуждения.[66] Считается, что дисбаланс между возбуждением и торможением связан с рядом факторов: нервно-психические расстройства.[67]
Нейроэндокринная система
В нейроэндокринная система это механизм, с помощью которого гипоталамус поддерживает гомеостаз, регулируя метаболизм, воспроизводство, пищевое и питьевое поведение, использование энергии, осмолярность и артериальное давление.
Регуляция обмена веществ осуществляется гипоталамический соединения с другими железами.[68] Три эндокринные железы из гипоталамо-гипофизарно-гонадная ось (Ось HPG) часто работают вместе и выполняют важные регулирующие функции. Две другие регуляторные эндокринные оси - это гипоталамо-гипофизарно-надпочечниковая ось (Ось HPA) и гипоталамо-гипофизарно-тироидная ось (Ось HPT).
В печень также выполняет множество регулирующих функций обмена веществ. Важная функция - производство и контроль желчные кислоты. Слишком большое количество желчной кислоты может быть токсичным для клеток, и ее синтез может подавляться активацией FXR а ядерный рецептор.[4]
Генная регуляция
На клеточном уровне гомеостаз осуществляется несколькими механизмами, включая транскрипционная регуляция это может изменить активность генов в ответ на изменения.
Энергетический баланс
Эта секция нуждается в расширении. Вы можете помочь добавляя к этому. (Ноябрь 2017 г.) |
Количество энергии, поступающей через питание должно соответствовать количеству используемой энергии. Для достижения энергетического гомеостаза аппетит регулируется двумя гормонами, Грехлин и лептин. Грехлин стимулирует чувство голода, а прием пищи и лептина сигнализирует о сытости (сытости).
Обзор вмешательств по изменению веса за 2019 год, включая на диете, упражнения и переедание, обнаружили, что гомеостаз массы тела не мог точно исправить «энергетические ошибки», потерю или прирост калорий в краткосрочной перспективе.[69]
Клиническое значение
Многие заболевания являются результатом нарушения гомеостаза. Практически любой гомеостатический компонент может выйти из строя либо в результате наследственный дефект, врожденная ошибка обмена веществ, или приобретенное заболевание. Некоторые гомеостатические механизмы имеют встроенную избыточность, которая гарантирует, что жизнь не подвергнется немедленной угрозе в случае выхода из строя какого-либо компонента; но иногда нарушение гомеостаза может привести к серьезному заболеванию, которое может быть фатальным, если его не лечить. Хорошо известный пример нарушения гомеостаза показан на сахарный диабет 1 типа. Здесь регулирование уровня сахара в крови не может работать, потому что бета-клетки из островки поджелудочной железы уничтожаются и не могут производить необходимые инсулин. Уровень сахара в крови повышается в состоянии, известном как гипергликемия.
Гомеостат кальция, ионизированный плазмой, может быть нарушен постоянным, неизменным, избыточным производством паратироидный гормон паращитовидными железами аденома что приводит к типичным особенностям гиперпаратиреоз, а именно высоко ионизированный плазмой Ca2+ уровни и резорбция кости, что может привести к спонтанным переломам. Аномально высокие концентрации ионизированного кальция в плазме вызывают конформационные изменения многих белков клеточной поверхности (особенно ионных каналов и рецепторов гормонов или нейротрансмиттеров).[70] вызывает вялость, мышечную слабость, анорексию, запор и лабильные эмоции.[71]
Гомеостат воды в организме может быть нарушен из-за неспособности выделять ADH в ответ даже на обычные ежедневные потери воды с выдыхаемым воздухом, кал, и бесчувственное потоотделение. При нулевом сигнале АДГ в крови почки производят огромные неизменные объемы очень разбавленной мочи, вызывая обезвоживание и смерть, если не лечить.
По мере старения организмов эффективность их систем управления снижается. Неэффективность постепенно приводит к нестабильной внутренней среде, что увеличивает риск заболевания и приводит к физическим изменениям, связанным со старением.[5]
Разные хронический Заболевания контролируются гомеостатической компенсацией, которая маскирует проблему, компенсируя ее (компенсируя ее) другим способом. Однако компенсирующие механизмы со временем изнашиваются или нарушаются новым усложняющим фактором (например, появлением сопутствующей острой вирусной инфекции), что заставляет тело раскачиваться через новый каскад событий. Такая декомпенсация выявляет основное заболевание, усугубляя его симптомы. Общие примеры включают декомпенсированные сердечная недостаточность, почечная недостаточность, и отказ печени.
Биосфера
в Гипотеза Гайи, Джеймс Лавлок[72] заявил, что вся масса живого вещества на Земле (или любой планете с жизнью) функционирует как огромное гомеостатическое суперорганизм который активно изменяет свою планетарную среду для создания условий окружающей среды, необходимых для собственного выживания. С этой точки зрения вся планета поддерживает несколько гомеостазов (первичный гомеостаз - температурный). Присутствует ли такая система на Земле, пока не ясно. Однако обычно принимаются некоторые относительно простые гомеостатические механизмы. Например, иногда утверждается, что при повышении уровня углекислого газа в атмосфере некоторые растения могут расти лучше и, таким образом, удаляют больше углекислого газа из атмосферы. Однако потепление усугубило засухи, в результате чего вода ограничивающий фактор на земле. Утверждается, что когда солнечного света много и температура воздуха повышается, фитопланктон поверхностных вод океана, действующих как глобальный солнечный свет и, следовательно, тепловые датчики, могут процветать и производить больше диметилсульфид (DMS). Молекулы ДМС действуют как облачные ядра конденсации, которые создают больше облаков и, таким образом, увеличивают атмосферный альбедо, и это способствует снижению температуры атмосферы. Однако повышение температуры моря привело к расслоению океанов, отделяя теплые, залитые солнцем воды от прохладных, богатых питательными веществами. Таким образом, питательные вещества стали ограничивающим фактором, и уровни планктона фактически упали за последние 50 лет, а не повысились. По мере того как ученые все больше узнают о Земле, обнаруживается огромное количество контуров положительной и отрицательной обратной связи, которые вместе поддерживают метастабильное состояние, иногда в очень широком диапазоне условий окружающей среды.
Прогнозирующий
Прогнозирующий гомеостаз - это упреждающий ответ на ожидаемый вызов в будущем, такой как стимуляция секреции инсулина гормонами кишечника, которые попадают в кровь в ответ на прием пищи.[38] Эта секреция инсулина происходит до повышения уровня сахара в крови, что снижает уровень сахара в крови в ожидании большого притока в кровь глюкозы в результате переваривания углеводов в кишечнике.[73] Такие упреждающие реакции представляют собой системы с разомкнутым контуром, которые, по сути, основаны на «догадках» и не самокорректируются.[74] Для упреждающих реакций всегда требуется замкнутая система отрицательной обратной связи, чтобы исправить «перевыпуски» и «недолеты», к которым склонны упреждающие системы.
Другие поля
Этот термин стал использоваться в других областях, например:
Риск
An актуарий может относиться к гомеостаз риска, где (например) люди с антиблокировочной системой тормозов не имеют более высоких показателей безопасности, чем люди без антиблокировочной системы тормозов, потому что первые подсознательно компенсируют более безопасное транспортное средство менее безопасными привычками вождения. До изобретения антиблокировочной системы тормозов некоторые маневры включали незначительные заносы, вызывая страх и избегание: теперь антиблокировочная система перемещает границу для такой обратной связи, и модели поведения расширяются в область, которая больше не карается. Также было высказано предположение, что экологические кризисы являются примером гомеостаза риска, при котором определенное поведение продолжается до тех пор, пока не наступят доказанные опасные или драматические последствия.[75][самостоятельно опубликованный источник? ]
Стресс
Социологи и психологи могут ссылаться на стрессовый гомеостаз, тенденция населения или отдельного человека оставаться на определенном уровне стресс, часто создавая искусственные стрессы, если «естественного» уровня стресса недостаточно.[76][самостоятельно опубликованный источник? ]
Жан-Франсуа Лиотар, теоретик постмодерна, применил этот термин к социальным «центрам власти», которые он описывает в Состояние постмодерна, как «управляемая принципом гомеостаза», например, научная иерархия, которая иногда годами игнорирует радикально новое открытие, потому что оно дестабилизирует ранее принятые нормы.
Технологии
К знакомым технологическим гомеостатическим механизмам относятся:
- А термостат работает путем включения и выключения обогревателей или кондиционеров в ответ на выходной сигнал датчика температуры.
- Круиз-контроль регулирует дроссельную заслонку автомобиля в ответ на изменение скорости.[77][78]
- An автопилот управляет рулевым управлением самолета или корабля в ответ на отклонение от предварительно установленного пеленга или маршрута по компасу.[79]
- Контроль над процессом системы в химический завод или же нефтеперегонный завод поддерживать уровни жидкости, давление, температуру, химический состав и т. д., управляя нагревателями, насосами и клапанами.[80]
- В центробежный регулятор из паровой двигатель, как разработано Джеймс Ватт в 1788, уменьшает дроссельную заслонку в ответ на увеличение скорости двигателя или открывает клапан, если скорость падает ниже заданного значения.[81][82]
Смотрите также
- Апоптоз - Запрограммированная гибель клеток в многоклеточных организмах
- Церебральная ауторегуляция
- Хронобиология
- Энантиостаз
- Геофизиология
- Гликобиология
- Гомеорез
- Гомеостатическая пластичность
- Гормезис
- Принцип Ле Шателье - Принцип прогнозирования воздействия изменения условий на химическое равновесие
- Закон Ленца
- Осмос - химический процесс
- Протеостаз
- Старение
- Устойчивое состояние
- Системная биология - Вычислительное и математическое моделирование сложных биологических систем.
- Vis medicatrix naturae
Рекомендации
- ^ Гордон., Беттс, Дж. Анатомия и психология. ДеСе, Питер., Джонсон, Эдди., Джонсон, Джоди Э., Корол, Оксана., Круз, Дин Х., По, Брэндон. Хьюстон, Техас. п. 9. ISBN 9781947172043. OCLC 1001472383.
- ^ Мартин, Элизабет (2008). Словарь биологии (6-е изд.). Оксфорд: Издательство Оксфордского университета. С. 315–316. ISBN 978-0-19-920462-5.
- ^ Биология онлайн. «Гомеостаз». Биология онлайн. Получено 27 октября 2019.
- ^ а б Калаани, штат Нью-Йорк; Мангельсдорф, DJ (2006). «LXRS и FXR: инь и янь холестерина и жирового обмена». Ежегодный обзор физиологии. 68: 159–91. Дои:10.1146 / annurev.physiol.68.033104.152158. PMID 16460270.
- ^ а б c Мариеб Э. Н., Хоэн К. Н. (2009). Основы анатомии и физиологии человека (9-е изд.). Сан-Франциско: Пирсон / Бенджамин Каммингс. ISBN 978-0321513427.
- ^ Ловингер, Дэвид М. (2008), "Пресинаптическая модуляция эндоканнабиноидами", в Südhof, Thomas C .; Старке, Клаус (ред.), Фармакология высвобождения нейротрансмиттера, Справочник по экспериментальной фармакологии, 184, Springer Berlin Heidelberg, стр. 435–477, Дои:10.1007/978-3-540-74805-2_14, ISBN 9783540748052, PMID 18064422
- ^ Фрейтас, Геркулес Резенде; Исаак, Алинни Розендо; Мальхер-Лопес, Ренато; Диас, Бруно Лоуренсо; Тревенцоли, Исида Хара; Рейс, Рикардо Аугусто Де Мело (26 ноября 2018 г.). «Полиненасыщенные жирные кислоты и эндоканнабиноиды в здоровье и болезнях». Пищевая неврология. 21 (10): 695–714. Дои:10.1080 / 1028415X.2017.1347373. ISSN 1028-415X. PMID 28686542. S2CID 40659630.
- ^ а б Кэннон, У. (1932). Мудрость тела. Нью-Йорк: У. В. Нортон. С. 177–201.
- ^ а б Кэннон, В. Б. (1926). «Физиологическая регуляция нормальных состояний: некоторые предварительные постулаты относительно биологической гомеостатики». В А. Петтит (ред.). A Charles Riches amis, ses collègues, ses élèves (На французском). Париж: Les Éditions Médicales. п. 91.
- ^ Смит, Джерард П. (2008). «Непризнанный вклад Павлова и Баркрофта в теорию гомеостаза Кэннона». Аппетит. 51 (3): 428–432. Дои:10.1016 / j.appet.2008.07.003. PMID 18675307. S2CID 43088475.
- ^ Зорея, Аарон (2014). Стероиды (здоровье и медицинские проблемы сегодня). Вестпорт, Коннектикут: Greenwood Press. п. 10. ISBN 978-1440802997.
- ^ Риггс, Д.С. (1970). Теория управления и механизмы физиологической обратной связи. Балтимор: Уильямс и Уилкинс.
- ^ а б Холл, Джон (2011). Гайтон и Холл учебник медицинской физиологии (12-е изд.). Филадельфия, Пенсильвания: Saunders / bich er. С. 4–9. ISBN 9781416045748.
- ^ а б Милсум, Дж. (1966). Анализ систем биологического контроля. Нью-Йорк: Макгроу-Хилл.
- ^ «Гомеостаз». Словарь Merriam-Webster.
- ^ «Гомеостаз». Dictionary.com Несокращенный. Случайный дом.
- ^ а б c d е ж грамм Тортора, Джерард Дж .; Анагностакос, Николас П. (1987). Основы анатомии и физиологии (Пятое изд.). Нью-Йорк: Harper & Row, Publishers. стр.315 –316, 475, 657–658. ISBN 978-0-06-350729-6.
- ^ Ханская академия. «Гомеостаз». Ханская академия. Получено 13 июля 2018.
- ^ Сведан, Надя Габриэле (2001). Женская спортивная медицина и реабилитация. Липпинкотт Уильямс и Уилкинс. п. 149. ISBN 978-0-8342-1731-7.
- ^ Вешлер, Тони (2002). Взять на себя ответственность за свою фертильность. Нью-Йорк: HarperCollins. стр.52, 316, 361–362. ISBN 978-0-06-093764-5.
- ^ Клюге, Мэтью Дж. (2015). Лихорадка: ее биология, эволюция и функции. Издательство Принстонского университета. п. 57. ISBN 9781400869831.
- ^ Гармель, Гус М. (2012). «Лихорадка у взрослых». In Mahadevan, S.V .; Гармель, Гас М. (ред.). Введение в неотложную клиническую медицину (2-е изд.). Кембридж: Издательство Кембриджского университета. п. 375. ISBN 978-0521747769.
- ^ Запад, Брюс Дж (2006). Где медицина пошла не так: заново открывая путь к сложности. Исследования нелинейных явлений в науках о жизни. 11. Нью-Джерси: World Scientific. Дои:10.1142/6175. ISBN 978-981-256-883-0.
- ^ Лонго, Джузеппе; Монтевиль, Маэль (2014). Взгляды на организмы. Конспект лекций по морфогенезу. Springer. Дои:10.1007/978-3-642-35938-5. ISBN 978-3-642-35937-8. S2CID 27653540.
- ^ Шеннон, Клод Э .; Уивер, Уоррен (1963). Математическая теория коммуникации (4-е изд.). Урбана: Университет Иллинойса Press. ISBN 978-0252725487.
- ^ Rucker, R. (1987). Инструменты разума: математика информации. Хармондсворт: Книги Пингвина. С. 25–30.
- ^ Koeslag, Johan H .; Сондерс, Питер Т .; Весселс, Джабус А. (1999). «Хромогранины и контррегулирующие гормоны: имеют ли они гомеостатический смысл?». Журнал физиологии. 517 (3): 643–649. Дои:10.1111 / j.1469-7793.1999.0643s.x. ЧВК 2269385. PMID 10358106.
- ^ а б c Уильямс, Питер Л .; Уорик, Роджер; Дайсон, Мэри; Баннистер, Лоуренс Х. (1989). Анатомия Грея (Тридцать седьмое изд.). Эдинбург: Черчилль Ливингстон. С. 691–692, 791, 10011–10012. ISBN 0443-041776.
- ^ Tansey, Etain A .; Джонсон, Кристофер Д. (2015). «Последние достижения в области терморегуляции». Достижения в физиологическом образовании. 39 (3): 139–148. Дои:10.1152 / advan.00126.2014. ISSN 1043-4046. PMID 26330029.
- ^ Стендринг, Сьюзен (7 августа 2015 г.). Анатомия Грея: анатомические основы клинической практики. Стендринг, Сьюзен (41-е изд.). [Филадельфия]. С. 141, 151–152. ISBN 9780702068515. OCLC 920806541.
- ^ Purves, Дейл (2011). Неврология (5-е изд.). Сандерленд, Массачусетс: Sinauer. п. 458. ISBN 978-0-87893-695-3.
- ^ а б c Кэмпбелл, Нил А. (1990). Биология (Второе изд.). Редвуд-Сити, Калифорния: Издательство Бенджамин / Каммингс. С. 897–898. ISBN 978-0-8053-1800-5.
- ^ Flouris, AD (январь 2011 г.). «Функциональная архитектура поведенческой терморегуляции». Европейский журнал прикладной физиологии. 111 (1): 1–8. Дои:10.1007 / s00421-010-1602-8. PMID 20711785. S2CID 9109352.
- ^ Гилрой, Энн М .; Макферсон, Брайан Р .; Росс, Лоуренс М. (2008). Атлас анатомии. Штутгарт: Thieme Medical Publishers. С. 318, 349. ISBN 978-1-60406-062-1.
- ^ Шмидт-Нильсен К. (1981). «Противоточные системы у животных». Scientific American. 244 (5): 118–28. Bibcode:1981SciAm.244e.118S. Дои:10.1038 / scientificamerican0581-118. PMID 7233149.
- ^ Стюарт, И. (2011). Физиология человека (Двенадцатое изд.). Нью-Йорк: Макгроу-Хилл. п. 667.
- ^ Бхагаван, Н. В. (2002). Медицинская биохимия (4-е изд.). Академическая пресса. п. 499. ISBN 978-0-12-095440-7.
- ^ а б c Koeslag, Johan H .; Сондерс, Питер Т .; Тербланш, Эльмари (2003). «Тематический обзор: переоценка гомеостата глюкозы в крови, которая всесторонне объясняет комплекс X синдрома диабета 2 типа». Журнал физиологии. 549 (Pt 2): 333–346. Дои:10.1113 / jphysiol.2002.037895. ЧВК 2342944. PMID 12717005.
- ^ Страйер, Люберт (1995). Биохимия (Четвертое изд.). Нью-Йорк: W.H. Фримен и компания. С. 164, 773–774. ISBN 0-7167-2009-4.
- ^ Aronoff, Стивен Л .; Берковиц, Кэти; Шрейнер, Барб; Хочу, Лаура (1 июля 2004 г.). «Метаболизм и регуляция глюкозы: помимо инсулина и глюкагона». Спектр диабета. 17 (3): 183–190. Дои:10.2337 / diaspect.17.3.183. ISSN 1040-9165.
- ^ Спайер, км; Гурин, А.В. (12 сентября 2009 г.). «Хемосенсорные пути в стволе мозга, контролирующие кардиореспираторную деятельность». Философские труды Лондонского королевского общества. Серия B, Биологические науки. 364 (1529): 2603–10. Дои:10.1098 / rstb.2009.0082. ЧВК 2865116. PMID 19651660.
- ^ Павлин, Эндрю Дж (17 октября 1998 г.). «Кислород на большой высоте». Британский медицинский журнал. 317 (7165): 1063–1066. Дои:10.1136 / bmj.317.7165.1063. ЧВК 1114067. PMID 9774298.
- ^ Янг, Эндрю Дж; Ривз, Джон Т. (2002). «Адаптация человека к большой высоте над землей» (PDF). Медицинские аспекты суровых условий окружающей среды. 2. Институт Бордена, Вашингтон, округ Колумбия. CiteSeerX 10.1.1.175.3270. Архивировано из оригинал (PDF) 16 сентября 2012 г.. Получено 5 января 2009.
- ^ Харрис, Н. Стюарт; Нельсон, Сара В. (16 апреля 2008 г.). «Высотная болезнь - церебральные синдромы». Медицинские специальности> Скорая помощь> Экология.
- ^ Альбертс, Брюс (2002). Молекулярная биология клетки (4-е изд.). Нью-Йорк [u.a.]: Гарланд. С. 1292–1293. ISBN 978-0-8153-4072-0.
- ^ а б Тортора, Джерард Дж .; Анагностакос, Николас П. (1987). Основы анатомии и физиологии (Пятое изд.). Нью-Йорк: Harper & Row, Publishers. стр.444–445. ISBN 978-0-06-350729-6.
- ^ Фишер Дж. В., Кури С., Дьюси Т., Мендель С. (1996). «Продукция эритропоэтина интерстициальными клетками почек гипоксической обезьяны». Британский журнал гематологии. 95 (1): 27–32. Дои:10.1046 / j.1365-2141.1996.d01-1864.x. PMID 8857934. S2CID 38309595.
- ^ Елкманн В (2007). «Эритропоэтин после столетия исследований: моложе, чем когда-либо». Европейский журнал гематологии. 78 (3): 183–205. Дои:10.1111 / j.1600-0609.2007.00818.x. PMID 17253966. S2CID 37331032.
- ^ (PDF). 27 февраля 2008 г. https://web.archive.org/web/20080227162001/http://www.orlandoregional.org/pdf%20folder/overview%20adult%20brain%20injury.pdf. Архивировано из оригинал (PDF) 27 февраля 2008 г. Отсутствует или пусто
| название =(помощь) - ^ Покок, Джиллиан; Ричардс, Кристофер Д. (2006). Физиология человека: основы медицины (3-е изд.). Оксфорд: Издательство Оксфордского университета. п. 4. ISBN 978-0-19-856878-0.
- ^ Тортора, Джерард Дж .; Анагностакос, Николас П. (1987). Основы анатомии и физиологии (Пятое изд.). Нью-Йорк: Harper & Row, Publishers. п.430. ISBN 978-0-06-350729-6.
- ^ Покок, Джиллиан; Ричардс, Кристофер Д. (2006). Физиология человека: основы медицины (3-е изд.). Оксфорд: Издательство Оксфордского университета. С. 299–302. ISBN 978-0-19-856878-0.
- ^ Брини М, Оттолини Д., Кали Т, Карафоли Э (2013). «Глава 4. Кальций в здоровье и болезнях». В Sigel A, Helmut RK (ред.). Взаимосвязь между ионами эссенциальных металлов и заболеваниями человека. Ионы металлов в науках о жизни. 13. Springer. С. 81–137. Дои:10.1007/978-94-007-7500-8_4. ISBN 978-94-007-7499-5. PMID 24470090.
- ^ Страйер, Люберт (1995). «Витамин D получается из холестерина в результате расщепления колец светом». В: Биохимия. (Четвертое изд.). Нью-Йорк: W.H. Фримен и компания. п. 707. ISBN 0-7167-2009-4.
- ^ Аяканну, Тангесверан; Тейлор, Энтони Х .; Marczylo, Timothy H .; Виллетс, Джонатон М .; Конье, Джастин С. (2013). «Эндоканнабиноидная система и рак, зависимый от половых стероидных гормонов». Международный журнал эндокринологии. 2013: 259676. Дои:10.1155/2013/259676. ISSN 1687-8337. ЧВК 3863507. PMID 24369462.
- ^ а б c d е Тортора, Джерард Дж .; Анагностакос, Николас П. (1987). Основы анатомии и физиологии (Пятое изд.). Нью-Йорк: Harper & Row, Publishers. стр.420–421. ISBN 978-0-06-350729-6.
- ^ Престон, Ричард А .; Materson, B.J .; Reda, D.J .; Уильямс, Д. В .; Hamburger, R.J .; Cushman, W. C .; Андерсон, Р. Дж. (1998). "Статья JAMA, январь 2012 г.". JAMA. 280 (13): 1168–72. Дои:10.1001 / jama.280.13.1168. PMID 9777817.
- ^ Уильямс Г. Х., Длухи Р. Г. (2008).«Глава 336: Заболевания коры надпочечников». In Loscalzo J, Fauci AS, Braunwald E, Kasper DL, Hauser SL, Longo DL (ред.). Принципы внутренней медицины Харрисона. Нью-Йорк: McGraw-Hill Medical. ISBN 978-0-07-146633-2.
- ^ Бауэр Дж. Х., Гаунтнер В. К. (март 1979 г.). «Влияние хлорида калия на активность ренина плазмы и альдостерон плазмы при ограничении натрия у нормального человека». Почка Int. 15 (3): 286–93. Дои:10.1038 / ки.1979.37. PMID 513492.
- ^ Ху Ц., Русин К.Г., Тан З., Гуальярдо Н.А., Барретт П.К. (июнь 2012 г.). «Клетки клубочковой зоны коры надпочечников мыши являются собственными электрическими осцилляторами». J Clin Invest. 122 (6): 2046–2053. Дои:10.1172 / JCI61996. ЧВК 3966877. PMID 22546854.
- ^ Палмер, LG; Фриндт, Г. (2000). «Секреция альдостерона и калия кортикальным собирательным протоком». Kidney International. 57 (4): 1324–8. Дои:10.1046 / j.1523-1755.2000.00970.x. PMID 10760062.
- ^ Линас С.Л., Петерсон Л.Н., Андерсон Р.Дж., Эйзенбрей Г.А., Саймон Ф.Р., Берл Т. (июнь 1979 г.). «Механизм почечной консервации калия у крысы». Kidney International. 15 (6): 601–11. Дои:10.1038 / ки.1979.79. PMID 222934.
- ^ а б Тортора, Джерард Дж .; Анагностакос, Николас П. (1987). Основы анатомии и физиологии (Пятое изд.). Нью-Йорк: Harper & Row, Publishers. стр.581 –582, 675–676. ISBN 978-0-06-350729-6.
- ^ Сакка, Л .; Coll, G .; Чазал, Дж. (Декабрь 2011 г.). «Анатомия и физиология спинномозговой жидкости». Европейские анналы оториноларингологии, болезней головы и шеи. 128 (6): 309–316. Дои:10.1016 / j.anorl.2011.03.002. PMID 22100360.
- ^ Саладин, Кеннет (2012). Анатомия и психология (6-е изд.). Макгроу Хилл. С. 519–20.
- ^ Флорес, CE; Мендес, П. (2014). «Формовочное ингибирование: зависимая от активности структурная пластичность ГАМКергических синапсов». Границы клеточной неврологии. 8: 327. Дои:10.3389 / fncel.2014.00327. ЧВК 4209871. PMID 25386117.
- ^ Эм, Джи Вон (13 ноября 2017 г.). «Роль глиальных клеток в формировании тормозных синапсов и нервных цепей». Границы молекулярной неврологии. 10: 381. Дои:10.3389 / fnmol.2017.00381. ЧВК 5694142. PMID 29180953.
- ^ Тони, Р. (2004). «Нейроэндокринная система: организация и гомеостатическая роль». Журнал эндокринологических исследований. 27 (6 Дополнение): 35–47. PMID 15481802.
- ^ Левицкий Д.А.; Сьюолл, А; Чжун, Y; Барре, L; Shoen, S; Агаронник, Н; LeClair, JL; Чжо, Вт; Пакановски, К. (1 февраля 2019 г.). «Количественная оценка неточности потребления энергии людьми для компенсации наложенных энергетических ошибок: проблема физиологического контроля за потреблением пищи человеком». Аппетит. 133: 337–343. Дои:10.1016 / j.appet.2018.11.017. PMID 30476522. S2CID 53712116.
- ^ Армстронг CM, Cota G (март 1999 г.). «Кальциевый блок Na+ каналов и их влияние на скорость закрытия ". Труды Национальной академии наук Соединенных Штатов Америки. 96 (7): 4154–7. Bibcode:1999PNAS ... 96.4154A. Дои:10.1073 / пнас.96.7.4154. ЧВК 22436. PMID 10097179.
- ^ Харрисон, Т. Принципы внутренней медицины (третье изд.). Нью-Йорк: Книжная компания Макгроу-Хилл. С. 170, 571–579.
- ^ Лавлок, Джеймс (1991). Healing Gaia: практическая медицина для планеты. Нью-Йорк: Книги Гармонии. ISBN 978-0-517-57848-3.
- ^ Бор WF, Boulpaep EL (2009). Медицинская физиология: клеточный и молекулярный подход (2-е международное изд.). Филадельфия, Пенсильвания: Saunders / Elsevier. ISBN 9781416031154.
- ^ Koeslag, J.H .; Saunders, P.T .; Wessels, J.A. (1997). «Гомеостаз глюкозы с бесконечной выгодой: дальнейшие уроки из притчи« Мир маргариток »?». Журнал эндокринологии. 134 (2): 187–192. Дои:10.1677 / joe.0.1540187. PMID 9291828.
- ^ Спенсер, Лачи (2015). Флотация: руководство по резервуарам для сенсорной депривации, релаксации и изоляции. Lulu.com. п. 29. ISBN 978-1329173750.[самостоятельно опубликованный источник ]
- ^ Спенсер, Лачи (29 мая 2015 г.). Флотация: руководство по резервуарам для сенсорной депривации, релаксации и изоляции. Lulu.com. ISBN 9781329173750.[самостоятельно опубликованный источник ]
- ^ «Американские моторы 1966 года». Автомобильная жизнь. 12: 46. 1965. Получено 9 марта 2015.
- ^ Ницца, Карим (15 января 2001 г.). «Как работают системы круиз-контроля». Как это работает. Получено 9 марта 2015.
- ^ Харрис, Уильям (10 октября 2007 г.). «Как работает автопилот». HowStuffWorks.com. Получено 14 апреля 2018.
- ^ Уайт, Дуглас (3 октября 2005 г.). «Передовая технология автоматизации снижает затраты на электроэнергию на НПЗ». Нефтегазовый журнал. Получено 13 июля 2018.
- ^ Максвелл, Джеймс Клерк (1868). «О губернаторах». Труды Лондонского королевского общества. 16: 270–283. Дои:10.1098 / rspl.1867.0055. JSTOR 112510.
- ^ Беннетт, Стюарт (1992). История техники управления, 1930-1955 гг.. ИЭПП. п.п. 48. ISBN 978-0-86341-299-8.CS1 maint: ref = harv (связь)
дальнейшее чтение
- Clausen, M. J .; Поульсен, Х. (2013). «Глава 3 гомеостаз натрия / калия, глава 5 гомеостаз кальция, глава 6 гомеостаз марганца». В Банчи, Лючия (ред.). Металломика и клетка. Ионы металлов в науках о жизни. 12. Springer. С. 41–67. Дои:10.1007/978-94-007-5561-1_3. ISBN 978-94-007-5560-4. PMID 23595670. электронная книга ISBN 978-94-007-5561-1 ISSN 1559-0836 электронный-ISSN 1868-0402
